
Genetic and Physiological Characterization of Soybean-Nodule-Derived Isolates from Bangladeshi Soils Revealed Diverse Array of Bacteria with Potential Bradyrhizobia for Biofertilizers

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
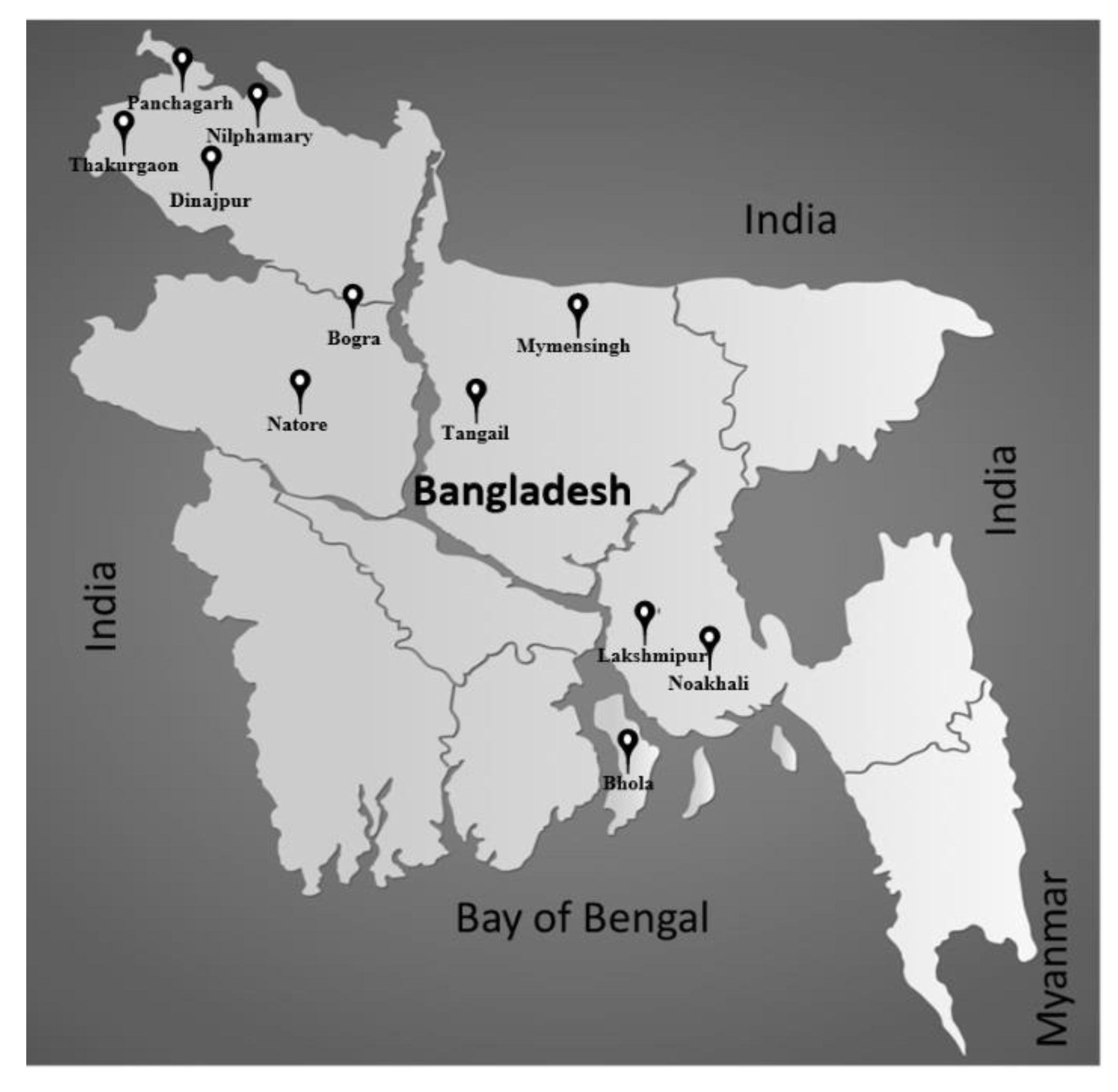
2.1. Collection of Soil Samples
2.2. Collection of Nodules Using Soil Samples in Pot Experiment
2.3. Temperature Tolerance Test of Isolates
2.4. pH Tolerance Test of Isolates
2.5. Salinity Tolerance Test of Isolates
2.6. DNA Extraction, Amplification and Sequencing
2.7. Analysis of DNA Sequences and Phylogeny Genomic
2.8. Analysis of Inoculation Effects of Selected Isolates on Plants
2.9. Acetylene Reduction Assay
2.10. Nucleotide Sequence Accession Numbers
2.11. Statistical Analysis
3. Results
3.1. Isolation of Bacteria from Root Nodule
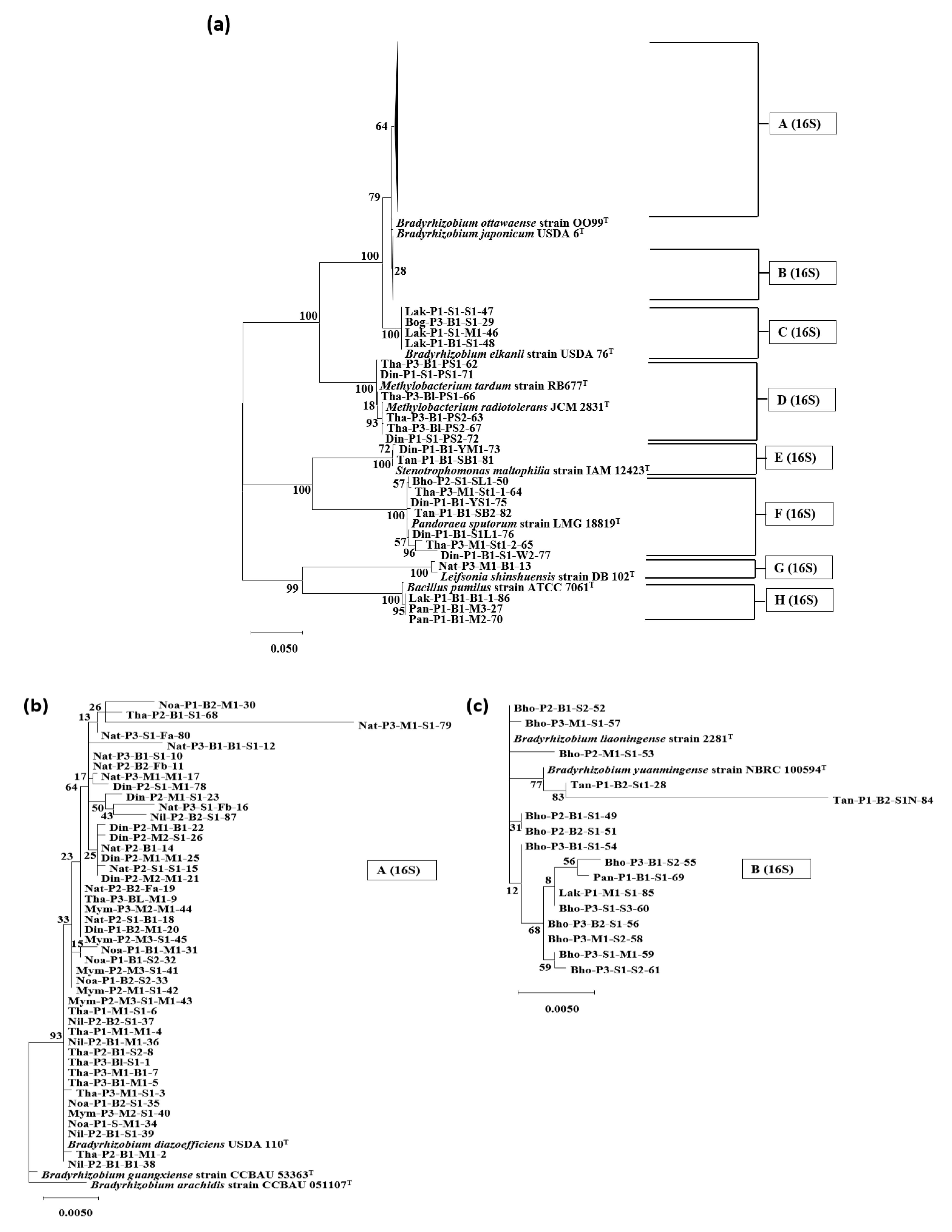
3.2. Phylogenetic Analysis Based on the 16S rRNA Genes
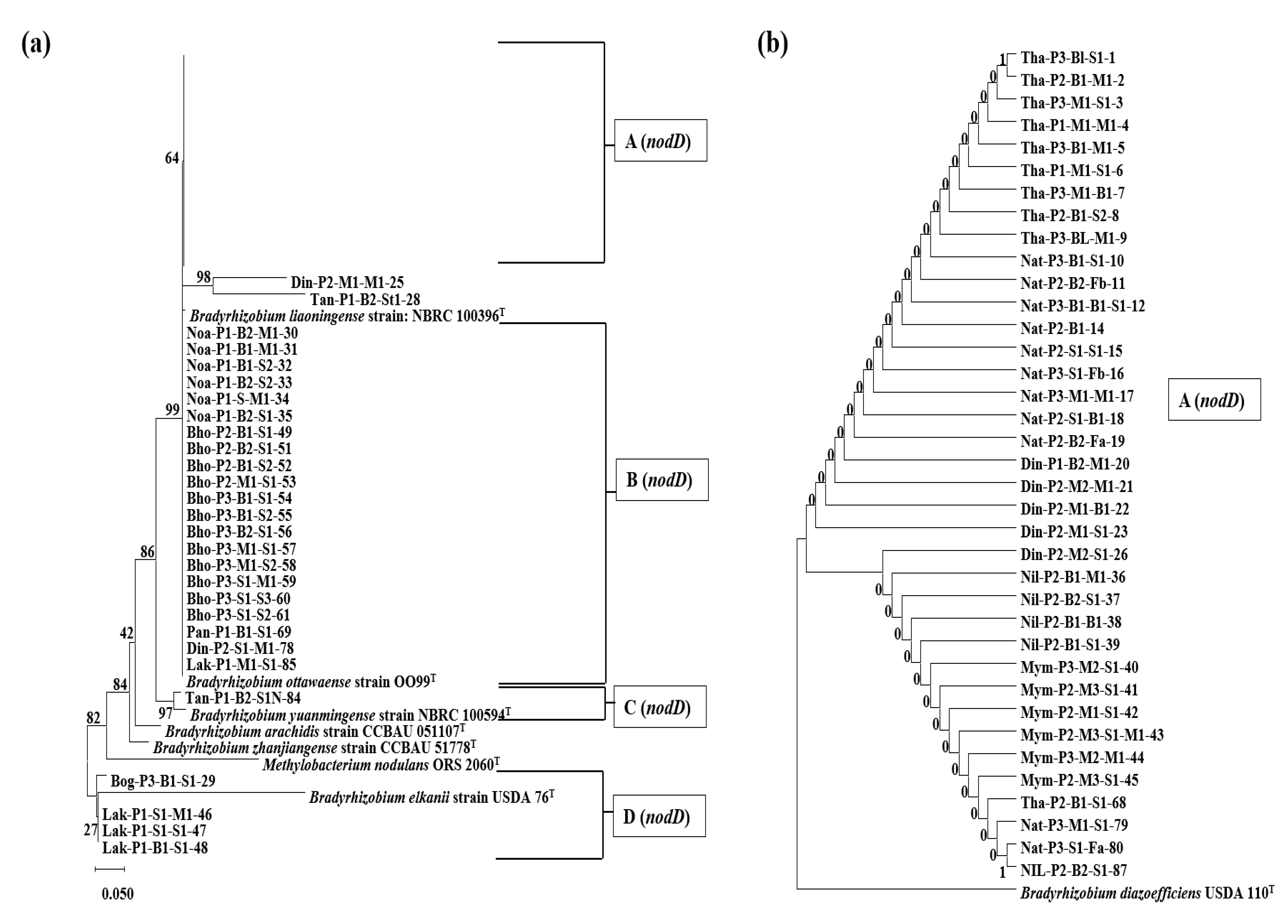
3.3. Phylogenetic Analysis Based on the nifH Gene
3.4. Phylogenetic Analysis Based on the nodD1 Gene
3.5. Selection of Isolates for Further Genetic and Physiological Characterization
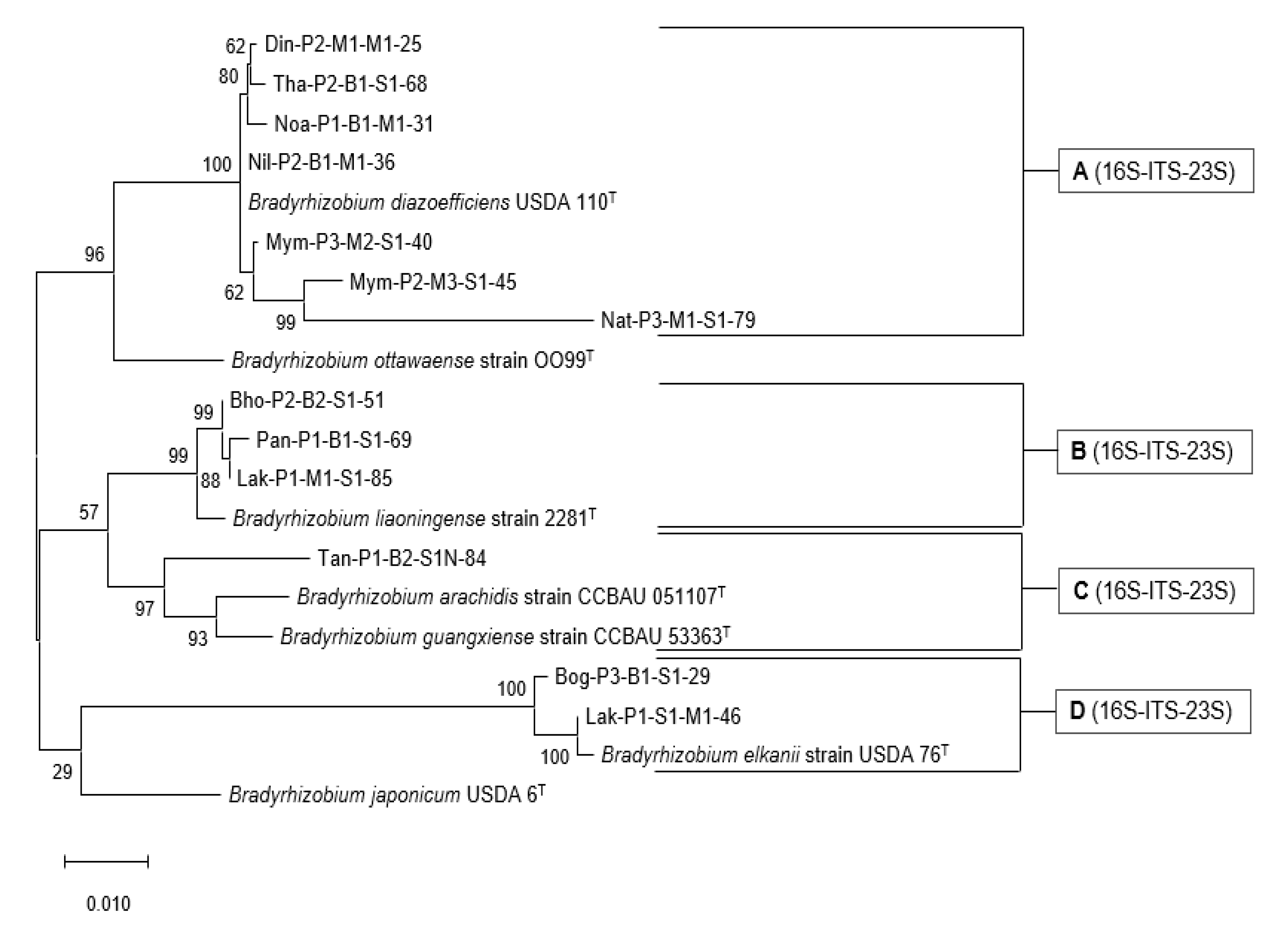
3.6. Phylogenetic Analysis of the rrn Operon (16S-ITS-23S)
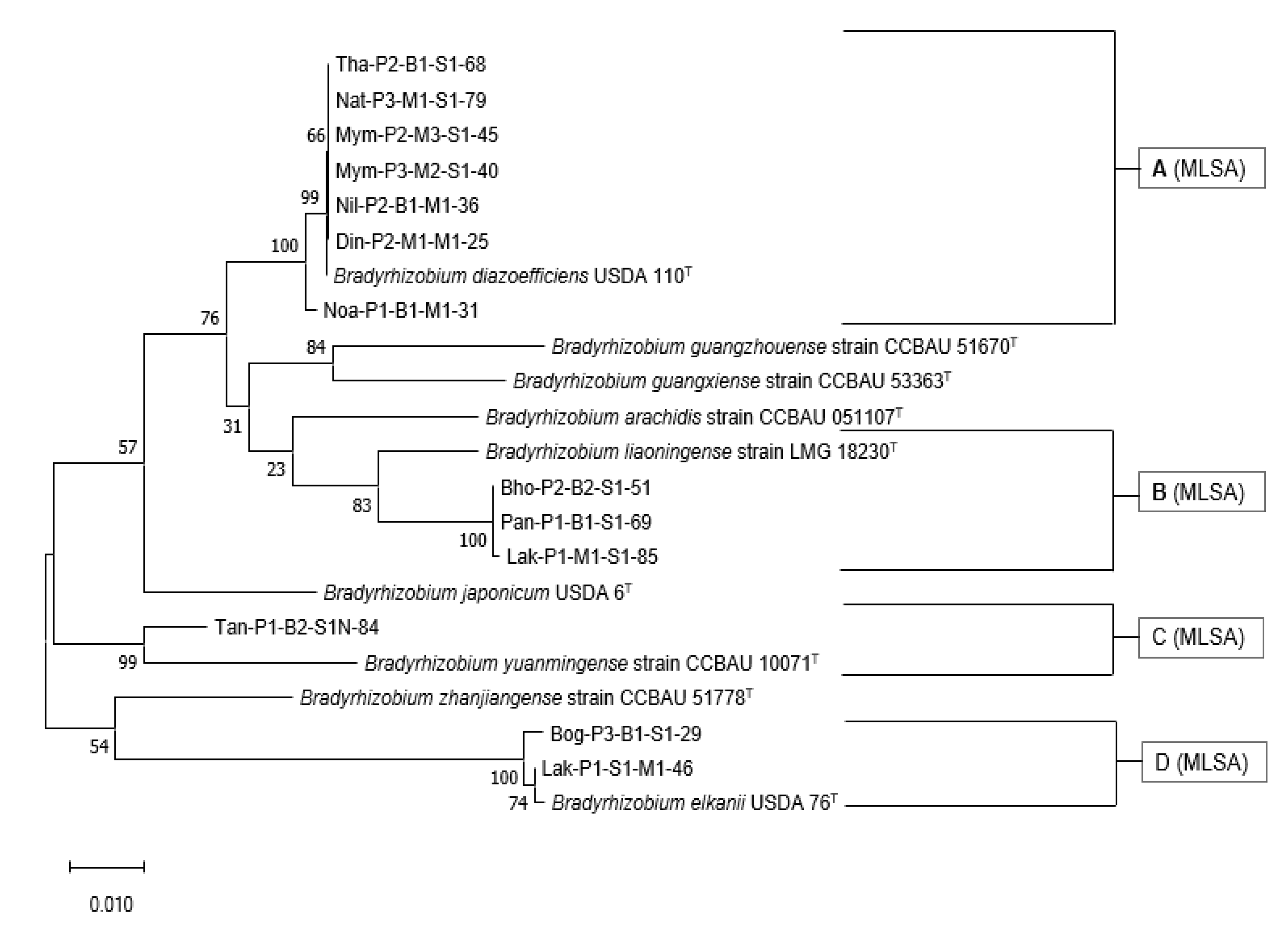
3.7. Multi-Locus Sequence Analysis of the Housekeeping Genes
3.8. Symbiotic Performances
3.9. Stress Tolerance Tests
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liu, M.; Shi, X.; Peng, K.; Xiao, F.; Sun, M.; Liu, X.; Shi, K.; Peng, F.; Xiao, Y. Effects of Root Zone Aeration on Soil Microbes Species in a Peach Tree Rhizosphere and Root Growth. Microorganisms 2022, 10, 1879. [Google Scholar] [CrossRef]
- Colwell, R.R. Microbial Diversity: The Importance of Exploration and Conservation. J. Ind. Microbiol. Biotechnol. 1997, 18, 302–307. [Google Scholar] [CrossRef] [PubMed]
- Wang, E.T.; Tian, C.F.; Chen, W.F.; Young, J.P.W.; Chen, W.X. Ecology and Evolution of Rhizobia; Springer: Singapore, 2019. [Google Scholar] [CrossRef]
- Martínez-Hidalgo, P.; Hirsch, A.M. The Nodule Microbiome: N2 fixing Rhizobia Do Not Live Alone. Phytobiomes J. 2017, 1, 70–82. [Google Scholar] [CrossRef] [Green Version]
- Wagner, S.C. Biological Nitrogen Fixation. Nat. Educ. Knowl. 2011, 3, 15. [Google Scholar]
- Binde, D.R.; Menna, P.; Bangel, E.V.; Barcellos, F.G.; Hungria, M. Rep-PCR Fingerprinting and Taxonomy Based on the Sequencing of the 16S RRNA Gene of 54 Elite Commercial Rhizobial Strains. Appl. Microbiol. Biotechnol. 2009, 83, 897–908. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.F.; Wang, E.T.; Ji, Z.J.; Zhang, J.J. Recent Development and New Insight of Diversification and Symbiosis Specificity of Legume Rhizobia: Mechanism and Application. J. Appl. Microbiol. 2021, 131, 553–563. [Google Scholar] [CrossRef]
- Cevallos, M.A.; Degli Esposti, M. New Alphaproteobacteria Thrive in the Depths of the Ocean with Oxygen Gradient. Microorganisms 2022, 10, 455. [Google Scholar] [CrossRef]
- Moulin, L.; Munive, A.; Dreyfus, B.; Boivin-Masson, C. Nodulation of Legumes by Members of the Beta-Subclass of Proteobacteria. Nature 2001, 411, 948–950. [Google Scholar] [CrossRef]
- Shiraishi, A.; Matsushita, N.; Hougetsu, T. Nodulation in Black Locust by the Gammaproteobacteria Pseudomonas Sp. and the Betaproteobacteria Burkholderia sp. Syst. Appl. Microbiol. 2010, 33, 269–274. [Google Scholar] [CrossRef]
- Estrada-de los Santos, P.; Palmer, M.; Chávez-Ramírez, B.; Beukes, C.; Steenkamp, E.; Briscoe, L.; Khan, N.; Maluk, M.; Lafos, M.; Humm, E.; et al. Whole Genome Analyses Suggests That Burkholderia Sensu Lato Contains Two Additional Novel Genera (Mycetohabitans gen. nov., and Trinickia gen. nov.): Implications for the Evolution of Diazotrophy and Nodulation in the Burkholderiaceae. Genes 2018, 9, 389. [Google Scholar] [CrossRef]
- Iturralde, E.T.; Covelli, J.M.; Alvarez, F.; Pérez-Giménez, J.; Arrese-Igor, C.; Lodeiro, A.R. Soybean-Nodulating Strains with Low Intrinsic Competitiveness for Nodulation, Good Symbiotic Performance, and Stress-Tolerance Isolated from Soybean-Cropped Soils in Argentina. Front. Microbiol. 2019, 10, 1061. [Google Scholar] [CrossRef] [PubMed]
- Jaiswal, S.K.; Dakora, F.D. Widespread Distribution of Highly Adapted Bradyrhizobium Species Nodulating Diverse Legumes in Africa. Front. Microbiol. 2019, 10, 310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.M.; Li, Y.; Chen, W.F.; Wang, E.T.; Tian, C.F.; Li, Q.Q.; Zhang, Y.Z.; Sui, X.H.; Chen, W.X. Biodiversity and Biogeography of Rhizobia Associated with Soybean Plants Grown in the North China Plain. Appl. Environ. Microbiol. 2011, 77, 6331–6342. [Google Scholar] [CrossRef] [Green Version]
- Mousavi, S.A.; Young, J.P.W. International Committee on Systematics of Prokaryotes, Subcommittee on the Taxonomy of Rhizobia and Agrobacteria, Minutes of the Annual Meeting by Videoconference, 5 July 2021, Followed by Online Discussion until 31 December 2021. Int. J. Syst. Evol. Microbiol. 2022, 72, 005453. [Google Scholar] [CrossRef]
- Parte, A.C.; Carbasse, J.S.; Meier-Kolthoff, J.P.; Reimer, L.C.; Göker, M. List of Prokaryotic Names with Standing in Nomenclature (LPSN) Moves to the DSMZ. Int. J. Syst. Evol. Microbiol. 2020, 70, 5607–5612. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Wang, E.; Wang, S.; Li, Y.; Chen, X.; Li, Y. Characteristics of Rhizobium tianshanense sp. nov., a Moderately and Slowly Growing Root Nodule Bacterium Isolated from an Arid Saline Environment in Xinjiang, People’s Republic of China. Int. J. Syst. Bacteriol. 1995, 45, 153–159. [Google Scholar] [CrossRef] [Green Version]
- Tian, C.F.; Zhou, Y.J.; Zhang, Y.M.; Li, Q.Q.; Zhang, Y.Z.; Li, D.F.; Wang, S.; Wang, J.; Gilbert, L.B.; Li, Y.R.; et al. Comparative Genomics of Rhizobia Nodulating Soybean Suggests Extensive Recruitment of Lineage-Specific Genes in Adaptations. Proc. Natl. Acad. Sci. USA 2012, 109, 8629–8634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, K.; Reckling, M.; Ramirez, M.D.A.; Djedidi, S.; Fukuhara, I.; Ohyama, T.; Yokoyama, T.; Bellingrath-Kimura, S.D.; Halwani, M.; Egamberdieva, D.; et al. Characterization of Rhizobia for the Improvement of Soybean Cultivation at Cold Conditions in Central Europe. Microbes Environ. 2020, 35, ME19124. [Google Scholar] [CrossRef] [Green Version]
- Somasegaran, P.; Hoben, H.J. Handbook for Rhizobia: Methods in Legume-Rhizobium Technology; Springer-Verlag: New York, NY, USA, 1994. [Google Scholar]
- Somasegaran, P.; Hoben, H.J.; Somasegaran, P.; Hoben, H.J. Quantifying the Growth of Rhizobia. In Handbook for Rhizobia; Springer: New York, NY, USA, 1994; pp. 47–57. [Google Scholar] [CrossRef]
- Djedidi, S.; Yokoyama, T.; Ohkama-Ohtsu, N.; Risal, C.P.; Abdelly, C.; Sekimoto, H. Stress Tolerance and Symbiotic and Phylogenic Features of Root Nodule Bacteria Associated with Medicago Species in Different Bioclimatic Regions of Tunisia. Microbes Environ. 2011, 26, 36–45. [Google Scholar] [CrossRef]
- Risal, C.P.; Yokoyama, T.; Ohkama-Ohtsu, N.; Djedidi, S.; Sekimoto, H. Genetic Diversity of Native Soybean Bradyrhizobia from Different Topographical Regions along the Southern Slopes of the Himalayan Mountains in Nepal. Syst. Appl. Microbiol. 2010, 33, 416–425. [Google Scholar] [CrossRef]
- van Berkum, P.; Fuhrmann, J.J. Evolutionary Relationships among the Soybean Bradyrhizobia Reconstructed from 16S RRNA Gene and Internally Transcribed Spacer Region Sequence Divergence. Int. J. Syst. Evol. Microbiol. 2000, 50 Pt 6, 2165–2172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mortuza, M.F.; Tomooka, N.; Habibi, S.; Akatsu, T.; Djedidi, S.; Naito, K.; Sekimoto, H.; Okazaki, S.; Ohkama-Ohtsu, N.; Yokoyama, T. Multiphase Characterization of Wild Vigna Associated Root Nodule Bacteria from Japanese Subtropical Islands Unveiled Novel High Temperature Resistant Bradyrhizobium Strains Having High Symbiotic Compatibility with Soybean and Mungbean. Soil Sci. Plant Nutr. 2020, 66, 285–298. [Google Scholar] [CrossRef]
- Laguerre, G.; Nour, S.M.; Macheret, V.; Sanjuan, J.; Drouin, P.; Amarger, N. Classification of Rhizobia Based on NodC and NifH Gene Analysis Reveals a Close Phylogenetic Relationship among Phaseolus vulgaris Symbionts. Microbiology 2001, 147, 981–993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martens, M.; Dawyndt, P.; Coopman, R.; Gillis, M.; De Vos, P.; Willems, A. Advantages of Multilocus Sequence Analysis for Taxonomic Studies: A Case Study Using 10 Housekeeping Genes in the Genus Ensifer (Including Former Sinorhizobium). Int. J. Syst. Evol. Microbiol. 2008, 58, 200–214. [Google Scholar] [CrossRef] [Green Version]
- Vinuesa, P.; Rojas-Jiménez, K.; Contreras-Moreira, B.; Mahna, S.K.; Prasad, B.N.; Moe, H.; Selvaraju, S.B.; Thierfelder, H.; Werner, D. Multilocus Sequence Analysis for Assessment of the Biogeography and Evolutionary Genetics of Four Bradyrhizobium Species That Nodulate Soybeans on the Asiatic Continent. Appl. Environ. Microbiol. 2008, 74, 6987–6996. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Habibi, S.; Djedidi, S.; Prongjunthuek, K.; Mortuza, M.F.; Ohkama-Ohtsu, N.; Sekimoto, H.; Yokoyoma, T. Physiological and Genetic Characterization of Rice Nitrogen Fixer PGPR Isolated from Rhizosphere Soils of Different Crops. Plant Soil 2014, 379, 51–66. [Google Scholar] [CrossRef]
- Woliy, K.; Degefu, T.; Frostegård, Å. Host Range and Symbiotic Effectiveness of N2O Reducing Bradyrhizobium Strains. Front. Microbiol. 2019, 10, 2746. [Google Scholar] [CrossRef]
- Naamala, J.; Jaiswal, S.K.; Dakora, F.D. Microsymbiont Diversity and Phylogeny of Native Bradyrhizobia Associated with Soybean (Glycine max L. Merr.) Nodulation in South African Soils. Syst. Appl. Microbiol. 2016, 39, 336–344. [Google Scholar] [CrossRef]
- Appunu, C.; N’Zoue, A.; Laguerre, G. Genetic Diversity of Native Bradyrhizobia Isolated from Soybeans (Glycine max L.) in Different Agricultural-Ecological-Climatic Regions of India. Appl. Environ. Microbiol. 2008, 74, 5991–5996. [Google Scholar] [CrossRef] [Green Version]
- Ardley, J.K.; Reeve, W.G.; O’Hara, G.W.; Yates, R.J.; Dilworth, M.J.; Howieson, J.G. Nodule Morphology, Symbiotic Specificity and Association with Unusual Rhizobia Are Distinguishing Features of the Genus Listia within the Southern African Crotalarioid Clade Lotononis s.l. Ann. Bot. 2013, 112, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madhaiyan, M.; Poonguzhali, S.; Senthilkumar, M.; Sundaram, S.; Sa, T. Nodulation and Plant-Growth Promotion by Methylotrophic Bacteria Isolated from Tropical Legumes. Microbiol. Res. 2009, 164, 114–120. [Google Scholar] [CrossRef] [PubMed]
- Renier, A.; Jourand, P.; Rapior, S.; Poinsot, V.; Sy, A.; Dreyfus, B.; Moulin, L. Symbiotic Properties of Methylobacterium Nodulans ORS 2060T: A Classic Process for an Atypical Symbiont. Soil Biol. Biochem. 2008, 40, 1404–1412. [Google Scholar] [CrossRef]
- Sy, A.; Giraud, E.; Jourand, P.; Garcia, N.; Willems, A.; De Lajudie, P.; Prin, Y.; Neyra, M.; Gillis, M.; Boivin-Masson, C.; et al. Methylotrophic Methylobacterium Bacteria Nodulate and Fix Nitrogen in Symbiosis with Legumes. J. Bacteriol. 2001, 183, 214–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dobritsa, A.P.; Samadpour, M. Transfer of Eleven Species of the Genus Burkholderia to the Genus Paraburkholderia and Proposal of Caballeronia gen. nov. to Accommodate Twelve Species of the Genera Burkholderia and Paraburkholderia. Int. J. Syst. Evol. Microbiol. 2016, 66, 2836–2846. [Google Scholar] [CrossRef]
- Gyaneshwar, P.; Hirsch, A.M.; Moulin, L.; Chen, W.M.; Elliott, G.N.; Bontemps, C.; De Los Santos, P.E.; Gross, E.; Dos Reis, F.B.; Janet, I.S.; et al. Legume-Nodulating Betaproteobacteria: Diversity, Host Range, and Future Prospects. Mol. Plant-Microbe Interact. 2011, 24, 1276–1288. [Google Scholar] [CrossRef] [Green Version]
- Hoque, M.S.; Broadhurst, L.M.; Thrall, P.H. Genetic Characterization of Root-Nodule Bacteria Associated with Acacia salicina and A. stenophylla (Mimosaceae) across South-Eastern Australia. Int. J. Syst. Evol. Microbiol. 2011, 61, 299–309. [Google Scholar] [CrossRef] [Green Version]
- Bai, Y.; D’Aoust, F.; Smith, D.L.; Driscoll, B.T. Isolation of Plant-Growth-Promoting Bacillus Strains from Soybean Root Nodules. Can. J. Microbiol. 2002, 48, 230–238. [Google Scholar] [CrossRef]
- Ampomah, O.Y.; Huss-Danell, K. Genetic Diversity of Root Nodule Bacteria Nodulating Lotus corniculatus and Anthyllis vulneraria in Sweden. Syst. Appl. Microbiol. 2011, 34, 267–275. [Google Scholar] [CrossRef]
- Palaniappan, P.; Chauhan, P.S.; Saravanan, V.S.; Anandham, R.; Sa, T. Isolation and Characterization of Plant Growth Promoting Endophytic Bacterial Isolates from Root Nodule of Lespedeza sp. Biol. Fertil. Soils 2010, 46, 807–816. [Google Scholar] [CrossRef]
- Barnawal, D.; Bharti, N.; Maji, D.; Chanotiya, C.S.; Kalra, A. ACC Deaminase-Containing Arthrobacter protophormiae Induces NaCl Stress Tolerance through Reduced ACC Oxidase Activity and Ethylene Production Resulting in Improved Nodulation and Mycorrhization in Pisum sativum. J. Plant Physiol. 2014, 171, 884–894. [Google Scholar] [CrossRef]
- Sreevidya, M.; Gopalakrishnan, S.; Kudapa, H.; Varshney, R.K. Exploring Plant Growth-Promotion Actinomycetes from Vermicompost and Rhizosphere Soil for Yield Enhancement in Chickpea. Brazilian J. Microbiol. 2016, 47, 85–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tokala, R.K.; Strap, J.L.; Jung, C.M.; Crawford, D.L.; Salove, M.H.; Deobald, L.A.; Bailey, J.F.; Morra, M.J. Novel Plant-Microbe Rhizosphere Interaction Involving Streptomyces lydicus WYEC108 and the Pea Plant (Pisum sativum). Appl. Environ. Microbiol. 2002, 68, 2161–2171. [Google Scholar] [CrossRef] [Green Version]
- Moulin, L.; Béna, G.; Boivin-Masson, C.; Stepkowski, T. Phylogenetic Analyses of Symbiotic Nodulation Genes Support Vertical and Lateral Gene Co-Transfer within the Bradyrhizobium Genus. Mol. Phylogenet. Evol. 2004, 30, 720–732. [Google Scholar] [CrossRef]
- Gaby, J.C.; Buckley, D.H. A Comprehensive Aligned NifH Gene Database: A Multipurpose Tool for Studies of Nitrogen-Fixing Bacteria. Database 2014, 2014, bau001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cuscó, A.; Catozzi, C.; Viñes, J.; Sanchez, A.; Francino, O. Microbiota Profiling with Long Amplicons Using Nanopore Sequencing: Full-Length 16S RRNA Gene and the 16S-ITS-23S of the Rrn Operon. F1000Research 2019, 7, 1755. [Google Scholar] [CrossRef]
- Nygaard, A.B.; Tunsjø, H.S.; Meisal, R.; Charnock, C. A Preliminary Study on the Potential of Nanopore MinION and Illumina MiSeq 16S RRNA Gene Sequencing to Characterize Building-Dust Microbiomes. Sci. Rep. 2020, 10, 3209. [Google Scholar] [CrossRef] [Green Version]
- De Lajudie, P.M.; Andrews, M.; Ardley, J.; Eardly, B.; Jumas-Bilak, E.; Kuzmanović, N.; Lassalle, F.; Lindström, K.; Mhamdi, R.; Martínez-Romero, E.; et al. Minimal Standards for the Description of New Genera and Species of Rhizobia and Agrobacteria. Int. J. Syst. Evol. Microbiol. 2019, 69, 1852–1863. [Google Scholar] [CrossRef]
- Graham, P.H.; Hungria, M.; Tlusty, B. Breeding for Better Nitrogen Fixation in Grain Legumes: Where Do the Rhizobia Fit In? Crop Manag. 2004, 3, 1–6. [Google Scholar] [CrossRef]
- Allito, B.B.; Ewusi-Mensah, N.; Logah, V.; Hunegnaw, D.K. Legume-Rhizobium Specificity Effect on Nodulation, Biomass Production and Partitioning of Faba Bean (Vicia faba L.). Sci. Rep. 2021, 11, 3678. [Google Scholar] [CrossRef]
- Habibi, S.; Ayubi, A.G.; Ohkama-Ohtsu, N.; Sekimoto, H.; Yokoyama, T. Genetic Characterization of Soybean Rhizobia Isolated from Different Ecological Zones in North-Eastern Afghanistan. Microbes Environ. 2017, 32, 71–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, R.; Zhang, J.; Huan, H.; Bai, C.; Chen, Z.; Liu, G. High Salt Tolerance of a Bradyrhizobium Strain and Its Promotion of the Growth of Stylosanthes guianensis. Int. J. Mol. Sci. 2017, 18, 1625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.S.; Figueredo, A.; Pedrosa, F.O.; Hungria, M. Genetic Characterization of Soybean Rhizobia in Paraguay. Appl. Environ. Microbiol. 2000, 66, 5099–5103. [Google Scholar] [CrossRef]
- Karanja, N.K.; Wood, M. Selecting Rhizobium phaseoli Strains for Use with Beans (Phaseolus vulgaris L.) in Kenya: Tolerance of High Temperature and Antibiotic Resistance. Plant Soil 1988, 112, 15–22. [Google Scholar] [CrossRef]
- Zhang, J.; Singh, D.; Guo, C.; Shang, Y.; Peng, S. Rhizobia at Extremes of Acidity, Alkalinity, Salinity, and Temperature. In Microbial Versatility in Varied Environments; Springer: Singapore, 2020; pp. 51–65. [Google Scholar] [CrossRef]
- Kulkarni, S.; Surange, S.; Shekhar Nautiyal, C. Crossing the Limits of Rhizobium Existence in Extreme Conditions. Curr. Microbiol. 2000, 41, 402–409. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Name of Site | Collection Date (dd/mm/yy) | Soil Type | Crop History | Soil pH | Latitude | Longitude | Soybean Cultivation |
|---|---|---|---|---|---|---|---|---|
| 1 | Bhola | 4 February 2018 | Clay loam | Soybean–rice–rice–soybean–rice–rice | 6.85 | 22.6883 | 90.5975 | Yes |
| 2 | Bogra | 2 February 2018 | Loamy | Mustard–rice–rice | 6.92 | 24.8881 | 89.3869 | No |
| 3 | Dinajpur | 5 February 2018 | Sandy loam | Soybean–rice–potato–vegetable | 6.30 | 25.8373 | 88.4794 | Yes |
| 4 | Lakshmipur | 6 February 2018 | Sandy loam | Soybean–rice–soybean–rice–so–bean | 6.19 | 22.7097 | 90.9952 | Yes |
| 5 | Mymensingh | 4 February 2018 | Loamy | Soybean–soybean–soybean | 6.49 | 24.7244 | 90.4300 | Yes |
| 6 | Natore | 2 February 2018 | Loamy | Soybean–rice–soybean–rice | 6.41 | 24.3720 | 88.8991 | Yes |
| 7 | Nilphamary | 28 January 2018 | Sandy loam | Corn–rice–jute–soybean | 6.87 | 26.1102 | 88.8506 | Yes |
| 8 | Noakhali | 6 February 2018 | Sandy loam | Soybean–rice–soybean–rice–soybean | 6.68 | 22.7388 | 91.0677 | Yes |
| 9 | Panchagarh | 5 February 2018 | Sandy loam | Soybean–rice–groundnut–rice–groundnut | 6.04 | 26.2440 | 88.5598 | Yes |
| 10 | Tangail | 8 February 2018 | loamy | Mustard–jute–rice–mustard–jute–rice | 6.04 | 24.2341 | 89.8598 | No |
| 11 | Thakurgaon | 3 February 2018 | Loamy | Mustard–rice–soybean–rice | 6.39 | 25.9208 | 88.4702 | Yes |
| Enrei Variety | Binasoybean-3 Variety | |||
|---|---|---|---|---|
| Bacteria Name | Medium Size Nodule | Small Size Nodule | Medium Size Nodule | Small Size Nodule |
| USDA110 | 10.00 ± 1.00 | 9.00 ± 6.93 | 16.33 ± 3.22 | 7.67 ± 4.04 |
| Bho-P2-B2-S1-51 | 13.00 ± 5.57 | 5.00 ± 4.36 | 19.33 ± 4.73 | 1.33 ± 0.58 |
| Nat-P3-M1-S1-79 | 12.67 ± 2.08 | 6.67 ± 2.89 | 14.00 ± 4.36 | 11.33 ± 4.73 |
| Lak-P1-M1-S1-85 | 10.33 ± 1.53 | 5.00 ± 3.61 | 12.33 ± 1.53 | 1.67 ± 0.58 |
| Lak-P1-S1-M1-46 | 9.67 ± 3.51 | 12.00 ± 5.20 | 11.67 ± 4.73 | 3.00 ± 3.00 |
| Tha-P2-B1-S1-68 | 8.67 ± 1.16 | 5.33 ± 4.51 | 16.33 ± 7.02 | 11.67 ± 13.28 |
| Nil-P2-B1-M1-36 | 8.33 ± 1.53 | 7.33 ± 0.58 | 13.67 ± 3.79 | 5.00 ± 2.65 |
| Pan-P1-B1-S1-69 | 7.67 ± 2.08 | 6.33 ± 4.73 | 12.33 ± 3.22 | 1.67 ± 2.89 |
| Din-P2-M1-M1-25 | 7.33 ± 2.08 | 13.00 ± 2.65 | 11.00 ± 3.00 | 14.33 ± 8.08 |
| Mym-P2-M3-S1-45 | 7.00 ± 4.00 | 14.33 ± 16.65 | 14.33 ± 3.79 | 8.00 ± 4.36 |
| Mym-P3-M2-S1-40 | 6.67 ± 1.16 | 4.00 ± 1.73 | 13.00 ± 6.93 | 10.67 ± 11.72 |
| Noa-P1-B1-M1-31 | 4.00 ± 1.00 | 1.00 ± 1.73 | 13.00 ± 1.00 | 7.00 ± 4.00 |
| Bog-P3-B1-S1-29 | 1.00 ± 0.00 | 6.67 ± 7.64 | 10.33 ± 3.06 | 13.00 ± 11.27 |
| Tan-P1-B2-S1N-84 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.33 ± 0.58 | 11.00 ± 12.29 |
| Enrei Variety | Binasoybean-3 Variety | |||
|---|---|---|---|---|
| Bacteria Name | Biomass DW (g) | ARA (µmol/h/g Nodule DW) | Biomass DW (g) | ARA (µmol/h/g Nodule DW) |
| USDA110 | 0.95 ± 0.04 | 22.37 ± 20.18 | 0.52 ± 0.04 | 35.83 ± 12.78 |
| Bho-P2-B2-S1-51 | 0.87 ± 0.20 | 109.76 * ± 7.04 | 0.57 ± 0.04 | 86.77 * ± 27.27 |
| Bog-P3-B1-S1-29 | 0.96 ± 0.05 | 50.83 ± 88.41 | 0.50 ± 0.02 | 44.98 ± 11.93 |
| Din-P2-M1-M1-25 | 1.02 ± 0.02 | 21.74 ± 29.17 | 0.46 ± 0.12 | 42.59 ± 31.99 |
| Lak-P1-M1-S1-85 | 0.96 ± 0.06 | 18.71 ± 28.87 | 0.54 ± 0.07 | 64.66 ± 16.53 |
| Lak-P1-S1-M1-46 | 0.95 ± 0.06 | 44.53 ± 41.29 | 0.49 ± 0.07 | 24.95 ± 12.07 |
| Mym-P2-M3-S1-45 | 0.93 ± 0.03 | 30.92 ± 27.52 | 0.52 ± 0.05 | 21.80 ± 9.19 |
| Mym-P3-M2-S1-40 | 0.89 ± 0.04 | 60.12 ± 20.58 | 0.50 ± 0.04 | 28.98 ± 7.37 |
| Nat-P3-M1-S1-79 | 0.95 ± 0.11 | 12.53 ± 21.65 | 0.51 ± 0.10 | 69.23 ± 21.22 |
| Nil-P2-B1-M1-36 | 0.98 ± 0.02 | 36.72 ± 39.18 | 0.60 ± 0.00 | 19.55 ± 11.68 |
| Noa-P1-B1-M1-31 | 0.92 ± 0.07 | 5.01 ± 4.48 | 0.57 ± 0.06 | 24.58 ± 2.93 |
| Pan-P1-B1-S1-69 | 0.79 ± 0.01 | 60.53 ± 60.33 | 0.59 ± 0.04 | 48.34 ± 21.62 |
| Tan-P1-B2-S1N-84 | 0.85 ± 0.08 | 0.00 ± 0.00 | 0.50 ± 0.04 | 6.43 ± 5.84 |
| Tha-P2-B1-S1-68 | 0.96 ± 0.08 | 30.77 ± 51.64 | 0.57 ± 0.05 | 13.20 ± 6.80 |
| NaCl | Temperature | pH | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 0% | 1% | 2% | 3% | 4% | 10 °C | 20 °C | 28 °C | 37 °C | 40 °C | 45 °C | 4.5 | 6 | 7 | 8.5 | 10 | |
| USDA110 | (+) | (+) | (+) | (+) | (−) | (−) | (+) | (+) | (+) | (−) | (−) | (+) | (+) | (+) | (+) | (+) |
| Bho-P2-B2-S1-51 | (+) | (+) | (+) | (+) | (+) | (−) | (+) | (+) | (+) | (+) | (+) | (+) | (+) | (+) | (+) | (+) |
| Bog-P3-B1-S1-29 | (+) | (+) | (+) | (+) | (+) | (−) | (+) | (+) | (+) | (+) | (+) | (+) | (+) | (+) | (+) | (+) |
| Din-P2-M1-M1-25 | (+) | (+) | (+) | (+) | (+) | (−) | (+) | (+) | (+) | (+) | (−) | (+) | (+) | (+) | (+) | (+) |
| Lak-P1-M1-S1-85 | (+) | (+) | (+) | (+) | (+) | (−) | (+) | (+) | (+) | (+) | (−) | (+) | (+) | (+) | (+) | (+) |
| Lak-P1-S1-M1-46 | (+) | (+) | (+) | (+) | (+) | (+) | (+) | (+) | (+) | (+) | (−) | (+) | (+) | (+) | (+) | (+) |
| Mym-P2-M3-S1-45 | (+) | (+) | (+) | (+) | (+) | (−) | (+) | (+) | (+) | (+) | (+) | (+) | (+) | (+) | (+) | (+) |
| Mym-P3-M2-S1-40 | (+) | (+) | (+) | (+) | (+) | (−) | (+) | (+) | (+) | (+) | (−) | (−) | (+) | (+) | (+) | (+) |
| Nat-P3-M1-S1-79 | (+) | (+) | (+) | (+) | (+) | (−) | (+) | (+) | (+) | (+) | (+) | (+) | (+) | (+) | (+) | (+) |
| Nil-P2-B1-M1-36 | (+) | (+) | (+) | (+) | (+) | (−) | (+) | (+) | (+) | (+) | (−) | (+) | (+) | (+) | (+) | (+) |
| Noa-P1-B1-M1-31 | (+) | (+) | (+) | (+) | (+) | (−) | (+) | (+) | (+) | (+) | (−) | (−) | (+) | (+) | (+) | (+) |
| Pan-P1-B1-S1-69 | (+) | (+) | (+) | (+) | (+) | (+) | (+) | (+) | (+) | (+) | (−) | (+) | (+) | (+) | (+) | (+) |
| Tan-P1-B2-S1N-84 | (+) | (+) | (+) | (+) | (+) | (+) | (+) | (+) | (+) | (+) | (−) | (+) | (+) | (+) | (+) | (+) |
| Tha-P2-B1-S1-68 | (+) | (+) | (+) | (+) | (+) | (−) | (+) | (+) | (+) | (+) | (+) | (+) | (+) | (+) | (+) | (+) |
| Bacteria Name | 16S rRNA Gene Similarity | ITS Region Similarity | 23S rRNA Gene Similarity | Symbiotic Genes Similarity | House-Keeping Gene Similarity | ||||
|---|---|---|---|---|---|---|---|---|---|
| nifH Gene Similarity | nodD1 Gene Similarity | atpD | glnII | gyrB | MLSA | ||||
| Bho-P2-B2-S1-51 | B. liaoningense | B. liaoningense | B. liaoningense | B. diazoefficiens | B. diazoefficiens | B. diazoefficiens | B. guangxiense | B. liaoningense | B. guangzhouense |
| Bog-P3-B1-S1-29 | B. elkanii | B. elkanii | B. elkanii | B. elkanii | B. zhanjiangense | B. japonicum | B. paxllaeri | B. elkanii | B. elkanii |
| Din-P2-M1-M1-25 | B. diazoefficiens | B. diazoefficiens | B. diazoefficiens | B. diazoefficiens | B. diazoefficiens | B. diazoefficiens | B. diazoefficiens | B. diazoefficiens | B. diazoefficiens |
| Lak-P1-M1-S1-85 | B. liaoningense | B. liaoningense | B. liaoningense | B. diazoefficiens | B. ottawaense | B. diazoefficiens | B. guangxiense | B. liaoningense | B. diazoefficiens |
| Lak-P1-S1-M1-46 | B. elkanii | B. elkanii | B. elkanii | B. elkanii | B. zhanjiangense | B. japonicum | B. paxllaeri | B. elkanii | B. elkanii |
| Mym-P2-M3-S1-45 | B. diazoefficiens | B. diazoefficiens | B. diazoefficiens | B. diazoefficiens | B. diazoefficiens | B. diazoefficiens | B. diazoefficiens | B. diazoefficiens | B. diazoefficiens |
| Mym-P3-M2-S1-40 | B. diazoefficiens | B. diazoefficiens | B. diazoefficiens | B. diazoefficiens | B. diazoefficiens | B. diazoefficiens | B. diazoefficiens | B. diazoefficiens | B. diazoefficiens |
| Nat-P3-M1-S1-79 | B. diazoefficiens | B. diazoefficiens | B. diazoefficiens | B. diazoefficiens | B. diazoefficiens | B. diazoefficiens | B. diazoefficiens | B. diazoefficiens | B. diazoefficiens |
| Nil-P2-B1-M1-36 | B. diazoefficiens | B. diazoefficiens | B. diazoefficiens | B. diazoefficiens | B. diazoefficiens | B. diazoefficiens | B. diazoefficiens | B. diazoefficiens | B. diazoefficiens |
| Noa-P1-B1-M1-31 | B. diazoefficiens | B. diazoefficiens | B. diazoefficiens | B. diazoefficiens | B. diazoefficiens | B. diazoefficiens | B. diazoefficiens | B. diazoefficiens | B. diazoefficiens |
| Pan-P1-B1-S1-69 | B. liaoningense | B. liaoningense | B. liaoningense | B. diazoefficiens | B. ottawaense | B. guangzhouense | B. guangxiense | B. liaoningense | B. guangzhouense |
| Tan-P1-B2-S1N-84 | B. yuanmingense | B. arachidis | B. japonicum | B. yuanmingense | B. yuanmingense | B. diazoefficiens | B. arachidis | B. diversitatis | B. diazoefficiens |
| Tha-P2-B1-S1-68 | B. diazoefficiens | B. diazoefficiens | B. diazoefficiens | B. diazoefficiens | B. diazoefficiens | B. diazoefficiens | B. diazoefficiens | B. diazoefficiens | B. diazoefficiens |
| Name | Dependent Variable | Independent Variable | Pearson Correlation/ Standardized Coefficient Beta | Significance (p-Value) |
|---|---|---|---|---|
| Enrei | Biomass DW | Nodule dry weight | 0.344 | 0.026 * |
| Nodule number | 0.219 | 0.163 | ||
| Medium size nodule | 0.032 | 0.841 | ||
| Small size nodule | 0.251 | 0.108 | ||
| ARA (in µmol/h/g nodule DW) | −0.164 | 0.301 | ||
| ARA (in µmol/h/plant) | −0.097 | 0.539 | ||
| ARA (in µmol/h/plant) | Nodule dry weight | 0.325 | 0.035 * | |
| Nodule number | 0.240 | 0.125 | ||
| Medium size nodule | 0.293 | 0.060 | ||
| Small size nodule | 0.106 | 0.506 | ||
| Binasoybean-3 | Biomass DW | Nodule dry weight | 0.337 | 0.029 * |
| Nodule number | −0.088 | 0.578 | ||
| Medium size nodule | 0.343 | 0.026 * | ||
| Small size nodule | −0.323 | 0.037 * | ||
| ARA (in µmol/h/g nodule DW) | −0.101 | 0.524 | ||
| ARA (in µmol/h/plant) | 0.037 | 0.817 | ||
| ARA (in µmol/h/plant) | Nodule dry weight | 0.358 | 0.020 * | |
| Nodule number | 0.213 | 0.176 | ||
| Medium size nodule | 0.431 | 0.004 * | ||
| Small size nodule | −0.093 | 0.558 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mortuza, M.F.; Djedidi, S.; Ito, T.; Agake, S.-i.; Sekimoto, H.; Yokoyama, T.; Okazaki, S.; Ohkama-Ohtsu, N. Genetic and Physiological Characterization of Soybean-Nodule-Derived Isolates from Bangladeshi Soils Revealed Diverse Array of Bacteria with Potential Bradyrhizobia for Biofertilizers. Microorganisms 2022, 10, 2282. https://doi.org/10.3390/microorganisms10112282
Mortuza MF, Djedidi S, Ito T, Agake S-i, Sekimoto H, Yokoyama T, Okazaki S, Ohkama-Ohtsu N. Genetic and Physiological Characterization of Soybean-Nodule-Derived Isolates from Bangladeshi Soils Revealed Diverse Array of Bacteria with Potential Bradyrhizobia for Biofertilizers. Microorganisms. 2022; 10(11):2282. https://doi.org/10.3390/microorganisms10112282
Chicago/Turabian StyleMortuza, Md Firoz, Salem Djedidi, Takehiro Ito, Shin-ichiro Agake, Hitoshi Sekimoto, Tadashi Yokoyama, Shin Okazaki, and Naoko Ohkama-Ohtsu. 2022. "Genetic and Physiological Characterization of Soybean-Nodule-Derived Isolates from Bangladeshi Soils Revealed Diverse Array of Bacteria with Potential Bradyrhizobia for Biofertilizers" Microorganisms 10, no. 11: 2282. https://doi.org/10.3390/microorganisms10112282