

Gender-Difference in Hair Length as Revealed by Crispr-Based Production of Long-Haired Mice with Dysfunctional FGF5 Mutations
 , , and
, , and 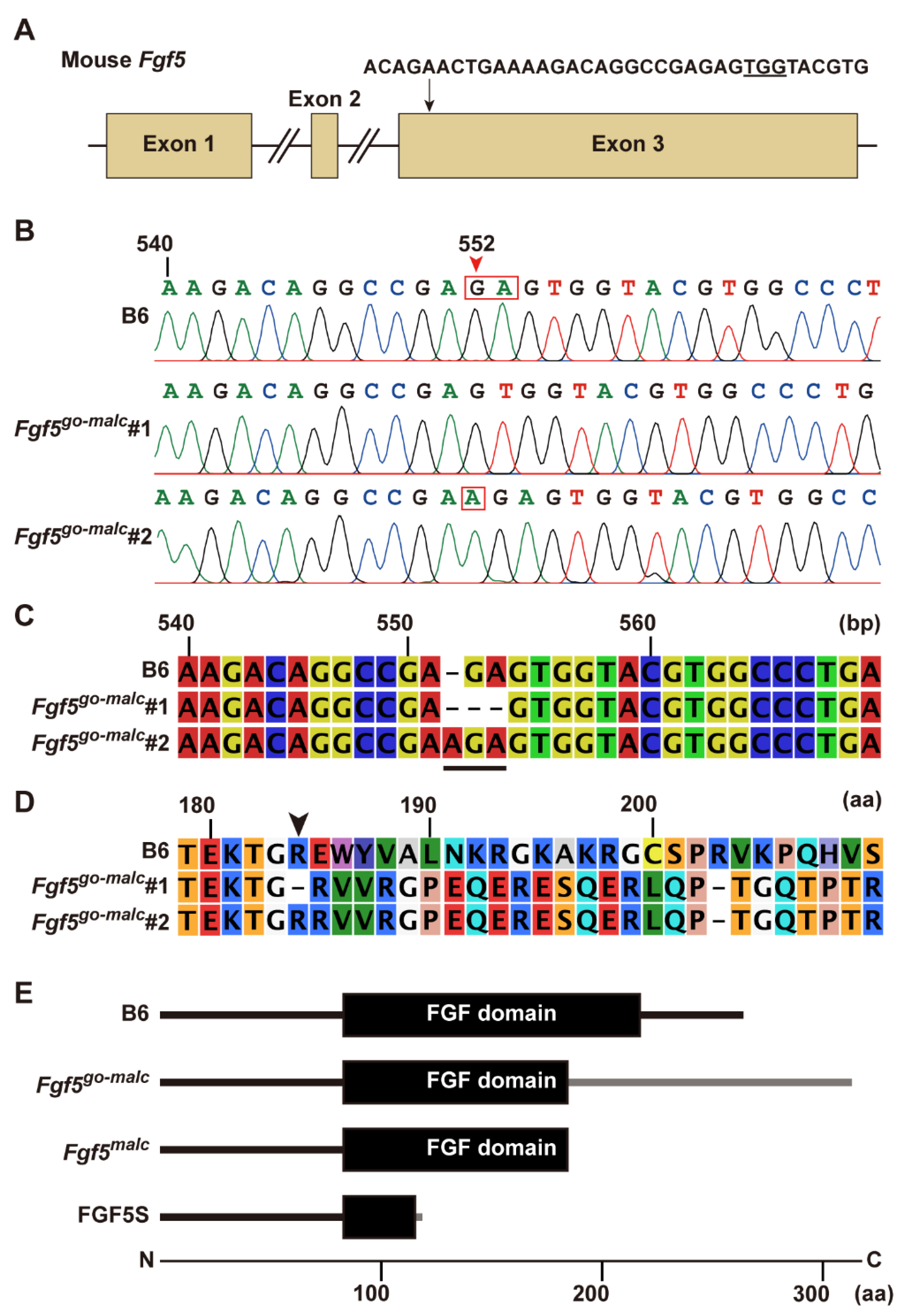{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Generation of Fgf5 Mutant Mice
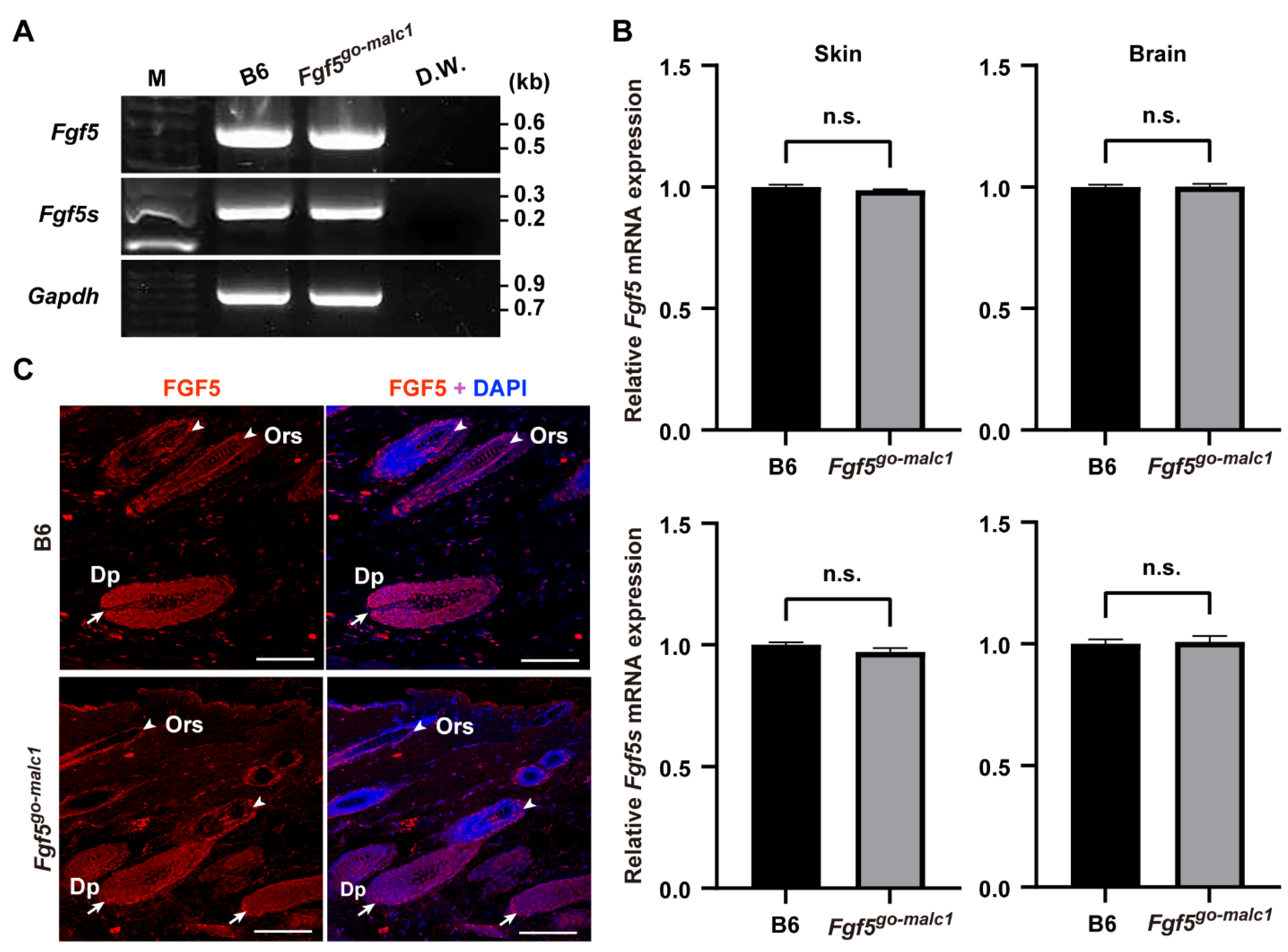
2.2. Analysis of Mutant FGF5 Protein in Fgf5go-malc1 Mice
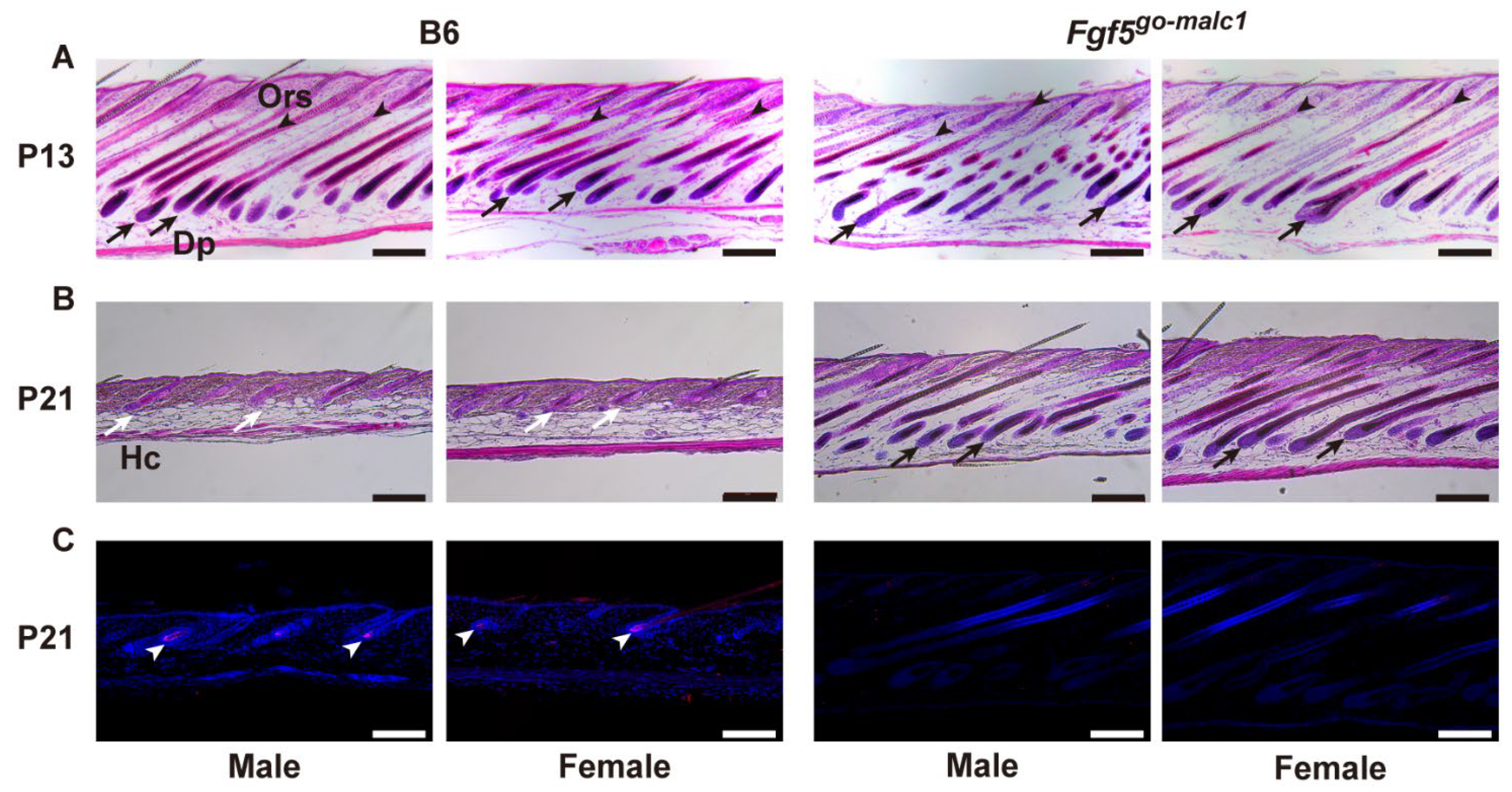
2.3. Prolonged Anagen Phase in Fgf5go-malc1 Mice
3. Discussion
4. Materials and Methods
4.1. Mice
4.2. Generation of Fgf5go-malc Mice
4.3. Measurement of Hair Length and Hair Follicles
4.4. Quantitative RT-PCR
4.5. Histological Analysis, Immunohistochemistry, and TUNEL Assay
4.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Paus, R.; Foitzik, K. In Search of the “Hair Cycle Clock”: A guided tour. Differentiation 2004, 72, 489–511. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, S.; Ota, Y.; Ozawa, K.; Imamura, T. Dual-mode regulation of hair growth cycle by two Fgf-5 gene products. J. Invest. Dermatol. 2000, 114, 456–463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ota, Y.; Saitoh, Y.; Suzuki, S.; Ozawa, K.; Kawano, M.; Imamura, T. Fibroblast Growth Factor 5 inhibits hair growth by blocking dermal papilla cell activation. Biochem. Biophys. Res. Commun. 2002, 290, 169–176. [Google Scholar] [CrossRef]
- Mizuno, S.; Iijima, S.; Okano, T.; Kajiwara, N.; Kunita, S.; Sugiyama, F.; Yagami, K.-I. Retrotransposon-mediated Fgf5(go-Utr) mutant mice with long pelage hair. Exp. Anim. 2011, 60, 161–167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pennycuik, P.R.; Raphael, K.A. The angora locus (go) in the mouse: Hair morphology, duration of growth cycle and site of action. Genet. Res. 1984, 44, 283–291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hébert, J.M.; Rosenquist, T.; Götz, J.; Martin, G.R. FGF5 as a regulator of the hair growth cycle: Evidence from targeted and spontaneous mutations. Cell 1994, 78, 1017–1025. [Google Scholar] [CrossRef]
- Drögemüller, C.; Rüfenacht, S.; Wichert, B.; Leeb, T. Mutations within the FGF5 gene are associated with hair length in cats. Anim. Genet. 2007, 38, 218–221. [Google Scholar] [CrossRef] [PubMed]
- Dierks, C.; Mömke, S.; Philipp, U.; Distl, O. Allelic heterogeneity of FGF5 mutations causes the long-hair phenotype in dogs. Anim. Genet. 2013, 44, 425–431. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Zhou, S.; Li, C.; Cai, B.; Yu, H.; Ma, B.; Huang, Y.; Ding, Y.; Liu, Y.; Ding, Q.; et al. Base pair editing in goat: Nonsense codon introgression into FGF5 results in longer hair. FEBS J. 2019, 286, 4675–4692. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Li, Y.; Jia, K.; Xu, X.; Li, Y.; Zhao, Y.; Zhang, X.; Zhang, J.; Liu, G.; Deng, S.; et al. Crosstalk between androgen and Wnt/β-Catenin leads to changes of wool density in FGF5-knockout sheep. Cell Death Dis. 2020, 11, 407. [Google Scholar] [CrossRef] [PubMed]
- Legrand, R.; Tiret, L.; Abitbol, M. Two recessive mutations in FGF5 are associated with the long-hair phenotype in donkeys. Genet. Sel. Evol. 2014, 46, 65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daverio, M.S.; Vidal-Rioja, L.; Frank, E.N.; Di Rocco, F. Molecular characterization of the llama FGF5 gene and identification of putative loss of function mutations. Anim. Genet. 2017, 48, 716–719. [Google Scholar] [CrossRef] [PubMed]
- Maraqa, T.; Alhajeri, B.H.; Alhaddad, H. FGF5 missense mutation is associated with dromedary hair length variation. Anim. Genet. 2021, 52, 848–856. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Liu, H.; Pan, H.; Wang, X.; Zhang, Y.; Yao, B.; Li, N.; Lai, L.; Li, Z. CRISPR/Cas9-mediated disruption of Fibroblast Growth Factor 5 in rabbits results in a systemic long hair phenotype by prolonging anagen. Genes 2020, 11, 297. [Google Scholar] [CrossRef] [Green Version]
- Higgins, C.A.; Petukhova, L.; Harel, S.; Ho, Y.Y.; Drill, E.; Shapiro, L.; Wajid, M.; Christiano, A.M. FGF5 is a crucial regulator of hair length in humans. Proc. Natl. Acad. Sci. USA 2014, 111, 10648–10653. [Google Scholar] [CrossRef] [Green Version]
- Yoshizawa, Y.; Wada, K.; Shimoi, G.; Kameyama, Y.; Wakabayashi, Y.; Fukuta, K.; Hashizume, R. A 1-bp deletion in Fgf5 causes male-dominant long hair in the Syrian hamster. Mamm. Genome 2015, 26, 630–637. [Google Scholar] [CrossRef] [PubMed]
- Grymowicz, M.; Rudnicka, E.; Podfigurna, A.; Napierala, P.; Smolarczyk, R.; Smolarczyk, K.; Meczekalski, B. Hormonal effects on hair follicles. Int. J. Mol. Sci. 2020, 21, 5342. [Google Scholar] [CrossRef]
- Randall, V.A. Hormonal regulation of hair follicles exhibits a biological paradox. Semin. Cell Dev. Biol. 2007, 18, 274–285. [Google Scholar] [CrossRef] [PubMed]
- Leirós, G.J.; Ceruti, J.M.; Castellanos, M.L.; Kusinsky, A.G.; Balañá, M.E. Androgens modify Wnt agonists/antagonists expression balance in dermal papilla cells preventing hair follicle stem cell differentiation in androgenetic alopecia. Mol. Cell. Endocrinol. 2017, 439, 26–34. [Google Scholar] [CrossRef]
- Ceruti, J.M.; Leirós, G.J.; Balañá, M.E. Androgens and androgen receptor action in skin and hair follicles. Mol. Cell. Endocrinol. 2018, 465, 122–133. [Google Scholar] [CrossRef] [PubMed]
- Ohtsuka, M.; Sato, M.; Miura, H.; Takabayashi, S.; Matsuyama, M.; Koyano, T.; Arifin, N.; Nakamura, S.; Wada, K.; Gurumurthy, C.B. i-GONAD: A robust method for in situ germline genome engineering using CRISPR nucleases. Genome Biol. 2018, 19, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gurumurthy, C.B.; Takahashi, G.; Wada, K.; Miura, H.; Sato, M.; Ohtsuka, M. GONAD: A novel CRISPR/Cas9 genome editing method that does not require ex vivo handling of embryos. Curr. Protoc. Hum. Genet. 2016, 88, 15.8.1–15.8.12. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, G.; Gurumurthy, C.B.; Wada, K.; Miura, H.; Sato, M.; Ohtsuka, M. GONAD: Genome-Editing via Oviductal Nucleic Acids Delivery system: A novel microinjection independent genome engineering method in mice. Sci. Rep. 2015, 5, 11406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gurumurthy, C.B.; Sato, M.; Nakamura, A.; Inui, M.; Kawano, N.; Islam, M.A.; Ogiwara, S.; Takabayashi, S.; Matsuyama, M.; Nakagawa, S.; et al. Creation of CRISPR-based germline-genome-engineered mice without ex vivo handling of zygotes by i-GONAD. Nat. Protoc. 2019, 14, 2452–2482. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Chao, Y.; Zhou, G.; Chen, Y. Fibroblast Growth Factor 5-short (FGF5s) inhibits the activity of FGF5 in primary and secondary hair follicle dermal papilla cells of cashmere goats. Gene 2016, 575, 393–398. [Google Scholar] [CrossRef]
- Ozawa, K.; Suzuki, S.; Asada, M.; Tomooka, Y.; Li, A.J.; Yoneda, A.; Komi, A.; Imamura, T. An alternatively spliced Fibroblast Growth Factor (FGF)-5 mRNA is abundant in brain and translates into a partial agonist/antagonist for FGF-5 neurotrophic activity. J. Biol. Chem. 1998, 273, 29262–29271. [Google Scholar] [CrossRef] [Green Version]
- Jo, S.K.; Kim, S.; Lee, Y.H. Expression and function of FGF5 isoform in hair growth. Sch. Int. J. Anat. Physiol. 2021, 4, 1–5. [Google Scholar] [CrossRef]
- Lykke-Andersen, S.; Jensen, T.H. Nonsense-mediated mRNA decay: An intricate machinery that shapes transcriptomes. Nat. Rev. Mol. Cell Biol. 2015, 16, 665–677. [Google Scholar] [CrossRef] [Green Version]
- Sundberg, J.P.; Rourk, M.H.; Boggess, D.; Hogan, M.E.; Sundberg, B.A.; Bertolino, A.P. Angora mouse mutation: Altered hair cycle, follicular dystrophy, phenotypic maintenance of skin grafts, and changes in keratin expression. Vet. Pathol. 1997, 34, 171–179. [Google Scholar] [CrossRef]
- Müller-Röver, S.; Handjiski, B.; van der Veen, C.; Eichmüller, S.; Foitzik, K.; McKay, I.A.; Stenn, K.S.; Paus, R. A comprehensive guide for the accurate classification of murine hair follicles in distinct hair cycle stages. J. Invest. Dermatol. 2001, 117, 3–15. [Google Scholar] [CrossRef]
- Seago, S.V.; Ebling, F.J. The hair cycle on the human thigh and upper arm. Br. J. Dermatol. 1985, 113, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Rook, A. Endocrine influences on hair growth. Br. Med. J. 1965, 1, 609–614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elder, M.J. Anatomy and physiology of eyelash follicles: Relevance to lash ablation procedures. Ophthal. Plast. Reconstr. Surg. 1997, 13, 21–25. [Google Scholar] [CrossRef] [PubMed]
- Mendonca, B.B.; Batista, R.L.; Domenice, S.; Costa, E.M.F.; Arnhold, I.J.P.; Russell, D.W.; Wilson, J.D. Steroid 5α-reductase 2 deficiency. J. Steroid Biochem. Mol. Biol. 2016, 163, 206–211. [Google Scholar] [CrossRef]
- Zhao, J.; Harada, N.; Okajima, K. Dihydrotestosterone inhibits hair growth in mice by inhibiting Insulin-like Growth Factor-I production in dermal papillae. Growth Horm. IGF Res. 2011, 21, 260–267. [Google Scholar] [CrossRef]
- Mezick, J.A.; Gendimenico, G.J.; Liebel, F.T.; Stenn, K.S. Androgen-induced delay of hair growth in the golden syrian hamster. Br. J. Dermatol. 1999, 140, 1100–1104. [Google Scholar] [CrossRef]
- Fu, D.; Huang, J.; Li, K.; Chen, Y.; He, Y.; Sun, Y.; Guo, Y.; Du, L.; Qu, Q.; Miao, Y.; et al. Dihydrotestosterone-induced hair regrowth inhibition by activating androgen receptor in C57BL6 mice simulates androgenetic alopecia. Biomed. Pharmacother. 2021, 137, 111247. [Google Scholar] [CrossRef]
- Li, Y.-H.; Zhang, K.; Ye, J.-X.; Lian, X.-H.; Yang, T. Wnt10b promotes growth of hair follicles via a canonical Wnt signalling pathway. Clin. Exp. Dermatol. 2011, 36, 534–540. [Google Scholar] [CrossRef]
- Lin, W.-H.; Xiang, L.-J.; Shi, H.-X.; Zhang, J.; Jiang, L.-P.; Cai, P.-T.; Lin, Z.-L.; Lin, B.-B.; Huang, Y.; Zhang, H.-L.; et al. Fibroblast Growth Factors stimulate hair growth through β-catenin and Shh expression in C57BL/6 mice. Biomed Res. Int. 2015, 2015, 730139. [Google Scholar]
- Papukashvili, D.; Rcheulishvili, N.; Liu, C.; Xie, F.; Tyagi, D.; He, Y.; Wang, P.G. Perspectives on miRNAs targeting DKK1 for developing hair regeneration therapy. Cells 2021, 10, 2957. [Google Scholar] [CrossRef]
- McLean, D.; Cunningham, K.; Ricke, W. MP68-20 establishing a negative feedback loop between Fibroblast Growth Factor 5 and androgen receptor in prostate cancer. J. Urol. 2019, 201, e983. [Google Scholar] [CrossRef]
- Naito, Y.; Hino, K.; Bono, H.; Ui-Tei, K. CRISPRdirect: Software for designing CRISPR/Cas guide RNA with reduced off-Target sites. Bioinformatics 2015, 31, 1120–1123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harms, D.W.; Quadros, R.M.; Seruggia, D.; Ohtsuka, M.; Takahashi, G.; Montoliu, L.; Gurumurthy, C.B. Mouse genome editing using the CRISPR/Cas system. Curr. Protoc. Hum. Genet. 2014, 83, 15.7.1–15.7.27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wada, K.; Matsushima, Y.; Tada, T.; Hasegawa, S.; Obara, Y.; Yoshizawa, Y.; Takahashi, G.; Hiai, H.; Shimanuki, M.; Suzuki, S.; et al. Expression of truncated PITX3 in the developing lens leads to microphthalmia and aphakia in mice. PLoS ONE 2014, 9, e111432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wada, K.; Saito, J.; Yamaguchi, M.; Seki, Y.; Furugori, M.; Takahashi, G.; Nishito, Y.; Matsuda, H.; Shitara, H.; Kikkawa, Y. Pde6brd1 mutation modifies cataractogenesis in Foxe3rct mice. Biochem. Biophys. Res. Commun. 2018, 496, 231–237. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, G.; Hasegawa, S.; Fukutomi, Y.; Harada, C.; Furugori, M.; Seki, Y.; Kikkawa, Y.; Wada, K. A novel missense mutation of Mip causes semi-dominant cataracts in the Nat mouse. Exp. Anim. 2017, 66, 271–282. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Takahashi, R.; Takahashi, G.; Kameyama, Y.; Sato, M.; Ohtsuka, M.; Wada, K. Gender-Difference in Hair Length as Revealed by Crispr-Based Production of Long-Haired Mice with Dysfunctional FGF5 Mutations. Int. J. Mol. Sci. 2022, 23, 11855. https://doi.org/10.3390/ijms231911855
Takahashi R, Takahashi G, Kameyama Y, Sato M, Ohtsuka M, Wada K. Gender-Difference in Hair Length as Revealed by Crispr-Based Production of Long-Haired Mice with Dysfunctional FGF5 Mutations. International Journal of Molecular Sciences. 2022; 23(19):11855. https://doi.org/10.3390/ijms231911855
Chicago/Turabian StyleTakahashi, Ryo, Gou Takahashi, Yuichi Kameyama, Masahiro Sato, Masato Ohtsuka, and Kenta Wada. 2022. "Gender-Difference in Hair Length as Revealed by Crispr-Based Production of Long-Haired Mice with Dysfunctional FGF5 Mutations" International Journal of Molecular Sciences 23, no. 19: 11855. https://doi.org/10.3390/ijms231911855