Characterization and Comparison of Human and Ovine Mesenchymal Stromal Cells from Three Corresponding Sources

 , ,
, ,
Abstract
:1. Introduction
2. Results
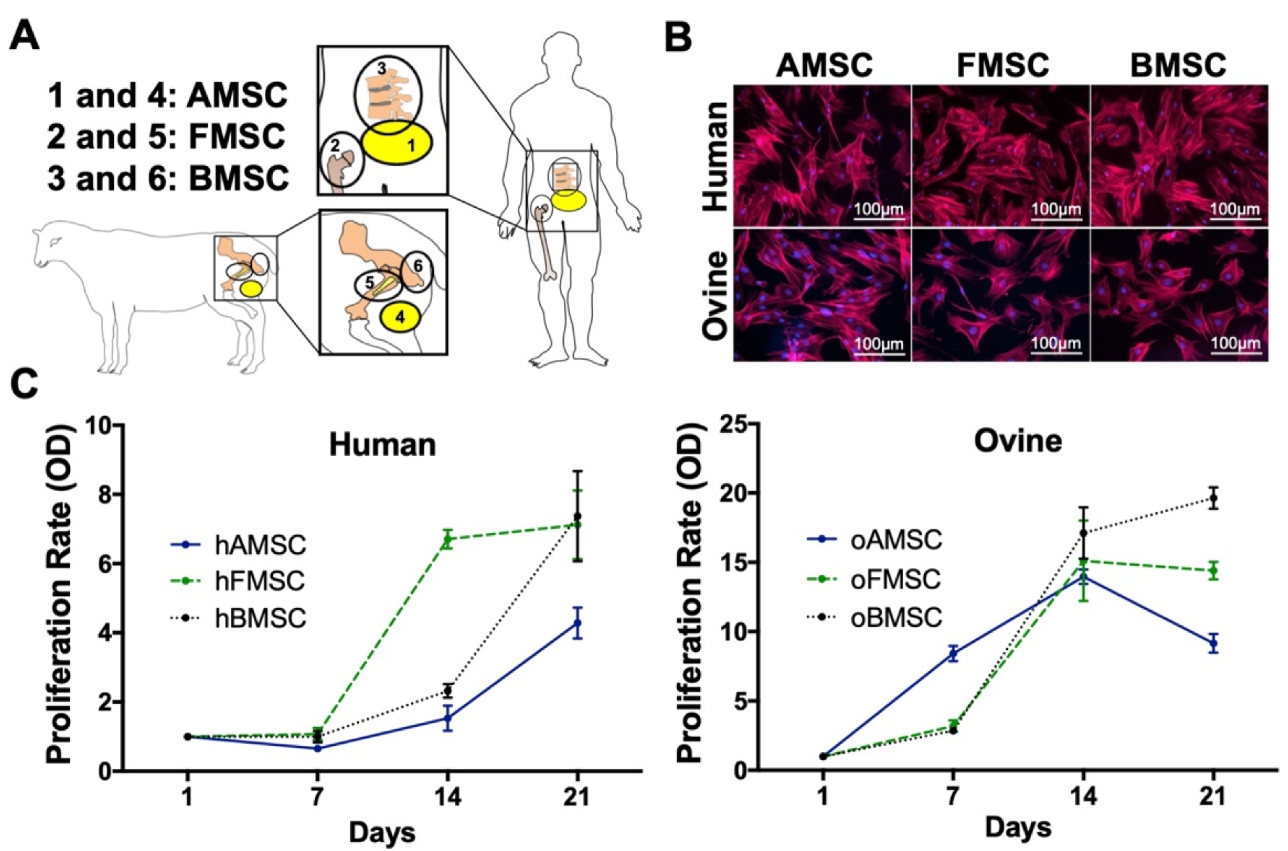
2.1. hMSC and oMSC Morphology and Proliferation Rates
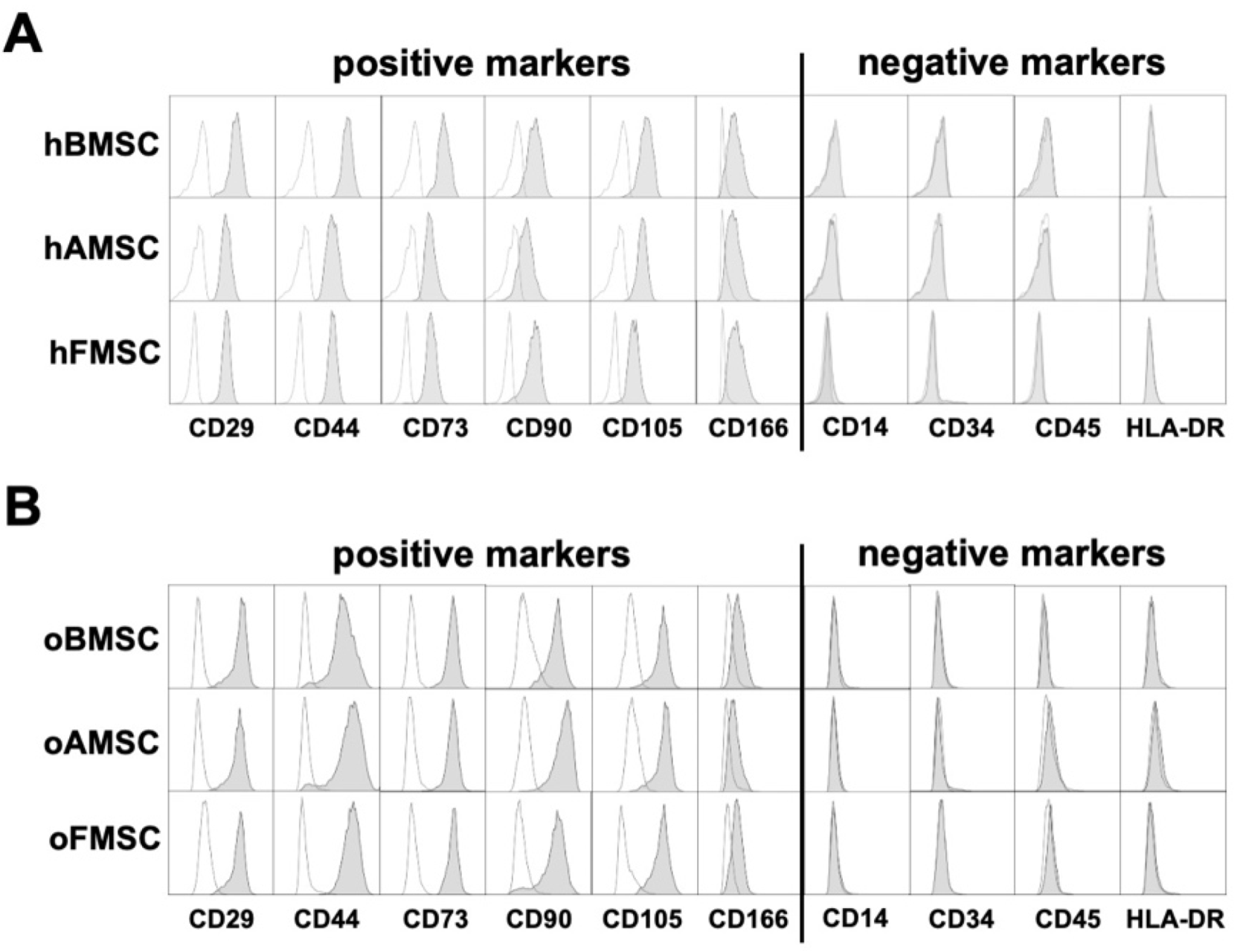
2.2. Determination of Surface Markers
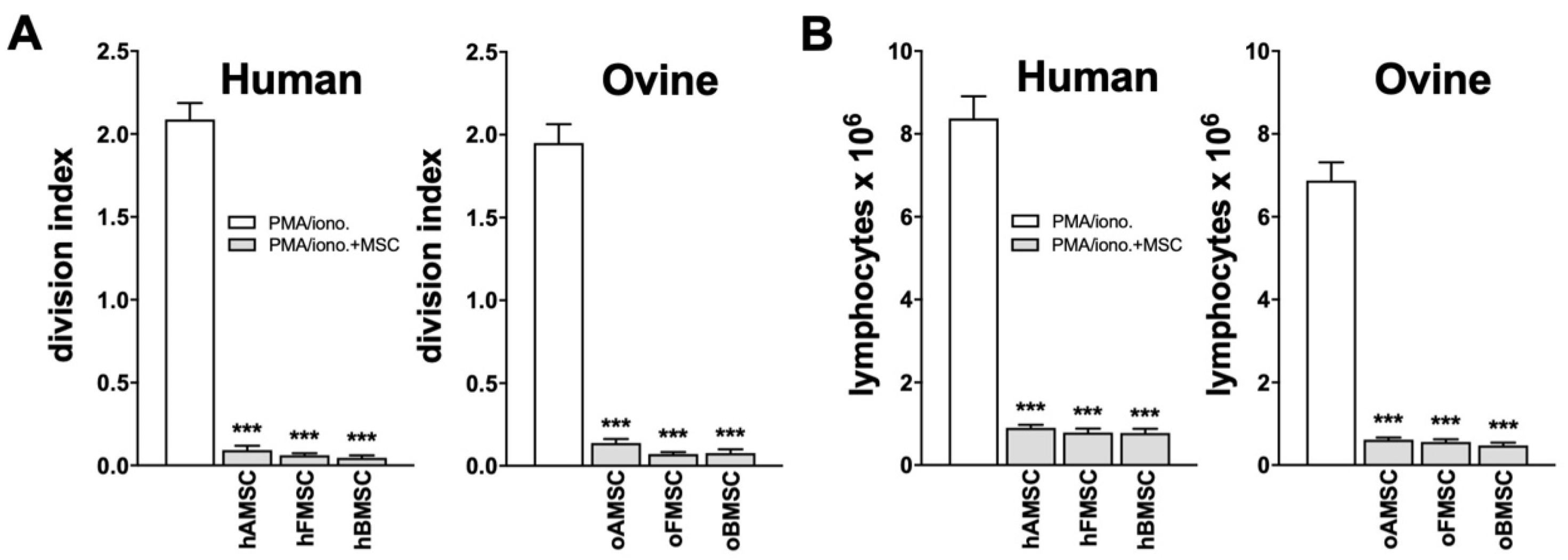
2.3. Immunomodulatory Capacity
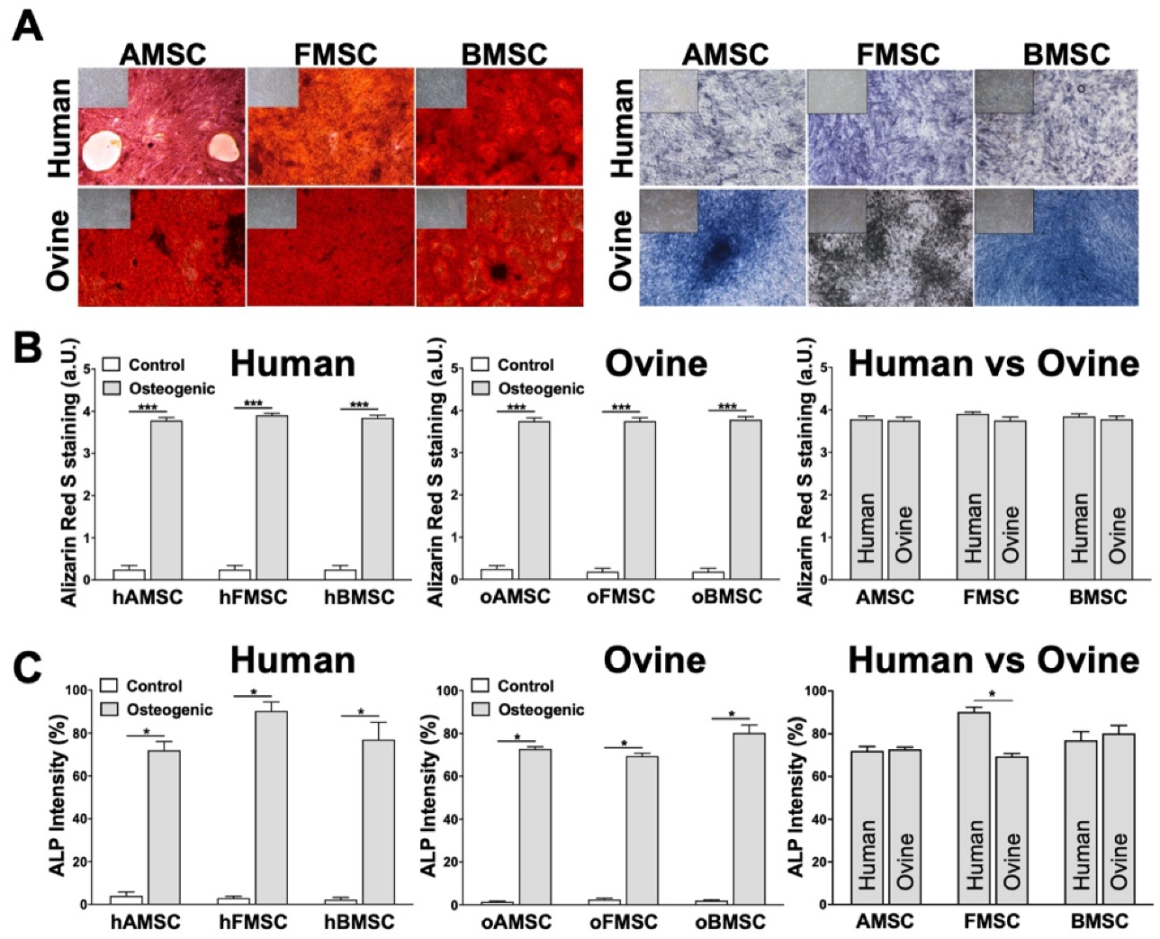
2.4. Differentiation towards Adipogenic and Chondrogenic Lineages
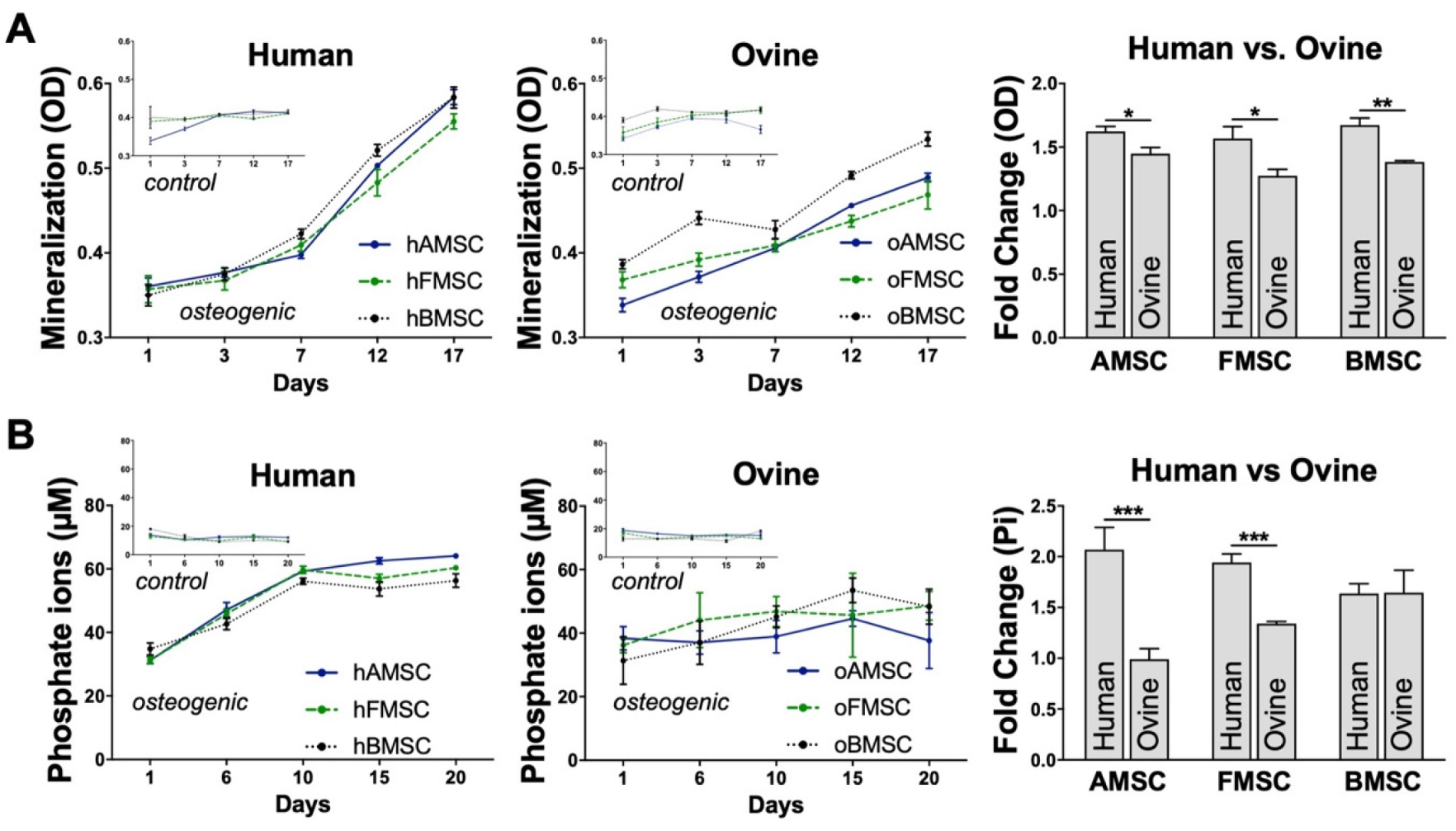
2.5. Assessment of Osteogenic Differentiation
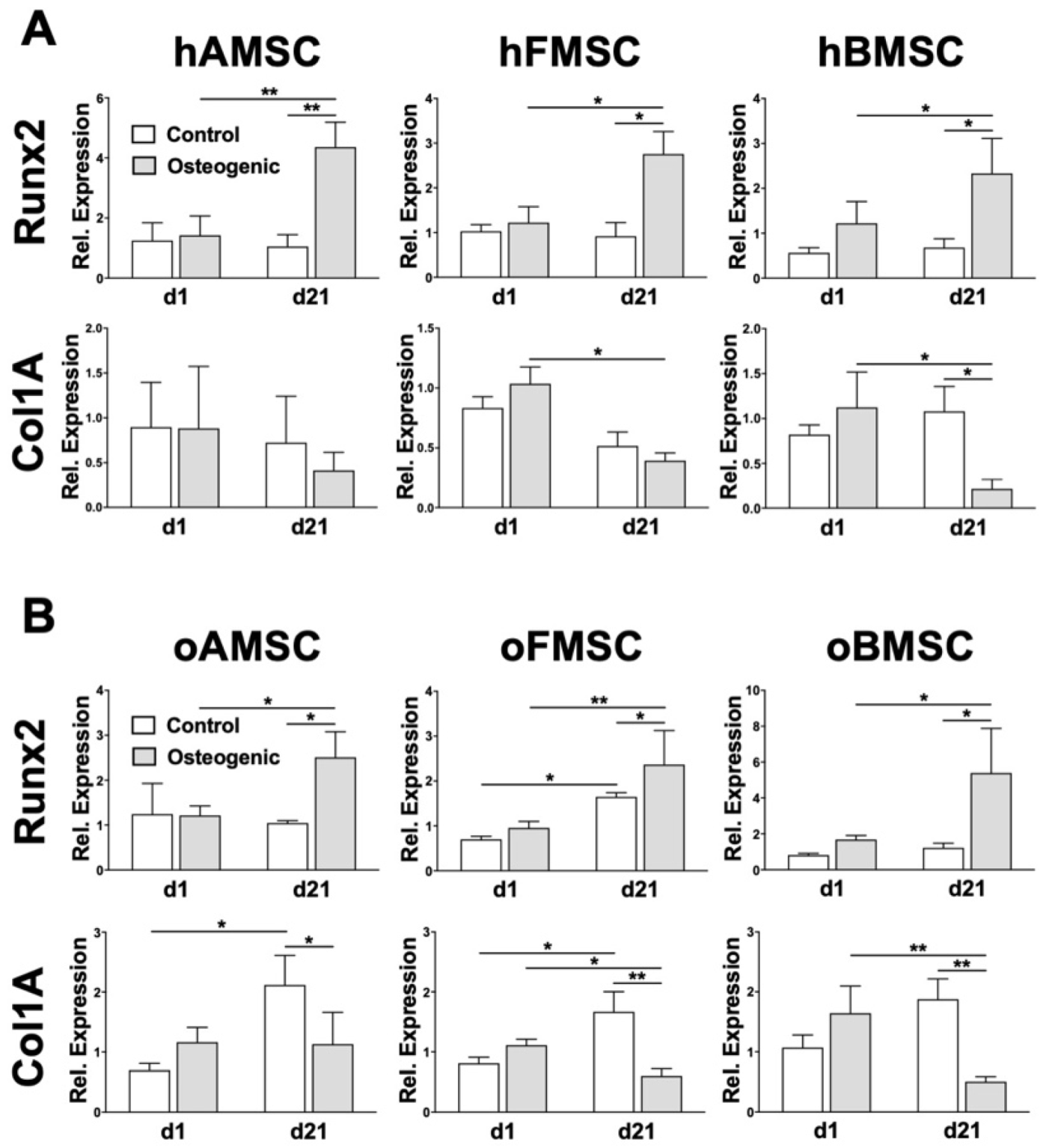
2.6. Osteogenic Lineage-Specific Gene Expression
3. Discussion
4. Materials and Methods
4.1. Tissue Donors and Study Design
4.2. MSC Isolation and Culture
4.3. MSC Morphology
4.4. MSC Proliferation
4.5. Immune Modulation
4.6. MSC Surface Marker Expression
4.7. Adipogenic Differentiation
4.8. Chondrogenic Differentiation
4.9. Osteogenic Differentiation
4.10. Optical Density and Free Phosphate Measurements
4.11. Real-Time Polymerase Chain Reaction (RT-PCR)
4.12. Statistics
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ALP | Alkaline phosphatase |
| AMSC | Adipose tissue-derived mesenchymal stromal cells |
| BMP-2 | Bone morphogenetic protein 2 |
| BMSC | Bone marrow-derived mesenchymal stromal cells |
| CD | Cluster of differentiation |
| cDNA | Complementary deoxyribonucleic acid |
| CFSE | Carboxyfluorescein succinimidyl ester |
| Col1A | Collagen, type I, alpha 1 |
| DAPI | 4′,6-Diamidino-2-phenylindole |
| ddCT | Delta-delta-Ct |
| DMEM | Dulbecco’s modified Eagle’s medium |
| DPBS | Dulbecco’s phosphate-buffered saline |
| EDTA | Ethylenediaminetetraacetic acid |
| FMSC | Femoral-derived mesenchymal stromal cells |
| GAPDH | Glyceraldehyde-3-phosphate dehydrogenase |
| hAMSC | Human adipose tissue-derived mesenchymal stromal cells |
| hBMSC | Human bone marrow-derived mesenchymal stromal cells |
| hFMSC | Human femoral-derived mesenchymal stromal cells |
| HLA-DR | Human leukocyte antigen - DR isotype |
| hMSC | Human mesenchymal stromal cells |
| ISCT | International Society for Cellular Therapy |
| mRNA | Messenger ribonucleic acid |
| MSC | Mesenchymal stromal cells |
| oAMSC | Ovine adipose tissue-derived mesenchymal stromal cells |
| oBMSC | Ovine bone marrow-derived mesenchymal stromal cells |
| OD | Optical density |
| oFMSC | Ovine femoral-derived mesenchymal stromal cells |
| oMSC | Ovine mesenchymal stromal cells |
| PBMC | Peripheral blood mononuclear cell |
| PBS | Phosphate-buffered saline |
| PFA | Paraformaldehyde |
| Pi | Phosphate ions |
| PMA | Phorbol myristate acetate |
| RNA | Ribonucleic acid |
| RT-PCR | Real-time polymerase chain reaction |
| Runx2 | Runt-related transcription factor 2 |
References
- Kon, T.; Cho, T.J.; Aizawa, T.; Yamazaki, M.; Nooh, N.; Graves, D.; Gerstenfeld, L.C.; Einhorn, T.A. Expression of osteoprotegerin, receptor activator of NF-kappaB ligand (Osteoprotegerin ligand) and related proinflammatory cytokines during fracture healing. J. Bone Min. Res. 2001, 16, 1004–1014. [Google Scholar] [CrossRef]
- Einhorn, T.A.; Gerstenfeld, L.C. Fracture healing: Mechanisms and interventions. Nat. Rev. Rheumatol. 2014, 11, 45–54. [Google Scholar] [CrossRef] [Green Version]
- Pape, H.C.; Evans, A.; Kobbe, P. Autologous bone graft: Properties and techniques. J. Orthop. Trauma 2010, 24, S36–S40. [Google Scholar] [CrossRef]
- Giannoudis, P.V.; Einhorn, T.A.; Marsh, D. Fracture healing: The diamond concept. Injury 2007, 38, S3–S6. [Google Scholar] [CrossRef]
- Fayaz, H.C.; Giannoudis, P.V.; Vrahas, M.S.; Smith, R.M.; Moran, C.; Pape, H.C.; Krettek, C.; Jupiter, J.B. The role of stem cells in fracture healing and nonunion. Int. Orthop. 2011, 35, 1587–1597. [Google Scholar] [CrossRef] [Green Version]
- Bernardo, M.E.; Fibbe, W.E. Mesenchymal stromal cells: Sensors and switchers of inflammation. Cell Stem Cell 2013, 13, 392–402. [Google Scholar] [CrossRef] [Green Version]
- Eggenhofer, E.; Luk, F.; Dahlke, M.H.; Hoogduijn, M.J. The life and fate of mesenchymal stem cells. Front. Immunol. 2014, 5, 148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wagers, A.J. The stem cell niche in regenerative medicine. Cell Stem Cell 2012, 10, 362–369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ivanovska, I.L.; Shin, J.W.; Swift, J.; Discher, D.E. Stem cell mechanobiology: Diverse lessons from bone marrow. Trends Cell Biol. 2015, 25, 523–532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, S.; Xie, N.; Li, W.; Yuan, B.; Shi, Y.; Wang, Y. Immunobiology of mesenchymal stem cells. Cell Death Differ. 2014, 21, 216–225. [Google Scholar] [CrossRef]
- Yagi, H.; Soto-Gutierrez, A.; Parekkadan, B.; Kitagawa, Y.; Tompkins, R.G.; Kobayashi, N.; Yarmush, M.L. Mesenchymal stem cells: Mechanisms of immunomodulation and homing. Cell Transpl. 2010, 19, 667–679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pittenger, M.F.; Mackay, A.M.; Beck, S.C.; Jaiswal, R.K.; Douglas, R.; Mosca, J.D.; Moorman, M.A.; Simonetti, D.W.; Craig, S.; Marshak, D.R. Multilineage potential of adult human mesenchymal stem cells. Science 1999, 284, 143–147. [Google Scholar] [CrossRef] [Green Version]
- Horwitz, E.M.; Le Blanc, K.; Dominici, M.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.C.; Deans, R.J.; Krause, D.S.; Keating, A.; International Society for Cellular Therapy. Clarification of the nomenclature for MSC: The International Society for Cellular Therapy position statement. Cytotherapy 2005, 7, 393–395. [Google Scholar] [CrossRef] [PubMed]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.; Krause, D.; Deans, R.; Keating, A.; Prockop, D.; Horwitz, E. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef]
- Pearce, A.I.; Richards, R.G.; Milz, S.; Schneider, E.; Pearce, S.G. Animal models for implant biomaterial research in bone: A review. Eur. Cell Mater. 2007, 13, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, M.B.; Guimaraes, J.A.; Casado, P.L.; Cavalcanti Ados, S.; Goncalves, N.N.; Ambrosio, C.E.; Rodrigues, F.; Pinto, A.C.; Miglino, M.A.; Duarte, M.E. The effect of bone allografts combined with bone marrow stromal cells on the healing of segmental bone defects in a sheep model. BMC Vet. Res. 2014, 10, 36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Viateau, V.; Guillemin, G.; Bousson, V.; Oudina, K.; Hannouche, D.; Sedel, L.; Logeart-Avramoglou, D.; Petite, H. Long-bone critical-size defects treated with tissue-engineered grafts: A study on sheep. J. Orthop. Res. 2007, 25, 741–749. [Google Scholar] [CrossRef] [PubMed]
- Martini, L.; Fini, M.; Giavaresi, G.; Giardino, R. Sheep model in orthopedic research: A literature review. CompMed 2001, 51, 292–299. [Google Scholar]
- O’Loughlin, P.F.; Morr, S.; Bogunovic, L.; Kim, A.D.; Park, B.; Lane, J.M. Selection and development of preclinical models in fracture-healing research. J. Bone Jt. Surg. Am. 2008, 90, 79–84. [Google Scholar] [CrossRef]
- Kolar, P.; Schmidt-Bleek, K.; Schell, H.; Gaber, T.; Toben, D.; Schmidmaier, G.; Perka, C.; Buttgereit, F.; Duda, G.N. The early fracture hematoma and its potential role in fracture healing. Tissue Eng. Part B Rev. 2010, 16, 427–434. [Google Scholar] [CrossRef]
- Bottagisio, M.; Lovati, A.B.; Lopa, S.; Moretti, M. Osteogenic Differentiation of Human and Ovine Bone Marrow Stromal Cells in response to beta-Glycerophosphate and Monosodium Phosphate. Cell. Reprogram. 2015, 17, 235–242. [Google Scholar] [CrossRef] [PubMed]
- Loebel, C.; Czekanska, E.M.; Bruderer, M.; Salzmann, G.; Alini, M.; Stoddart, M.J. In vitro osteogenic potential of human mesenchymal stem cells is predicted by Runx2/Sox9 ratio. Tissue Eng. Part A 2015, 21, 115–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaneto, C.M.; Lima, P.S.; Zanette, D.L.; Prata, K.L.; Pina Neto, J.M.; de Paula, F.J.; Silva, W.A., Jr. COL1A1 and miR-29b show lower expression levels during osteoblast differentiation of bone marrow stromal cells from Osteogenesis Imperfecta patients. BMC Med. Genet. 2014, 15, 45. [Google Scholar] [CrossRef] [Green Version]
- Okolicsanyi, R.K.; Camilleri, E.T.; Oikari, L.E.; Yu, C.; Cool, S.M.; van Wijnen, A.J.; Griffiths, L.R.; Haupt, L.M. Human Mesenchymal Stem Cells Retain Multilineage Differentiation Capacity Including Neural Marker Expression after Extended In Vitro Expansion. PLoS ONE 2015, 10, e0137255. [Google Scholar] [CrossRef]
- Schildberg, F.A.; Donnenberg, V.S. Stromal cells in health and disease. Cytoma A 2018, 93, 871–875. [Google Scholar] [CrossRef]
- Ghesquiere, B.; Wong, B.W.; Kuchnio, A.; Carmeliet, P. Metabolism of stromal and immune cells in health and disease. Nature 2014, 511, 167–176. [Google Scholar] [CrossRef] [Green Version]
- Galipeau, J.; Krampera, M.; Barrett, J.; Dazzi, F.; Deans, R.J.; DeBruijn, J.; Dominici, M.; Fibbe, W.E.; Gee, A.P.; Gimble, J.M.; et al. International Society for Cellular Therapy perspective on immune functional assays for mesenchymal stromal cells as potency release criterion for advanced phase clinical trials. Cytotherapy 2016, 18, 151–159. [Google Scholar] [CrossRef] [Green Version]
- Pasumarthy, K.K.; Doni Jayavelu, N.; Kilpinen, L.; Andrus, C.; Battle, S.L.; Korhonen, M.; Lehenkari, P.; Lund, R.; Laitinen, S.; Hawkins, R.D. Methylome Analysis of Human Bone Marrow MSCs Reveals Extensive Age- and Culture-Induced Changes at Distal Regulatory Elements. Stem Cell Rep. 2017, 9, 999–1015. [Google Scholar] [CrossRef] [Green Version]
- Lam, A.T.; Li, J.; Toh, J.P.; Sim, E.J.; Chen, A.K.; Chan, J.K.; Choolani, M.; Reuveny, S.; Birch, W.R.; Oh, S.K. Biodegradable poly-epsilon-caprolactone microcarriers for efficient production of human mesenchymal stromal cells and secreted cytokines in batch and fed-batch bioreactors. Cytotherapy 2017, 19, 419–432. [Google Scholar] [CrossRef] [PubMed]
- Ercal, P.; Pekozer, G.G.; Kose, G.T. Dental Stem Cells in Bone Tissue Engineering: Current Overview and Challenges. Adv. Exp. Med. Biol. 2018, 1107, 113–127. [Google Scholar] [PubMed]
- Heino, T.J.; Hentunen, T.A. Differentiation of osteoblasts and osteocytes from mesenchymal stem cells. Curr. Stem Cell Res. 2008, 3, 131–145. [Google Scholar] [CrossRef] [PubMed]
- Faiella, W.; Atoui, R. Immunotolerant Properties of Mesenchymal Stem Cells: Updated Review. Stem Cells Int. 2016, 2016, 1859567. [Google Scholar] [CrossRef] [Green Version]
- Berebichez-Fridman, R.; Gomez-Garcia, R.; Granados-Montiel, J.; Berebichez-Fastlicht, E.; Olivos-Meza, A.; Granados, J.; Velasquillo, C.; Ibarra, C. The Holy Grail of Orthopedic Surgery: Mesenchymal Stem Cells-Their Current Uses and Potential Applications. Stem Cells Int. 2017, 2017, 2638305. [Google Scholar] [CrossRef] [PubMed]
- Giannoni, P.; Mastrogiacomo, M.; Alini, M.; Pearce, S.G.; Corsi, A.; Santolini, F.; Muraglia, A.; Bianco, P.; Cancedda, R. Regeneration of large bone defects in sheep using bone marrow stromal cells. J. Tissue Eng. Regen. Med. 2008, 2, 253–262. [Google Scholar] [CrossRef] [PubMed]
- McCarty, R.C.; Gronthos, S.; Zannettino, A.C.; Foster, B.K.; Xian, C.J. Characterisation and developmental potential of ovine bone marrow derived mesenchymal stem cells. J. Cell. Physiol. 2009, 219, 324–333. [Google Scholar] [CrossRef]
- Menicanin, D.; Mrozik, K.M.; Wada, N.; Marino, V.; Shi, S.; Bartold, P.M.; Gronthos, S. Periodontal-ligament-derived stem cells exhibit the capacity for long-term survival, self-renewal, and regeneration of multiple tissue types in vivo. Stem Cells Dev. 2014, 23, 1001–1011. [Google Scholar] [CrossRef]
- Koobatian, M.T.; Liang, M.S.; Swartz, D.D.; Andreadis, S.T. Differential effects of culture senescence and mechanical stimulation on the proliferation and leiomyogenic differentiation of MSC from different sources: Implications for engineering vascular grafts. Tissue Eng. Part A 2015, 21, 1364–1375. [Google Scholar] [CrossRef] [Green Version]
- Lovati, A.B.; Lopa, S.; Recordati, C.; Talo, G.; Turrisi, C.; Bottagisio, M.; Losa, M.; Scanziani, E.; Moretti, M. In Vivo Bone Formation Within Engineered Hydroxyapatite Scaffolds in a Sheep Model. Calcif. Tissue Int. 2016, 99, 209–223. [Google Scholar] [CrossRef]
- Adamzyk, C.; Emonds, T.; Falkenstein, J.; Tolba, R.; Jahnen-Dechent, W.; Lethaus, B.; Neuss, S. Different Culture Media Affect Proliferation, Surface Epitope Expression, and Differentiation of Ovine MSC. Stem Cells Int. 2013, 2013, 387324. [Google Scholar] [CrossRef] [Green Version]
- Kalaszczynska, I.; Ruminski, S.; Platek, A.E.; Bissenik, I.; Zakrzewski, P.; Noszczyk, M.; Lewandowska-Szumiel, M. Substantial differences between human and ovine mesenchymal stem cells in response to osteogenic media: How to explain and how to manage? Biores. Open Access 2013, 2, 356–363. [Google Scholar] [CrossRef]
- Music, E.; Futrega, K.; Doran, M.R. Sheep as a model for evaluating mesenchymal stem/stromal cell (MSC)-based chondral defect repair. Osteoarthr. Cartil. 2018, 26, 730–740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rentsch, C.; Hess, R.; Rentsch, B.; Hofmann, A.; Manthey, S.; Scharnweber, D.; Biewener, A.; Zwipp, H. Ovine bone marrow mesenchymal stem cells: Isolation and characterization of the cells and their osteogenic differentiation potential on embroidered and surface-modified polycaprolactone-co-lactide scaffolds. In Vitro Cell. Dev. Biol. Anim. 2010, 46, 624–634. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.R.; Chandrashekran, A.; Smith, R.K.; Dudhia, J. Immunophenotypic characterization of ovine mesenchymal stem cells. Cytoma A 2016, 89, 443–450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vivas, D.; Caminal, M.; Oliver-Vila, I.; Vives, J. Derivation of Multipotent Mesenchymal Stromal Cells from Ovine Bone Marrow. Curr. Protoc. Stem Cell Biol. 2018, 44. [Google Scholar] [CrossRef] [PubMed]
- Gao, F.; Chiu, S.M.; Motan, D.A.; Zhang, Z.; Chen, L.; Ji, H.L.; Tse, H.F.; Fu, Q.L.; Lian, Q. Mesenchymal stem cells and immunomodulation: Current status and future prospects. Cell Death Dis. 2016, 7, e2062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mrugala, D.; Bony, C.; Neves, N.; Caillot, L.; Fabre, S.; Moukoko, D.; Jorgensen, C.; Noel, D. Phenotypic and functional characterisation of ovine mesenchymal stem cells: Application to a cartilage defect model. Ann. Rheum. Dis. 2008, 67, 288–295. [Google Scholar] [CrossRef]
- Sanjurjo-Rodriguez, C.; Castro-Vinuelas, R.; Hermida-Gomez, T.; Fernandez-Vazquez, T.; Fuentes-Boquete, I.M.; de Toro-Santos, F.J.; Diaz-Prado, S.M.; Blanco-Garcia, F.J. Ovine Mesenchymal Stromal Cells: Morphologic, Phenotypic and Functional Characterization for Osteochondral Tissue Engineering. PLoS ONE 2017, 12, e0171231. [Google Scholar] [CrossRef] [Green Version]
- Loebel, C.; Czekanska, E.M.; Staudacher, J.; Salzmann, G.; Richards, R.G.; Alini, M.; Stoddart, M.J. The calcification potential of human MSCs can be enhanced by interleukin-1beta in osteogenic medium. J. Tissue Eng. Regen. Med. 2017, 11, 564–571. [Google Scholar] [CrossRef]
- Suzuki, A.; Ghayor, C.; Guicheux, J.; Magne, D.; Quillard, S.; Kakita, A.; Ono, Y.; Miura, Y.; Oiso, Y.; Itoh, M.; et al. Enhanced expression of the inorganic phosphate transporter Pit-1 is involved in BMP-2-induced matrix mineralization in osteoblast-like cells. J. Bone Min. Res. 2006, 21, 674–683. [Google Scholar] [CrossRef]
- Jikko, A.; Harris, S.E.; Chen, D.; Mendrick, D.L.; Damsky, C.H. Collagen integrin receptors regulate early osteoblast differentiation induced by BMP-2. J. Bone Min. Res. 1999, 14, 1075–1083. [Google Scholar] [CrossRef]
- Kulterer, B.; Friedl, G.; Jandrositz, A.; Sanchez-Cabo, F.; Prokesch, A.; Paar, C.; Scheideler, M.; Windhager, R.; Preisegger, K.H.; Trajanoski, Z. Gene expression profiling of human mesenchymal stem cells derived from bone marrow during expansion and osteoblast differentiation. BMC Genom. 2007, 8, 70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schildberg, F.A.; Wojtalla, A.; Siegmund, S.V.; Endl, E.; Diehl, L.; Abdullah, Z.; Kurts, C.; Knolle, P.A. Murine hepatic stellate cells veto CD8 T cell activation by a CD54-dependent mechanism. Hepatology 2011, 54, 262–272. [Google Scholar] [CrossRef] [PubMed]
- Schildberg, F.A.; Hegenbarth, S.I.; Schumak, B.; Scholz, K.; Limmer, A.; Knolle, P.A. Liver sinusoidal endothelial cells veto CD8 T cell activation by antigen-presenting dendritic cells. Eur. J. Immunol. 2008, 38, 957–967. [Google Scholar] [CrossRef] [PubMed]
- Langenbach, F.; Berr, K.; Naujoks, C.; Hassel, A.; Hentschel, M.; Depprich, R.; Kubler, N.R.; Meyer, U.; Wiesmann, H.P.; Kogler, G.; et al. Generation and differentiation of microtissues from multipotent precursor cells for use in tissue engineering. Nat. Protoc. 2011, 6, 1726–1735. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Human | Ovine |
|---|---|---|
| GAPDH | fwd: 5′CTCTGCTCCTCCTGTTCGAC3′ rev: 5′ACCAAATCCGTTGACTCCGA3′ | fwd: 5′TCACCATCTTCCAGGAGCGA3′ rev: 5′GGTGCAGAGATGATGACCCT3′ |
| Col1A | fwd: 5′TGCTCGTGGAAATGATGGTG3′ rev: 5′CCTCGCTTTCCTTCCTCTCC3′ | fwd: 5′CATGACCGAGACGTGTGGAA3′ rev: 5′CATTCGTCCGTGGGGACTTT3′ |
| Runx2 | fwd: 5′GCGCATTCCTCATCCCAGTA3′ rev: 5′GGCTCAGGTAGGAGGGGTAA3′ | fwd: 5′ CCGCCGGACTCGAACTG3′ rev: 5′GAGAGGCGCAGGTCTTGATG3′ |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Haddouti, E.-M.; Randau, T.M.; Hilgers, C.; Masson, W.; Walgenbach, K.J.; Pflugmacher, R.; Burger, C.; Gravius, S.; Schildberg, F.A. Characterization and Comparison of Human and Ovine Mesenchymal Stromal Cells from Three Corresponding Sources. Int. J. Mol. Sci. 2020, 21, 2310. https://doi.org/10.3390/ijms21072310
Haddouti E-M, Randau TM, Hilgers C, Masson W, Walgenbach KJ, Pflugmacher R, Burger C, Gravius S, Schildberg FA. Characterization and Comparison of Human and Ovine Mesenchymal Stromal Cells from Three Corresponding Sources. International Journal of Molecular Sciences. 2020; 21(7):2310. https://doi.org/10.3390/ijms21072310
Chicago/Turabian StyleHaddouti, El-Mustapha, Thomas M. Randau, Cäcilia Hilgers, Werner Masson, Klaus J. Walgenbach, Robert Pflugmacher, Christof Burger, Sascha Gravius, and Frank A. Schildberg. 2020. "Characterization and Comparison of Human and Ovine Mesenchymal Stromal Cells from Three Corresponding Sources" International Journal of Molecular Sciences 21, no. 7: 2310. https://doi.org/10.3390/ijms21072310