 Cristina Andrés-Barrao1
Cristina Andrés-Barrao1 Feras F. Lafi1,2†
Feras F. Lafi1,2† Intikhab Alam2
Intikhab Alam2 Axel de Zélicourt1
Axel de Zélicourt1 Abdul A. Eida1
Abdul A. Eida1 Ameerah Bokhari1
Ameerah Bokhari1 Hanin Alzubaidy1
Hanin Alzubaidy1 Vladimir B. Bajic2
Vladimir B. Bajic2 Heribert Hirt1*
Heribert Hirt1* Maged M. Saad1
Maged M. Saad1- 1Biological and Environmental Sciences and Engineering Division, King Abdullah University of Science and Technology, Thuwal, Saudi Arabia
- 2Computational Bioscience Research Center, King Abdullah University of Science and Technology, Thuwal, Saudi Arabia
Enterobacter sp. SA187 is an endophytic bacterium that has been isolated from root nodules of the indigenous desert plant Indigofera argentea. SA187 could survive in the rhizosphere as well as in association with different plant species, and was able to provide abiotic stress tolerance to Arabidopsis thaliana. The genome sequence of SA187 was obtained by using Pacific BioScience (PacBio) single-molecule sequencing technology, with average coverage of 275X. The genome of SA187 consists of one single 4,429,597 bp chromosome, with an average 56% GC content and 4,347 predicted protein coding DNA sequences (CDS), 153 ncRNA, 7 rRNA, and 84 tRNA. Functional analysis of the SA187 genome revealed a large number of genes involved in uptake and exchange of nutrients, chemotaxis, mobilization and plant colonization. A high number of genes were also found to be involved in survival, defense against oxidative stress and production of antimicrobial compounds and toxins. Moreover, different metabolic pathways were identified that potentially contribute to plant growth promotion. The information encoded in the genome of SA187 reveals the characteristics of a dualistic lifestyle of a bacterium that can adapt to different environments and promote the growth of plants. This information provides a better understanding of the mechanisms involved in plant-microbe interaction and could be further exploited to develop SA187 as a biological agent to improve agricultural practices in marginal and arid lands.
Introduction
Worldwide agriculture is currently facing big challenges posed by the increase in global population and climate change, and plant growth-promoting bacteria (PGPB) are becoming an important alternative for sustainable crop production (De Zélicourt et al., 2013; Timmusk et al., 2017). The increase in global temperature is drastically affecting the amount of available arable lands, particularly in dryland areas (40% of world land surface), where approximately half of the poorest people live (rural areas in developing countries) (El-Beltagy and Madkour, 2012). Due to climate change, dryland areas (hyper-arid, arid, and semi-arid lands) are expected to rapidly increase, at the same time as crops, cropping systems, rotations and biota will undergo a deep transformation (El-Beltagy and Madkour, 2012).
Plant growth-promoting bacteria are a group of bacteria that are taxonomically unrelated and can establish symbiotic associations with plants to promote their growth under harsh environmental conditions. PGPB can live in the rhizosphere, epiphytically attached to the surface of roots or leaves, or as endophytic bacteria, living inside the plant tissues. PGPB affect plant growth by directly acquiring nutrients (phosphate, nitrogen, iron) or modulating plant hormone levels (auxins, ethylene), and also by indirectly inhibiting pathogenic bacteria (antibiotics) or insects (pesticides) (Glick, 2012).
In order to identify a beneficial strain that might help crops to cope with various environmental challenges, researchers have started to look for halophilic and halotolerant bacteria inhabiting salty and arid ecosystems, which have the potential to promote plant growth under salinity and drought conditions (Jorquera et al., 2012; Mapelli et al., 2013; Liu et al., 2016). In the last decade, the desert has become a niche of growing research interest for bacteria that are adapted to water scarcity, high salinity and high temperature, and could be used to promote growth of crops under these conditions (Goswami et al., 2014; Vurukonda et al., 2016). Desert soils from across the world typically contain a number of ubiquitous bacterial phyla including Actinobacteria, Bacteroidetes, and Proteobacteria (Makhalanyane et al., 2015), and Halomonas, Salicola, Bacillus, Paenibacillus, Klebsiella, Enterobacter, Serratia, Cellulosimicrobium, Ochrobactrum, and Pseudomonas have been identified among the most abundant genera in saline and hypersaline rhizospheric soils (Bhatnagar and Bhatnagar, 2005; Hedi et al., 2009; Köberl et al., 2011; Marasco et al., 2012; Hanna et al., 2013; Köberl et al., 2013; Mapelli et al., 2013; Goswami et al., 2014). Microbes found in desert soils have been subjected to evolutionary adaptation to extreme conditions and show higher abundance of gene functions related to dormancy and stress response than microbes in non-arid environments (Fierer et al., 2012). Moreover, these microbes have been found to enhance soil fertility and promote the growth of plants (Herman et al., 1995; Marasco et al., 2012; Köberl et al., 2013; Goswami et al., 2014).
Following this idea, our group launched the Darwin21 project1, with the goal of exploring the microbial diversity of desert pioneer plants and their use in improving agricultural sustainability in dryland and marginal areas. Preliminary results revealed a large diversity of bacteria that have the potential to promote the growth of different crop plants under diverse biotic and abiotic stresses. In the frame of this project, our group also obtained the draft genomes of several PGPB (Lafi et al., 2016a,b,c, 2017a,c).
Among the isolates, one particular endophytic bacterium, SA187, showed the highest consistency in terms of growth promotion among all performed laboratory tests. SA187 was isolated from root nodules of Indigofera argentea, an indigenous desert plant growing in the Kingdom of Saudi Arabia. Based on the analysis of the almost complete 16S rRNA gene (>1400 bp), SA187 was classified as member of the family Enterobacteriaceae and named as Enterobacter sp. SA187 (Lafi et al., 2017b). The family Enterobacteriaceae contains a large number of genera that are found in different environmental niches and are biochemically and genetically closely related. Strains belonging to the genus Enterobacter have been isolated from the Atacama Desert, northern Chile, showing tolerance to arsenic (Azua-Bustos and González-Silva, 2014), in the desert soil in Algeria showing capability to degrade glyphosate (Benslama and Boulahrouf, 2013, 2016), or in the rhizosphere of desert plants from oil-polluted soils (Diab et al., 2017). Moreover, members of this genus have been found as endophytic plant growth promoters in date palms (Phoenix dactylifera L.) under saline conditions or in association with other diverse plants growing in arid lands (Park et al., 2005; Shankar et al., 2011; Yaish et al., 2015). Members of the genus Enterobacter have also been reported to show PGP traits, e.g., Enterobacter sp. EnB1, isolated from semidesert soil in Mexico, which has been reported to be able to solubilize phosphate (Delgado et al., 2014), or Enterobacter sp. B6, isolated from desert soil in Algeria, which showed biopesticide properties against Locusta migratoria L5 nymphs (Oulebsir-Mohandkaci et al., 2015).
Over the past few years, a number of Enterobacter sp. and close relatives in the family Enterobacteriaceae showing PGP under abiotic stress have been also characterized. For example, Enterobacter cloacae SBP-8 (formerly Klebsiella sp. SBP-8), which induced systemic tolerance in wheat under salt stress (Singh et al., 2015, 2017), E. cloacae UW5, which was able to produce high-levels of indole-3-acetic acid (IAA) (Coulson and Patten, 2015), or E. oryzae Ola 51T, which was capable to fix atmospheric nitrogen (Peng et al., 2009). Additionally, members of the genera Enterobacter, Klebsiella, and Leclercia have also been reported as phosphate-solubilizers, nitrogen-fixers, and producers of antifungal compounds, phytohormones and siderophores (Melo et al., 2016).
SA187 promoted the growth of the model plant Arabidopsis thaliana under diverse abiotic stresses such as salinity, drought or high temperature, demonstrating an important potential for application as PGPB to improve abiotic resistance and yield of crops in arid lands. Here we present the analysis of the complete genome sequence and the biochemical characterization of SA187, and highlight different metabolic pathways that potentially contribute to plant growth and stress tolerance.
Materials and Methods
Genomic DNA Isolation and PCR Amplification
Genomic DNA was extracted from fresh bacterial cultures by using Qiagen’s DNeasy blood and tissue kit (Qiagen GmbH, Hilden, Germany) and treated with RNAse A (MO BIO Laboratories, Carlsbad, CA, United States). The extracted DNA was further purified by using Mo Bio PowerClean Pro DNA Clean-Up kit (MO BIO Laboratories, Carlsbad, CA, United States) following the manufacturer’s instructions. DNA quality and quantity were assessed by using Nanodrop 2000 (Thermo Fisher Scientific, Wilmington, DE, United States) and Qubit 2.0 (Life Technologies, Invitrogen division, Darmstadt, Germany). The purified DNA was shipped to the sequencing facilities by using the DNAstable® Plus (Biomatrica, San Diego, CA, United States).
(GTG)5-rep-PCR amplification was carried out in a total volume of 25 μl as described by Andrés-Barrao et al. (2016) by using the GTG5 primer (Rademaker et al., 1998) (Supplementary Table S1). The amplification of the 16S rRNA gene was carried out in a total volume of 20 μl by using the universal primers 27F and 1429R (Supplementary Table S1): 200 ng DNA template was added to 18 μl of PCR mixture [2X Taq Polymerase Master Mix (Promega), 0.75 μM each primer] by using the following program: A initial denaturation of 95°C for 5 min followed by 35 cycles with steps of 94°C for 30 s, 55°C for 45 s and 72°C for 1 min 30 s, and a final extension of 5 min at 72°C. PCR amplification of the housekeeping genes was carried out in a total volume of 50 μl as follows: 100 ng DNA template was added to 49 μl of PCR mixture [Taq DNA polymerase (Invitrogen), 1.5 μM MgCl2, 125 μM dNTPs, 0.2 μM each primer, 1.5 μl DMSO] by using the following program: A initial denaturation of 95°C for 3 min followed by 35 cycles with steps of 95°C for 30 s, 57°C for 45 s and 72°C for 1 min 30 s, and a final extension of 5 min at 72°C. All PCR reactions were carried out in a C1000 Touch Thermal Cycler (Bio-Rad, United States), run in 0.8-1.5% agarose gel electrophoresis and visualized in a ChemiDocTM MP Imaging System (Bio-Rad, United States).
Whole Genome Sequencing and Annotation
Prior to sequencing, DNA was size selected to 20 kb by using the BluePippin system (Sage Science, Beverly, MA, United States). Whole genome sequencing was performed by DNA Link Korea (Seoul, South Korea) using the PacBio RS II sequencing platform (Pacific Biosciences, Menlo Park, CA, United States). Large-insert libraries were sequenced in single-molecule real-time (SMRT) sequencing cells by using P6-C4 chemistry. PacBio reads were assembled into one single contig by using the de novo Hierarchical Genome Assembly Process (HGAP.2) algorithm with default parameters. Genome annotation was conducted by using the in-house Automatic Annotation of Microbial Genomes (AAMG) pipeline (Alam et al., 2013). Briefly, AAMG annotated the genome by first predicting RNAs and open reading frames (ORFs), followed by BLAST and Interproscan. Ribosomal RNAs (rRNAs), transfer RNAs (tRNAs) and other non-coding RNAs (ncRNAs) were predicted by using RNAmmer, tRNAscan-SE and Infernal software, respectively (Alam et al., 2013). ORFs were predicted by using FragGene Scan (Rho et al., 2010) with the training model specific to complete genomes. ORFs were annotated based on BLAST against UniprotKB and Kyoto Encyclopedia of Genes and Genomes (KEGG) databases. Protein core signatures, domains and associated Gene Ontology (GO) were assigned using InterProscan. Circular chromosome and GC skew were computed by using CGViewer Server (Grant and Stothard, 2008) and GenSkew2, respectively. Chromosomes of related bacteria were aligned by progressive MAUVE (Darling et al., 2010). Function and pathway analysis was performed by using BlastKOALA web tool of KEGG database (Kanehisa et al., 2016). Function analysis by Cluster of Orthologous Genes (COG) was done by using WebMGA (Wu et al., 2011). In-house blast searches were performed through INDIGO-Desert v1.1 (Alam et al., 2013). Toxin-antitoxin (T/A) systems were retrieved by using TA finder (Shao et al., 2011). Gene clusters for the biosynthesis of secondary metabolites were identified by using antiSMASH v.4.0.1 (Weber et al., 2015).
Phylogenetic Analysis
Phylogenetic analyses based on 16S rRNA gene and multi-locus sequence analysis (MLSA) were used to evaluate the taxonomical affiliation of SA187. To construct the 16S and MLSA phylogenetic trees, sequences of individual genes of SA187, along with those from reference strains of the genus Enterobacter and other closely related genera retrieved from public databases3, were aligned by using ClustalW (Thompson et al., 2002). For MLSA analysis, blunt end alignments of partial sequences were obtained for each gene: 615 bp (infB), 742 bp (gyrB), 642 bp (atpD), 637 bp (rpoB). The concatenated gyrB-rpoB-atpD-infB sequences were used to construct the Neighbor-Joining phylogenetic tree. Phylogenetic and molecular evolutionary analyses were conducted by using MEGA 6 (Tamura et al., 2013). Accession numbers of the sequences used in this study are shown in Supplementary Table S2. Similarity matrices showing the pairwise percentage identity between 16S rRNA gene sequences and housekeeping genes concatenated sequences, as well as the correspondent distance matrices are shown in Supplementary Tables S3 and S4.
For further whole-genome phylogenetic analysis, we obtained related genome sets initially based on megablast and later additional members of selected genera, so that a resolved species tree could be constructed. We gathered 55 genomes of 12 different genera. Once at least 3–5 genomes were obtained for each genera, we predicted genes using FragGeneScan to carry out clustering of protein coding genes using OrthoMCL. This provided us with a list of clusters common and unique to the analyzed genomes. We separated the core set of gene clusters (1250 genes) and aligned each of these using MAFFT aligner. Alignments were then concatenated and FastTree was used with default parameters to obtain a species tree with bootstrap values. Two-way ANIs calculation was performed on http://enve-omics.ce.gatech.edu/ani/index, considering a minimum length of 700 bp and minimum identity of 70%. The fragment options were set to 1000 bp for window size and 200 bp for step size. Accession numbers of the genome sequences used in this study, together with % similarity and ANI values are shown in Supplementary Table S5 and Figure S4.
Data Deposition
The whole-genome shotgun project was deposited in NCBI/DDBJ/EMBL database under the accession no CP019113.
Arabidopsis Salt Stress Resistance Screening
Arabidopsis thaliana seeds were surface sterilized by 10 min shaking in 70% EtOH + 0.05% sodium dodecyl sulfate (SDS), then washed 2–3 times in milliQ water and let to dry. Sterilized seeds where stratified for 2 days on half-strength Murashige-Skoog Basal Salt broth pH 5.8 (½MS) (Murashige and Skoog, 1962) (Sigma, Germany) agar plates, at 4°C and darkness, then transferred vertically to a Percival (22°C, 16 h light cycle) for germination. Five-day old seedlings were then transferred to fresh ½MS+100 mM NaCl agar plates (6 seedlings/plate) next to a LB agar plug with (+B, treated) or without (-B, control) SA187. Plant growth was followed for 12 additional days, until the plant roots reached the bottom of the plates. To evaluate the effect of the bacterial treatment in the plant growth, aerial fresh weight (AFW), root fresh weight (RFW), total fresh weight (FW), and lateral root density (LRD) were determined at the end of the experiment.
Quantitative PCR (RT-qPCR)
To evaluate the gene expression of several key genes identified during SA187 genome analysis, the total RNA of SA187 treated roots (dual samples) were extracted by using the NucleoSpin® RNA Plant kit (Macherey-Nagel), following manufacturer’s instructions. SA187 grown overnight in LB liquid was used as a control. Total RNA of control SA187 was extracted by using the RiboPureTM RNA Purification Kit, bacteria (Ambion), following the manufacturer’s instructions. A total of 1 mg of total RNA from dual and control samples was retrotranscribed by using SuperScript® III First-Strand Synthesis System for RT-PCR (Invitrogen), using random hexamers and following manufacturer’s instructions. Two μl of the obtained cDNA solution diluted 10-fold was mixed with specific primers (Supplementary Table S1) and SsoAdvancedTM Universal SYBR® Green Supermix (Bio-Rad), and the qPCR reactions were performed in a CFX96 TouchTM Real-Time PCR Detection System (Bio-Rad) as follows: 95°C for 3 min, 40x [95°C for 10 s and 60°C for 40 s], 95°C for 10 s and a final melting curve [65–95°C, 0.5°C increment, for 5 s]. All reactions were performed in three biological replicates. Gene expression values were calculated relative to the housekeeping gene infB, by using the ΔΔCt method.
Biochemical Assays
Evaluation of Plant Growth Promoting Traits and Tolerance to Abiotic Stresses
Plant growth promoting (PGP) traits were evaluated by using clearing assays. The ability of SA187 to solubilize phosphate was assessed on Pikovskaya’s (PVK) agar plates (M520, Himedia). The production of siderophores was determined by Blue Agar CAS assay, as described by Louden et al. (2011). Zinc solubilization was assessed on modified PVK agar plates, supplemented with 0.1% ZnCO3, as described by Bapiri et al. (2013). Assays were performed by inoculating 30 μl of overnight LB bacterial culture into cavities of ∼0.5 cm in diameter. The production of indole-3-acetic acid was qualitatively determined according to Bric et al. (1991), with the following modifications: LB broth supplemented with 2.5 mM L-tryptophan were used in 96-well plates, which were incubated at 28°C and 190 rpm for 2 days. Tolerance to drought and salt stresses were evaluated by growing SA187 in LB broth (Lennox L Broth Base, Invitrogen) supplemented with 20% polyethylene-glycol (PEG) 6000 and 3% or 5% NaCl, respectively. Tolerance to heat stress was assessed by using LB agar plates incubated at 37°C or 42°C. Assays were performed in triplicates and plates and liquid cultures were incubated at 28°C for 2–5 days.
Evaluation of Bacterial Growth in Different Carbon Sources and Salt Concentrations
SA187 was grown overnight in LB broth (Lennox L Broth Base, Invitrogen) at 28°C with 190 rpm until the culture reached the exponential growth phase. Cells were harvested by centrifugation, washed twice with 10 mM MgSO4, and finally resuspended in 10 mM MgSO4 to a final OD600 of 0.5. Thirty microliters of this cell suspension were inoculated by triplicate in 96-well plates, in 300 μl of LB supplemented with increasing concentrations of NaCl (0–4 M) and ½MS alone or supplemented with 1% of the following carbon sources: arabinose, fructose, glucose, glycerol, lactose, maltose, raffinose, sucrose, acetic acid, citric acid, or lactic acid. Bacterial growth was monitored by using a Varioskan Flash microplate reader (Thermo Scientific), where the 96-well plates were incubated for 2 days at 28°C and 300 rpm.
Evaluation of Antibiotic Resistance
Antibiotic sensitivity tests were evaluated by clearing assay. Hundred μl of overnight SA187 culture were spread on LB agar plates, where antibiotic impregnated disks were then placed (Fischer Scientific). Clearing rings around the disks indicated inhibition of bacterial growth, hence antibiotic sensitivity (S). When SA187 was able to grow normally and no clearing ring was observed, the bacterium contained the corresponding antibiotic resistance marker (R).
Results and Discussion
Genome Sequence Assembly and General Features
Previously, our group published a SA187 draft genome, using Illumina MiSeq technology and obtaining a final assembly of 13 scaffolds (Lafi et al., 2017b). In the present work, PacBio reads were de novo assembled by using the Hierarchical Genome Assembly Process (HGAP.2) software and the PBcR pipeline, resulting in 4,793 filtered and preassembled sequence reads with a mean length of 13,471 bp and 275X genome coverage. Consensus polished assembly yielded one circular contig (Figure 1). Accordingly, the genome of SA187 consists of a single circular chromosome of 4,429,597 bp, with an average 56% GC content and no plasmids (Table 1). A clear GC skew transition was observed, and the origin of replication (oriC) and terminus (terC) were identified at the positions 1,071,819 and 3,184,452, respectively (Figure 1). A total of 4,606 ORFs were identified, including 4,347 protein coding DNA genes (CDS), 153 ncRNAs, 7 complete rRNAs, one additional 5S rRNA, and 84 tRNAs. This number of rRNAs and tRNAs is typical of soil bacteria and an indication of positive selection (Trujillo et al., 2014). Indeed, a high number of rRNAs is a typical characteristic of soil microorganisms, which are able to respond rapidly to changing availability of nutrients (Klappenbach et al., 2000; Shrestha et al., 2007; Lee et al., 2008). Among the CDS, a total of 3,779 (82%) were annotated as genes with a biological function, while 568 (12.3%) were annotated as hypothetical proteins or proteins with unknown function (Table 1). 27 ncRNAs were identified as clustered regularly interspaced short palindromic repeat (CRISPR) RNA direct repeat elements, part of the CRISPR/Cas system, which provides acquired resistance against bacteriophages. This result suggests that the genome of SA187 may have been shaped by interaction with bacteriophages (Horvath and Barrangou, 2010; Al-Attar et al., 2011). AntiSMASH analysis revealed 18 clusters for the biosynthesis of secondary metabolites (Supplementary Table S6). Among them, biosynthetic clusters were identified for lipopolysaccharides, emulsan, O- and K-antigens, colonic acid, carotenoids, streptomycin, asukamycin and turnerbactin.
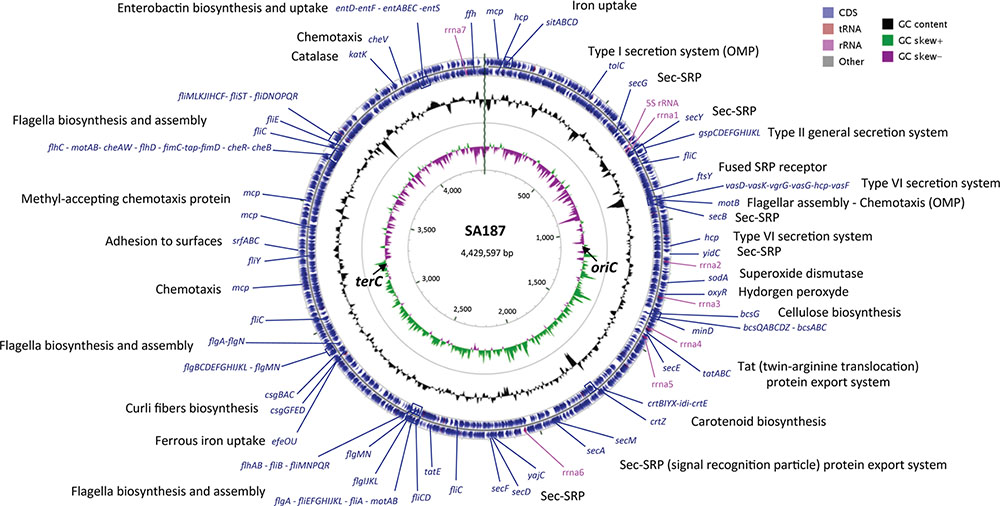
FIGURE 1. Genome map of the SA187. The bacterial chromosome is 4.4 Mb in size. The outer concentric circles (blue) include the annotation, location and direction of expression of predicted genes, the middle circle (black) indicates the % GC content, and the inner circle indicates de GC skew [(G–C)/(G+C)] positive (green) and negative (purple). A number of interesting genes are highlighted.
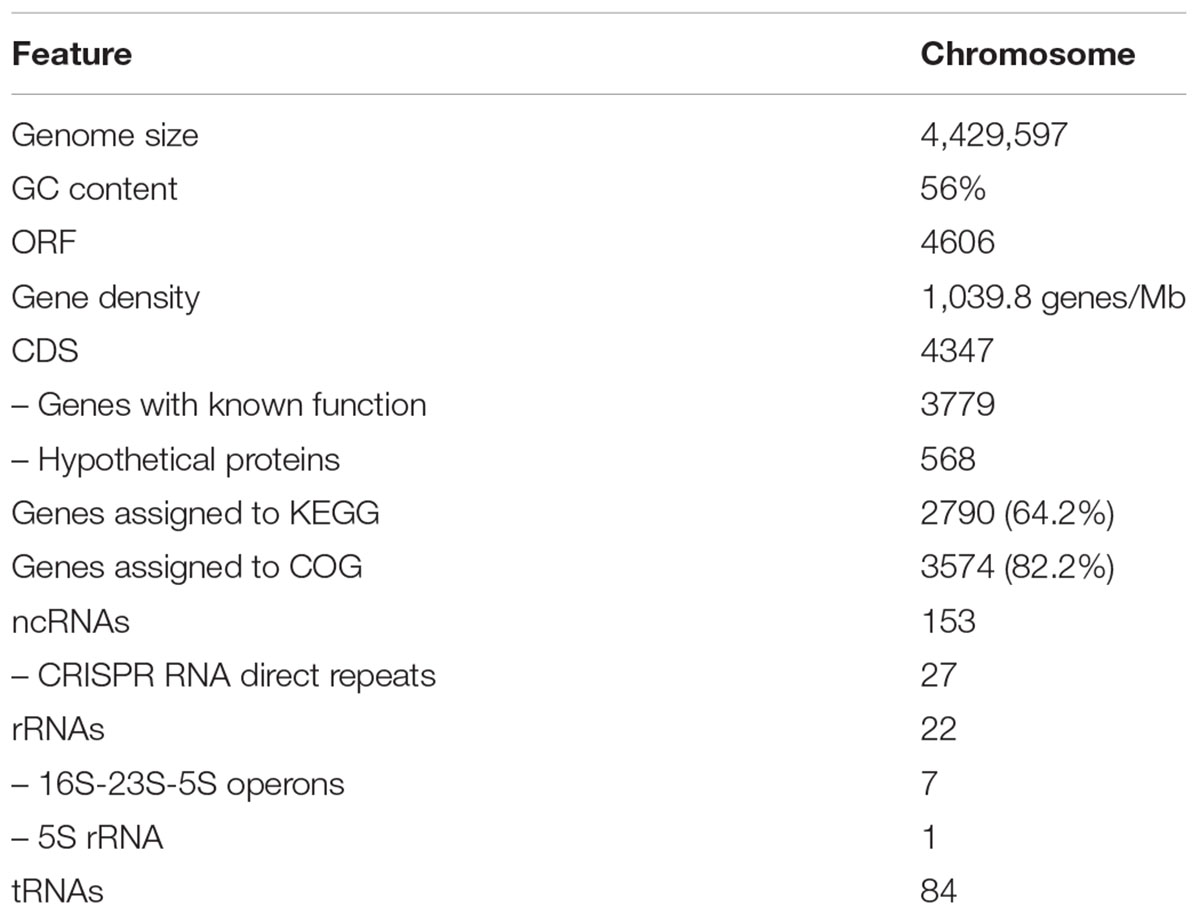
TABLE 1. SA187 genome structure and general features.
We performed functional analysis by Cluster of Orthologous Groups (COGs) (Table 2A and Supplementary Figure S1A). 3574 out of the 4347 predicted CDSs (82.2%) were assigned to a COG category. This result revealed three main functional gene classes: Carbohydrate transport and metabolism (G), amino acid transport and metabolism (E) and transcription (K), representing 26.85% of the predicted CDS. Another high percentage cluster (20.54%) represented genes involved in energy production and conversion (C), cell wall/membrane/envelope biogenesis (M), inorganic ions transport and metabolism (P) and signal transduction (T). Almost 17% of the predicted CDS are poorly characterized: general function prediction only (R) and with function unknown (S). The high proportion of genes involved in transport and metabolism of carbohydrates, amino acids and inorganic ions, indicate the inherent capacity of SA187 to compete with other microorganisms and survive in the rhizosphere (Niazi et al., 2014). Essential genes from the COG functional categories G and K have been found to have a lower rate of evolution compared with the corresponding non-essential genes (Luo et al., 2015).
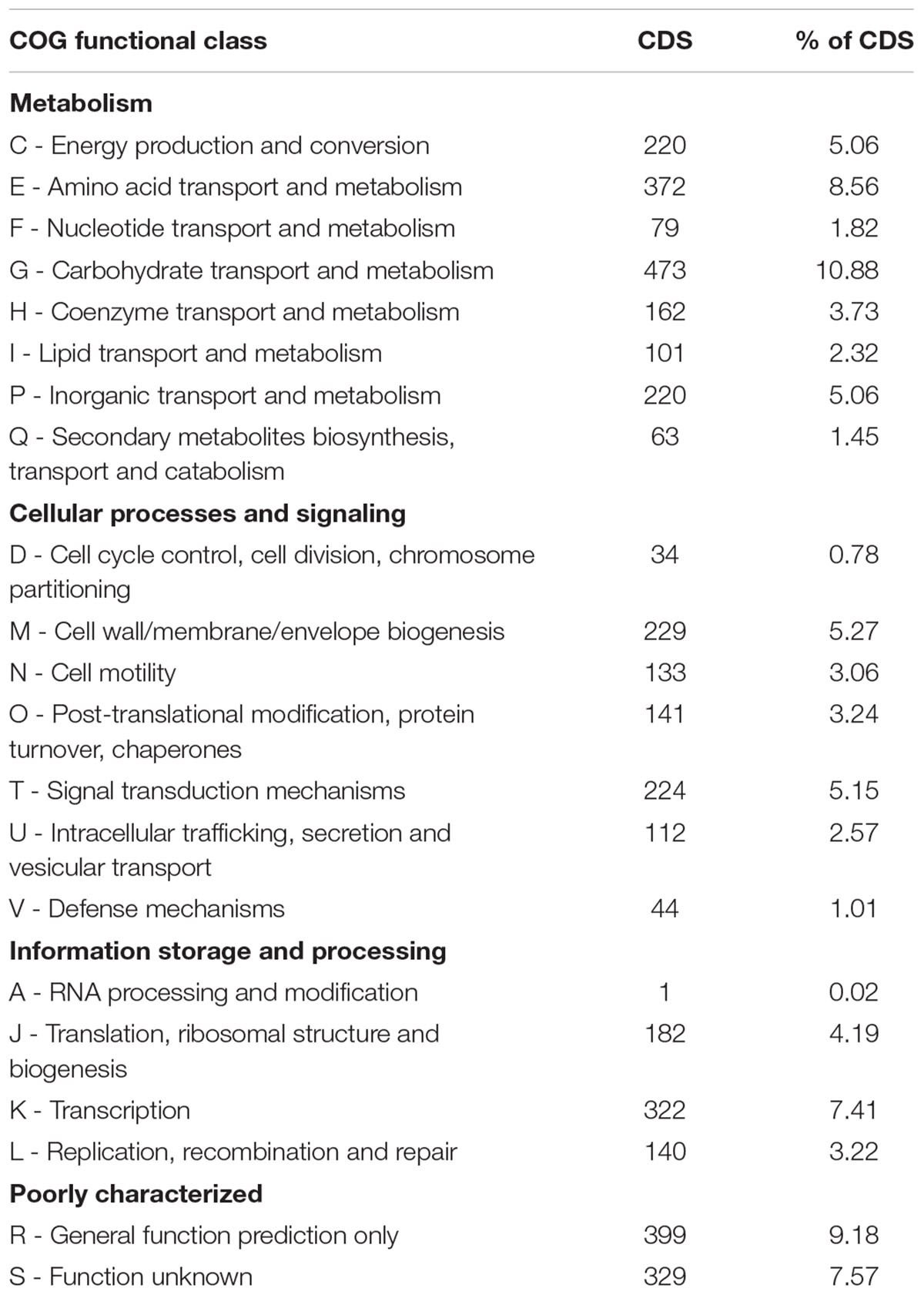
TABLE 2A. Functional cluster of orthologous genes (COG) classification of predicted genes in SA187.
The functional analysis performed by using the KEGG identified 2790 genes (64.2% of all CDSs) involved in any of the metabolic pathways included in the knowledgebase (Table 2B and Supplementary Figure S1B). The analysis revealed the largest number of identified genes as unclassified (10.42%). From those genes that were classified among the KEGG pathway categories, the largest number was involved in metabolism of carbohydrates (7.84%), amino acids (4.23%) and cofactors and vitamins (3.69%). Most of the remaining genes were involved in processes related to environmental and information processing: Membrane transport [ABC transporters 4.88%, phosphotransferase systems (PTS) 2.32%, secretion systems 0.85%] and signal transduction (two-component systems 4.32%). These results confirmed a preference toward metabolism and transport of carbohydrates and amino acids, as well as signal transduction, consistent with the previous results from COG functional analysis.
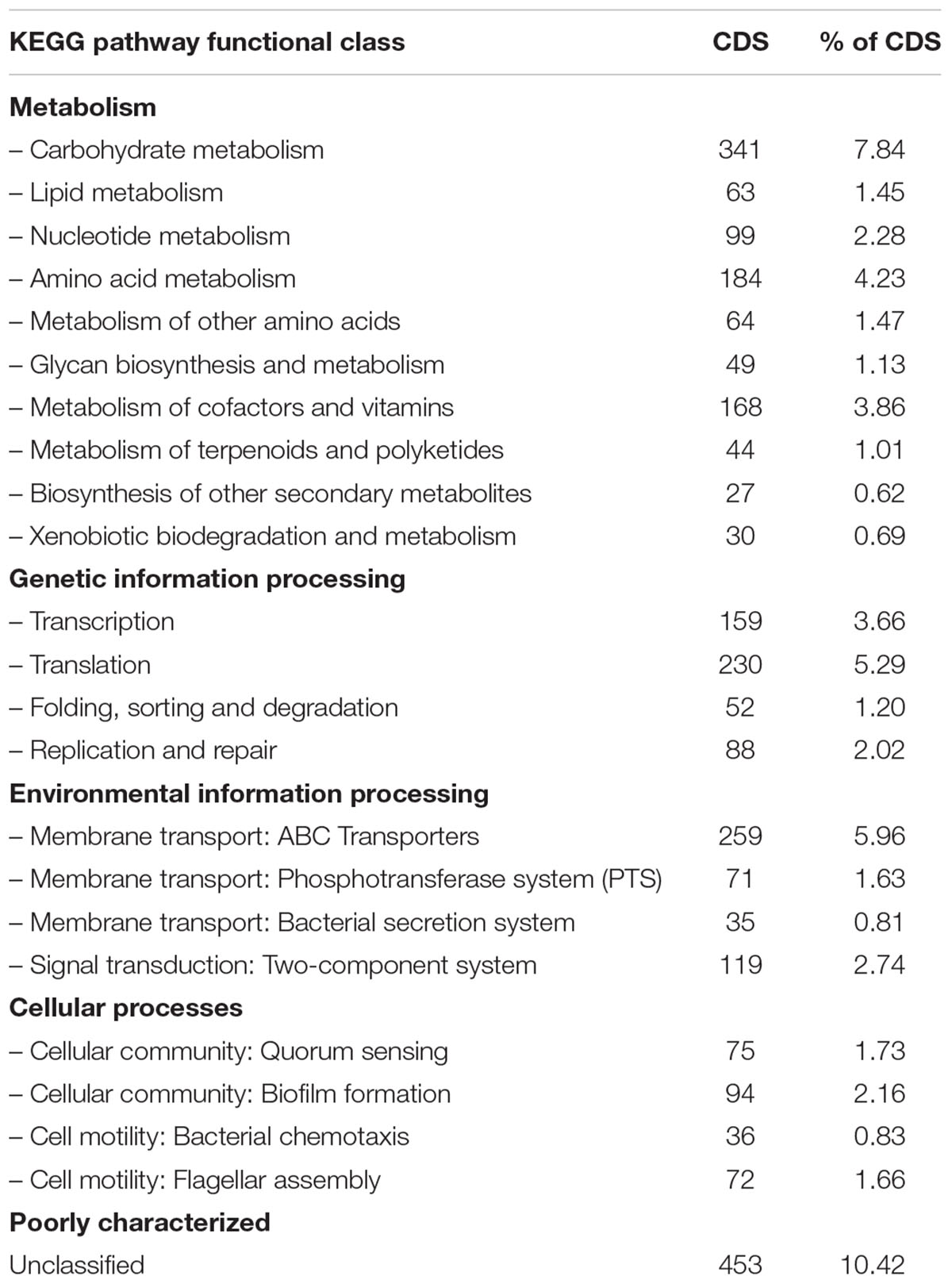
TABLE 2B. Functional Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway classification of predicted genes in SA187.
Taxonomic Affiliation
SA187 was first identified as Enterobacter sp., closely related to E. kobei, based on the full 16S rRNA gene (Lafi et al., 2017b) (Figure 2A and Supplementary Tables S3A, S4A). However, based on the 16S rRNA gene alone, the polyphyletic nature of the genus Enterobacter makes its classification very difficult. Brady et al. (2008) developed the use of Multilocus Sequence Analysis (MLSA) based on four housekeeping genes (gyrB-rpoB-atpD-infB) to obtain a more accurate classification within the family Enterobacteriaceae. By applying MLSA, SA187 appeared in a separate clade, distant from the group Enterobacter (Figure 2B). The taxonomic position of SA187 appeared in a cluster of strains related to the genus Lelliottia, with the closest relative Leclercia adecarboxylata LMG 2803T (Figure 2B). Nevertheless, the low similarity (92%) and the high evolutionary distance (82%) (Supplementary Tables S3B, S4B and Figure S2) between the concatenated sequences of SA187 and LMG 2803T, together with the low robustness of the clade (bootstrap 53%) (Figure 2B), do not allow the classification of the two strains as members of the same genus. These results suggest that SA187 represents a novel taxon within the family Enterobacteriaceae.
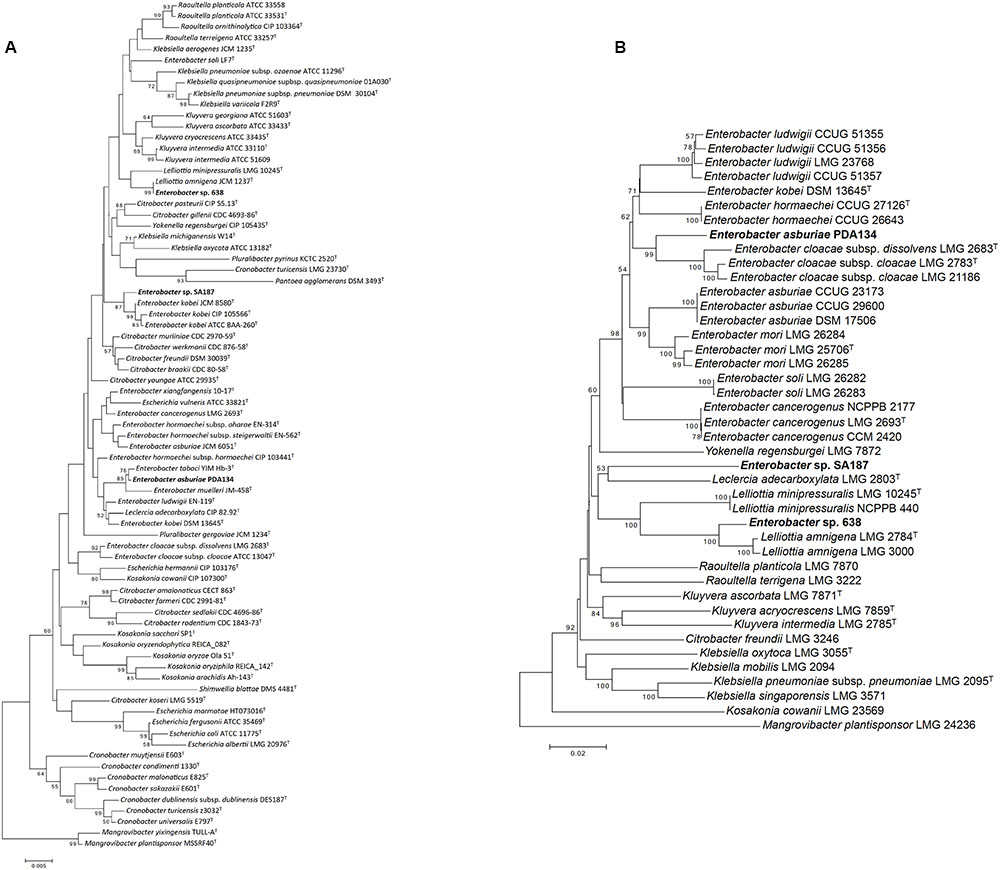
FIGURE 2. Taxonomic analysis. (A) 16S rRNA based phylogenetic tree. (B) Multilocus sequence analysis (MLSA) based on four housekeeping genes gyrB-rpoB-atpD-infB. The phylogenetic tree was inferred by using the Neighbor-Joining method (Saitou and Nei, 1987), and the optimal tree is shown. The percentage of replicate trees in which the associated taxa clustered together in the bootstrap test (1000 replicates) is shown next to the branches (bootstrap >50 is shown) (Felsenstein, 1985). The tree is drawn to scale, with branch lengths in the same units as those of the evolutionary distances used to infer the phylogenetic tree. The evolutionary distances (number of nucleotide substitutions per site) were computed using the Kimura-2-parameter method (Kimura, 1980). All ambiguous positions were removed for each sequence pair. There was a total of 2,636 positions in the final dataset.
Similarly, strains 638 and PDA 134, which were used as reference for comparative genomic analyses, were initially identified based on partial 16S rRNA as Enterobacter sp. and Klebsiella oxytoca, respectively (Taghavi et al., 2009; Gu et al., 2014), and strain PDA 134 was further reclassified as E. asburiae (Yaish, 2016). Unlike this, the position of these strains in the MLSA phylogenetic tree (Figure 2B) reveals Enterobacter sp. 638 as a new species in the genus Lelliottia, and PDA 134 in a different cluster from that formed by the E. asburiae group. The evolutionary distance between E. asburiae PDA 134 and its closer relatives suggests the strain could represent a new species within the genus Enterobacter (Figure 2B).
Further whole-genome analysis confirmed these results (Supplementary Figure S3). In this case, SA187 seemed to be closely related to the recently proposed genus Kosakonia (Brady et al., 2013), although cannot yet be taxonomically affiliated to any of the closest known species, as ANI values among all the sequences included in the phylogenetic tree ranged 78–81%. Consistently with the MLSA results, Enterobacter sp. 638 (Supplementary Table S5 and Figure S4) appeared to belong the genus Lelliottia, and E. asburiae PDA134 was closely related to the E. cloacae group (Supplementary Figure S3).
Arabidopsis Plant Growth Promotion under Salt Stress
The treatment of A. thaliana under salt stress conditions (½MS+100 mM) with SA187 showed a clear PGPB effect. After 12 days growth, seedlings treated with SA187 showed bigger shoots and more developed root systems (Figures 3A,B) and an increment of 50% in all measured parameters (Figure 3C).

FIGURE 3. Plant growth promotion under salt stress. (A) Arabidopsis thaliana growing in ½ MS+100 mM NaCl, with no bacterial treatment (–B, control). (B) A. thaliana growing in ½ MS+100 mM NaCl, treated with SA187 bacterialized plug (+B). (C) Radar chart representing the effect of SA187 treatment in the growth of Arabidopsis. AFW, aerial fresh weight; RFW, root fresh weight; FW, total fresh weight; LRD, lateral root density.
Gene expression of several key genes later identified in this report, gave hints to elucidate a possible mechanism for plant growth promotion: the gene coding for iron(III) ABC transporter substrate-binding protein, afuA; phytoene synthase, crtB; MFS transporter ENTS family enterobactin (siderophore) exporter, entS; and PTS system, sucrose-specific IIB component, srcA, were highly expressed when SA187 was associated with roots, compared with the pure bacterial culture, suggesting their role in plant growth promotion (Figure 4A). The gene coding for catalase, katE, did not show significant differences (Figure 4A), and 2 genes coding for the protein flagellin, fliC, a structural part of the bacterial flagellum, decreased its expression upon association with Arabidopsis roots (Figure 4B). The decrease in expression of fliC genes is consistent with the transition from a highly aerated shacked liquid culture to a situation where SA187 has stably colonized the plant root.
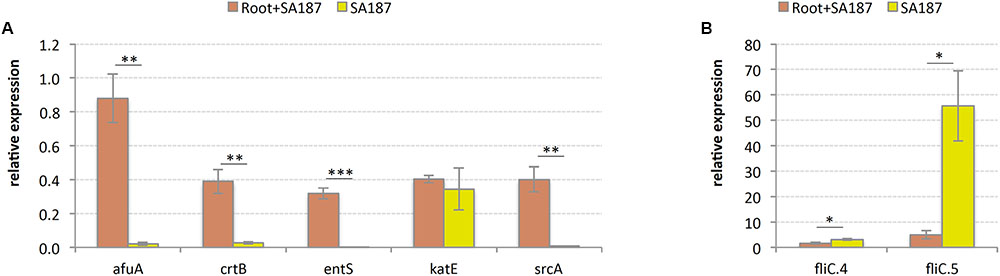
FIGURE 4. Relative gene expression by RT-qPCR. (A) SA187 genes increasing their expression upon association with Arabidopsis roots. afuA = iron(III) ABC transporter substrate-binding protein, crtB = phytoene synthase, entS = MFS transporter ENTS family enterobactin (siderophore) exporter, katE = catalase, srcA = PTS system, sucrose-specific IIB component. (B) SA187 genes decreasing their expression upon association with Arabidopsis roots. fliC = flagellin. ∗ = significant (p < 0.05), ∗∗ = very significant (p < 0.01), ∗∗∗ = extremely significant (p < 0.001).
Biochemical Characteristics of SA187
The qualitative evaluation of PGP traits showed that SA187 was able to produce siderophores and also to solubilize zinc, but was unable to solubilize phosphate (Table 3). These results revealed some of the possible strategies that the bacterium might employ when interacting with its host plant, contributing to plant growth promotion. The obtained antibiogram showed that SA187 is resistant to both ampicillin and penicillin G, but sensitive to chloramphenicol, erythromycin, kanamycin, oxytetracycline, streptomycin, tetracycline and rifampicin (Table 4). The resistance to ampicillin and penicillin G is providing SA187 with an additional strategy to compete with other coexisting bacteria and fungi in the desert soil and in the rhizosphere.

TABLE 3. Biochemical characteristics of SA187.

TABLE 4. Resistance of SA187 to different antibiotics.
SA187 grew well under osmotic and heat stress conditions, as well as under salt stress, up to a concentration of 1 M NaCl (Table 5). When comparing the growth of the bacterium under increasing concentrations of NaCl, we observed that supplementing LB broth with low concentrations of NaCl (0.1–0.5 M) had a beneficial effect on bacterial growth. The maximum cell density in these cases (OD600), when compared with LB with no additional NaCl added, increased about 0.2 units. The growth of SA187 was in contrast slightly delayed when 1 M NaCl was added, and completely inhibited at higher concentrations (LB+2 M NaCl and LB+4 M NaCl) (Table 5 and Supplementary Figure S5A). These results indicate that SA187 is moderately halophilic, being able to resist NaCl concentrations up to 1 M. Similar results were recently reported for Klebsiella sp. BRL6-2, which was able to grow under salt conditions up to 1.5 M NaCl (Woo et al., 2014). The capability to resist moderate salt stress would contribute to the survival of SA187 as a free-living bacterium in desert soils.
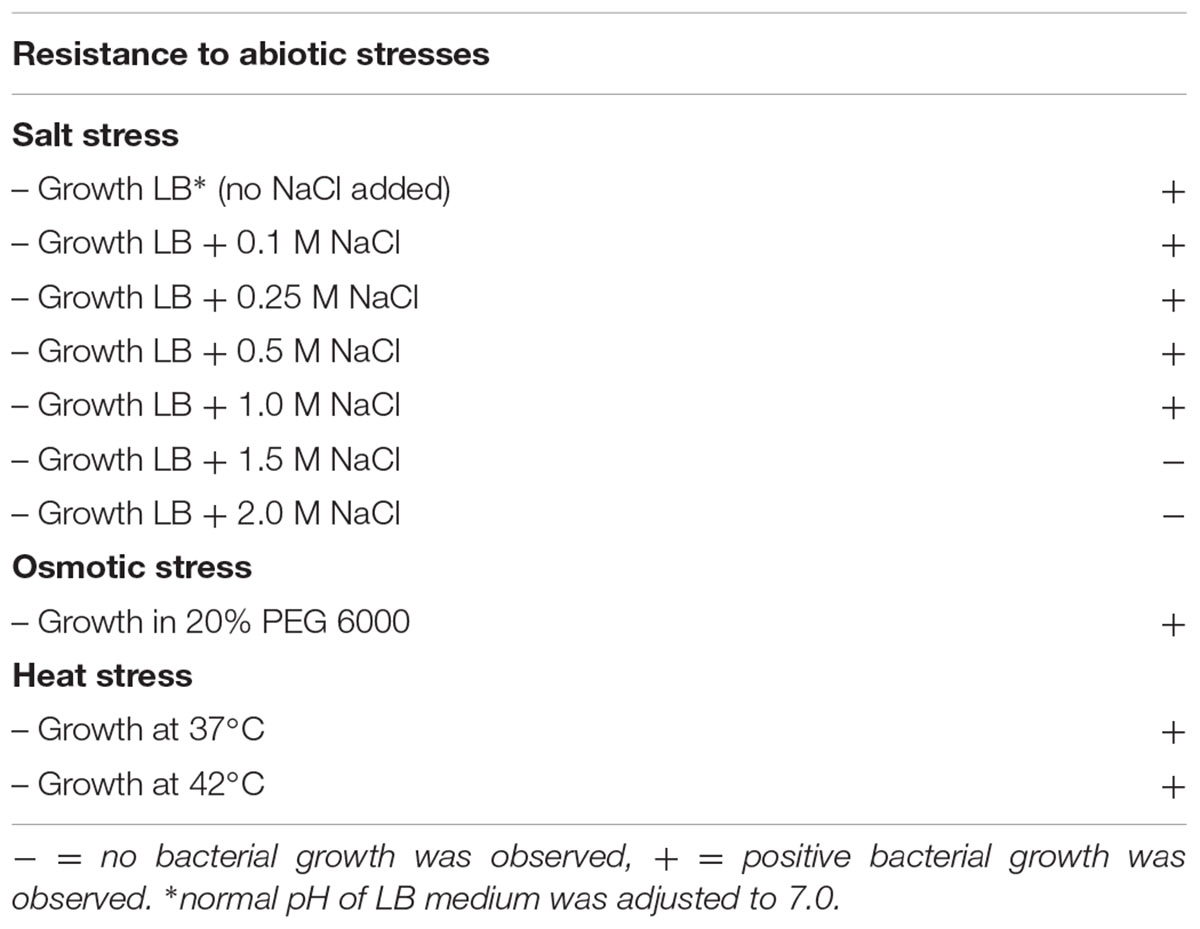
TABLE 5. Resistance of SA187 to different abiotic stresses.
The evaluation of the growth of SA187 in different carbon sources showed that the bacterium grows fast in LB broth (containing 0.5% NaCl), as expected for this nutrient rich medium. Consistently with the lack of a utilizable carbon source in ½MS, SA187 did not show any growth when no sugar was added to this medium. SA187 did not grow in ½MS supplemented with 1% acetic acid or 1% lactic acid. SA187 did nevertheless grow in ½MS supplemented with 0.1–1% sucrose, 1% arabinose or 1% citric acid, although the log phase was clearly delayed (>12 h) compared to LB (Supplementary Figure S5B). SA187 was also able to grow in ½MS+1% glycerol and ½ MS+1% lactose to some extent, although the growth rates in these cases were strongly reduced. Finally, compared with ½MS the growth of SA187 was slightly stimulated when 1% fructose, 1% raffinose, 1% glucose, or 1% maltose were added (Supplementary Figure S5B).
Additionally, the pH of the culture broth has a clear influence on the bacterial growth. The good growth in LB compared with all other conditions might be also due to the higher pH to which the medium is adjusted (pH 7.0), compared with the pH in ½ MS (pH 5.8). Interestingly, no big difference was observed when SA187 was grown in LB pH 7.0 or LB pH 5.8 (Supplementary Figure S5B), indicating the bacterium is capable to resist certain acid stress due to a lower pH.
Metabolic Features of SA187 Involved in the Dual Life-Style: Free-Living and Plant-Associated
A general overview of the main metabolic pathways and transport systems involved in the interactions between SA187 and its plant-host is presented in Figure 5.
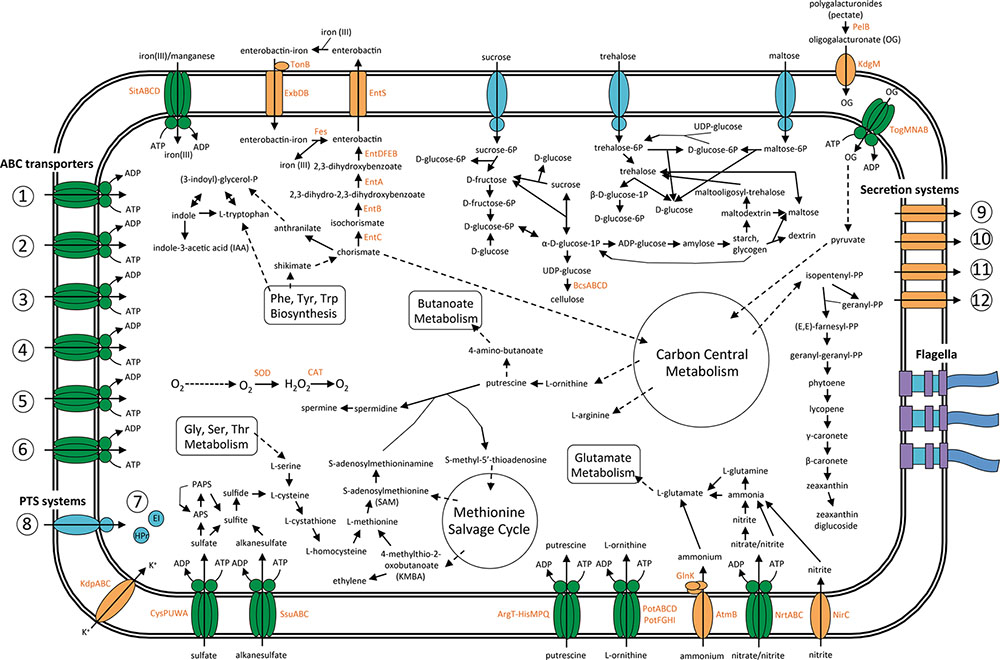
FIGURE 5. Overview of the metabolism and transport in endophytic bacterium SA187. Predicted pathways involved in plant–microbe interaction, based on the current annotation of the complete genome sequence SA187. Metabolic flow through each pathway is designated by the direction of the arrow connecting two metabolites. Import or export of solutes is designated by the direction of the arrow through the transporter. ABC transporters: (1) Mineral and organic ion transporters: Sulfate, molybdate, nitrate/nitrite, taurine, sulfonate, iron(III), thiamine, spermidine/putrescine, betaine/proline, osmoprotectant; (2) Saccharide, polyol, lipids transporters: Maltose/maltodextrine, L-arabinose, oligogalacturonide, methyl-galactoside, D-xylose, autoinducer-2 (AI-2), rhamnose, ribose, glycerol-3-phosphate, phospholipid, multiple sugar, simple sugar; (3) Phosphate and amino acid transporters: Phosphate, histidine, glutamine, arginine, glutamate/aspartate, general L-amino acid, cysteine, branched-chain amino acid, urea, D-methionine, polar amino acid; (4) Peptide and nickel transporters: Oligopeptide, dipeptide, cationic peptide, nickel, glutathione, peptide/nickel, microcin C; (5) Metallic cation, iron-siderophore, vitamin B12 transporters: Iron-complex transporter, vitamin B12, zinc, manganese/iron; (6) ABC-2 type and other transporters: lipopolysaccharide, lipoprotein-releasing, putative ABC, ABC-2 type. Phosphotranspherase systems (PTS): (7) Enzyme I and phosphocarrier protein (HPr); (8) Enzyme II: Glucose, N-acetyl-glucosamine, maltose/glucose, maltose, sucrose, beta-glucoside, trehalose, alpha-glucoside, fructose, mannitol, 2-O-A-mannosyl-glycerate, cellobiose, glucitol/sorbitol, galactitol, mannose, fructoselysine, ascorbate. Secretion systems: (9) General (Sec-dependent) secretion system, (10) Tat-dependent secretion system, (11) type 2 secretion system (T2SS), (12) type 6 secretion system (T6SS).
Survival under Extreme Conditions
SA187 was isolated from I. argentea, an indigenous desert plant growing in areas where heat, drought and salt are key environmental stresses with which bacteria must cope during their free-living lifestyle in the soil. Betaine (also called glycine-betaine) and proline are known to confer salt tolerance to organisms (Ren et al., 2010; Hayat et al., 2012). P-blast genome mining revealed that SA187 contains the complete pathway for the biosynthesis of proline. However, the genes coding for choline dehydrogenase (betA) and betaine dehydrogenase (betB) are not present, indicating that SA187 is unable to synthesize betaine. On the other hand, we found a set of genes coding for membrane transporters: ATP-binding cassette (ABC) transporters ProVWX (SA187PBcda_000000076-000000079) and OpuABCD (SA187PBcda_000004092-000004095), and 3 copies of an major facilitator superfamily (MFS) transporter metabolite:H+ symporters (MHS) family, ProP (SA187PBcda_000002035, SA187PBcda_000002396, SA187PBcda_000004343), which can be used to internalize these osmoprotectants, which might be released into the rhizosphere by other microorganisms and plants. A novel role for ProP as carnitine uptake system has been recently described in Cronobacter sakazakii BAA-894 (member of the family Enterobacteriaceae). The uptake of carnitine by ProP provided the strain with a higher osmotolerance, being able to grow under salt concentrations far in excess of that afforded by proline (Feeney and Sleator, 2015).
Trehalose is another important osmoprotectant produced under environmental stresses. Five pathways for the biosynthesis of trehalose have been described: TreS, OtsA/OtsB, TreP, TreT, TreY/TreZ (Garg et al., 2002; Paul et al., 2008). In the genome of SA187 we found genes coding for trehalose 6-phosphate synthase and trehalose 6-phosphate phosphatase (otsAB, SA187PBcda_000004793-000004794), 2 copies of trehalose-6-phosphate hydrolase (treC, SA187PBcda_000000633, SA187PBcda_000002209), malto-oligosyltrehalose trehalohydrolase (treZ, SA187PBcda_000002137) and the transcriptional regulator treR (SA187PBcda_000002211). Additionally, we found 2 copies of treA (SA187PBcda_000001180, SA187PBcda_000004582) coding for the enzyme trehalase, which catalyzes the hydrolysis of trehalose into glucose, as well as 2 copies of treB (SA187PBcda_000002210, SA187PBcda_000002946) a PTS that is specific for the uptake of trehalose. The role of trehalose in osmotolerance has been reported recently in Klebsiella sp. BRL6-2. The growth rate of this strain in 6% NaCl containing medium supplemented with trehalose increased significantly when compared with the growth in the absence the of osmoprotectants (Woo et al., 2014).
A recent study reported an ATP-dependent potassium (K+) uptake system (KdpFABC) to be essential for survival of Halobacterium salinarum, an extreme halophilic Gram-negative archeon, under desiccation and high salinity (Kixmüller and Greie, 2012). Homologs of kdpFABC are widely distributed among the family Enterobacteriaceae and other Gram-negative bacteria and cyanobacteria and have been reported to increase their expression in response to salinity (Walderhaug et al., 1989; Jung et al., 1997; Solheim et al., 2014). One of the prominent responses of Salmonella enterica to high ion concentrations has been reported to be the transcriptional induction by more than 100-fold of 2 operons: proU (proVWX), and the kdpABC system (Balaji et al., 2005). The genome of SA187 contains the genes kdp forming an operon that also included the genes for the corresponding two-component system response regulator (KdpED) (kdpEDCBAF, SA187PBcda_000003311-000003316).
Other mechanisms conferring salt tolerance to halophiles is the presence of cation transport systems for the controlled uptake of sodium (Na+), potassium (K+) and chloride (Cl-) (Flowers et al., 2015). The genome of SA187 contains genes coding for cation/proton (H+) antiporters that contribute to osmoregulation: K+/H+ antiporter NhaP2 (SA187PBcda_000004591) and Na+/H+ antiporter NhaA (SA187PBcda_000002407). We also found a transcriptional regulator of the family LysR, activator of NhaA (nhaR, SA187PBcda_000002407-000002408). The NhaP2 and NhaA transport systems allow bacteria to import H+ while pumping K+ and Na+, thus preventing excessive cation accumulation, and have been recently found to resist hyperosmotic stress in the genome of the alkalotolerant plant growth-promoting rhizobacterium Klebsiella sp. D5A (Liu et al., 2016).
In addition to the biosynthesis of different osmoprotectants, the production of carotenoids has also been reported to be important for the survival of the bacteria in the rhizosphere and its protection against UV radiation, as well as for the bacterial-plant association (Mohammadi et al., 2012; Bible et al., 2016). On synthetic media, SA187 produced a yellow pigment that could be due to the biosynthesis of carotenoids. We found that the genome of SA187 contains all 7 genes of the entire carotenoid biosynthesis pathway, which is organized in a gene cluster (crtE-idi-crtXYIBZ, SA187PBcda_000002248-000002254) (Figure 6A) that is syntenic with the cluster identified in C. sakazakii BAA894, a close relative of the genus Enterobacter (Zhang et al., 2014). This Cronobacter strain also forms yellow colonies when grown on agar plates, due to the production of two carotenoids: zeaxanthin-monoglycoside and zeaxanthin-diglucoside (Zhang et al., 2014). The conserved synteny of the gene cluster present in SA187 and the one identified in C. sakazakii BAA894 suggest that the yellow pigment produced by SA187 could be due to the same zeaxanthins. The importance of carotenoids for an effective plant–microbe interaction was revealed by Pantoea sp. YR343 ΔcrtB mutant. This mutant strain was defective in the production of carotenoids, due to the lack of phytoene synthase (crtB), a homolog of this gene is found in SA187, and was reported to show increased sensitivity to oxidative stress, impaired biofilm formation and production of IAA, and the reduced colonization of plant roots (Bible et al., 2016). By homology with the same genes in C. sakazakii BAA894 and other Enterobacteriaceae strains, including Pantoea sp. (Misawa et al., 1990; Sedkova et al., 2005; Conlan et al., 2014; Zhang et al., 2014), we hypothesized that the yellow pigment produced by SA187 could be a derivate of the carotenoid zeaxanthin, and could have a role in Arabidopsis root colonization. Additionally, zeaxanthin is a precursor in the plant biosynthetic pathway to produce salicylic acid (SA), a plant hormone that could have a role in the PGP provided by SA187.
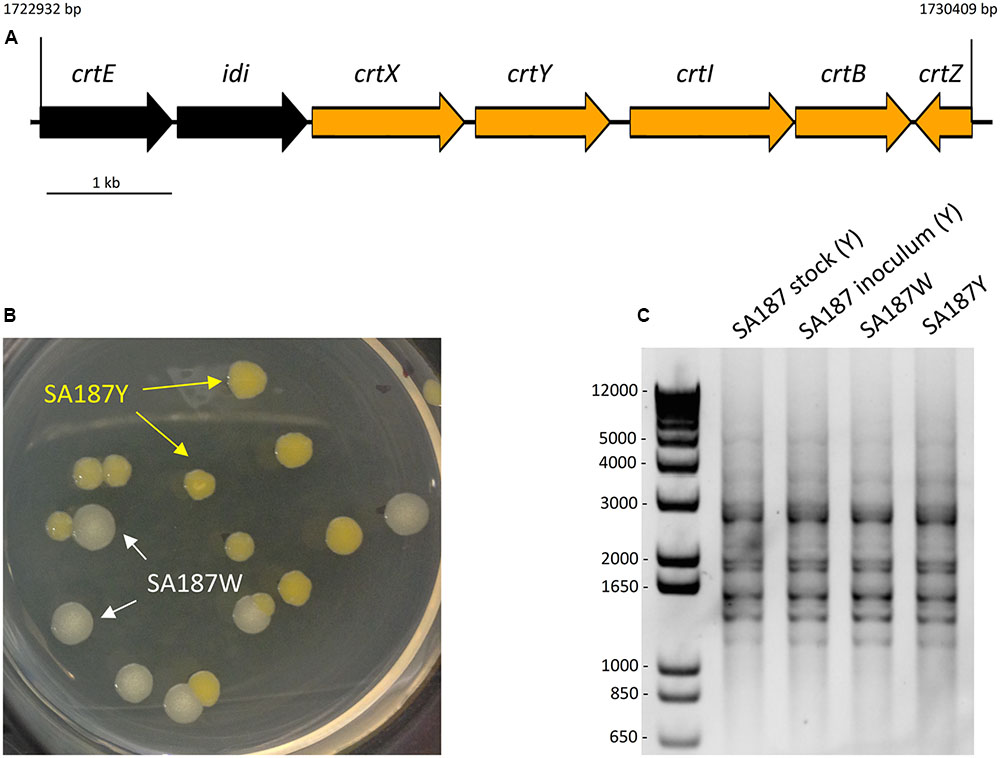
FIGURE 6. (A) Carotenoid biosynthesis gene cluster. (B) SA187 multi-phenotypic complex. After a certain period of interaction between SA187 and the plant host Arabidopsis, 2 morphologies differing in the pigmentation are observed: yellow (SA187Y) and white (SA187W). (C) (GTG)5-rep-PCR fingerprinting. Genotypic characterization of the SA187Y and SA187W isolates, comparison with the original SA187 stock and the inoculum used for one of the screening experiments. The same amplification pattern shown by the four samples shown in the panel indicates that both phenotypes, yellow and white, are genetically identical.
Two-component systems (TCS) are signaling pathways that allow bacteria to sense and respond rapidly to changes in their environment. TCS consist of a sensor membrane-bound histidine kinase (HK) and a corresponding response regulator (RR) (Mitrophanov and Groisman, 2008). In SA187, a high number of genes are involved in TCS systems and signal transduction (Supplementary Table S7). Most of the SA187 TCS belong to the OmpR family, but also systems belonging to the NarL and NtrC families were identified. Among them, the KdpD/KdpE system (mentioned before), one of the most distributed HK/RR systems in bacteria, which is typically activated under K+ limitation or osmotic stress (Heermann and Jung, 2012). We also found the CpxA/CpxR TCS, which controls the envelope stress response in Gram-negative bacteria (cpxAR, SA187PBcda_000001596-000001597). In Escherichia coli, the CpxA/CpxR system, jointly with the sigmaE and sigma32 response pathways, regulates gene expression in response to adverse conditions (De Wulf and Lin, 2000). A large number of TCS genes is typical for bacteria living in rapidly changing or diverse environments (Capra and Laub, 2012), and correlates with the dualistic life style of SA187 as free-living and plant-associated microorganism.
Transport and Exchange of Nutrients
Bacteria living in endophytic association need to exchange nutrients (Chibucos and Tyler, 2009). Consistently, we found that SA187 codes for a large diversity of transporters to allow the exchange of bacterial metabolites and plant-produced nutrients. Among these transporters, we identified more than 200 genes coding for ABC transporters, which among other things, are involved in the uptake of metals (iron, manganese, nickel, molybdate, zinc), phosphate, sulphate, nitrate/nitrite, urea, sugars (glycerol-3P, ribose, rhamnose, xylose, maltose/maltodextrine, arabinose), amino acids (glycine-betaine/proline, methionine, cysteine, arginine, branched-chain amino acids, glutamine/aspartate, histidine, lysine/arginine/ornithine) polyamines (spermidine/putrescine), or quorum sensing autoinducer-2 (AI-2) (Supplementary Table S8). AI-2 and LuxS, the enzyme that catalyzes the production of the signal precursor for AI-2 mediated quorum sensing, has been reported in Enterobacteriaceae of the genera Enterobacter, Klebsiella, and Pantoea that live in close association with plants (Rezzonico et al., 2012).
SA187 is also able to incorporate a wide plethora of sugars through PTS. We identified 101 genes involved in the uptake of glucitol/sorbitol, lactose/cellobiose, galactitol, mannitol, fructose, ascorbate, trehalose, or mannose, among others (Supplementary Table S9). We also found 53 genes coding for members of the MFS transporters, such as the MHS family proline/betaine transporter ProP (mentioned before) (Supplementary Table S10). These results are consistent with the capacity of SA187 to grow on different carbon sources, as described previously. SA187 was able to grow in media with arabinose and, to some extent, with glycerol or lactose. SA187 is also able to incorporate sucrose, fructose, glucose or maltose, what is consistent with its growth on ½MS when the corresponding carbon sources were added (Supplementary Figure S5B).
Additionally, bacteria have developed several specific mechanisms to compete for iron, an essential element whose availability often limits bacterial growth. These mechanisms include specific iron uptake transporters, the secretion of large numbers of diverse siderophores and the synthesis of siderophore receptors to utilize siderophores produced by other microorganisms. The presence of an efficient iron uptake system can also contribute to protect the host plant against pathogen infections, by depriving iron from the pathogenic microorganisms (Taghavi et al., 2010). We found 54 genes involved in iron- and manganese-uptake by SA187 (Supplementary Table S11). Among them, several iron ABC transporters: SitABCD, 2 AfuABC, 4 FhuDBC as well as six genes coding for the iron complex-outer membrane receptor FhuA; two ferrous iron uptake transporters: FeoABC and EfeUO; and the ferrous-ion efflux pump FieF (Supplementary Table S11). An important role of iron uptake in the interaction between SA187 and the plant is supported by the observed increase in gene expression of the genes afuA and entS (Figure 4A). These transporters for iron uptake are common among members of the Enterobacteriaceae family, including PGPR Enterobacter sp. 638 and Klebsiella sp. D5A (Boyer et al., 2002; Taghavi et al., 2010; Liu et al., 2016).
Similarly to other Enterobacteriaceae (Taghavi et al., 2010; Carpenter and Payne, 2014), SA187 contains the genes necessary to synthesize the siderophore (ferric chelator) enterobactin: entD (SA187PBcda_000005352), entF (SA187PBcda_000005348), entABEC (SA187PBcda_000005338-000005341), to secrete it through an MFS transporter (entS, SA187PBcda_000005343), and to recover the enterobactin-iron complex through the TonB-dependent transporter ExbDB (SA187PBcda_000000488-000000489). Finally, the enterobactin esterase Fes (SA187PBcda_ 000005350) will liberate the iron molecule. An additional TonB-dependent outer membrane iron-enterobactin/colicin (fepA, SA187PBcda_000005351) was also identified. The production of enterobactin has been reported in E. coli BW25113 and E. cloacae ATCC 13047, were it has been found to be positively regulated by the peroxiredoxin AhpC (Ma and Payne, 2012; Carpenter and Payne, 2014).
Phosphorus (P) is another element that is an essential macronutrient for the growth of all biota, including plants, and, together with nitrogen, it is one of the major limiting macronutrients for crop production (Bergkemper et al., 2016). Plants are only able to take up free orthophosphate (PO43-), but phosphate in the soil is mostly present in the form of insoluble compounds. Therefore, specialized microorganisms such as PGPB play an important role in providing available inorganic P to the plant, in the form of PO43- (Bergkemper et al., 2016). In most bacteria, a mineral phosphate-dissolving capacity has been shown to be related to the production of organic acids, and the direct oxidation of glucose to gluconic acid (GA) has been proposed as the main mechanism for mineral phosphate solubilization in Gram-negative bacteria. This oxidation is carried out by the glucose dehydrogenase (GDH) enzyme and the cofactor pyrroloquinoline quinone (PQQ) and has been identified, among others, in the phosphate solubilizers E. asburiae and Leclercia sp. QAU-66 (Gyaneshwar et al., 1999; Rodríguez et al., 2007; Naveed et al., 2014). The genome of SA187 contains only the gene coding for PqqF. The lack of the operon pqqABCDE eliminates this pathway as the strategy used for phosphate solubilization. Nevertheless, we found that SA187 can synthetize 2 exopolyphosphatases (ppx-gppA, SA187PBcda_000001262, SA187PBcda_000005395) and an inorganic pyrophosphatase (ppa, SA187PBcda_000002182), which are three enzymes that have been shown to be involved in making insoluble phosphorus available for to plants (Bhattacharyya et al., 2017). The genome of SA187 contains genes coding for the TCS PhoB/PhoR involved in the phosphate starvation response (phoBR, SA187PBcda_000002805-000002806) and an ABC transporter for phosphate uptake (pstSCAB, SA187PBcda_000001427- SA187PBcda_000001430). Moreover, we also identified a low-affinity inorganic phosphate transporter (pit, SA187PBcda_000001153). The genes phoA and phoD, coding for enzymes alkaline phosphatases, which release PO43- and acts downstream the PhoBR system, were not found. The Pst transporter is repressed by phosphate and induced under phosphate limitation, while the Pit system is constitutive (Jansson, 1988). Despite the potential capability of SA187 to incorporate phosphate (PO43-) through these systems, the bacterium was unable to experimentally solubilize phosphate, as shown by our biochemical analysis (Table 3), suggesting that the transporters might be non-functional or not expressed under the experimental conditions.
Together with P and K+, nitrogen is one of the most important micronutrients for the plant. A number of Enterobacteriaceae, including Enterobacter oryzae Ola51T, E. agglomerans, E. cloacae, or L. adecarboxylata STUPM20, have been reported to be nitrogen-fixers (Kreutzer et al., 1991; Peng et al., 2009; Laili et al., 2017). Interestingly, the genome of SA187 lacks genes coding for the nitrogenase enzyme (nifDHK), required for nitrogen fixation, but contains genes for dissimilatory nitrate reduction (Supplementary Table S12): narLXK (SA187PBcda_000004490-000004493), narGHIJ (SA187PBcda_000004486-000004489), nirBD (SA187PBcda_000001018-000001019), nirC (SA187PBcda_000001020) and nitrate assimilation: nasAB (SA187PBcda_000004496-000004497). We also found an ammonium uptake transporter and its regulator (amtB and glnK, SA187PBcda_000002879-000002880), a periplasmic nitrate reductase (napA, SA187PBcda_000005261), a nitrate/nitrite ABC transporter (nasDEF/nrtABC, SA187PBcda_000004498-000004500) and the nitrate RR NasR (SA187PBcda_000004502). These results indicate that SA187 is potentially able to incorporate nitrate and nitrite for assimilation into ammonia, as well as to incorporate ammonia directly.
Secretion of Effector Proteins
Besides the uptake and exchange of nutrients, bacteria also need a set of different protein secretion systems that are essential for their growth and for their interaction with plants. Through these systems, bacteria secrete enzymes, peptides, toxins, antibiotics or secondary metabolites to the surrounding environment, to compete with nearby microorganisms or to be incorporated and used by their host plant (Green and Mecsas, 2016). Among the bacterial secretion systems, the general secretion (Sec) and the twin-arginine translocation (Tat) pathways are most commonly used to transport proteins across the plasma membrane (Natale et al., 2008). The Sec pathway primarily secretes unfolded proteins, while the Tat pathway is mostly used to secrete folded proteins (Robinson and Bolhuis, 2004). Most of the proteins transported by these pathways remain inside the cell, but in Gram-negative bacteria, they can either stay in the periplasm or the inner membrane, or they can be secreted outside through the type II (T2SS) or type V (T5SS) secretion systems (Green and Mecsas, 2016). In the genome of SA187, we identified 35 genes involved in bacterial secretion systems. Among them, we identified a complete Sec and Tat secretion pathways and most of the T2SS and type VI (T6SS) secretion systems (Supplementary Table S13). We did not find any of the genes required for the biosynthesis of type III (T3SS) nor T5SS.
The T2SS is conserved in most Gram-negative bacteria, and is unique in its ability to promote secretion of large and sometimes multimeric proteins that are folded in the periplasm and transport them into the extracellular environment (Douzi et al., 2012; Green and Mecsas, 2016). Although T2SS has been found in many plant pathogens such as Pseudomonas fluorescens, Erwinia, or Xanthomonas spp., T2SS is also important for non-pathogenic bacteria, such as the metal reducing bacteria Shewanella oneidensis (Douzi et al., 2012; Korotkov et al., 2013). In the genome of SA187 we found almost all core genes coding for the T2SS, which were organized in an operon (gspCDEFGHIJKLM, SA187PBcda_000000965-000000975). Only the genes gspO, coding for a prepilin peptidase, and gspS, coding for an accessory pilotin, were not found. As non-core component, genes encoding GspS have not been found in all bacterial species. Nevertheless, the T2SS core protein GspO seems to be essential for the secretion system to be functional (Douzi et al., 2012). Although no gene coding for GspO was found in the genome of SA187, we found two genes showing a high homology, coding for a type IV prepilin-like proteins peptidase (pilD, SA187PBcda_000000976, SA187PBcda_000000320), which could be used instead, rendering SA187 with a fully functional T2SS secretion system.
The T6SS, on the other hand, is the most recent bacterial secretion system to be discovered and also fairly well conserved among Gram-negative bacteria. Although it is still poorly characterized, T6SS translocates effector proteins into a variety of recipient cells, including eukaryotic cells and other bacteria and has been reported in a well-studied PGPB strain P. fluorescens (Decoin et al., 2014; Green and Mecsas, 2016). Many of these effectors are directed against the bacterial cell wall and membrane, supporting a role in bacterial competition with other microorganisms (Russell et al., 2011, 2014). Similarly to T2SS, T6SS has been found in pathogenic as well as in non-pathogenic bacteria (Shyntum et al., 2014). In the case of SA187, we identified most of the genes coding for T6SS, including three copies of the gene coding for the Hcp protein, (SA187PBcda_000000063, SA187PBcda_000001145, SA187PBcda_000001383), but none of the three post-translational regulators: PpkA, Fha1 and Stp1. These regulatory proteins belong to the non-core set of genes and are not essential for the biosynthesis of a functional secretion system (Shyntum et al., 2014).
Chemotaxis and Bacterial Mobility
Motility is an important characteristic for plant-associated bacteria and endophytes, enabling bacteria to move and colonize plants and also to systematically spread within the plant (Hardoim et al., 2008). We identified 156 genes involved in chemotaxis and biosynthesis and assembly of flagella (Supplementary Table S14). The most widespread bacterial chemotaxis signaling pathway centers on a fixed core of signaling genes, consisting in the TCS CheA/CheY, methyl-accepting chemoreceptor proteins (MCP) and an adaptor protein CheW (Capra and Laub, 2012). Consistently with its chemotactic nature, we found that SA187 is able to synthetize the TCS CheA/CheY (cheA, SA187PBcda_000004788; cheY, SA187PBcda_000004777), as well as CheW (SA187PBcda_000004787) and a wide variety of MCPs (10 mcp, tap, 3 tar, 2 tgr, 4 tsr) (Supplementary Tables S6, S14). We also found genes coding for additional chemotaxis proteins: CheZYBR (SA187PBcda_000004776-000004779), CheWA (SA187PBcda_000004787-000004788), the TCS RR CheV (SA187PBcda_000005252) and CheB/CheR fusion protein (cheBR, SA187PBcda_000000470), and also several copies of the genes coding for fimbrial proteins FimA, FimC, and FimD (Supplementary Table S14).
The mobile nature of SA187, which allows the bacterium to move through the soil matrix and inside the plant, was confirmed by the presence of a large number of genes involved in the biosynthesis and assembly of flagella, such as 2 operons flgABCDEFGHIJKLMN, 2 flhAB, flhCD, flhE, 2 fliA, fliB, 2 fliCD, 3 fliC, fliEFGHIJKLMN, fliEFGHIJKLMNOPQRST, as well as and 2 copies of genes coding for the flagellar motor proteins MotA and MotB. We also found genes involved in the biosynthesis and assembly of the type IV pilus system (T4PS) (hofBC, hofMNOPQ, tcpC, tcpD, tcpE, tcpT) and pilin (2 pilD, ppdABC, tcpA, tcpB) (Supplementary Table S14).
It is known that T2SS and T4PS are evolutionary related and shared several structural and functional features, such as the prepilin peptidase PilD, as mentioned before (Korotkov et al., 2013). Interestingly, we observed that, in the genome of SA187, most of the genes involved in the biosynthesis of flagella, as well as a set of genes coding for T4PS are grouped in clusters (Figure 1). The high number of flagella synthetized by SA187 can be seen in the negative-stained transmission electron microscopy sections shown in Figure 7A. Interestingly, 5 flagellin (FliC) paralogs containing the N-terminal conserved flg22 motif were found in the genome of SA187. This Flg22 motif is the main pathogen-associated molecular pattern (PAMP) motif that induces PAMP-triggered immunity when plants sense bacteria, such as P. aeruginosa or S. enterica (García and Hirt, 2014; Bigeard et al., 2015) (Figure 7B). Since different host plants including Arabidopsis recognize SA187 as a beneficial bacterium and not a pathogen, it is clear that FliC-induced PAMP-triggered defense responses must be suppressed by SA187 through a yet unknown mechanism.
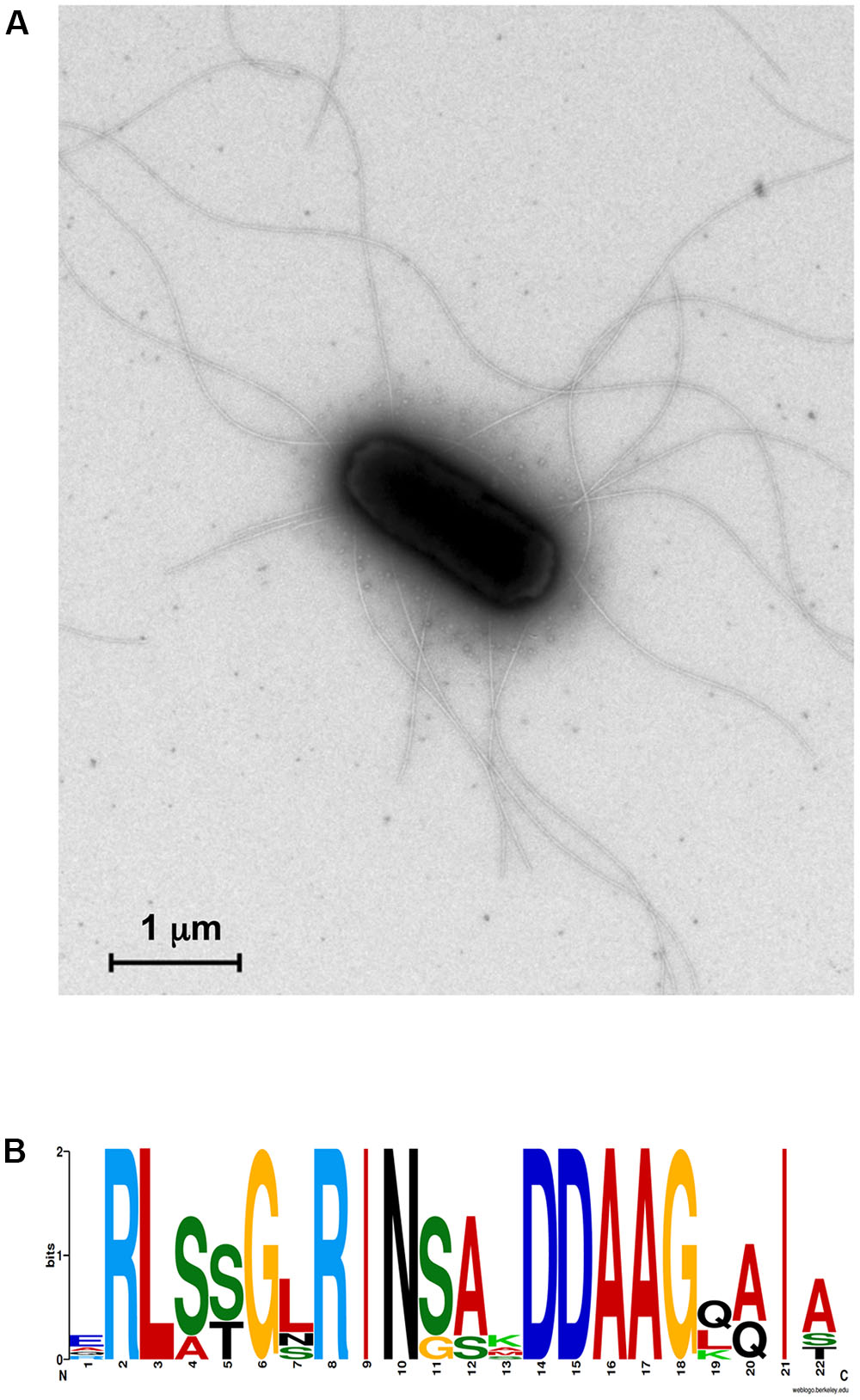
FIGURE 7. (A) Transmission electron microscopy section of SA187. Bacteria were cultured in LB broth before fixation and negative staining. SA187 shows a high number of peritrichous flagella. (B) Conservation of flg22 motif. The N-terminal of FliC proteins of SA187 shown a highly conserved motif shared with Pseudomonas flg22. Diagram obtained by using WebLogo on-line tool (Crooks et al., 2004).
Plant Colonization
To mediate the adhesion and colonization of plant roots, a variety of plant-associated bacteria produce cellulose and other exopolysaccharides (Römling and Galperin, 2015). The genome of SA187 contains all genes necessary to synthesize cellulose (bcsABCD) (Supplementary Table S15). In some members of the Enterobacteriaceae, cellulose is usually co-expressed with curli fibers (Römling, 2007). In the genome of SA187 we found 2 operons coding for curli fibers: csgGFED (SA187PBcda_000003728-000003731) and csgBAC (SA187PBcda_000003733-000003735), which were found adjacent, separated by a hypothetical protein (SA187PBcda_000003732).
Colanic acid is another exopolysaccharide produced by many Enterobacteriaceae and critical for biofilm formation (Rättö et al., 2006). The genome of SA187 contains all genes necessary for the biosynthesis of colanic acid, organized in a gene cluster, together with the proteins required for its translocation to the bacteria cell surface: Wza, Wzb, and Wzc (SA187PBcda_000005052-000005070). The colanic acid transcriptional regulator McbR (SA187PBcda_000004274) was also identified in a separate genomic region. Additionally, we found several genes involved in the putative adhesion to roots, as well as the operon srfABC (SA187PBcda_000004281-000004283), which codes for virulence effectors homologs to those identified in S. enterica, which are believed to be involved in host colonization (Worley et al., 2000; Frye et al., 2006).
It has been reported that the endophytic bacterium E. asburiae JM22 is able to hydrolyze plant cell wall-bound cellulose to gain access to the plant cell (Hallmann et al., 1997). The genome of SA187, however, does not encode any endo-/exo- or hemi-cellulases, suggesting that SA187 could gain entry into the host plant through injuries, points of damaged tissue or natural openings, as described for Enterobacter sp. 638 (Taghavi et al., 2010). Similarly to this well characterized PGPB, we found that SA187 is able to degrade and utilize pectin, as a gene coding for pectate lyase (pelB, SA187PBcda_000002287), an enzyme involved in the cleavage of pectate into oligosaccharides, was identified next to an oligogalacturonate-specific porin (kdgM, SA187PBcda_000002288) (Taghavi et al., 2010). We also found an oligogalacturonide ABC transporter (togMNAB, SA187PBcda_000000314-000000317) involved in the translocation of this pectic oligosaccharide across the inner membrane and genes involved in the degradation of oligogalacturonide (ogl, SA187PBcda_000000332; kduDI, SA187PBcda_000000330-000000331; kdgKA, SA187PBcda_000001733-000001734), was well as the transcriptional regulator kdgR, SA187PBcda_000001735) and 2 copies of an additional galacturonate uptake transporter exuT (SA187PBcda_000000661, SA187PBcda_000003517), and its negative regulator exuR (SA187PBcda _000000662) (Valmeekam et al., 2001). This strategy has been also reported for others PGPB, such as Bacillus amyloliquefaciens subsp. plantarum B9601-Y2 (He et al., 2012). Alternatively, we found that SA187 may degrade galacturonate through a pathway involving the enzymes coded by uxaA (SA187PBcda_000000658), uxaB (SA187PBcda_000004197-000004198), and uxaC (SA187PBcda_000000659). Enzymatic complexes UxaABC and Uxa AB has been found to be normally used in E. coli to degrade galacturonate and glucuronate (Rothe et al., 2013).
As described previously, carotenoids play an important role in the survival in the rhizosphere and plant colonization of Pantoea sp. YR343 to A. thaliana and Populus deltoids (Bible et al., 2016). Interestingly, during our experiments to screen the effect of SA187 in the growth and development of Arabidopsis seedlings, we monitored the amount and viability of bacterial cells (CFU/μl) that were associated with plant roots, and observed the appearance of a multiphenotypic complex in showing yellow and white colonies (Figure 6B). The ratio between white colonies (SA187W) and those showing the original yellow phenotype (SA187Y) increased with longer periods of incubation with plants (data not shown). To eliminate the possibility of contamination, we performed analysis of the 16S rRNA gene sequence as well as genotyping of both white and yellow isolates by (GTG)5-rep-PCR fingerprinting. The results obtained from these analyses confirmed both morphologies corresponded to the same bacterial strain (Figure 6C). Based on these results, we can hypothesize that a modification in the metabolism of SA187 occurred upon colonization of the plant, leading to a decrease in the production of carotenoids. Although a clear role of carotenoids in the interaction between SA187 and Arabidopsis was suggested by the increase in gene expression of crtB upon association with the plant (Figure 4A), further investigations are needed to clarify a role of these carotenoids in the interaction of SA187 with its host–plant.
Defense against Oxidative Stress
Upon contact with bacteria, a major plant defense reaction is the production of reactive oxygen species (ROS), nitric oxide and phytoalexins (Hammond-Kosack and Jones, 1996; Zeidler et al., 2004). Therefore, during colonization, endophytes have to survive in a highly oxidative environment. Accordingly, in the genome of SA187 we found a wide variety of enzymes and regulators that help bacteria to cope with oxidative stress, including superoxide dismutase (sod, SA187PBcda_000001593), catalase (katE, SA187PBcda_000005240), Mn-containing catalase (katN, SA187PBcda_000000181), 4 peroxiredoxins (ahpC, SA187PBcda_000002814, SA187PBcda_000003138; bcp, SA187PBcda_000005379; ahpF, SA187PBcda_000003139), 2 osmotically inducible proteins (osmC, SA187PBcda_000000190, SA187PBcda_000002308), iron-dependent peroxidase (SA187PBcda_000003030), cloroperoxidase (SA187PBcda_000001174) and thiol peroxidase (SA187PBcda_000004379). Synteny analysis demonstrated that the gene SA187PBcda_000003918, annotated as hypothetical protein, was in reality a homolog of katE.
Additionally, we found a gene coding for the hydrogen peroxidase sensor OxyR (SA187PBcda_000001637), which activates the expression of genes such as glutathione reductase (katG, gor, SA187PBcda_000001159), ahpC, ahpF, a DNA-protection during starvation protein (dpsA, SA187PBcda_000003479), the transcriptional regulator of ferric uptake (fur, SA187PBcda_000003300) and glutaredoxin (grxA, SA187PBcda_000003553). We also found 3 glutathione S-transferases (GTS) (gts, SA187PBcda_000001274, SA187PBcda_000002004, SA187PBcda_000003538), a glutathione ABC transporter (gsiABCD, SA187PBcda_000003528-000003531), 3 glutathione peroxidases (btuE, SA187PBcda_000003945, SA187PBcda_000004145) and a γ-glutamyl transpeptidase (GGT) (SA187PBcda_000001086). The operon coding for an RND family multidrug efflux pump (acrAB, SA187PBcda_000002911-000002912) that is required for a successful colonization of the host plant (Taghavi et al., 2010; Burse et al., 2004), as well as its transcriptional regulator (acrC, SA187PBcda_000002913), were also encoded in the genome of SA187.
Production of Antimicrobial Compounds and Toxins
Many beneficial bacteria also produce a variety of antimicrobial compounds, thereby enhancing the plant resistance against pathogens. The genome of SA187 contain phzF (SA187PBcda_000004183) and ubiC (SA187PBcda_000001974), two enzymes involved in the biosynthesis of phenazine and 4-hydroxybenzoate, respectively, which are antibiotics against plant pathogenic bacteria (Duan et al., 2013; Gupta et al., 2014). Furthermore, we identified six genes coding for chitinase (gene.SA187PBcda_000000184, gene.SA187PBcda_000000547,gene.SA187PBcda_000000978, gene.SA187PBcda_000000980, gene.SA187PBcda_000003882, gene.SA187PBcda_000005622), a potent enzyme against insects and fungi (Hamid et al., 2013), which has been also identified in PGPB of the genera Enterobacter, Klebsiella, Pantoea, or Serratia (Dinesh et al., 2015; Rodrigues et al., 2016). SA187 can also synthetize proteins involved in resistance against several antimicrobial compounds: ß-lactam, vancomycin and cationic antimicrobial peptide (CAP) (Supplementary Table S16). We also found a number of toxin/antitoxin (T/A) systems (symE/R, relE/B, hipA/B, cptA/B, chpBK/BI, vapB/C, fic/yhfG, hicA/B) (Supplementary Table S17) and the toxin-coregulated proteins TcpA (gene.SA187PBcda_000000329) and TcpE (gene.SA187PBcda_000000321), what is consistent with the dualistic life-style of SA187 as free-living in the rhizosphere and associated with the host–plant (Pandey and Gerdes, 2005).
Plant Hormone Modulation and Promotion of Plant Growth
Many beneficial bacteria have PGPB activity that is mediated by a variety of mechanisms, including the production or inactivation of plant hormones, such as aminocyclopropane-1-carboxilate (ACC) deaminase. ACC deaminase is involved in the metabolism of the immediate precursor of ethylene in the ethylene biosynthesis, and one of the most well-known PGPB traits (Loper et al., 2012; Shen et al., 2013). ACC deaminase has been found predominantly in Pseudomonas and Mesorhizobium strains, but also reported in member of the genus Enterobacter: E. cloacae UW4, E. cloacae CAL2, and rhizospheric E. cloacae and E. cancerogenus, among others (Shah et al., 1998; Holguin and Glick, 2001; Glick, 2014). Interestingly, acdS, coding for ACC deaminase, was not found in the genome of SA187 (Supplementary Figure S6).
Acetoin and 2,3-butanediol are volatile organic compounds (VOCs) emitted by many PGPB to enhance plant growth (Ryu et al., 2003). The main pathway for the production of these VOCs by Enterobacter sp. 638, a PGPB strain closely related to SA187, is via the sequential action of enzymes coded by the operon budABC (Taghavi et al., 2010). The genome sequence of SA187 contains 3 paralogs of the dimeric enzyme coded by budB (SA187PBcda_000001378-000001379, SA187PBcda_000001698-000001699, SA187PBcda_000002514-000002515), acetolactate synthase, but no budA or budC are present. The lack of these genes makes SA187 capable to transform pyruvate into acetolactate, but no further transformation into acetoin or 2,3-butanediol is possible.
Another strategy that PGPB use to enhance plant growth is the synthesis of auxin indole-3-acetic acid (IAA) from tryptophan through indolepyruvate (Taghavi et al., 2009). We found that SA187 contains most of the genes coding for enzymes involved in this pathway, but it lacks the gene coding for indolepyruvate decarboxylase (ipdC). Instead, SA187 encodes the enzyme tryptophanase (tnaA, SA187PBcda_000000047), which was also found in biofilm forming E. coli (Hu et al., 2010), which can transform tryptophan into indole, but cannot produce salicylic acid (SA). There is nevertheless the possibility that IAA could be produced from indole, although the mechanism of this reaction is not yet understood. We also found a gene coding for nitrilase (SA187PBcda_000002715), which could be a possible alternative tryptophan-independent pathway for the biosynthesis of IAA from indole-3-acetonitrile (Bhattacharyya et al., 2017). Alternatively, SA187 could supply the plant with tryptophan itself, which is the source for the de novo synthesis of IAA in plants, through the intermediate indole-3-pyruvate (Zhao, 2012).
Additionally, we found that the genome of SA187 also contains genes coding for arginine decarboxylase (SpeA), agmatinase (SpeB) and spermidine synthase (SpeE) (speABE, SA187PBcda_000002462-000002464). These enzymes allow the transformation of amino acids into PGP substances, the polyamines putrescine, spermine, and spermidine, respectively, which contribute to bacterial fitness, and have been reported in PGPB strains such as B. subtilis OKB105 or Klebsiella sp. D5A (Xie et al., 2014; Liu et al., 2016).
Although SA187 lacks the former common PGP mechanisms, its beneficial effect in promoting the growth of plants under stress has been suggested to be due to the production of ethylene. Several mechanisms have been described for the production of ethylene in microbes, as ethylene-forming enzyme (EFE) or spontaneous oxidation of 2-keto-4-methylthiobutyric acid (KMBA), an intermediate of the methionine salvage pathway (MSP) (Eckert et al., 2014). In the genome of SA187, we identified all genes involved in the MSP, suggesting that SA187 has the potential to produce ethylene in this manner (Supplementary Figure S6).
SA187 Central Metabolism
The genome of SA187 contains genes involved in the central carbon metabolism, including glycolysis (Embden–Meyerhof and Entner–Doudoroff pathways), pyruvate oxidation, tricarboxylic acid cycle, pentose phosphate pathway and glyoxylate cycle (Supplementary Table S18). The presence of these metabolic pathways should provide SA187 with the capacity to metabolize sugars and other carbon sources present in the plant root exudates. We also found that SA187 can utilize lactose, a differentiating characteristic of the genera Escherichia, Enterobacter, Citrobacter, Klebsiella, and Serratia (Guentzel, 1996), and also fructose, mannose and malonate (mdcABCDEFGH, SA187PBcda_000000997-000001004; and the transcriptional regulator mdcR, SA187PBcda_000000996). The capacity of SA187 to metabolize these sugars is consistent with its uptake through ABC transporters or PTS, as described before, and with the capability of SA187 to grow in ½ MS+1% fructose and ½ MS+0.1–1% sucrose (Supplementary Figure S5B). The role of sucrose as carbon source utilized by the bacteria in association with the plant is also supported by the observed increase in gene expression of the sucrose transporter scrA (Figure 4A).
Conclusion
The results of our taxonomic analysis supports the notion that SA187 represents a novel taxon closely related to the genus Kosakonia. Although this suggests the classification of SA187 as a new genus, further investigation is needed to fully characterize its taxonomic position. Additionally, in the genome of SA187, we successfully identified a number of genes conferring the bacterium the characteristics of a dualistic life style, allowing SA187 to survive in the soil under harsh conditions, as well as to colonize, internalize and provide growth promotion to plants through diverse metabolic strategies. However, the mechanism of interaction between SA187 and its host-plant is not yet fully understood. The combination of the present genomic data with comparative studies on gene expression and metabolite production in SA187, alone or in association with plants, will deepen our understanding which specific genes and pathways are induced during the beneficial interaction. Once important genes will be identified, the further phenotypic screening of mutant strains will reveal their role in the plant-bacterial interaction and plant growth promotion. The knowledge obtained can be further translated into comprehensive and more sophisticated strategies to establish sustainable agricultural practices in marginal and arid lands by using endophytic bacteria as biofertilizers to improve crop production.
Author Contributions
MS, FL, and HH conceived the overall study. AdZ established the plant screening assay that led to the selection of SA187. FL extracted and purified SA187 genomic DNA. IA performed gene prediction and annotation of complete genome data. CA-B performed taxonomical analysis based on 16S and MLSA. FL and IA performed whole-genome phylogenetic analysis. AE performed the biochemical characterization of SA187 and growth curves. AB realized the plant growth promotion screening. HA extracted RNA from dual samples CA-B extracted RNA from bacterial samples, performed qPCR and analyzed the genomic data. CA-B and MS wrote the manuscript. HH, VB, and MS coordinated the experiments and the writing of the manuscript.
Funding
This publication is based upon work supported by the King Abdullah University of Science and Technology (KAUST) to HH No. BAS/1/1062-01-01, and by the KAUST Office of Sponsored Research (OSR) under Awards No. URF/1/1976-02 and FCS/1/2448-01 to VB.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We would like to thank all members of the Hirt lab, CDA management team and green house facility in KAUST for the technical assistance and for their help in many aspects of this work. We would like to also acknowledge Dr. Michael F. Mette for the valuable comments and Dr. Lukas Synek for his help to build and format the figures of this paper.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2017.02023/full#supplementary-material
Footnotes
References
Alam, I., Antunes, A., Kamau, A. A., Ba Alawi, W., Kalkatawi, M., Stingl, U., et al. (2013). INDIGO - INtegrated data warehouse of microbial genomes with examples from the red sea extremophiles. PLOS ONE 8:e82210. doi: 10.1371/journal.pone.0082210
Al-Attar, S., Westra, E. R., van der Oost, J., and Brouns, S. J. (2011). Clustered regularly interspaced short palindromic repeats (CRISPRs): the hallmark of an ingenious antiviral defense mechanism in prokaryotes. Biol. Chem. 392, 277–289. doi: 10.1515/BC.2011.042
Andrés-Barrao, C., Saad, M. M., Cabello Ferrete, E., Bravo, D., Chappuis, M. L., Ortega Pérez, R., et al. (2016). Metaproteomics and ultrastructure characterization of Komagataeibacter spp. involved in high-acid spirit vinegar production. Food Microbiol. 55, 112–122. doi: 10.1016/j.fm.2015.10.012
Azua-Bustos, A., and González-Silva, C. (2014). Biotechnological applications derived from microorganisms of the atacama desert. BioMed Res. Int. 2014:909312. doi: 10.1155/2014/909312
Balaji, B., O’Connor, K., Lucas, J. R., Anderson, J. M., and Csonka, L. N. (2005). Timing of induction of osmotically controlled genes in Salmonella enterica serovar Typhimurium, determined with quantitative real-time reverse transcription-PCR. Appl. Environ. Microbiol. 71, 8273–8283. doi: 10.1128/aem.71.12.8273-8283.2005
Bapiri, A., Asgharzadeh, A., Mujallali, H., Khavazi, K., and Pazira, E. (2013). Evaluation of Zinc solubilization potential by different strains of fluorescent Pseudomonads. J. Appl. Sci. Environ. Manage. 16, 295–298.
Bhatnagar, A., and Bhatnagar, M. (2005). Microbial diversity in desert ecosystems. Curr. Sci. 89, 91–100.
Benslama, O., and Boulahrouf, A. (2013). Isolation and characterization of glyphosate-degrading bacteria from different soils of Algeria. Afr. J. Microbiol. Res. 7, 5587–5595. doi: 10.5897/ajmr2013.6080
Benslama, O., and Boulahrouf, A. (2016). High-quality draft genome sequence of Enterobacter sp. Bisph2, a glyphosate-degrading bacterium isolated from a sandy soil of Biskra, Algeria. Genomics Data 8, 61–66. doi: 10.1016/j.gdata.2016.03.005
Bergkemper, F., Schöler, A., Engel, M., Lang, F., Krüger, J., Schloter, M., et al. (2016). Phosphorus depletion in forest soils shapes bacterial communities towards phosphorus recycling systems. Environ. Microbiol. 18, 1988–2000. doi: 10.1111/1462-2920.13188
Bhattacharyya, C., Bakshi, U., Mallick, I., Mukherji, S., Bera, B., and Ghosh, A. (2017). Genome-guided insights into the plant growth promotion capabilities of the physiologically versatile Bacillus aryabhattai strain AB211. Front. Microbiol. 8:411. doi: 10.3389/fmicb.2017.00411
Bible, A. N., Fletcher, S. J., Pelletier, D. A., Schadt, C. W., Jawdy, S. S., Weston, D. J., et al. (2016). A Carotenoid-Deficient Mutant in Pantoea sp. YR343, a bacteria isolated from the rhizosphere of Populus deltoides, is defective in root colonization. Front. Microbiol. 7:491. doi: 10.3389/fmicb.2016.00491
Bigeard, J., Colcombet, J., and Hirt, H. (2015). Signaling mechanisms in pattern-triggered immunity (PTI). Mol. Plant 8, 521–539. doi: 10.1016/j.molp.2014.12.022
Boyer, E., Bergevin, I., Malo, D., Gros, P., and Cellier, M. F. M. (2002). Acquisition of Mn(II) in addition to Fe(II) is required for full virulence of Salmonella enterica serovar Typhimurium. Infect. Immun. 70, 6032–6042. doi: 10.1128/iai.70.11.6032-6042.2002
Brady, C., Cleenwerck, I., Venter, S., Coutinho, T., and De Vos, P. (2013). Taxonomic evaluation ofthe genus Enterobacter based on multilocus sequence analysis (MLSA): proposal to reclassify E. nimipressuralis and E. amnigenus into Lelliottia gen. nov. as Lelliottia nimipressuralis comb. nov. and Lelliottia amnigena comb. nov., respectively, E. gergoviae and E. pyrinus into Pluralibacter gen. nov. as Pluralibacter gergoviae comb. nov. and Pluralibacter pyrinus comb. nov., respectively, E. cowanii, E. radicincitans, E. oryzae and E. arachidis into Kosakonia gen. nov. as Kosakonia cowanii comb. nov., Kosakonia radicincitans comb. nov., Kosakonia oryzae comb. nov. and Kosakonia arachidis comb. nov., respectively, and E. turicensis, E. helveticus and E. pulveris into Cronobacter as Cronobacter zurichensis nom. nov., Cronobacter helveticus comb. nov. and Cronobacter pulveris comb. nov., respectively, and emended description of the genera Enterobacter and Croinobacter. Syst. Appl. Microbiol. 36, 309–319. doi: 10.1016/j.syapm.2013.03.005
Brady, C., Cleenwerck, I., Venter, S., Vancanneyt, M., Swings, J., and Coutinho, T. (2008). Phylogeny and identification of Pantoea species associated with plants, humans and the natural environment based on multilocus sequence analysis (MLSA). Syst. Appl. Microbiol. 31, 447–460. doi: 10.1016/j.syapm.2008.09.004
Bric, J. M., Bostock, R. M., and Silverstone, S. E. (1991). Rapid in situ assay for indoleacetic acid production by bacteria immobilized on a nitrocellulose membrane. Appl. Environ. Microbiol. 57, 535–538.
Burse, A., Weingart, H., and Ullrich, M. S. (2004). The phytoalexin-inducible multidrug efflux pump AcrAB contributes to virulence in the fire blight pathogen, Erwinia amylovora. Mol. Plant Microbiol. Interact. 17, 43–54. doi: 10.1094/MPMI.2004.17.1.43
Capra, E. J., and Laub, M. T. (2012). The evolution of two-component signal transduction systems. Annu. Rev. Microbiol. 66, 325–347. doi: 10.1146/annurev-micro-092611-150039
Carpenter, C., and Payne, S. M. (2014). Regulation of iron transport systems in Enterobacteriaceae in response to oxygen and iron availability. J. Inorg. Biochem. 133, 110–117. doi: 10.1016/j.jinorgbio.2014.01.007
Chibucos, M. C., and Tyler, B. M. (2009). Common themes in nutrient acquisition by plant symbiotic microbes, described by the Gene Ontology. BMC Microbiol. 9(Suppl. 1):S6. doi: 10.1186/1471-2180-9-S1-S6
Conlan, S., Thomas, P. J., Deming, C., Park, M., Lau, A. F., Dekker, J. P., et al. (2014). Single-molecule sequencing to track plasmid diversity of hospital-associated carbapenemase-producing Enterobacteriaceae. Sci. Transl. Med. 6, 254ra126. doi: 10.1126/scitranslmed.3009845
Coulson, T. J., and Patten, C. L. (2015). Complete genome sequence of Enterobacter cloacae UW5, a rhizobacterium capable of high levels of indole-3-acetic acid production. Genome Announc. 3:e00843–15. doi: 10.1128/genomeA.00843-15
Crooks, G. E., Hon, G., Chandonia, J. M., and Brenner, S. E. (2004). WebLogo: a sequence logo generator. Genome Res. 14, 1188–1190. doi: 10.1101/gr.849004
Darling, A. E., Mau, B., and Perna, N. T. (2010). progressiveMauve: multiple genome alignment with gene gain, loss and rearrangement. PLOS ONE 5:e11147. doi: 10.1371/journal.pone.0011147
De Wulf, P., and Lin, E. C. (2000). Cpx two-component signal transduction in Escherichia coli: excessive CpxR-P levels underlie CpxA∗ phenotypes. J. Bacteriol. 182, 1423–1426. doi: 10.1128/JB.182.5.1423-1426.2000
De Zélicourt, A., Al-Yousif, M., and Hirt, H. (2013). Rhizosphere microbes as essential partners for plant stress tolerance. Mol Plant 6, 242–245. doi: 10.1093/mp/sst028
Decoin, V., Barbey, C., Bergeau, D., Latour, X., Feuilloley, M. G., Orange, N., et al. (2014). A type VI secretion system is involved in Pseudomonas fluorescens bacterial competition. PLOS ONE 9:e89411. doi: 10.1371/journal.pone.0089411
Delgado, M., Mendez, J., Rodriìguez-Herrera, R., Aguilar, C. N., Cruz- Hernaìndez, M., and Balagurusamy, N. (2014). Characterization of phosphate-solubilizing bacteria isolated from the arid soils of a semi-desert region of north-east Mexico. Biol. Agric. Hortic. 30, 211–217. doi: 10.1080/01448765.2014.909742
Diab, A., Ageez, A., and Sami, S. (2017). Enterobacter cloacae MSA4, a new strain isolated from the rhizosphere of a desert plant, produced potent biosurfactant used for enhancing the biodegradation of polycyclic aromatic hydrocarbons (PAHs) during the bioremediation of spent motor oil-polluted sandy soil. Int. J. Sci. Res. 6, 2726–2734. doi: 10.21275/art20173636
Dinesh, R., Anandaraj, M., Kumar, A., Bini, Y. K., Subila, K. P., and Aravin, R. (2015). Isolation, characterization, and evaluation of multi-trait plant growth promoting rhizobacteria for their growth promoting and disease suppressing effects on ginger. Microbiol. Res. 173, 34–43. doi: 10.1016/j.micres.2015.01.014
Douzi, B., Filloux, A., and Voulhoux, R. (2012). On the path to uncover the bacterial type II secretion system. Philos. Trans. R. Soc. B Biol. Sci. 367, 1059–1072. doi: 10.1098/rstb.2011.0204
Duan, J., Juang, W., Cheng, Z., Heikkila, J. J., and Glick, B. R. (2013). The complete genome sequence of the plant-growth-promoting bacterium Pseudomonas sp. UW4. PLOS ONE 8:e58640. doi: 10.1371/journal.pone.0058640
Eckert, C., Xu, W., Xiong, W., Lynch, S., Ungerer, J., Tao, L., et al. (2014). Ethylene-forming enzyme and bioethylene production. Biotechnol. Biofuels 7:33. doi: 10.1186/1754-6834-7-33
El-Beltagy, A., and Madkour, M. (2012). Impact of climate change on arid lands agriculture. Agric. Food Secur. 1:3. doi: 10.1186/2048-7010-1-3
Feeney, A., and Sleator, R. D. (2015). Functional creening of the Cronobacter sakazakii BAA-894 genome reveals a role for ProP (ESA_02131) in carnitine uptake. Bioengineered 6, 161–165. doi: 10.1080/21655979.2015.1043500
Felsenstein, J. (1985). Confidence limits on phylogenies: an approach using the bootstrap. Evolution. 39, 783–791. doi: 10.2307/2408678
Fierer, N., Leff, J. W., Adams, B. J., Nielsen, U. N., Bates, S. T., Lauber, C. L., et al. (2012). Cross-biome metagenomic analyses of soil microbial communities and their functional attributes. Proc. Natl. Acad. Sci. U.S.A. 109, 21390–21395. doi: 10.1073/pnas.1215210110
Flowers, T. J., Munns, R., and Colmer, T. D. (2015). Sodium chloride toxicity and the cellular basis of salt tolerance in halophytes. Ann. Bot. 115, 419–431. doi: 10.1093/aob/mcu217
Frye, J., Karlinsey, J. E., Felise, H. R., Marzolf, B., Dowidar, N., McClelland, M., et al. (2006). Identification of new flagellar genes of Salmonella enterica serovar Typhimurium. J. Bacteriol. 188, 2233–2243. doi: 10.1128/JB.188.6.2233-2243.2006
García, A. V., and Hirt, H. (2014). Salmonella enterica induces and subverts the plant immune system. Front. Microbiol. 5:141. doi: 10.3389/fmicb.2014.00141
Garg, A. K., Kim, J. K., Owens, T. G., Ranwala, A. P., Choi, Y. D., Kochian, L. V., et al. (2002). Trehalose accumulation in rice plants confers high tolerance levels to different abiotic stresses. Proc. Natl. Acad. Sci. U.S.A. 99, 15893–15903. doi: 10.1073/pnas.252637799
Glick, B. R. (2012). Plant growth-promoting bacteria: mechanisms and applications. Scientifica (Cairo) 2012:963401. doi: 10.6064/2012/963401
Glick, B. R. (2014). Bacteria with ACC deaminase can promote plant growth and help to feed the world. Microbiol. Res. 169, 30–39. doi: 10.1016/j.micres.2013.09.009
Goswami, D., Dhandhukia, P., Patel, P., and Thakker, J. N. (2014). Screening of PGPR from saline desert of Kutch: growth promotion in Arachis hypogea by Bacillus licheniformis A2. Microbiol. Res. 169, 66–75. doi: 10.1016/j.micres.2013.07.004
Grant, J. R., and Stothard, P. (2008). The CGView server: a comparative genomics tool for circular genomes. Nucleic Acids Res. 36, W181–W184. doi: 10.1093/nar/gkn179
Green, E. R., and Mecsas, J. (2016). Bacterial secretion systems – an overview. Microbiol. Spectr. 4:10.1128/microbiolspec.VMBF-0012-2015. doi: 10.1128/microbiolspec.VMBF-0012-2015
Gu, C. T., Li, C. Y., Yang, L. J., and Huo, G. C. (2014). Enterobacter xiangfangensis sp. nov., isolated from Chinese traditional sourdough, and reclassification of Enterobacter sacchari Zhu et al., 2013 as Kosakonia sacchari comb. nov. Int. J. Syst. Evol. Microbiol. 64, 2650–2656. doi: 10.1099/ijs.0.064709-0
Guentzel, N. M. (1996). “Escherichia, Klebsiella, Enterobacter, Serratia, Citrobacter, and Proteus,” in Medical Microbiology, 4th Edn, ed. S. Baron (Galveston TX: University of Texas Medical Branch at Galveston).
Gupta, A., Gopal, M., Thomas, G. V., Manikandan, V., Gajewski, J., Thomas, G., et al. (2014). Whole genome sequencing and analysis of plant growth promoting bacteria isolated from the rhizosphere of plantation crops coconut, cocoa and arecanut. PLOS ONE 9:e104259. doi: 10.1371/journal.pone.0104259
Gyaneshwar, P., Parekh, L. J., Archana, G., Poole, P. S., Collins, M. D., Hutson, R. A., et al. (1999). Involvement of a phosphate starvation inducible glucose dehydrogenase in soil phosphate solubilization by Enterobacter asburiae. FEMS Microbiol. Lett. 171, 223–229. doi: 10.1111/j.1574-6968.1999.tb13436.x
Hallmann, J., Quadt-Hallmann, A., Mahaffee, W. F., and Kloepper, J. W. (1997). Bacterial endophytes in agricultural crops. Can. J. Microbiol. 43, 895–914. doi: 10.1139/m97-131
Hamid, R., Khan, M. A., Ahmad, M., Ahmad, M. M., Abdin, M. Z., Musarrat, J., et al. (2013). Chitinases: an update. J. Pharm. Bioallied Sci. 5, 21–29. doi: 10.4103/0975-7406.106559
Hammond-Kosack, K. E., and Jones, J. D. (1996). Resistance gene-dependent plant defense responses. Plant Cell 8, 1773–1791. doi: 10.1105/tpc.8.10.1773
Hanna, A. L., Youssef, H. H., Amer, W. M., Monib, M., Fayez, M., and Hegazi, N. A. (2013). Diversity of bacteria nesting the plant cover of north Sinai deserts. Egypt. J. Adv. Res. 4, 13–26. doi: 10.1016/j.jare.2011.11.003
Hardoim, P. R., van Overbeek, L. S., and Elsas, J. D. (2008). Properties of bacterial endophytes and their proposed role in plant growth. Trends Microbiol. 16, 463–471. doi: 10.1016/j.tim.2008.07.008
Hayat, S., Hayat, Q., Alyemeni, M. N., Wani, A. S., Pichtel, J., and Ahmad, A. (2012). Role of proline under changing environments: a review. Plant Signal. Behav. 7, 1456–1466. doi: 10.4161/psb.21949
He, P., Hao, K., Blom, J., Rückert, C., Vater, J., Mao, Z., et al. (2012). Genome sequence of the plant growth promoting strain Bacillus amyloliquefaciens subsp. plantarum B9601-Y2 and expression of mersacidin and other secondary metabolites. J. Biotechnol. 164, 281–291. doi: 10.1016/j.jbiotec.2012.12.014
Hedi, A., Sadfi, N., Fardeau, M. L., Rebib, H., Cayol, J. L., Ollivier, B., et al. (2009). Studies on the biodiversity of halophilic microorganisms isolated from El-Djerid salt lake (Tunisia) under aerobic conditions. Int. J. Microbiol. 2009, 17. doi: 10.1155/2009/731786
Heermann, R., and Jung, K. (2012). “K(+) supply, osmotic stress, and the KdpD/KdpE two-component system,” in Two-Component Systems in Bacteria, eds R. Gross and D. Beier (Würzburg: University of Würzburg).
Herman, R., Provencio, K. R., Herrera-Matos, J., and Torres, R. J. (1995). Resource islands predict the distribution of heterotrophic bacteria in chihuahuan desert soils. Appl. Environ. Microbiol. 61, 1816–1821.
Holguin, G., and Glick, B. R. (2001). Expression of the ACC deaminase gene from Enterobacter cloacae UW4 in Azospirillum brasilense. Microb. Ecol. 41, 281–288. doi: 10.1007/s002480000040
Horvath, P., and Barrangou, R. (2010). CRISPR/Cas, the immune system of bacteria and archaea. Science 327, 167–170. doi: 10.1126/science.117955
Hu, M., Zhang, C., Mu, Y., Shen, Q., and Feng, Y. (2010). Indole affects biofilm formation in bacteria. Indian J. Microbiol. 50, 362–368. doi: 10.1007/s12088-011-0142-1
Jansson, M. (1988). Phosphate uptake and utilization by bacteria and algae. Hydrobiologia 170, 177–189. doi: 10.1007/BF00024904
Jorquera, M. A., Shaharoona, B., Nadeem, S. M., de la Luz Mora, M., and Crowley, D. E. (2012). Plant growth- promoting rhizobacteria associated with ancient clones of creosote bush (Larrea tridentata). Microb. Ecol. 64, 1008–1017. doi: 10.1007/s00248-012-0071-5
Jung, K., Tjaden, B., and Altendorf, K. (1997). Purification, reconstitution, and characterization of KdpD, the turgor sensor of Escherichia coli. J. Biol. Chem. 272, 10847–10852. doi: 10.1074/jbc.272.16.10847
Kanehisa, M., Sato, Y., Kawashima, M., Furumichi, M., and Tanabe, M. (2016). KEGG as a reference resource for gene and protein annotation. Nucleic Acids Res. 44, D457–D462. doi: 10.1093/nar/gkv1070
Kimura, M. (1980). A simple method for estimating evolutionary rate of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 16, 111–120. doi: 10.1007/BF01731581
Kixmüller, D., and Greie, J. C. (2012). An ATP-driven potassium pump promotes long-term survival of Halobacterium salinarum within salt crystals. Environ. Microbiol. Rep. 4, 234–241. doi: 10.1111/j.1758-2229.2012.00326.x
Klappenbach, J. A., Dunbar, J. M., and Schmidt, T. M. (2000). rRNA operon copy number reflects ecological strategies of bacteria. Appl. Environ. Microbiol. 66, 1328–1333. doi: 10.1128/AEM.66.4.1328-1333.2000
Korotkov, K. V., Sandkvist, M., and Hol, W. G. J. (2013). The type II secretion system: biogenesis, molecular architecture and mechanism. Nat. Rev. Microbiol. 10, 336–351. doi: 10.1038/nrmicro2762
Köberl, M., Müller, H., Ramadan, E. M., and Berg, G. (2011). Desert farming benefits from microbial potential in arid soils and promotes diversity and plant health. PLOS ONE 6:e24452. doi: 10.1371/journal.pone.0024452
Köberl, M., Ramadan, E. M., Adam, M., Cardinale, M., Hallmann, J., Heuer, H., et al. (2013). Bacillus and Streptomyces were selected as broad-spectrum antagonists against soil borne pathogens from arid areas in Egypt. FEMS Microbiol. Lett. 342, 168–178. doi: 10.1111/1574-6968.12089
Kreutzer, R., Steibl, H. D., Dayananda, S., Dippe, R., Halda, L., Buck, M., et al. (1991). “Genetic characterization of nitrogen fixation in Enterobacter strains from the rhizosphere of cereals,” in Nitrogen Fixation. Developments in Plant and Soil Sciences, Vol. 48, eds M. Polsinelli, R. Materassi, and M. Vincenzini (Dordrecht: Springer).
Lafi, F. F., Al-Bladi, M. L., Salem, N. M., Al-Banna, L., Alam, I., Bajic, V. B., et al. (2017a). Draft genome sequence of the plant growth-promoting Pseudomonas punonensis strain D1-6 isolated from the desert plant Erodium hirtum in Jordan. Genome Announc. 5:e01437–16. doi: 10.1128/genomeA.01437-16
Lafi, F. F., Alam, I., Geurts, R., Bisseling, T., Bajic, V. B., Hirt, H., et al. (2016a). Draft genome sequence of the phosphate-solubilizing bacterium Pseudomonas argentinensis strain SA190 Isolated from the desert plant Indigofera argentea. Genome Announc. 4:e01431–16. doi: 10.1128/genomeA.01431-16
Lafi, F. F., Alam, I., Geurts, R., Bisseling, T., Bajic, V. B., Hirt, H., et al. (2017b). Draft genome sequence of Enterobacter sp. Sa187, an endophytic bacterium isolated from the desert plant Indigofera argentea. Genome Announc. 5:e01638–16. doi: 10.1128/genomeA.01638-16
Lafi, F. F., Bokhari, A., Alam, I., Bajic, V. B., Hirt, H., and Saad, M. M. (2016b). Draft genome sequence of the plant growth-promoting Cupriavidus gilardii strain JZ4 isolated from the desert plant Tribulus terrestris. Genome Announc. 4:e00678–16. doi: 10.1128/genomeA.01520-16
Lafi, F. F., Ramirez-Prado, J. S., Alam, I., Bajic, V. B., Hirt, H., and Saad, M. M. (2016c). Draft genome sequence of Halomonas elongata strain K4, an endophytic growth-promoting bacterium enhancing salinity tolerance in planta. Genome Announc. 4:e01214–16. doi: 10.1128/genomeA.01214-16
Lafi, F. F., Ramirez-Prado, J. S., Alam, I., Bajic, V. B., Hirt, H., and Saad, M. M. (2017c). Draft genome sequence of plant growth-promoting Micrococcus luteus strain K39 isolated from Cyperus conglomeratus in Saudi Arabia. Genome Announc. 5:e01520–16. doi: 10.1128/genomeA.01520-16
Laili, N. S., Radziah, O., and Zaharah, S. S. (2017). Isolation and characterization of plant growth-promoting rhizobacteria (PGPR) and their effects on growth of strawberry (Fragaria Ananassa Duch.). Banglad. J. Bot. 46, 277–282.
Lee, Z. M., Bussema, C. III, and Schmidt, T. M. (2008). rrnDB: documenting the number of rRNA and tRNA genes in bacteria and archaea. Nucleic Acids Res. 37, D489–D493. doi: 10.1093/nar/gkn689
Liu, W., Wang, Q., Hou, J., Tu, C., Luo, Y., and Christie, P. (2016). Whole genome analysis of halotolerant and alkalotolerant plant growth-promoting rhizobacterium Klebsiella sp. D5A. Sci. Rep. 6:26710. doi: 10.1038/srep26710
Loper, J. E., Hassan, K. A., Mavrodi, D. V., Davis, E. W. II, Lim, C. K., Shaffer, B. T., et al. (2012). Comparative genomics of plant-associated Pseudomonas spp.: insights into diversity and inheritance of traits involved in multitrophic interactions. PLOS Genet. 8:e1002784. doi: 10.1371/journal.pgen.1002784
Louden, B. C., Haarmann, D., and Lynne, A. (2011). Use of blue agar CAS assay for siderophore detection. J. Microbiol. Biol. Educ. 12, 51–53. doi: 10.1128/jmbe.v12i1.249
Luo, H., Gao, F., and Lin, Y. (2015). Evolutionary conservation analysis between the essential and nonessential genes in bacterial genomes. Sci. Rep. 5:13210. doi: 10.1038/srep13210
Ma, L., and Payne, S. M. (2012). AhpC is required for optimal production of enterobactin by Escherichia coli. J. Bacteriol. 194, 6748–6757. doi: 10.1128/JB.01574-12
Makhalanyane, T. P., Valverde, A., Gunnigle, E., Frossard, A., Ramond, J. B., and Cowan, A. (2015). Microbial ecology of hot desert edaphic systems. FEMS Microb. Rev. 39, 203–221. doi: 10.1093/femsre/fuu011
Mapelli, F., Marasco, R., Rolli, E., Barbato, M., Cherif, H., Guesmi, A., et al. (2013). Potential for plant growth promotion of rhizobacteria associated with Salicornia growing in Tunisian hypersaline soils. Biomed. Res. Int 2013:248078. doi: 10.1155/2013/248078
Marasco, R., Rolli, E., Ettoumi, B., Vigani, G., Mapelli, F., Borin, S., et al. (2012). A drought resistance-promoting microbiome is selected by root system under desert farming. PLOS ONE 7:e48479. doi: 10.1371/journal.pone.0048479
Melo, J., Carolino, M., Carvalho, L., Correia, P., Tenreiro, R., Chaves, S., et al. (2016). Crop anagement as a driving force of plant growth promoting rhizobacteria physiology. SpringerPlus 5, 1574. doi: 10.1186/s40064-016-3232-z
Misawa, N., Nakagawa, M., Kobayashi, K., Yamano, S., Izawa, Y., Nakamura, K., et al. (1990). Elucidation of the Erwinia uredovora carotenoid biosynthetic pathway by functional analysis of gene products expressed in Escherichia coli. J. Bacteriol. 172, 6704–6712. doi: 10.1128/jb.172.12.6704-6712.1990
Mitrophanov, A. Y., and Groisman, E. A. (2008). Signal integration in bacterial two-component regulatory systems. Genes Dev. 22, 2601–2611. doi: 10.1101/gad.1700308
Mohammadi, M., Burbank, L., and Roper, M. C. (2012). Biological role of pigment production for the bacterial phytopathogen Pantoea stewartii subsp. stewartii. Appl. Environ. Microbiol. 78, 6859–6865. doi: 10.1128/AEM.01574-12
Murashige, T., and Skoog, F. (1962). A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant. 15, 473–497. doi: 10.1111/j.1399-3054.1962.tb08052.x
Natale, P., Bruser, T., and Driessen, A. J. (2008). Sec- and Tat-mediated protein secretion across the bacterial cytoplasmic membrane–distinct translocases and mechanisms. Biochim. Biophys. Acta 1778, 1735–1756. doi: 10.1016/j.bbamem.2007.07.015
Naveed, M., Ahmed, I., Khalid, N., and Mumtaz, A. S. (2014). Bioinformatics based structural characterization of glucose dehydrogenase (gdh) gene and growth promoting activity of Leclercia sp. QAU-66. Braz. J. Microbiol. 45, 603–611. doi: 10.1590/S1517-83822014000200031
Niazi, A., Manzoor, S., Asari, S., Bejai, S., Meijer, J., and Bongcam-Rudloff, E. (2014). Genome analysis of Bacillus amyloliquefaciens subsp. plantarum UCMB5113: a rhizobacterium that improves plant growth and stress management. PLOS ONE 9:e104651. doi: 10.1371/journal.pone.0104651
Oulebsir-Mohandkaci, H., Khemili-Talbi, S., Benzina, F., and Halouane, F. (2015). Isolation and identification of entomopathogenic bacteria form Algerian desert soil and their effects against the migratory locust, Locusta migratoria (Linnaeus, 1758) (Orthoptera: Acrididae). Egypt. J. Biol. Pest Control. 25, 739–746.
Pandey, D. P., and Gerdes, K. (2005). Toxin-antitoxin loci are highly abundant in free-living but lost from host-associated prokaryotes. Nucleic Acids Res. 33, 966–976. doi: 10.1093/nar/gki201
Park, M. S., Jung, S. R., Lee, M. S., Kim, K. O., Do, J. O., Lee, K. H., et al. (2005). Isolation and characterization of bacteria associated with two sand dune plant species, Calystegia soldanella and Elymus mollis. J. Microbiol. 43, 219–227.
Paul, M., Primavesi, L. F., Jhurreea, D., and Zhang, Y. (2008). Trehalose metabolism and signaling. Annu. Rev. Plant Biol. 59, 417–441. doi: 10.1146/annurev.arplant.59.032607.092945
Peng, G., Zhang, W., Luo, H., Xie, H., Lai, W., and Tan, Z. (2009). Enterobacter oryzae sp. nov., a nitrogen-fixing bacterium isolated from the wild rice species Oryza latifolia. Int. J. Syst. Evol. Microbiol. 59, 1650–1655. doi: 10.1099/ijs.0.65484-0
Rademaker, J. L. W., Louws, F. J., and de Brujin, F. J. (1998). “Characterization of the diversity of ecologically important microbes by rep-PCR genomic fingerprinting,” in Molecular Microbial Ecology Manual, eds A. D. L. Akkermans, J. D. van Elsas, and F. J. de Brujin (Dordrecht: Kluwer Academic Publishers), 1–26.
Rättö, M., Verhoef, R., Suihko, M. L., Blanco, A., Schols, H. A., Voragen, A. G., et al. (2006). Colanic acid is an exopolysaccharide common to many enterobacteria isolated from paper-machine slimes. J. Ind. Microbiol. Biotechnol. 33, 359–367. doi: 10.1007/s10295-005-0064-1
Ren, W., Wang, Z. F., and Xu, A. K. (2010). Advancement in the research of salt-alkali tolerance genes in alkaligrass. Acta Pratacult. Sin. 19, 260–266.
Rezzonico, F., Smits, T. H. M., and Duffy, B. (2012). Detection of AI-2 receptors in genomes of Enterobacteriaceae suggests a role of type-2 quorum sensing in closed ecosystems. Sensors 12, 6645–6665. doi: 10.3390/s120506645
Rho, M., Tang, H., and Ye, Y. (2010). FragGeneScan: predicting genes in short and error-prone reads. Nuceic. Acids Res. 38:e191. doi: 10.1093/nar/gkq747
Robinson, C., and Bolhuis, A. (2004). Tat-dependent protein targeting in prokaryotes and chloroplasts. Biochim. Biophys. Acta 1694, 135–147. doi: 10.1016/j.bbamcr.2004.03.010
Rodrigues, A. A., Forzani, M. V., Soares, R. S., Sibov, S. T., and Vieira, J. D. G. (2016). Isolation and selection of plant growth-promoting bacteria associated with sugarcane. Pesqui. Agropecu. Trop. 46, 149–158. doi: 10.1590/1983-40632016v4639526
Rodríguez, H., Fraga, R., Gonzalez, T., and Bashan, Y. (2007). “Genetics of phosphate solubilization and its potential applications for improving plant growth-promoting bacteria,” in First International Meeting on Microbial Phosphate Solubilization. Developments in Plant and Soil Sciences, Vol. 102, eds E. Velázquez and C. Rodríguez-Barrueco (Dordrecht: Springer), 15–21. doi: 10.1007/978-1-4020-5765-6_2
Römling, U. (2007). “Cellulose biosynthesis in Enterobacteriaceae,” in Cellulose: Molecular and Structural Biology, eds M. Brown Jr. and I. M. Saxena (Dordrecht: Springer), 107–122. doi: 10.1007/978-1-4020-5380-1_7
Römling, U., and Galperin, M. Y. (2015). Bacterial cellulose biosynthesis: diversity of operons, subunits, products, and functions. Trends Microbiol. 23, 545–557. doi: 10.1016/j.tim.2015.05.005
Rothe, M., Alpert, C., Loh, G., and Blaut, M. (2013). Novel insights into E. coli’s hexuronate metabolism: kduI facilitates the conversion of galacturonate and glucuronate under osmotic stress conditions. PLOS ONE 8:e56906. doi: 10.1371/journal.pone.0056906
Russell, A. B., Hood, R. D., Bui, N. K., LeRoux, M., Vollmer, W., and Mougous, J. D. (2011). Type VI secretion delivers bacteriolytic effectors to target cells. Nature 475, 343–347. doi: 10.1038/nature10244
Russell, A. B., Peterson, S. B., and Mougous, J. D. (2014). Type VI secretion system effectors: poisons with a purpose. Nat. Rev. Microbiol. 12, 137–148. doi: 10.1038/nrmicro3185
Ryu, C. M., Farag, M. A., Hu, C. H., Reddy, M. S., Wei, H. X., Paré, P. W., et al. (2003). Bacterial volatiles promote growth in Arabidopsis. Proc. Natl. Acad. Sci. U.S.A. 100, 4927–4932. doi: 10.1073/pnas.0730845100
Saitou, N., and Nei, M. (1987). The neighbor-joining method: a new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 4, 406–425. doi: 10.1093/oxfordjournals.molbev.a040454
Sedkova, N., Tao, L., Rouvière, P. E., and Cheng, Q. (2005). Diversity of carotenoid synthesis gene clusters from environmental Enterobacteriaceae strains. Appl. Environ. Microbiol. 71, 8141–8146. doi: 10.1128/AEM.71.12.8141-8146.2005
Shah, S., Li, J., Moffatt, B. A., and Glick, B. R. (1998). Isolation and characterization of ACC deaminase genes from two different plant growth-promoting rhizobacteria. Can. J. Microbiol. 44, 833–843. doi: 10.1139/w98-074
Shankar, M., Ponraj, P., Ilakkiam, D., and Gunasekaran, P. (2011). Root colonization of a rice growth promoting strain of Enterobacter cloacae. J. Basic Microbiol. 51, 523–530. doi: 10.1002/jobm.201000342
Shao, Y., Harrison, E. M., Bi, D., Tai, C., He, X., Ou, H. Y., et al. (2011). TADB: a web-based resource for Type 2 toxin-antitoxin loci in bacteria and archaea. Nucleic Acids Res. 39, D606–D611. doi: 10.1093/nar/gkq908
Shen, X., Hu, H., Peng, H., Wang, W., and Zhang, X. (2013). Comparative genomic analysis of four representative plant growth-promoting rhizobacteria in Pseudomonas. BMC Genomics 14:271. doi: 10.1186/1471-2164-14-271
Shrestha, P. M., Noll, M., and Liesack, W. (2007). Phylogenetic identity, growth-response time and rRNA operon copy number of soil bacteria indicate different stages of community succession. Environ. Microbiol. 9, 2464–2474. doi: 10.1111/j.1462-2920.2007.01364.x
Shyntum, D. Y., Venter, S. N., Moleleki, L. N., Toth, I., and Coutinho, T. A. (2014). Comparative genomics of type VI secretion systems in strains of Pantoea ananatis from different environments. BMC Genomics 15:163. doi: 10.1186/1471-2164-15-163
Singh, R. P., Nalwaya, S., and Jha, P. N. (2015). The plant-growth promoting bacterium Klebsiella sp. SBP-8 confers induced systemic tolerance in wheat (Triticum aestivum) under salt stress. J. Plant. Physiol. 184, 57–67. doi: 10.1016/j.jplph.2015.07.002
Singh, R. P., Nalwaya, S., and Jha, P. N. (2017). The draft genome sequence of the plant growth promoting rhizospheric bacterium Enterobacter cloacae SBP-8. Genomics Data 12, 81–83. doi: 10.1016/j.gdata.2017.03.006
Solheim, M., La Rosa, S. L., Mathisen, T., Snipen, L. G., Nes, I. F., and Brede, D. A. (2014). Transcriptomic and functional analysis of NaCl-induced stress in Enterococcus faecalis. PLOS ONE 9:e94571. doi: 10.1371/journal.pone.0094571
Taghavi, S., Garafola, C., Monchy, S., Newman, L., Hoffman, A., Weyens, N., et al. (2009). Genome survey and characterization of endophytic bacteria exhibiting a beneficial effect on growth and development of poplar trees. Appl. Environ. Microbiol. 75, 748–757. doi: 10.1128/AEM.02239-08
Taghavi, S., van der Lelie, D., Hoffman, A., Zhang, Y. B., Walla, M. D., Vangronsveld, J., et al. (2010). Genome sequence of the plant growth promoting endophytic bacterium Enterobacter sp. 638. PLOS Genet. 6:e1000943. doi: 10.1371/journal.pgen.1000943
Tamura, K., Stecher, G., Peterson, D., Filipski, A., and Kumar, S. (2013). MEGA6: Molecular Evolutionary Genetics Analysis version 6.0. Mol. Biol. Evol. 30, 2725–2729. doi: 10.1093/molbev/mst197
Thompson, J. D., Gibson, T. J., and Higgins, D. G. (2002). Multiple sequence alignment using ClustalW and ClustalX. Curr. Protoc. Bioinformatics 2, 2.3.1–2.3.22. doi: 10.1002/0471250953.bi0203s00
Timmusk, S., Behers, L., Muthoni, J., Muraya, A., and Aronsson, A. C. (2017). Perspectives and challenges of microbial application for crop improvement. Front. Plant. Sci. 8:49. doi: 10.3389/fpls.2017.00049
Trujillo, M. E., Bacigalupe, R., Pujic, P., Igarashi, Y., Benito, P., Riesco, R., et al. (2014). Genome features of the endophytic actinobacterium Micromonospora lupini strain Lupac 08: on the process of adaptation to an endophytic life style? PLOS ONE 9:e108522. doi: 10.1371/journal.pone.0108522
Valmeekam, V., Loh, Y. L., and San Francisco, M. J. (2001). Control of exuT activity for galacturonate transport by the negative regulator ExuR in Erwinia chrysanthemi EC16. Mol. Plant Microbe Interact. 14, 816–820. doi: 10.1094/MPMI.2001.14.6.816
Vurukonda, S. S., Vardharajula, S., Shrivastava, M., and SkZ, A. (2016). Enhancement of drought stress tolerance in crops by plant growth promoting rhizobacteria. Microbiol. Res. 184, 13–24. doi: 10.1016/j.micres.2015.12.003
Walderhaug, M. O., Litwack, D. E., and Epstein, W. (1989). Wide distribution of homologs of Escherichia coli Kdp K+-ATPase among gram-negative bacteria. J. Bacteriol. 171, 1192–1195. doi: 10.1128/jb.171.2.1192-1195.1989
Weber, T., Blin, K., Duddela, S., Krug, D., Kim, H. U., Bruccoleri, R., et al. (2015). antiSMASH 3.0-a comprehensive resource for the genome mining of biosynthetic gene clusters. Nucleic Acids Res. 43, W237–W243. doi: 10.1093/nar/gkv437
Woo, H. L., Ballor, N. R., Hazen, T. C., Fortney, J. L., Simmons, B., Davenport, K. W., et al. (2014). Complete genome sequence of the lignin-degrading bacterium Klebsiella sp. strain BRL6-2. Stand. Genomic Sci. 9:19. doi: 10.1186/1944-3277-9-19
Worley, M. J., Chingm, K. H., and Heffron, F. (2000). Salmonella SsrB activates a global regulon of horizontally acquired genes. Mol. Microbiol. 36, 749–761. doi: 10.1046/j.1365-2958.2000.01902.x
Wu, S., Zhu, Z., Fu, L., Niu, B., and Li, W. (2011). WebMGA: a customizable web server for fast metagenomic sequence analysis. BMC Genomics 12:444. doi: 10.1186/1471-2164-12-444
Xie, S. S., Wu, H. J., Zang, H. Y., Wu, L. M., Zhu, Q. Q., and Gao, X. W. (2014). Plant growth promotion by spermidine-producing Bacillus subtilis OKB105. Mol. Plant Microbe Interact. 27, 655–663. doi: 10.1094/MPMI-01-14-0010-R
Yaish, M. W. (2016). Draft Genome sequence of endophytic bacterium Enterobacter asburiae PDA134, isolated from date palm (Phoenix dactylifera L.) roots. Genome Announc. 4:e00848–16. doi: 10.1128/genomeA.00848-16
Yaish, M. W., Antony, I., and Glick, B. R. (2015). Isolation and characterization of endophytic plant growth-promoting bacteria from date palm tree (Phoenix dactylifera L.) and their potential role in salinity tolerance. Antonie Van Leeuwenhoek 107, 1519–1532. doi: 10.1007/s10482-015-0445-z
Zeidler, D., Zähringer, U., Gerber, I., Dubery, I., Hartung, T., Bors, W., et al. (2004). Innate immunity in Arabidopsis thaliana: lipopolysaccharides activate nitric oxide synthase (NOS) and induce defense genes. Proc. Natl. Acad. Sci. U.S.A. 101, 15811–15816. doi: 10.1073/pnas.0404536101
Zhang, W., Hu, X., Wang, L., and Wang, X. (2014). Reconstruction of the carotenoid biosynthetic pathway of Cronobacter sakazakii BAA894 in Escherichia coli. PLOS ONE 9:e86739. doi: 10.1371/journal.pone.0086739
Keywords: Indigofera argentea, root endophytes, plant–microbe interaction, plant growth-promoting bacteria (PGPB), salt stress
Citation: Andrés-Barrao C, Lafi FF, Alam I, de Zélicourt A, Eida AA, Bokhari A, Alzubaidy H, Bajic VB, Hirt H and Saad MM (2017) Complete Genome Sequence Analysis of Enterobacter sp. SA187, a Plant Multi-Stress Tolerance Promoting Endophytic Bacterium. Front. Microbiol. 8:2023. doi: 10.3389/fmicb.2017.02023
Received: 05 June 2017; Accepted: 04 October 2017;
Published: 20 October 2017.
Edited by:
Devin Coleman-Derr, University of California, Berkeley, United StatesReviewed by:
Oswaldo Valdes-Lopez, National Autonomous University of Mexico, MexicoJavier Alejandro Izquierdo, Hofstra University, United States
Copyright © 2017 Andrés-Barrao, Lafi, Alam, de Zélicourt, Eida, Bokhari, Alzubaidy, Bajic, Hirt and Saad. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Heribert Hirt, heribert.hirt@kaust.edu.sa
†Present address: Feras F. Lafi, Department of Medical Laboratories, Faculty of Science, American University of Madaba, Madaba, Jordan