Neuroscience: Multitasking on the run
- The Rockefeller University, United States
- Duke University School of Medicine, United States
The learning abilities of the cerebellum, also known as the ‘little cortex’, enable us to learn and execute complex motor movements without even thinking about them. Thus, our abilities to play a musical instrument, for example, or even just to reach for a cup of coffee without looking up from our computer, all depend on the cerebellum. One of the hallmarks of the cerebellum, which is hidden under the much larger cerebral cortex, is its simple, stereotyped architecture and well-understood circuitry. This simplicity means that analysis of the structure and function of the cerebellum offers neuroscientists the opportunity to understand the basic principles of cortical function.
For many years, it has been known that the cerebellum relies on two fundamental principles of operation. First, it is able to compare sensory signals (which arise in sensory receptors and report what has happened in the outside world) and motor signals (which are corollaries of the motor commands sent to the spinal cord). Second, ‘plasticity’ in the cerebellum mediates the learning of motor skills. Here, plasticity refers to changes in the strength of the synapses that allow one neuron to communicate with another, or to changes in the number of action potentials emitted by a neuron when it receives a given set of synaptic inputs.
Writing in eLife, Adam Hantman and colleagues—including Cheng-Chiu Huang as first author—provide direct evidence that link these two principles. They show that sensory and motor inputs to the cerebellum converge at the earliest stage of processing, on the cerebellar granule neuron (Huang et al., 2013). Their findings have important implications for how the cerebellum does its job, including how and where synaptic plasticity causes learning of motor skills.
The cerebellar cortex consists of three layers—an innermost layer of granule neurons, a central layer of Purkinje cells, and an outer molecular layer—and it receives two major sources of input: mossy fibers and climbing fibers. Mossy fibers originate in multiple regions, relaying motor and sensory inputs to synapses on the dendrites of granule neurons (Palay and Chan-Palay, 1974). The axons of the granule neurons ascend through the Purkinje neuron layer into the molecular layer, where they bifurcate into ‘parallel fibers’ and form synapses with the dendrites of Purkinje neurons (Figure 1). Tens, or maybe hundreds, of thousands of parallel fibers can form synapses with a single Purkinje cell.
Figure 1
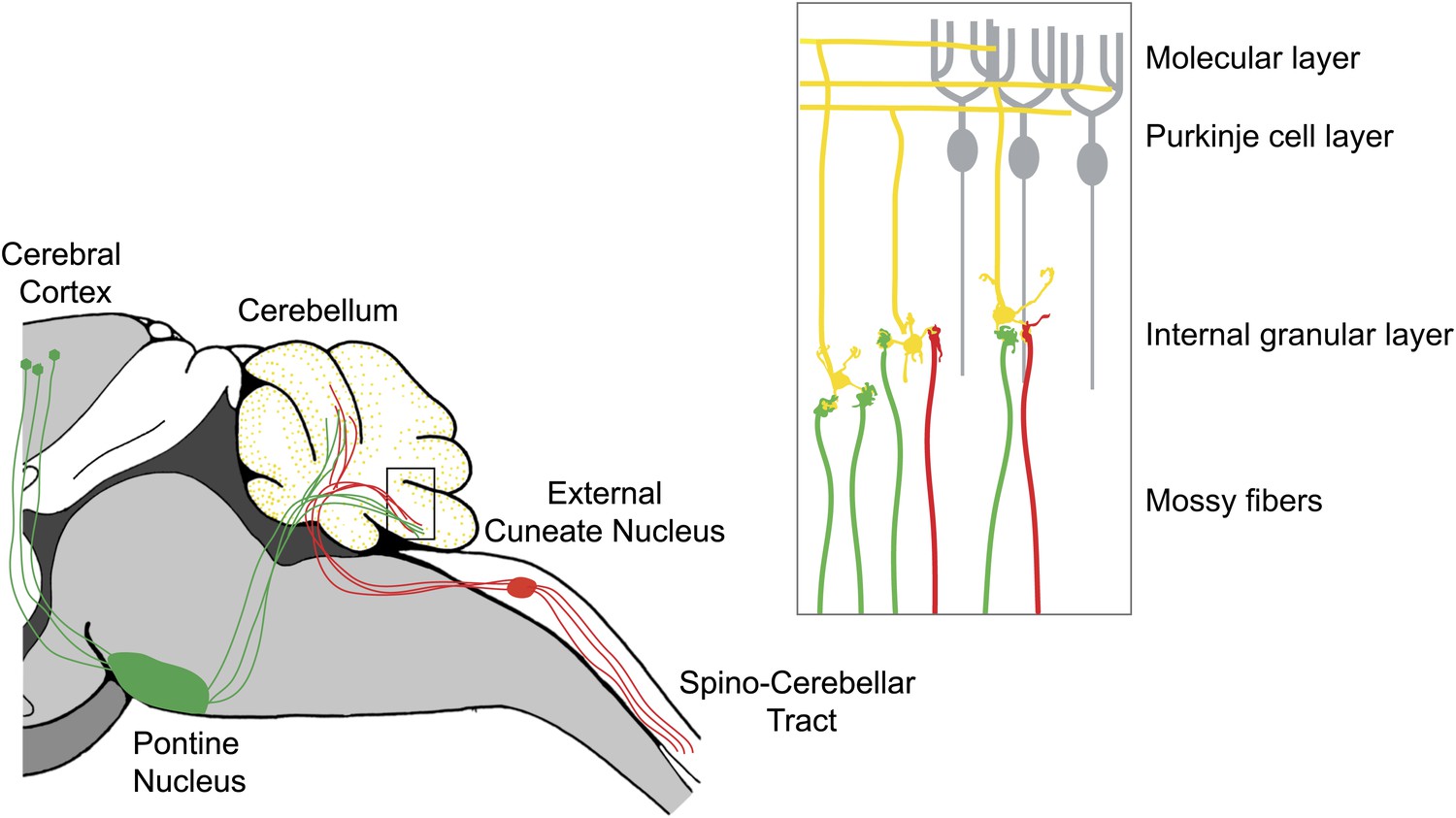
The cerebellum integrates motor and sensory signals, which allows us to learn and perfect complex motor skills.
The signals are relayed to the cerebellum by mossy fibers: copies of motor commands sent by the cerebral cortex are relayed via the pontine nucleus (green), while sensory signals are relayed from the spinal cord via the external cuneate nucleus (red). Inset: Expanded view of the cerebellum showing mossy fibers (red and green) that have formed synapses with the dendrites of granule neurons (yellow). More than 40% of the granule cells studied by Huang et al. received inputs from both sensory (red) and motor (green) mossy fibers. The axons of the granule neurons (yellow) ascend into the molecular layer of the cerebellum, where they bifurcate to form parallel fibers: large numbers of parallel fibers can form synapses with a single Purkinje cell (gray). The climbing fibers are not shown.
FIGURE: MARY E HATTEN/JIM DUFFY
Climbing fibers, on the other hand, originate from the inferior olivary nucleus in the brainstem. Moreover, each Purkinje cell receives many synaptic contacts from only a single climbing fiber. This unusual anatomical feature has driven much of the research into the function of climbing fibers.
Classical models of cerebellar function—first proposed independently by David Marr in 1969, James Albus in 1971 and Masao Ito in 1972 (Marr, 1969; Albus, 1971; Ito, 1972)—hold that the electrical activity of mossy fibers acts on a millisecond-by-millisecond basis to create a pattern of Purkinje cell action potentials that guides accurate movement, while climbing fibers produce action potentials on a longer time scale (about once per second) to guide plasticity in the cerebellum. Plasticity adjusts the strength of each parallel fiber input and attempts to create the correct commands for accurate, skilled movement. Indeed, when inputs from parallel fibers and climbing fibers arrive at a given Purkinje neuron at the same time, the strength of the synapse between the parallel fibers and the Purkinje cell is reduced (Ito et al., 1982; Linden et al., 1991). This reduction in the strength of the synapse is called ‘long-term depression'.
Huang, Hantman and colleagues—who are based at the Janelia Farm Research Campus of the Howard Hughes Medical Institute and at Brandeis University—now reveal a previously unknown feature of cerebellar organization that speaks loudly to the possible mechanisms by which the cerebellum can ‘learn' new motor skills. Through deployment of a tour de force combination of neuroanatomical, genetic and imaging tools, they show that mossy fibers from sensory and motor relay centers converge onto individual granule neurons. Moreover, they demonstrate that this multimodal convergence occurs across the cerebellum.
Huang et al. injected different reporter virus constructs into two mouse brainstem nuclei that are sources of mossy fiber input to the cerebellum. The construct tagged with a red reporter gene was injected into the external cuneate nucleus, which belongs to the pathway that carries proprioceptive sensory information (that is, information about the position of our limbs in space), and the construct tagged with a green reporter gene was injected into the pontine nucleus, which is part of the pathway that carries copies of motor commands from the cortex to the cerebellum. Confocal imaging of the fluorescently tagged mossy fiber projections revealed considerable convergence of the motor and sensory pathways within the cerebellar granule cell layer.
To determine whether the pathways converged onto individual granule cells, Huang et al. used a second transgenic mouse line, the TCGO line, which expresses a yellow reporter in a random subset of granule cells. Using high-resolution confocal imaging, they were able to home in on the sparsely labeled granule cells, and to visualize individual mossy fibers terminating on individual granule neuron dendrites (yellow). Remarkably, up to 40% of the granule cells that they analyzed received inputs from both sensory (green) and motor (red) fibers. This is the first evidence for the convergence of sensory and motor afferents at such an early stage of processing.
The most important implication from the paper of Huang et al. is that sensory–motor integration might occur where the mossy fibers meet the granule neurons, and not where the parallel fibers form synapses with the Purkinje cells in the molecular layer. This raises questions about our current models of cerebellar learning. If learning occurs through changes in the strength of the parallel fiber/Purkinje cell synapse, then the early convergence of motor and sensory signals on granule cells means that the cerebellum may have limited ability to adjust the strengths of sensory and motor signals independently. Or, there might be a critical role for plasticity mechanisms at other sites in the cerebellar circuit (Hansel et al., 2001; Carey and Lisberger, 2002), even at the synapses between mossy fibers and granule cells.
The findings provide elegant support for the unifying model of cerebellar function proposed more than 40 years ago by Marr, Albus and Ito. But while the truth may follow the fundamental principles these pioneering authors espoused, it may be more complex than they imagined. Nevertheless, the application of modern biological technology to known neural circuits, as pioneered by Hantman and co-workers, will boost our understanding of how behavior and learning are organized at the circuitry level.
References
-
Climbing fibre induced depression of both mossy fibre responsiveness and glutamate sensitivity of cerebellar Purkinje cellsJ Physiol 324:113–134.
Article and author information
Author details
Publication history
- Version of Record published: March 19, 2013 (version 1)
Copyright
© 2013, Hatten and Lisberger
This article is distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use and redistribution provided that the original author and source are credited.
Metrics
-
- 797
- views
-
- 72
- downloads
-
- 3
- citations
Views, downloads and citations are aggregated across all versions of this paper published by eLife.
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
Neuroscience: Multitasking on the run
eLife 2:e00641.
https://doi.org/10.7554/eLife.00641
Further reading
-
- Neuroscience
In the ‘double-drift’ illusion, local motion within a window moving in the periphery of the visual field alters the window’s perceived path. The illusion is strong even when the eyes track a target whose motion matches the window so that the stimulus remains stable on the retina. This implies that the illusion involves the integration of retinal signals with non-retinal eye-movement signals. To identify where in the brain this integration occurs, we measured BOLD fMRI responses in visual cortex while subjects experienced the double-drift illusion. We then used a combination of univariate and multivariate decoding analyses to identify (1) which brain areas were sensitive to the illusion and (2) whether these brain areas contained information about the illusory stimulus trajectory. We identified a number of cortical areas that responded more strongly during the illusion than a control condition that was matched for low-level stimulus properties. Only in area hMT+ was it possible to decode the illusory trajectory. We additionally performed a number of important controls that rule out possible low-level confounds. Concurrent eye tracking confirmed that subjects accurately tracked the moving target; we were unable to decode the illusion trajectory using eye position measurements recorded during fMRI scanning, ruling out explanations based on differences in oculomotor behavior. Our results provide evidence for a perceptual representation in human visual cortex that incorporates extraretinal information.
-
- Neuroscience
The flow of neural activity across the neocortex during active sensory discrimination is constrained by task-specific cognitive demands, movements, and internal states. During behavior, the brain appears to sample from a broad repertoire of activation motifs. Understanding how these patterns of local and global activity are selected in relation to both spontaneous and task-dependent behavior requires in-depth study of densely sampled activity at single neuron resolution across large regions of cortex. In a significant advance toward this goal, we developed procedures to record mesoscale 2-photon Ca2+ imaging data from two novel in vivo preparations that, between them, allow for simultaneous access to nearly all 0f the mouse dorsal and lateral neocortex. As a proof of principle, we aligned neural activity with both behavioral primitives and high-level motifs to reveal the existence of large populations of neurons that coordinated their activity across cortical areas with spontaneous changes in movement and/or arousal. The methods we detail here facilitate the identification and exploration of widespread, spatially heterogeneous neural ensembles whose activity is related to diverse aspects of behavior.




{kind=link}