









-
The MYB transcription factor (TF) is one of the earliest discovered functional factors; the first MYB transcription factor identified in plants was ZmMYBC1 cloned from maize in 1987[1], and its main function was found to be involved in the synthesis of anthocyanins. Since the discovery of the MYB transcription factor in plants, it has been demonstrated that these factors play a significant role in the development and maintenance of plants[2,3].
The functions of the MYB transcription factors are mainly due to their diversity of structures. The N-terminal region of the family contains a highly conserved domain, which is composed of 50 to 53 amino acids. This region is responsible for forming a three-dimensional HTH structure by linking the second and third helices to each other. The main role of the tryptophan (W) residues in this process is to form a hydrophobic core. Due to the number of repeats and repeat prototypes, they are divided into four subclasses: 1R-MYB (R3 and R1/2), R2R3-MYB(2R-MYB), 3R-MYB(R1, R2, R3) and 4R-MYB(R1, R2, R2, R1/2)[4−6]. Among them, the R1MYB protein contains a unique MYB-binding domain spanning 53 amino acid residues, including three isometric tryptophans (W). Substituting an alanine (A) residue for a third tryptophan (W) within the MYB domain can form a SHAQKYF motif that recognizes the TATC/GATA core[7]. The C-terminal motif's composition determines the function of MYB transcription factors, and contains one or two distinct motifs, such as the EAR motif (ERF - associated amphipathic repression)[8,9], the SID motif (sensitive to ABA and drought 2 protein interaction motifs[10], and the TLLLFR motif sequence[11]. Structural genes that[6] bind to MYB transcription factors typically contain two MYB transcription factor recognition elements, MRE (C/T)G (A/G)(A/C/G)GTT(A/G) and ACE (ACGT - containing element)[12]. These structural genes provide more possibilities for the diversity in the functions of MYB transcription factors (Fig. 1).
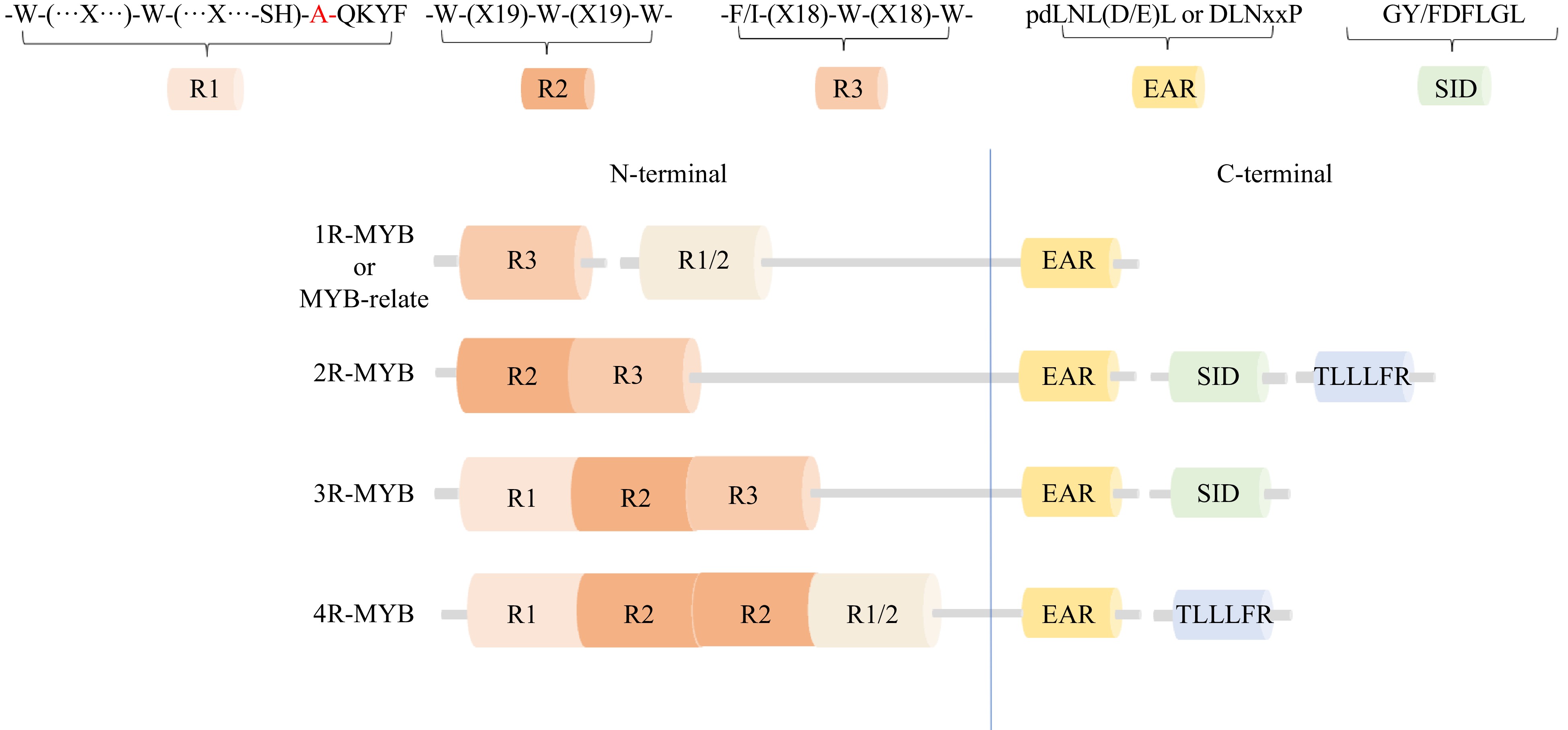
Figure 1.
Classification and structural characteristics of plant MYB transcription factors.
Functional characteristics of MYB subfamily members
-
The MYB family includes the R2R3-MYB, 1R-MYB, 3-MYB, and 4-MYB subfamilies. Existing research has shown that R2R3-MYB is the most abundant subgroup of the MYB transcription factor family, which is widely involved in environmental stress, hormone response, secondary metabolism, and cell differentiation processes. Examples of this subgroup include the R2R3-MYB transcription factor in rice[13], Arabidopsis, bamboo[14], Populus, etc.[15]. With increasing attention being paid to fruit crops, large quantities of functional studies on R2R3-MYB have been reported on fruit such as grape[16], apple[17,18], and pear[19−21], the R2R3-MYB are concerned with the biosynthesis of nutritional tissues, pericarp anthocyanins, flavonoids and lignans. Kiwifruit R2R3-MYB regulates chlorophyll and carotenoid accumulation[22], strawberry R2R3-MYB is concerned with controlling the synthesis of strawberry eugenol[23] and sweet cherry R2R3-MYB improves salt stress tolerance and infective agent resistance[24].
In addition to R2R3-MYB, functional studies of other subfamilies have also been reported. The 3R-MYB subfamily is predominantly involved in the cell cycle, cell differentiation, and plant stress. For example, MnMYB3R1 in mulberry is participating in regulating the polyphenol oxidase genes expression[25]. 3R-MYB in kiwifruit and rice enhance drought and salt tolerance in Arabidopsis. Additionally[26,27], 3R-MYB is involved in the synthesis of plant anthocyanins. e.g. BrMYBL2.1 negatively regulates anthocyanin biosynthesis in Chinese cabbage by inhibiting the MYB–bHLH–WD40 complex activity[28]. The 1R-MYB subfamily is of concern in the regulation of plant seed germination and resistance to stress, such as 1R-MYB transcription factors in barley and chickpea[7,29]. On the other hand, 4R-MYB is currently less researched, and its specific function is still unclear. Only member classifications have been reported, as the two 4R-MYB members in Arabidopsis and one in rice[30].
MYB transcription factor's diverse biological function mechanism
-
The regulation of MYB transcription factor expression is mainly manifested in transcription and translation. At the transcriptional level, the MYB transcription factor is mainly regulated by two small RNAs, miRNA[31] and TA-siRNA[32,33], after transcription. At the translational level, the MYB transcription factor is mainly regulated through histone acetylation, methylation, ubiquitin-mediated protein degradation, SUMO modification[34−36] and deacetylation[37]. The study of the function mechanism of the MYB transcription factor has become more advanced in recent years, and there are three common approaches: one is to cooperate with other transcription factors and then co-regulate them with structural genes; the other is to directly combine them with downstream target genes to exert regulatory effects; and the third is to regulate other transcription factors.
The MYB transcription factor binds to WD40 (WD40 repeat proteins) and the bHLH (basic helix–loop–helix) to regulate plant secondary metabolism, including the biosynthesis of flavonoids and glucosinolates, as well as some basic plant tissues, such as the formation of unicellular trichomes and the differentiation of non-hairy root epidermal cells and stomata[38]. Arabidopsis thaliana seed coat development is regulated by the MYB-bHLH-WDR transcription factor complex (MBW)[39]. Liu et al.[40] found that the ternary complex formed by CsWD40 (Camellia sinensis) and MYB, BHLH, etc., can regulate the biosynthesis of anthocyanins and procyanidins as well as the production by trichomes. JAZs (Jasmonate ZIM-domain) inhibit the bHLH–MYB complex to regulate JA-mediated stamen development[41]. At the same time, MYB transcription factors are also found to form a NAC–MYB-based gene regulatory network with NAC transcription factors or to form a BHLH–MYB–AP2 ternary complex with BHLH and AP2 to jointly regulate the synthesis of plant lignin[42]. The loquat ternary complex EjbHLH1–EjMYB2–EjAP2-1 delays the process of low temperature-induced loquat fruit lignification[43].
MYB transcription factors bind with specific binding elements in the promoter regions of many functional genes that activate or inhibit the expression of stress-responsive genes, and regulate the silencing interference and overexpression of downstream-related functional genes such as those related to disease resistance and stress resistance, so as to mitigate the consequences of disease and environmental stress or threats. Based on yeast one-hybrid screening, Zhang et al.[44] found that the Medicago truncatula R2R3-MYB transcription factor, MtMYB3, directly binds to the MYB cis-factor in the MtCBF4 promoter, thereby inhibiting the MtCBF4 promoter. At the same time, the MtMYB61 transcription factor and MtMYB3 antagonize each other to regulate MtCBF4, resulting in the upregulation of the downstream target gene MtCAS15 in the cold acclimation of Mycobacterium frondosa. Tobacco NtMYB305a binds to the Jasmonic acid-responsive GAG region in the NtPMT1a promoter to regulate nicotine biosynthesis[45].
MYB transcription factors can also regulate and be regulated by other transcription factors. OsMYB46 and ZmMYB46 promoters contain secondary wall NAC binding element (SNBEs) that can be bound and activated by rice OsSWN and maize ZmSWN[46]. Populus tomentosa PtrMYB161 can bind to multiple sets of target genes, acting as both an activator and a repressor, and it can simultaneously inhibit its upper-level regulatory transcription factor NAC Domain1 (PtrSND1) and its transcription factor family member. Moreover, the gene PtrMYB102, which is directly regulated by family members, can also directly transactivate five wood-forming genes[47]. Glutathione can regulate the expression of heat shock proteins by activating the promoters of HSP70B and BiP3 through MYB21[48].
-
There are currently 63 species of tropical fruit worldwide (
www.tasteatlas.com/tropical-fruits ). Pineapple, mango and banana are the major tropical fruit. Aside from lychee, kiwifruit, guava, and jackfruit, minor tropical fruit also include lesser-known fruits such as mangosteen, tree tomato, lutqua, carambola, passion fruit, elephant apple, water apple, rambutan, persimmon, indian gooseberry, bayberry, sapodilla, longan, pepino, bhawa, loquat, silverberry, annona, durian, jaboticaba, pulsan, and pitaya[49,50] (Supplemental Table S1) . The high quantity of flavonoids, anthocyanins and dietary fiber in tropical fruit has made them increasingly popular in recent years, while flavonoids (proanthocyanidins, anthocyanidins, flavonols,etc.) are also involved in the regulation of flower and fruit color and the enhancement of plant stress resistance[51,52]. MYB is the primary transcription factor that activates or inhibits these pathways. However, research on the MYB gene in tropical fruit is generally in the family classification stage, and research on its function is still in its infancy. Some of the subfamilies of fruit have been classified and analyzed. For example, based on mango leaf and fruit transcriptome analysis, there are 125 MYB family transcription factors were identified, including 1 4R-MYB, 1 3R-MYBs, 51 R2R3-MYBs, and 8 and 64 R1-MYBs and atypical MYBs, respectively[53]. A total of 233 MYBs were screened based on pineapple (Ananas comosus) genome information, including 126 1R-MYBs, 103 2R-MYBs, 3 3R-MYBs and 1 4R-MYB[54]. A genome-wide analysis of Chinese bayberry (Morella rubra) identified 174 MrMYB transcription factors, including 43 1R-MYBs, 122 R2R3-MYBs, two 3R-MYBs, one 4R-MYB, and six atypical[55] MYBs. A total of 293 MaMYBs, including 285 R2R3-MYB proteins, six 3R-MYB proteins, and two 4R-MYB proteins, were obtained from whole-genome analysis of bananas[56]. Xie et al.[57] obtained 105 R2R3-MYBs based on the whole genome data analysis of pitaya. A total of 119 DlR2R3-MYBs were obtained by analyzing the whole genome data of longan (Dimocarpus longan Lour.)[58]. Jue et al.[59] previously found 60 MYBs involved in the longan regulation of floral transformation. There are 25 R2R3-MYB and nine 1R-MYB-like transcription factors among the 34 MYB proteins in papaya (Carica papaya) fruit ripening[60]. Only some of the family members of the MYB subfamily are introduced. A total of 56 MYBs in lychee (Litchi chinensis) may be the main transcription factors of high temperature-induced flower abortion based on hormone control[61]. Lai et al.[62] found that 53 R2R3-MYB genes were involved in chlorophyll degradation and flavonoids synthesis during lychee ripening. Wax apple has 20 MYB genes related to lignin synthesis[63]. A total of 4.6% of the 843 transcriptional family members found in early somatic embryos of mangosteen belong to the MYB gene family[64]. Transcriptomic comparative analysis of mangosteen (Garcinia mangostana) fruit with translucent pulp disorder under different water conditions revealed 152 1R-MYBs[65]. Using the lifemap website (
https://lifemap-ncbi.univ-lyon1.fr/# ) to perform phylogenetic tree analysis on the whole genomes of known R2R3 MYB transcription factor family members: banana, pineapple, mango, longan, lichi, papaya, pitaya and bayberry, the results are shown in Fig. 2. As more research work is reported, more MYB transcription factor family members of tropical fruit will be discovered, and the functions of these MYB transcription factor family members will be different. The results were analyzed to serve as a basis for a more thorough a theoretical rationale for the further in-depth study on the biofunction of the MYB in tropical fruit. The MYB genes cited in this paper and their main functions are summarized in Table 1.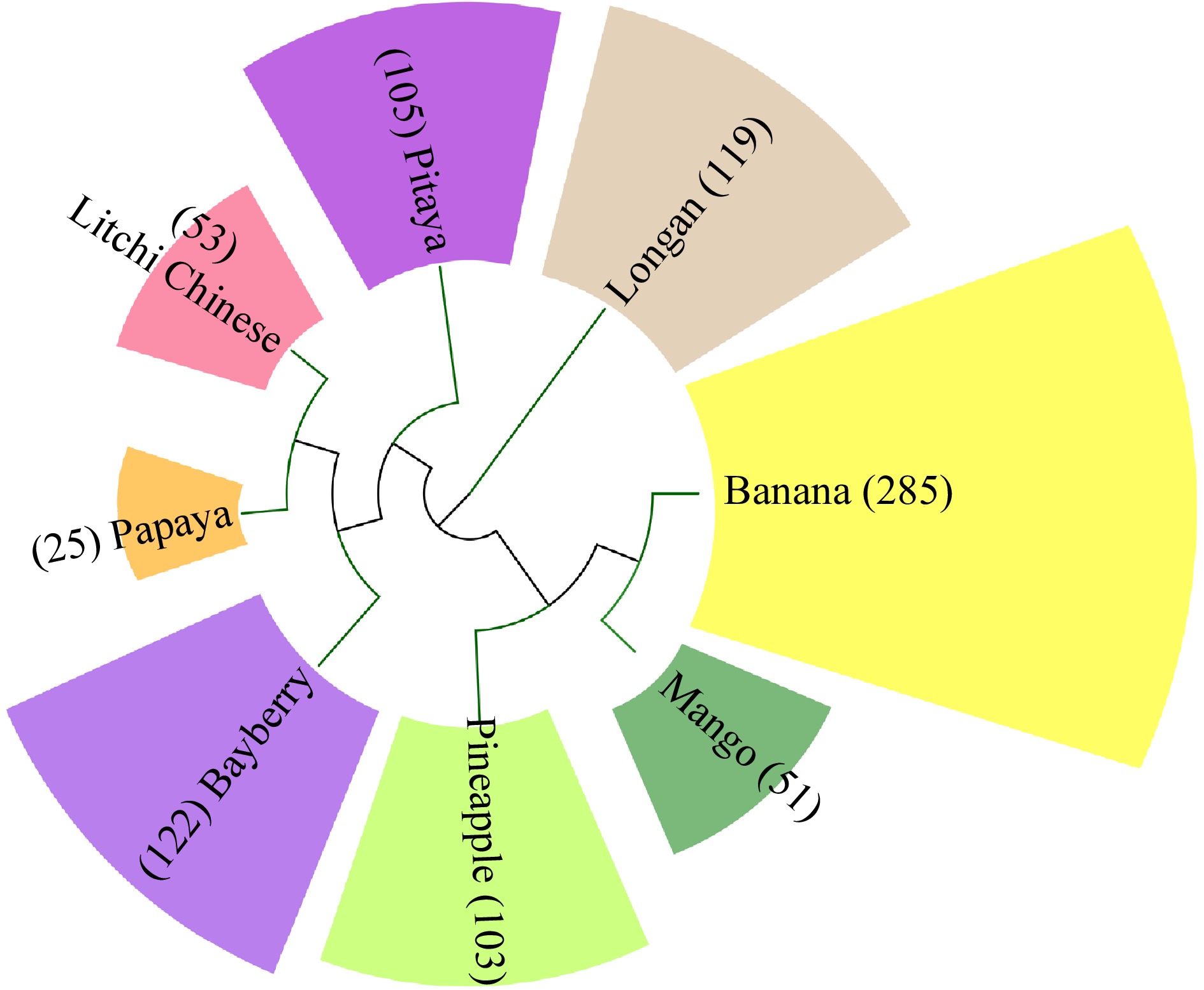
Figure 2.
Number of R2R3-MYB subfamily members in tropical fruit.
Table 1. MYB genes and their main functions in tropical fruit.
Latin name Plant name Gene Gene founction Reference Litchi chinensis Litch LcMYB1 (KY302802.1) Regulates the accumulation of anthocyanins [68−72] LcR1MYB1 (QCF47524.1) Negative regulation of anthocyanin biosynthesis [73] LcMYB5 (Unigene 0002103) Anthocyanin biosynthesis, tissue acidification [127] LcMYBx (UCU01159.1) Negatively correlated with the synthesis of leaf anthocyanins [74] Mangifera indica Mango MinMYB10 Possible involvement in the regulation of anthocyanin accumulation in fruits [91] MinMYBPA Possible involvement in the regulation of anthocyanin accumulation in peels [92] MiMYB1 (NC_056567.1) Peel anthocyanin synthesis [93] Musa acuminata Banana MaMYB1 Response to drought, low temperature, salt stress [101] MaMYB14/63/110 Response process of salt stress [105] MaMYBR1 Resistance to fusarium wilt. [103] MaMYB3/4/32 Fruit ripening [119] MaMYB4 (Ma03_g21920) Inhibits banana anthocyanin biosynthesis and fruit ripeing; Negatively regulates the synthesis of banana Regulates the anthocyanins and synthesis of fatty acids [75,106,118] MaMYB3 Delay banana fruit ripening [116] MusaMYB31 Inhibits banana lignin and polyphenol biosynthesis [107] MaMYB16L Inhibited fruit ripening [115] MaMYB16S Promotes ripening of banana fruit [115] Musa spp. MpMYBS3 (KM379134) Participate in the regulation of cold signaling pathways. [108] Musa paradisiaca MYB108 (Ma09_g15940) Regulates the growth and development of banana leaves and roots under long-term magnesium deficiency conditions [104] Ananas comosus Pineapple AcoMYB1 (XM_020230319) Regulates early development and late ripening in pineapples in response to drought and low temperature stress [109] AcoMYB4 Inhibit ABA synthesis [110] Hylocereus polyrhizus Pitaya HuMYB1814 Synthesis of Anthocyanins [83] Hylocereus undatus Unigene0036816 May be to relevant the color difference of dragon fruit pulp [84] Unigene0032395 May be to relevany the color difference of dragon fruit pulp [84] Unigene0034590 May be to relevant the color difference of dragon fruit pulp [84] Hylocereus polyrhizus HuMYB1 Inhibits the expression of β-galactose-related genes. [121] Garcinia mangostana Mangosteen GmMYB10 (ACM62751.1) Regulation of anthocyanin biosynthesis before and after fruit harvest [85] GmMYB30 (AID69234.1) Cell wall thickening, lignin
biosynthesis and stress response.
[128]Myrica rubra Bayberry MrMYB1 (GQ340767) Expression is induced by light signals, which then affects anthocyanin accumulation [76−80] MrMYB6 (c24596_g1) May negatively regulate the anthocyanins and PA (proanthocyanidin)accumulation [81] Macadamia
integrifoliaMacadamia nuts MiMYB1 (QIJ70646.1) Involved in stress response [111] MiMYB2 (QIJ70647.1) Related to growth [133] Carica papaya Papaya CPMYB1 Fruit softening, carotenoid accumulation, fruit ripening [120] CPMYB-2 Fruit softening, carotenoid accumulation, fruit ripening [120] Theobroma cacao Cocoa TcMYB113 Regulation of flavonoid synthesis genes to determine red pod color in cocoa beans [86] Tc-MYBPA Involved in the synthesis of anthocyanins and procyanidins in cocoa [88] MYB(gene18079) Involved in the regulation of cocoa pod color [87] Citrus unshiu Citrus CitMYB52 Transcriptional regulation involved in sucrose metabolism [11] Citrus reticulata CitMYB52 Negatively regulate citrate accumulation, Inhibits the breakdown of sucrose [121] Fortunella japonica CitMYB20 (MN689609) Associated with citrus canker resistance [112] C. reticulata/C. sinensis CiMYB42 (Ciclev10021695m) Involved in the biosynthesis of citrus limonoids [113] Dimocarpus Longan Longan MYB-related-R1 May be related to chromosome telomere structure and anthocyanin synthesis [89] MYB14 May be to relevant the flowering process of longan [134] Syzygium samarangense Wax apple SsMYB1/4/20/54/77 (Transcriptome database:PRJNA284092) Promote lignin synthesis [63] SsMYB108/114/308 Promote lignin synthesis [63] SsMYB3/5/6/10/33/39
(comp40847_c1/
comp186052_c0/
comp17591_c0/
comp_45853_c0/
comp_44103_c0Promote lignin synthesis [63] SsMYB44/48/86/306/330/C1 Related to postharvest lignin synthesis [63] SsMYB48 Inhibit lignin synthesis [63] Eriobotrya japonica Loquat EjMYB1 (KF767453) Fruit chilling lignification [123,124] EjMYB2 (KF767454) Lignification Negative Regulator [43,123,124] EjMYB8 Promote lignin synthesis [125] EjMYB9 Promote lignin synthesis [125] After downloading the mango genome data from the NCBI database, both hmmsearch and local blast were used to screen and identify the members of the mango MYB gene family, initially yielding 290 candidate MYB protein sequences, using NCBI CDD (
www.ncbi.nlm.nih.gov/Structure/bwrpsb/bwrpsb.cgi ), PFAM (https://pfam.xfam.org/ ), and SMART (http://smart.embl-heidelberg.de/ )[66], we carried out further analysis of candidate protein sequences. After testing and removing incomplete and repetitive sequences, a final preliminary determination of 206 members of the Mango MYB gene family was made. The sequences of 293 members of the banana MYB transcription factor family were obtained from the study by Pucker et al.[56]. Since most MYB functions are conserved in the MYB branch, the relationship obtained after cluster analysis is close enough that the MYB can identify similar/identical target genes with cooperative, overlapping or redundant functions. In order to predict the putative functions of banana and mango MYBs, we combined them with 87 MYBs with known functions from A. thaliana to perform phylogenetic tree analysis using the MEGAX NJ method for the prediction of the functions of MYB transcription factor family members. For easier distinction, 87 known functional A. thaliana are marked with green circles, MYBs clustered with known functional A. thaliana are given a color background, branches are marked as various columns with functional annotations given. The results are shown in Fig. 3.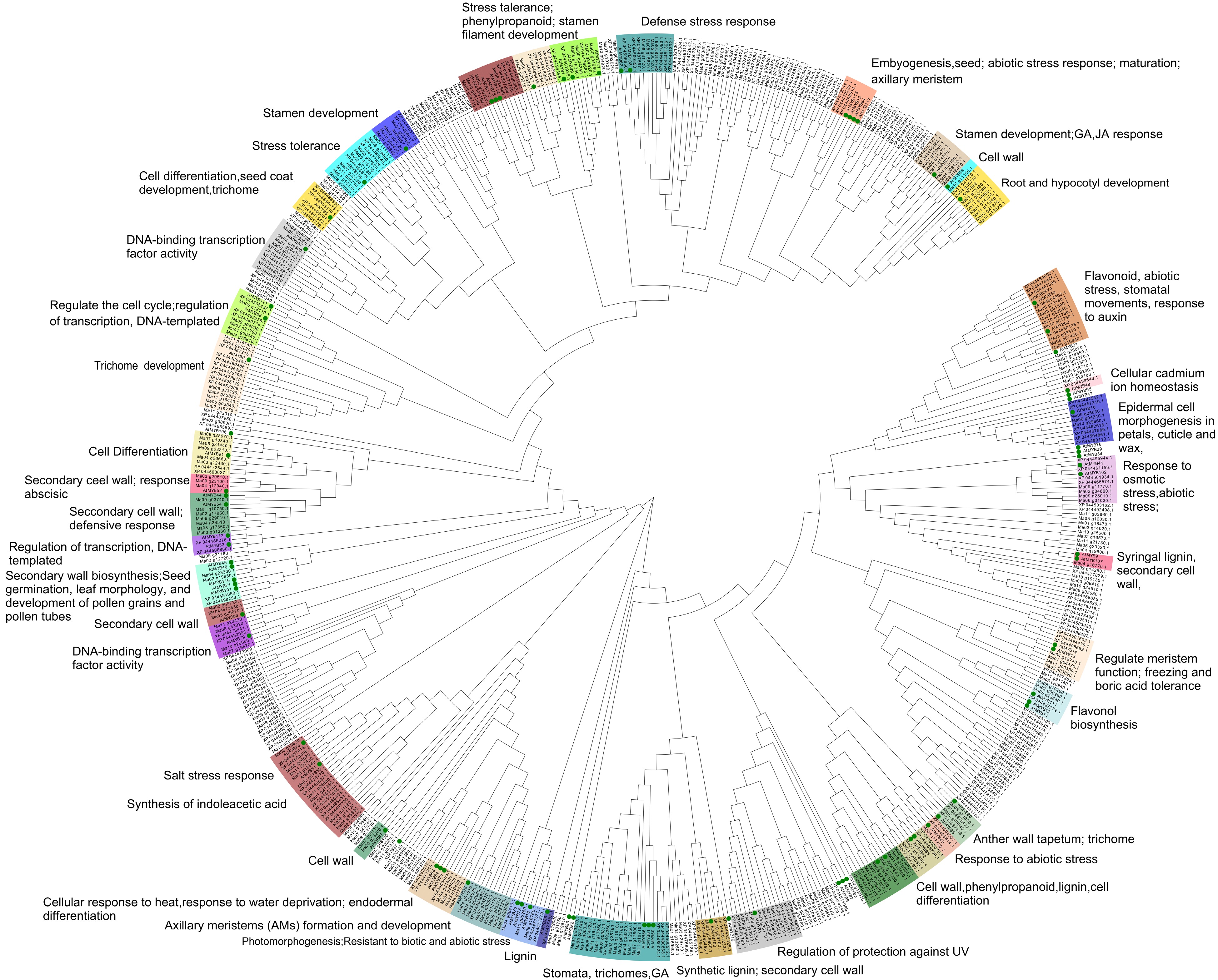
Figure 3.
Phylogenetic Neighbor-Joining (NJ) Tree.
Application of the tropical fruit MYB transcription factors in flavonoid and anthocyanin synthesis
-
The MYB transcription factor regulates secondary metabolic processes which mainly include glucosinolates, flavonoids, terpenes and lignin. Two conserved elements at the C-terminus of MYB transcription factors, SG7 and SG7-2, were found to be involved in the synthesis of flavonols. MYB transcription factors primarily work by activating or inhibiting downstream structural genes to control the synthesis of anthocyanins and flavonoids. Structural genes involved in the plant anthocyanin biosynthesis (Fig. 4) include chalcone synthase (CHS), chalcone isomerase (CHI), flavanone 3-hydroxylase (F3H), the flavonoid 3′-hydroxylase enzyme (F3′H), dihydroflavonol 4-reductase (DFR), anthocyanin synthase (ANS) and UDP glucose:flavonoid 3-O-glucosyltransferase (UFGT)[67]. Both anthocyanins and flavonoids are substrates for phytochrome synthesis and antioxidants[68]. Therefore, MYB transcription factors can regulate flower and fruit color and enhance plant stress resistance. Because the advantage of tropical fruit is that they contain more flavonoids and anthocyanins, the functional research of the MYB transcription factor in flavonoids and anthocyanins will have better prospects in tropical fruit (Fig. 5).
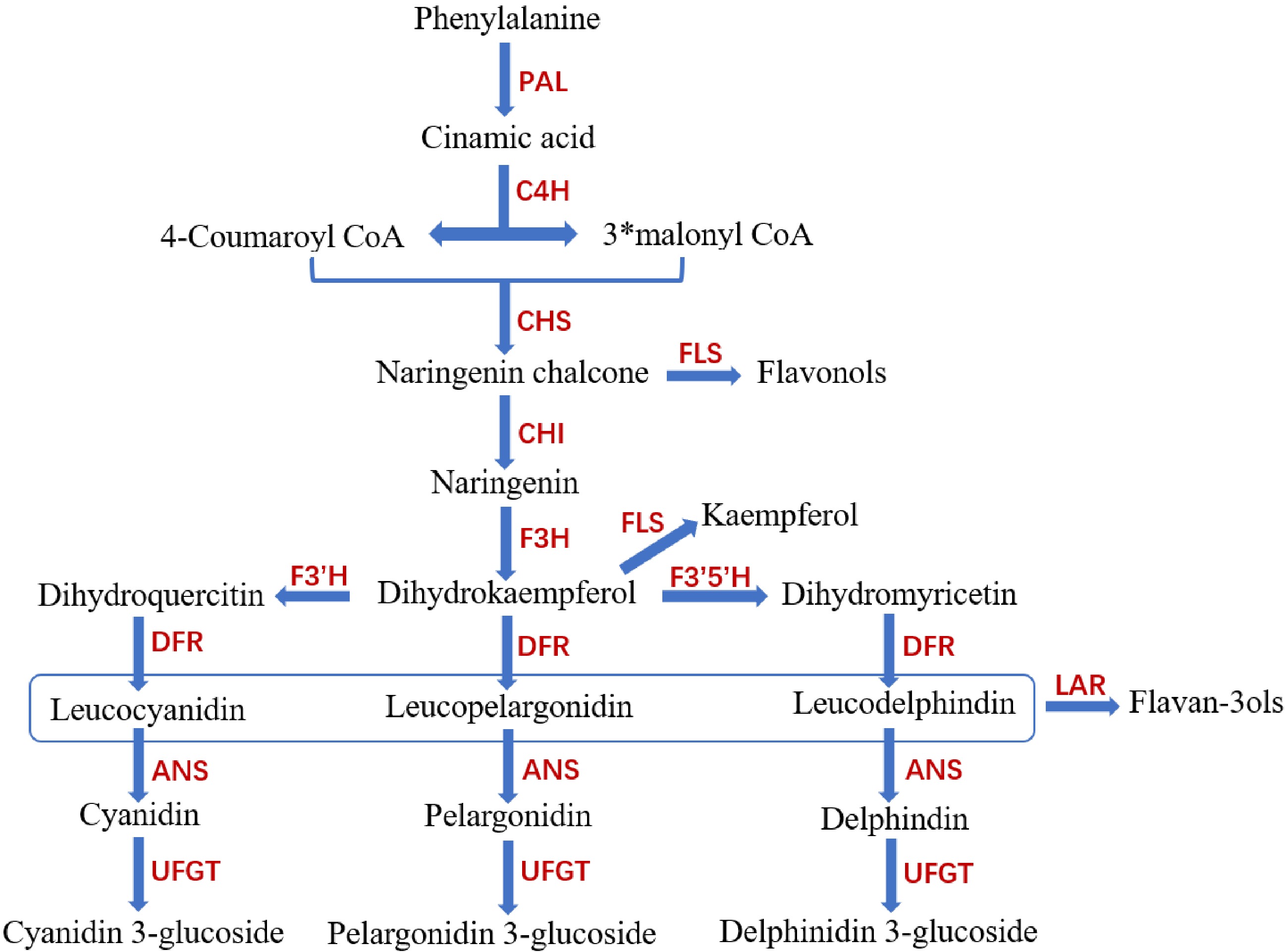
Figure 4.
Plant anthocyanin synthesis pathway. CHS (chalcone synthase), CHI (chalcone isomerase), F3H(flavanone 3-hydroxylase), F3′H (flavonoid 3′-hydroxylase Enzyme), DFR (dihydroflavonol 4-reductase), ANS (anthocyanin synthase) and UFGT (UDP glucose: flavonoid 3-O-glucosyltransferase), GST (Glutathione S-transferase), EAR (ERF-associated amphiphilic repression; Ethylene reaction factor related reaction inhibitory motifs).
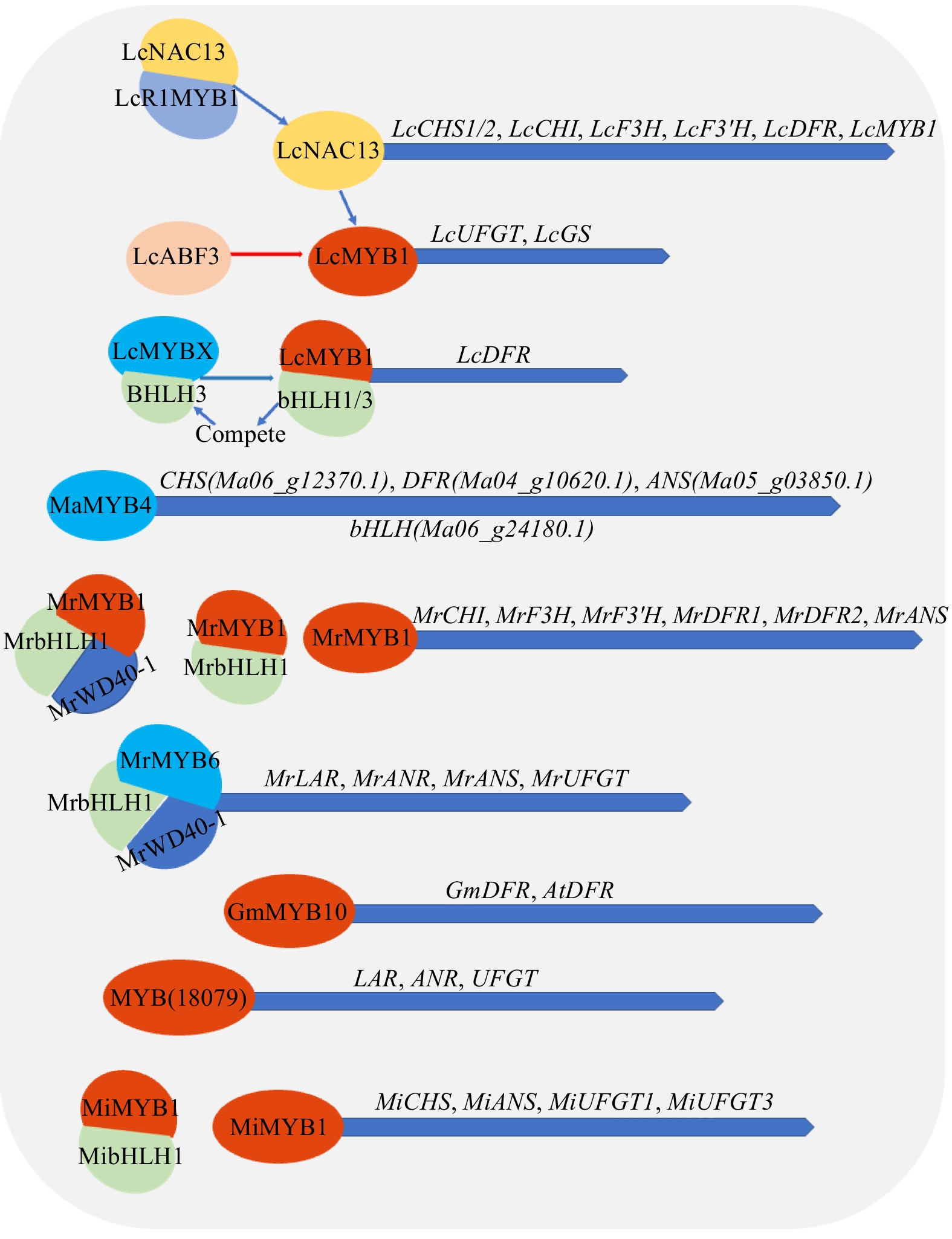
Figure 5.
Synthesis or decomposition of anthocyanins and flavonoids etc. by MYB transcription factor in tropical fruit. BHLH (basic helix-loop-helix), WD40 (WD40 repeat proteins), NAC (NAM, ATAF1, ATAF2 and CUC2). Note: blue represents inhibition, red represents activation and promotion.
The expression of the R2R3-MYB gene LcMYB1 and the accumulation of anthocyanins in litchi pericarp can be promoted by ABA and light exposure treatments. Lai et al.[69] found that this process is closely related to LcUFGT, which is probably one of the structural genes targeted and regulated by LcMYB1. LcMYB1 overexpression in tobacco can induce the accumulation of anthocyanin in almost all parts of tobacco such as pedicel, flower, ovary, seeds, and filament. The accumulation of anthocyanins in the reproductive tissues of LcMYB1-transfected tobacco is independent of the endogenous MYB and bHLH transcription factors in tobacco, and the accumulation of anthocyanins in tobacco leaves and pedicels appears to arise from the upregulation of NtAn1b (bHLH) by the overexpression of LcMYB1. LcBHLH1 and LcBHLH3 can both form LcMYB1–LcbHLH1 or LcMYB1–LcbHLH3 complexes with LcMYB1 and regulate the expression of DFR and ANS, which are the anthocyanin biosynthesis genes, thereby promoting the biosynthesis of anthocyanins[70]. Qin et al.[71] used the Agrobacterium rhizogenes-mediated hairy root transgenic system to overexpress LcMYB1 in the stem segments of 'pink guiwei' seedlings to obtain pink hairs with a higher content of anthocyanins, procyanidins and flavonols in the shaped root. In addition, another study found that LcMYB1 can directly activate LcGST4 (glutathione S-transferase) with ABA-responsive and light-responsive elements, operating in the accumulation of lychee anthocyanins[72]. At the same time, because LcMYB1 contains an ABRE motif, it can also be directly regulated by LcABF3 to mediate anthocyanin biosynthesis[73]. The internal regulatory network of plants is extremely complex. LcMYB1 can promote the accumulation of anthocyanins, and LcNAC13 can negatively regulate its expression and inhibit the accumulation of anthocyanins. In turn, LcR1MYB1, an R1-MYB transcription factor, can physically interact with LcNAC13 to antagonize the anthocyanin synthesis-related genes LcCHI, LcCHS1/2, LcF3H, LcDFR, LcF3'H, and LcMYB1 during maturation of lychee fruit, thereby reversing the effects of LcNAC13[74]. Some studies have found that MYB genes that negatively regulate secondary metabolic pathways such as anthocyanins also exist in lychee. LcMYBx contains a conserved [D/E]Lx2[R/K]x3Lx6Lx3R motif encodes a typical 3R-MYB protein, for interaction with bHLH proteins. At the later phases of leaf development, the synthesis of anthocyanins decreased and the expression level increased, presumably in negative agreement with the anthocyanins synthesis in leaves. Zhao et al.[75] found LcMYBx could inhibit the ability of LcMYB1–LcbHLH3 complex to activate the LcDFR genes and compete with LcMYB1 for LcbHLHs, thereby preventing the LcMYB1–LcbHLHs complex's activation of LcDFR and negatively controlling anthocyanin biosynthesis. MaMYB4 inhibits banana anthocyanin biosynthesis, which may be due to a two-stage inhibitory mechanism, including the expression of CHS (Ma06_g12370.1), DFR (Ma04_g10620.1), and ANS (Ma05_g03850.1). The inhibition and parallel down regulation of bHLH resulted in interference with the correct aggregation of the MYB–bHLH–WD40 activation complex and MaMYB4 has no physically interaction with bHLH (Ma06_g24180.1)[76]. Blue light plays an important position in the early stage of plant growth and development, and can encourage plants to produce more chlorophyll and help seedlings better absorb and utilize the energy of photosynthesis. The blue light treated bayberry rose the expression of MrMYB1 and the structural genes which participate in anthocyanin biosynthesis, such as MrCHI, MrF3'H, MrF3H, MrDFR1, MrANS and MrDFR2[77]. The opposite is true that bagging inhibited the expression of all anthocyanin biosynthesis genes and MrMYB1[78]. MrMYB1 can form binary complexes with MrbHLH1, and they can selectively activate the promoters of five of the eight anthocyanin biosynthesis genes from Myrica rubra[79]. Subsequent studies revealed an earlier and stronger induction of anthocyanin accumulation by ternary expression of MrMYB1–MrbHLH1–MrWD40-1 than by binary expression of MrMYB1–MrbHLH1[80]. The allele of Chinese bayberry, MrMYB1 and MrMYB1d, is associated with the accumulation of anthocyanin in myricetin and has an inhibitory effect on anthocyanins[81]. Shi et al.[82] found that MrMYB6 may negatively regulate the accumulation of anthocyanins and PA (proanthocyanidin) in myricetus. The interactions of MrMYB6 with MrbHLH1 and MrWD40-1 form a functional complex that directly inhibits both the PA-specific genes MrLAR and MrANR and the anthocyanin-specific genes MrANS and MrUFGT in terms of promoter activity. Pitaya MYB regulates anthocyanin synthesis[83]. HuMYB1814 is related to anthocyanin synthesis[84]. Zeng et al.[85] conducted in-depth research on the screened pitaya MYB transcription factor genes Unigene0036816, Unigene0032395, and Unigene0034590, and found that the expression levels of these three MYB transcription factors in the red-fleshed pitaya at the ripening stage were remarkably higher than in the green ripening stage, and Unigene0021844, Unigene0027949 and Unigene0010156 in the phylogenetic tree analysis have high homology with the sugar beet BvMYB1 transcription factor already considered to be involved in regulating the metabolic pathway of beta lain. It is speculated that their functions may be the same, but further verification is required. Palapol et al.[86] studied mangosteen GmMYB10 and found that it can activate GmDFR and AtDFR promoters. The degree of activation of DFR is similar to that of AtPAP1, which regulates pigmentation in A. thaliana and exert an essential role in the regulation of anthocyanin biosynthesis on crops, especially postharvest. At the same time, it was revealed that the ethylene impact on anthocyanin biosynthesis could be achieved by regulating GmMYB10 expression. Cocoa (Theobroma cacao L.) MYB may be involved in regulating the color change of cocoa pods from light green or green to lavender and regulating flavonoid synthesis genes[87]. The change in cocoa pod color from light green or green to lavender correlates with high levels of anthocyanins (anthocyanin 3-O-galactoside, anthocyanin 3-O-glucoside and anthocyanin O-syringic acid). Li et al.[88] found that MYB (gene 18079) positively regulates late genes (LAR, ANR, UFGT) that participate in the flavonoid biosynthetic pathway. The Tc-MYBPA gene, which is an R2R3-type MYB transcription factor, is an Arabidopsis TT2-like transcription factor that may be involved in controlling the synthesis of anthocyanins and proanthocyanidins in cocoa[89]. TcMYB113 may also be a part of regulation of the color of cocoa pods[87]. A synonymous mutation of a SNP (position 20878891) significantly associated with pod color was found in the coding region of TcMYB113, which is also located in the target site of small interfering RNAs (tasiRNAs), this SNP site appears to affect the transcriptional level of TcMYB113[87]. The research of Li et al.[90] on longan MYB-related-R1 preliminarily speculated that the function of this gene is related to the structure of chromosome telomeres and anthocyanin synthesis. Coconut MYB is related to the biosynthesis of flavonoids[91]. Papaya MYB is involved in anthocyanin synthesis[60]. Bai et al.[92,93] cloned the mango MinMYB10 and MinMYBPA genes, and constructed the corresponding overexpression vector and silencing vector, predicting that MiMYBPA may be related to the synthesis of mango peel anthocyanins. As an R2R3-MYB transcription factor MiMYB1 regulates four genes, MiCHS, MiANS, MiUFGT1 and MiUFGT3, and plays an important role in mango anthocyanin accumulation. Later studies concluded that MiMYB1 expression levels were affected by light stimulation, as they increased with light intensity, and that the increased intensity paralleled the accumulation of anthocyanins[94]. Tobacco transient expression assays showed that MiMYB1 and MibHLH2 co-infiltration induced the accumulation of anthocyanins in tobacco, but they did not accumulate anthocyanins when infiltrated alone.
In summary, most of the MYB transcription factors in tropical fruit with known functions are involved in regulating the anthocyanin synthesis pathway by regulating the structural genes DFR and ANS. The main function of these two structural genes is to catalyze the oxidative dehydration of colorless anthocyanins to form colored anthocyanins, which endow flowers and fruits with color. However, the functional research reports of other genes such as MiMYBPA and MIMYB10 are still in the stage of gene cloning and vector construction. Current reports regarding the functionality of MYB transcription factors like papaya are still focused on profiling of transcriptomic and metabolomic data, and the function of an MYB transcription factor has not been studied in depth.
Tropical fruit tree MYB transcription factors in response to environmental stresses
-
The most suitable flowering temperature for tropical and subtropical fruit is 18−25 °C, and climatic conditions have a huge impact on the flowering of these plants[95]. Abiotic stresses lead to flowering obstacles by affecting the process of dormancy and flower bud differentiation in tropical fruit. It also reduces the accumulation of carbohydrates in shoots, leading to reduced tree vigor and even death[96]. Combined with the slightly abnormal change in the global climate in recent years[97], it is necessary to conduct in-depth research on the MYB transcription factor as the main transcription factor regulating stress resistance (Fig. 6). Some MYB transcription factors can be localized directly in guard cells. MYB transcription factors can widely participate in the plant stress resistance process and regulate plant stress tolerance by regulating plant secondary metabolites such as hormones[98], flavonoids, waxes and cutin[99,100]. Stresses such as ABA, drought and salinity can also provoke transcription and translation of MYB genes, then regulate secondary cell wall formation by promoting/repressing lignin, cellulose and xylan biosynthesis via the ABA signaling pathway to regulate plant stomatal size[101].
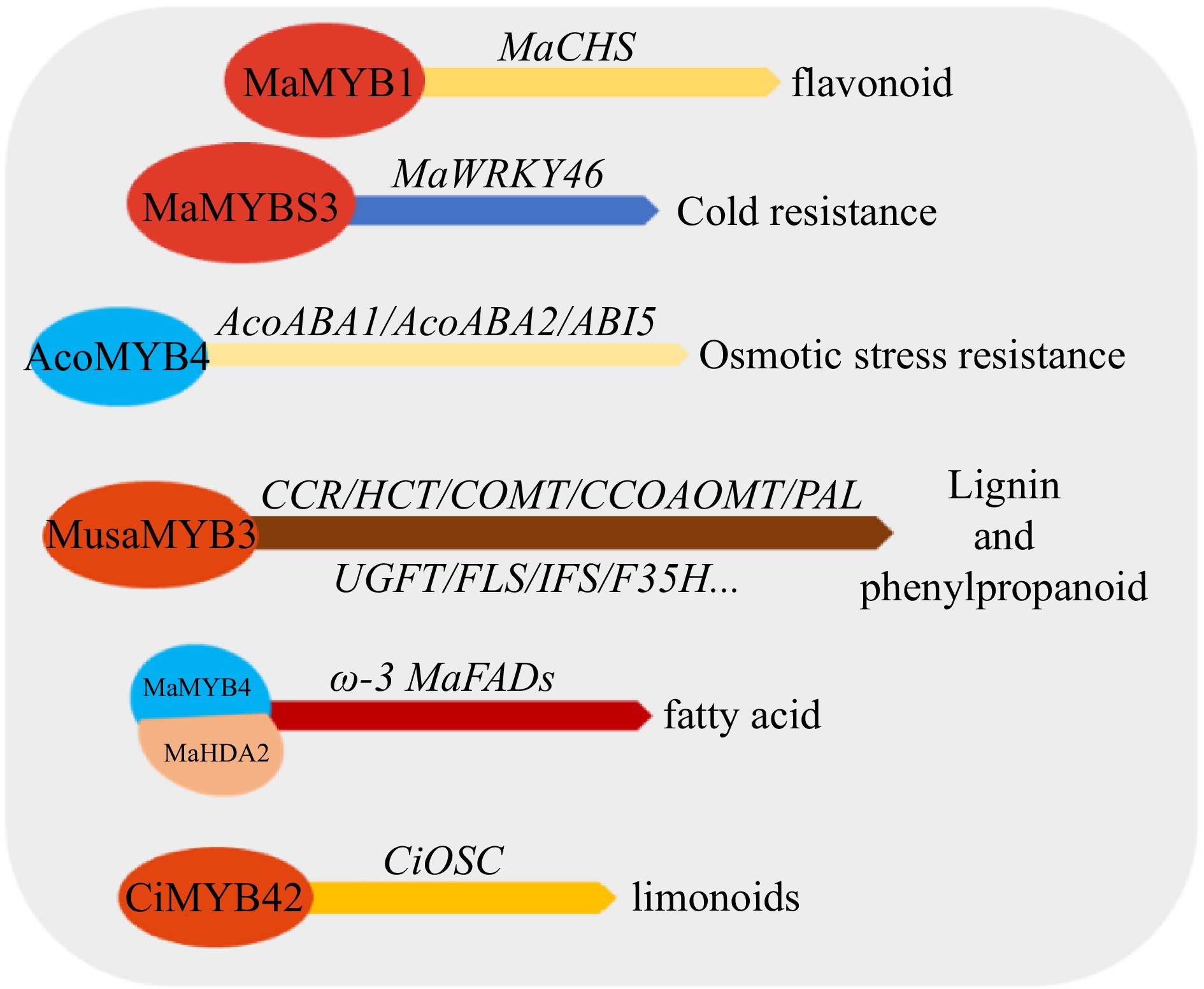
Figure 6.
Effects of MYB transcription factors on abiotic and biotic stress tolerance in tropical fruit. ABA1/ABA2 (genes responsible for ABA synthesis), ABI5 (Abscisic acid-insensitive 5, Key genes in response to ABA signals), CBF (C-repeat binding transcription factor/dehydrate responsive element binding factor, DREB), CHS (chalcone isomerase), FAD (fatty acid desaturase), CCR (Cinnamoyl CoA reductase), HCT (hydroxycinnamoyl CoA: shikimate hydroxycinnamoyl transferase), COMT (caffeic acid 3-O-methyltransferase), PAL (phenylalanine ammonia-lyase), CCOAOMT (caf-feoyl-CoA O-methyltransferase), UGFT (UDP-glucose: flavonoid-3-O-glucosyltransferase), FLS (flavonol synthase), IFS (isoflavone synthase), F35H (flavonoid 3,5-hydroxylases), HDA (histone deacetylase), OSC (Oxidosqualene cyclase).
MYB transcription factors can both regulate abiotic stress and participate in biotic stress response. Fusarium wilt is a serious disease in bananas. Wang et al.[102] found that MaMYB1 in banana responds to stress treatments such as drought, low temperature, salt and Fusarium wilt, and performs an essential regulatory function in the resistance of banana to stress. MaMYB1 can bind to the MaCHS promoter region with cis-acting elements such as drought, ABA, and salicylic acid activation and induction to regulate flavonoid synthesis[103]. Yang et al.[104] found that MaMYBR1 gene expression was significantly upregulated in resistant varieties inoculated with Fusarium wilt, while significantly downregulated in susceptible varieties, indicating that the MaMYBR1 gene was possibly implicated in the process of Fusarium wilt resistance in banana. Long-term magnesium deficiency (MD) inhibits the development of bananas, resulting in the chlorosis of leaves. Yang et al.[105] found that the MusaMYB108 mutant was hypersensitive to magnesium deficiency than the wild type and had lower root hair density than the wild type, suggesting that it is involved in regulating the growth and development of banana leaves and roots under long-term magnesium deficiency conditions. Mazumdar et al.[106] found that the expression of banana MaMYB14, MaMYB63 and MaMYB110 was substantially enhanced in banana roots in response to salinity stress. Fatty acid synthesis is related to plant cold stress tolerance, and MaMYB4 was discovered to recruit MaHDA2 (histone deacetylase) by Song et al.[107] and inhibit the transcription of ω-3 MaFADs (MaFAD3-1, MaFAD3-3, MaFAD3-4 and MaFAD3-7) by affecting their acetylation levels. Tak et al.[108] cloned the R2R3-MYB transcription factor gene MusaMYB31, which is highly expressed in the bulb and root tissue of the banana 'Rasthali'. Overexpression of MaMYB31 can cause a reduction in cell wall lignin in banana vascular tissue, and the transgenic line showed that the lignin was included. Alterations in the transcription levels of general phenylpropane pathway genes, including biosynthetic pathway genes, indicate that MaMYB31 has a potential role in inhibiting banana lignin and polyphenol biosynthesis. Overexpression of the 1R-MYB transcription factor gene MaMYBS3 in banana plants can enhance the cold tolerance of banana effectively, but the process is independent of the ICE1–CBF cold signaling pathway. Conversely, the overexpression of MaMYBS3 suppressed the transcription of MaCBF1 and MaCBF2 in the ICE1–CBF cold signaling pathway. It achieves cold tolerance by stimulating the transcription of MaWRKY46[109]. Pineapple AcoMYB1, as a positive regulator, participates in the response process of pineapple chilling injury and drought stress, and has a regulatory function in the early development and late ripening of pineapple fruit[110]. Chen et al.[111] found that under osmotic stress, AcoMYB4 inhibits ABA biosynthesis and negatively regulates osmolality by reducing the transcription of genes responsible for ABA synthesis (AcoABA1, and AcoABA2) and the ABA signaling factor AcoABI5. Water stress may be an important cause of mangosteen translucent flash disease (TFD). Matra et al.[65] applied transcriptome analysis to identify whether the MYB gene family responds to water stress in mangosteen. MiMYB1, a typical plant R2R3–MYB transcription factor, instrumental in the resistance of macadamia[112]. Citrus CitMYB20 is a transcription factor associated with citrus canker resistance[113]. Coconut MYB has been implicated in flavonoid biosynthesis and salt stress[91]. Limonin has anthelmintic, antibacterial and antiviral properties. CiMYB42 is an R2R3-MYB transcription factor in which binds to a type II MYB core sequence (TTGTTG) in the CiOSC promoter to induces its transcription[114].
In summary, relatively few studies have been conducted on the stress function of MYB transcription factors in tropical fruits, and the regulatory mechanism is still unclear. Moreover, most of the fruit and leaves of tropical fruit contain thicker waxy layers and cuticles, which are two natural stress-resistant structures. Therefore, in future research of the MYB transcription factor family in tropical fruit, in addition to expanding the functional study of stress response, we can also explore the relationship between MYB transcription factors and plant cuticle and waxy layer structures.
Tropical fruit MYB transcription factors in fruit ripening, fruit quality and fruit storage
-
Tropical fruit have a short time-to-market and vary in fruit quality. It is an important focus of the fruit tree market to cultivate tropical fruit with different stages of maturity and different tastes to meet market demands in different periods. For example, PbrMYB169 actively regulates stone cell lignification in pear fruit[21]. CgMYB58 is an IAA and ABA-responsive transcription factor that activates lignin biosynthesis and causes granulation of grapefruit juice sacs[115]. Therefore, it is very important to explore the application of the MYB genes in the fruit ripening as well as fruit quality of tropical fruit (Fig. 7).
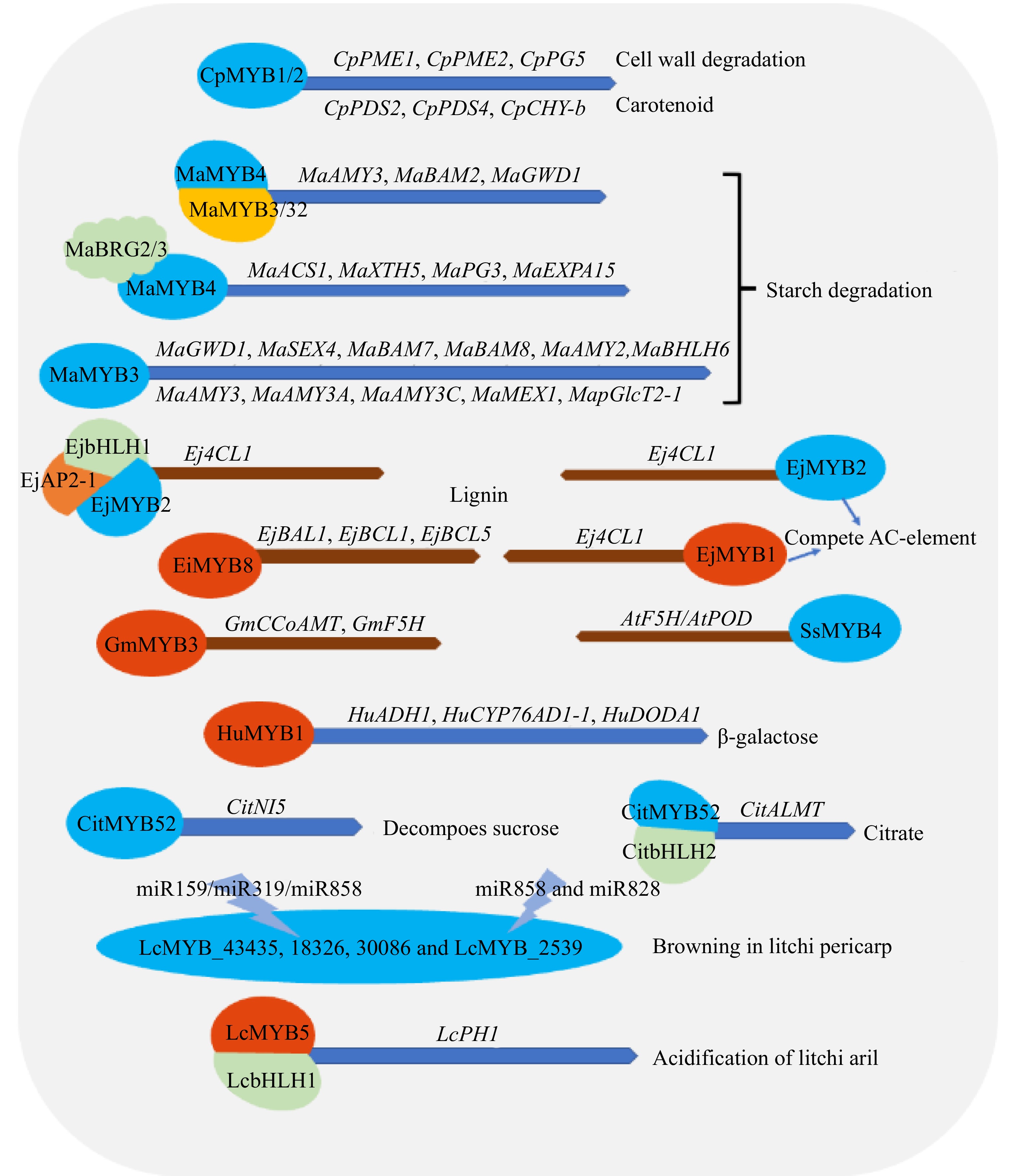
Figure 7.
Regulatory effects of MYB transcription factor on fruit ripening and fruit quality in tropical fruit crops. GWD (glucan-water dikinase), SEX4 (phosphoglucan phosphatases starch excess 4), BAM (Beta-amylase), AMY (α-amylases), MEX (maltose excess protein), pGlcT (plastidic glucose translocator), F5H (ferulate 5-hydroxylase), POD (Peroxidase), 4CL (4-coumarate;coenzyme A ligase), PAL (phenylalanine ammonia-lyase), PDS (Phytoene desaturase), CHY (Chymopapain), PME (pectinmethylesterase), PGs (polygalacturonases), NI (neutral/alkaline invertase), PGK (Phosphoglycerate kinase), Ca2+-ATPase (A multiregulated enzyme that is stimulated by calmodulin (CaM), KCI and lysophospholipids), ALMT (aluminum-activated malate transporter), ADH (Alcohol dehydrogenase), CYP76AD1 (cytochrome P 450 76AD1), DODA ((DOPA)-4,5-dioxygenase), AP2 (APETALA2), BRG2 (RING finger E3 ligases). MaACS1 (ACC synthase), MaXTH5 (cell wall modifying enzyme), MaPG3 (exopolymeric galacturonidase), MaEXPA15 (α-expansion protein) Note: blue represents inhibition, red represents activation and promotion.
Jiang et al.[116] found that the R1-type MYB transcription factor MaMYB16L is a starch degradation-related gene, which negatively regulates the transcription of the fruit ripening-related genes MaAMY3, MaBAM7, MaISA2 and MaDREB2 during the regulation of banana fruit ripening, and negatively regulates fruit ripening. MaMYB16L is alternatively spliced to produce two transcripts in banana fruit: the full-length isoform, MaMYB16L, and the truncated form, MaMYB16S. Down-regulation of MaMYB16L and up-regulation of MaMYB16S exacerbated the alternative splicing process during banana fruit ripening. Since a DNA binding domain is lacking. MaMBY16S competes with functional MYB16L for binding and forms a non-functional heterodimer MaMYB16L–MaMYB16S, thereby reducing the binding ability of MaMYB16L to fruit ripening-related genes. So the alternative splicing of MaMYB16L may result in a self-regulating circuit to participate in the regulation of banana fruit ripening.
Fan et al.[117] concluded that the MYB gene MaMYB3 regulates maturation and ethylene repression. It can interact with MaGWD1 (a member of the glucan water dikinase (GWD) family, which is an important enzyme in starch degradation), MaSEX4, MaBAM7–MaBAM8, MaAMY2B, MaAMY3, MaAMY3A, MaAMY3C, MaMEX1 and MapGlcT2-1. MabHLH6 (activator of starch degradation-related genes) inhibits the transcription of all 11 starch degradation-related genes, resulting in delayed ripening of banana fruit. HuMYB1 regulates fruit ripening by inhibiting the β-galactose biosynthesis-related genes HuADH1, HuCYP76AD1-1 and HuDODA1[57]. Sucrose is an influential carbohydrate in plants; after sucrose is decomposed, fructose and glucose are the source of sweetness in fruits. At the same time, sucrose has a major effect on plant growth, development and resistance to stress. The expression pattern of CitNI5 is related to the accumulation of sucrose, to which it is negatively correlated. CitMYB52 can bind to the ATTGTC core element in the CitNI5 promoter and transcriptionally inhibit the expression of CitNI5, thereby inhibiting the decomposition of fruit sucrose[118]. Anohylene biosynthesis and cell wall-modifying genes such as MaACS1 (ACC synthase), MaXTH5 (cell wall-modifying enzyme), MaPG3 (exopolymeric galacturonidase), and MaEXPA15 (α-expansion protein). MaMYB4 is a fruit ripening repressor inhibiting the binding of promoters of various ripening-related genes, among them MaACS1, MaXTH5, MaPG3, MAEXPA15. In addition, MaBRG2/3, two RING finger E3 ligases, were found to interact with MaMYB4 in tomatoes exogenously expressing it to ubiquitinate it, resulting in reduced MaMYB4 accumulation at fruit ripening[119]. Ba[120] found that MaMYB4 can form a protein complex with MaMYB3/32, enhancing the transcriptional repression of the starch degradation-related genes MaAMY3, MaBAM2, and MaGWD1, as well as MaPWD1 promoters. CpMYB1 and CpMYB2 are transcription factor inhibitors that regulate papaya fruit softening and carotenoid biosynthesis-related genes by binding to the promoters of the cell wall degradation genes CpPME1, CpPME2 and CpPG5 and the carotenoid biosynthesis genes CpPDS2, CpPDS4 and CpCHY-b[121]. HuMYB1 is an R2R3-type MYB transcription factor; its R3 domain contains the bHLH-binding motif, and its C-terminal contains the C1 motif (KLIsrGDPxT/SHRxI/L), C2 motif (pdLNLD/ELxiG/S or LxLxL), C3 Motif (Cx1–2Cx7–12Cx2C) and C4 motif sequence (FLGLx4-7V/ LLD/GF/YR/Sx1LEMK). Among them, C1 and C2 motifs participate in bHLH interactions and are considered as promoter repression domains. C4 motif was confirmed to be a negative regulator of floral volatile benzoic acid/phenylpropionic acid compounds. CitMYB52 was found to interact with CitbHLH2 and synergistically transduce CitALMT to negatively regulate citrate accumulation, providing an important basis for future improvement of citrus fruit quality[122]. Shen et al.[123] found that papaya MYB is connected to fruit ripening, EjbHLH1–EjMYB2–EjAP2-1 was associsted with the regulatory of loquat fruit lignification and affects the quality of loquat fruit[43], and EjMYB1 and EjMYB2 of Loquat directly regulate related genes through the AC-element that binds to the promoter region of the Ej4CL1 gene and is involved in chilling lignification of fruits. EjMYB1 is a positive regulator of fruit lignification, while EjMYB2 is a negative regulator. EjMYB2 can bind to the promoter of lignin synthesis genes directly to repress their transcription, as well as compete with EjMYB1 for AC-elements to inhibit corresponding gene expression. Low-temperature conditions (LTC) can simultaneously regulate EjMYB1 and EjMYB2 to reduce chilling-induced lignification, while heat treatment (HT) specifically inhibits EjMYB1 to regulate fruit lignification[124,125]. The above two treatments also inhibited the expression of EjMYB8. EjMYB8 transactivates the lignin-associated genes EjBAL1, EjBCL1 and EjBCL5 promoters to stimulate lignin synthesis. EjMYB1 and EjMYB8 work together to lead to a noticeable rise in the expression of the Ej4CL1 promoter (~21-fold). However, the findings of yeast two-hybrid analysis showed an inability of EjMYB8 to interact directly with EjMYB1[126]. The expression levels of wax apple (Syzygium samarangense) MYB genes SsMYB1, SsMYB4, SsMYB20, SsMYB54, SsMYB77, SsMYB108, SsMYB114 and SsMYB308 showed an overall upward trend during storage, and nitric oxide (NO) treatment slowed down the upward trend. It is hypothesized these transcription factors may be implicated in the synthesis of lignin. However, the expression levels of SsMYB3, SsMYB5, SsMYB6, SsMYB10, SsMYB33, SsMYB39, SsMYB44, SsMYB48, SsMYB86, SsMYB306, SsMYB330 and SsMYBC1 decreased continuously during storage, and nitric oxide treatment inhibited the downward trend. The transcriptional repressor slows down the synthesis of lignin. Nie[63] continued to study SsMYB48 and concluded that the gene has an repressive effect on key enzyme genes of lignin synthesis, AtF5H and AtPOD. Specific MYB genes (LcMYB_43435, 18326 and 30086, LcMYB_2539) targeted by miR159/319/858/828 and energy-related genes targeted by miR2118 might relate to regulating differences in storability among litchi varieties[127]. The expression of the binary complex composed of LcMYB5 and LcbHLH1 is coincident with the presumptive expression of the gene LcPH1 for tissue acidification. Litchi LcMYB5 is comply with the expression pattern of the major organic acid malic acid in litchi arils and is involved in cellular acidification[128]. The firmness and lignin content of damaged mangosteen pericarp increased rapidly after fresh mangosteen (Garcinia mangostana L.) impact, which was due to significantly increased transcript levels of GmCCoAMT and GmF5H and the stress-related R2R3-MYB transcription factor gene GmMYB30 after impact, but this process can be delayed with increasing nitrogen concentration[129].
At present, bananas are the most studied tropical fruit in regard to the regulation mechanism of fruit ripening based on the MYB transcription factor family. There are few related studies on other tropical fruit. The fruit maturity period and the length of the postharvest preservation time have a huge impact on the market price of fruit. Therefore, it is necessary to study the regulation of fruit ripening based on MYB transcription factors.
Tropical fruit's MYB transcription factors influence its growth and development, especially flower development
-
In addition to participating in the process of stress resistance and fruit ripening, and regulating fruit quality, MYB transcription factors can additionally regulate the flower organ and other tissue growth and development in plants through the regulation of the expression of related functional genes. For example, MYB transcription factors can promote microspore mother cells and their development into normal fertile pollen[130]. Wheat TaMYB72 can regulate wheat flowering time[131]. MULTI-FLORET SPIKELET 2, a MYB transcription factor, characteristics the differentiation direction of rice spikelet meristems and the characteristics of floral organs[132]. The overexpression of MdMYB3 in apple resulted in longer pedicels and pistil styles intransgenic plants than in wild-type plants[133].
The increase in petal color may be due to higher anthocyanin or lower pH, and the flowers of LcMYB5-transformed tobacco showed remarkable changes in phenotype compared to wild-type tobacco, such as darker petals, filaments, ovaries, and stamens. The flowers were much larger than the wild type. The flower and leaf PH of the transgenic strain were significantly lower than those of the transgenic tobacco and petunia, but the accumulation of anthocyanins was only found in the reproductive tissues of the tobacco. These changes were caused by LcMYB5 through the direct activation of the key anthocyanin synthesis genes DFR, CHI, and F3H, or the indirect upregulation of the expression of the endogenous gene bHLH[128]. LcMYB1 can regulate the accumulation of anthocyanins in tobacco pedicels, flower ovaries, filaments, and petals[69] (Fig. 8). Some MYB genes among 103 pineapple R2R3-MYB genes may participate the process of pineapple flower formation and induction in response to ethylene stimulation[54]. The MiMYB2 gene can influence the growth and development process of macadamia and affect macadamia yield[134]. Longan MYB14 may be related to longan flowering[135]. Whole genome transcriptome analysis of the inflorescences of 'Gnomitz' and 'Sophora' during high and low-temperature conditions suggest that MYBs may be the main transcription factors for high temperature-induced, hormone-controlled floral abortion[136], The jackfruit MYB gene is participateed in the expression of carbon fixation and energy metabolism genes in albino jackfruit seedlings, affecting their light response or photomorphogenesis and normal growth[137]. Loquat MYBs are involved in flower buddifferentiation[138]. Jamaluddin identified MYB (CP00006G00950 and CP00005G03080) as a possible transcription factor important in papaya callus that may work in the tissue differentiation of papaya embryogenic callus[64].
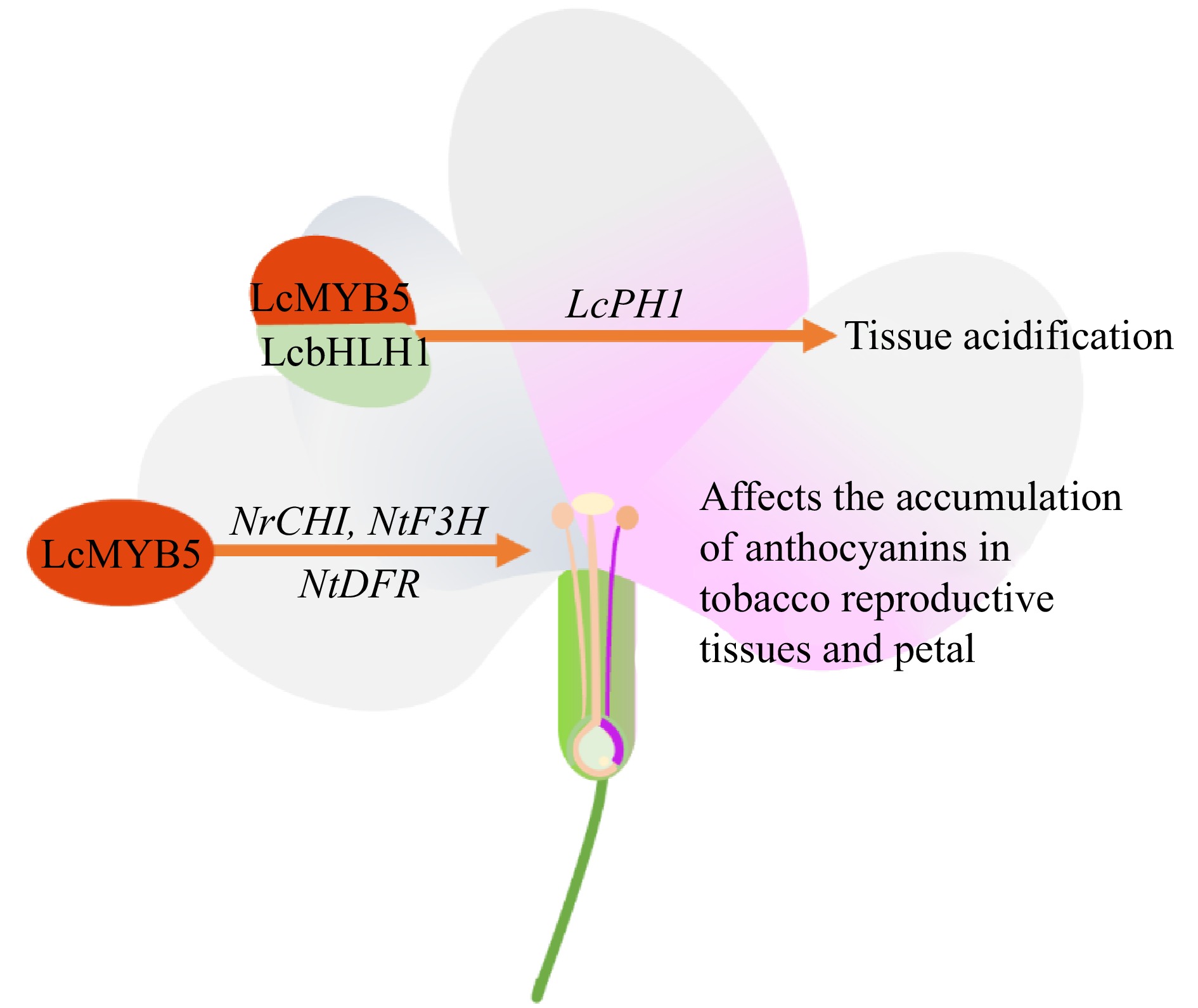
Figure 8.
Effects of MYB transcription factors on tropical fruit crops flowers. PH1 (P-ATPase), CHI (chalcone isomerase), F3H (flavanone 3-hydroxylase), DFR (dihydroflavonol 4-reductase).
MYB transcription factors, which are candidate genes for the regulation of sex differentiation in floral organs, is essential in dealing with the problem of abnormal flower development in tropical fruit crops that may be caused by the unstable global climate.
-
The main objectives of fruit tree breeding are to obtain varieties with a shorter juvenile non-bearing period, high yield, long ripening period, regular fruit shape, good fruit color, good flavor, consistent harvesting period[139], rich in micro/macronutrients, contaning low levels of antinutrients[140]. In addition to this, new varieties need to be tolerant to biotic and abiotic stresses to cope with the vagaries of the global climate and the ability to adapt effectively to different planting and postharvest regimes[141]. Because the traditional fruit breeding cycle is too long and the traits are influenced by environmental factors, biological techniques for fruit breeding have become very efficient ways to breed fruit. A large amount of research have been conducted to demonstrate that MYB transcription factors can satisfy the above-mentioned breeding needs. An MYB transcription factor is regulated as a large network that contributes to plant growth and development through a complex regulatory mechanism (Table 1).
Nonetheless, scarcity of studies on MYB transcription factors in tropical fruit, and these studies mainly focus on the process of flavonoids, anthocyanins, and resistance to abiotic stress. Part of the research on MYB transcription factors in tropical fruit is still in its infancy, and the regulatory mechanisms of MYB transcription factors in tropical fruits are not sufficiently studied. The research on the MYB transcription factor family is also mainly focused on R2R3-MYB and less research has been conducted on 1R-MYB, 3R-MYB, and 4R-MYB. In future research, we can use multi-omics joint analysis, combined with yeast single-hybrid screening, yeast two-hybrid screening, electrophoretic mobility shift assays[18], genetic transformation, and other experiments to discover more powerful MYB genes that can solve practical production problems, which can improve tropical fruit varieties. Moreover, such approaches could provide a reference for solving problems such as the fact that in some international regions, the location of tropical fruit planting and the growth characteristics for planting tropical fruit (not high suitability) are not sufficiently reasonable and the planting varieties are insufficiently rich. These factors result in wasting advantageous climatic resources brought about by geography.
This project was supported by Hainan Provincial Natural Science Foundation of China (321RC1088, 321MS091, 320QN305 and 321RC686),the project of Sanya Yazhou Bay Science and Technology City (SCKJ-JYRC-2022-84, SCKJ-JYRC-2022-93).
-
The authors declare that they have no conflict of interest.
-
accompanies this paper at (https://www.maxapress.com/article/doi/10.48130/TP-2022-0005)
-
Received 30 June 2022; Accepted 11 August 2022; Published online 30 August 2022
-
# These authors contributed equally: Yanshu Zhang, Yi Xu
- Supplemental Table S1 List of tropical fruit.
- Copyright: © 2022 by the author(s). Published by Maximum Academic Press on behalf of Hainan University. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Zhang Y, Xu Y, Huang D, Xing W, Wu B, et al. 2022. Research progress on the MYB transcription factors in tropical fruit. Tropical Plants 1:5 doi: 10.48130/TP-2022-0005 

Research progress on the MYB transcription factors in tropical fruit
- Received Date: 30 June 2022
- Accepted Date: 11 August 2022
- Published Online: 30 August 2022
Abstract: Tropical fruits, such as bananas, passion fruit, mangoes, and lychees, are rich in vitamins, flavonoids and non-flavonoid phenols, and are the healthiest way to intake bioactive compounds in one's daily diet. Coupled with their unique taste, tropical fruits are becoming more increasingly popular. The production of bananas is second only after grapes, while other tropical fruit are limited by the planting environment, resulting in a relatively low planting area and yield. With the improvement in breeding technology and planting technology, the development of tropical fruit crops will have broad prospects. MYB plays a major regulatory role in several biological processes, including growth and development of plants, quality formation, and stress resistance. There is a comparatively junior level of research on tropical fruit crops compared to grain fruit and model plants. Despite the progress that has been made in the development of molecular biology methods that can transform the genetic makeup of tropical fruit crops, the functional understanding of the multiple MYB genes is still in its initial stages. This paper aims to review the current status of the research on the development of these genes and their prospects. It is expected to provide a reference for the study of MYB transcription factors in tropical fruit crops and to provide new ideas for the functional resolution of MYB transcription factors in different biological processes.
-
Key words:
- Tropical fruit crops /
- MYB transcription factor /
- Anthocyanins /
- Flavonoids /
- Stress resistance /
- Fruit ripening /
- Flower development