
Abstract
An increasing bulk of evidence shows that through different mechanisms, experienced-based or voluntary, reactive or proactive, human beings can attenuate the distracting impact of salient visual, albeit irrelevant, stimuli. Current mechanisms assume that this is achieved by suppressing the salient distractor’s features or location at the priority map level, or at lower dimension-based maps levels. However, this functional architecture has so far ignored the role of time in distractors filtering, a key question that we have addressed in the present study. We found that during a visual discrimination task, a “standard” onset distractor, always appearing at the same interval from the beginning of the trial, was subject to habituation. Crucially, however, when the onset distractor was unfrequently presented with an unexpected 1-second delay, it reboosted capture at full strength, while the “standard” distractor continued to remain overall habituated. As predicted by Sokolov’s (1963, Annual Review of Physiology, 25[1], 545–580) theory, our results show that habituation mechanisms filter the irrelevant distracting sensory input also on the basis of its temporal parameters. We conclude that habituation to onsets is controlled also by time-based expectation mechanisms and suggest that more recently proposed theories of distractors filtering should also incorporate the temporal parameter among the factors that allow an efficient handling of visual distraction.
Similar content being viewed by others
Introduction
In natural conditions, the abrupt appearance of a new salient event in the visual field is accompanied by a sudden luminance change called visual transient. Onsets are a specific class of visual transients capable of triggering a robust attentional capture (Breitmeyer & Ganz, 1976; Jonides & Yantis, 1988; Lamy & Egeth, 2003), which serves the purpose of identifying the new event and its motivational significance, or alternatively its irrelevance. Hence, whatever the activity one may be engaged in, the initial involuntary orienting toward the onset stimulus is generally a highly adaptive response allowing a rapid inspection of a potentially relevant event. Yet if the onset is irrelevant, the initially adaptive capture response becomes a source of distraction when emitted in a stereotyped fashion on subsequent onset presentations, with potentially dangerous consequences. Luckily enough, evolution has equipped the brain with learning mechanisms that allow to filter, and therefore to cease to respond to, insignificant repetitive sensory signals, a phenomenon known as habituation (Harris, 1943; Thompson, 2009).
In particular, a response that has long been shown to be subject to habituation is the orienting reflex (OR), the covert and overt orienting toward a novel or significant stimulation (Pavlov, 1927). The mechanism controlling the OR and its habituation has been described by Sokolov in his stimulus-model comparator theory (Sokolov, 1960, 1963). According to Sokolov, habituation of the OR emerges when the current sensory input matches the brain’s expectation about the upcoming stimulation, with expectation arising from the statistics of the past events. By contrast, a surprising event generates a prediction error, or put differently conveys information entropy (Shannon, 1948), which in turn triggers the OR (Sokolov et al., 2002). In agreement with the possibility that the visual distraction caused by repetitive onsets is subject to habituation, previous studies have documented that onsets interference on a discriminative task declines with repeated exposure to the irrelevant transient event (Dukewich, 2009; Turatto, Bonetti, & Pascucci, 2018a; Turatto, Bonetti, Pascucci, & Chelazzi, 2018b; Turatto & Pascucci, 2016). Furthermore, as anticipated by Sokolov’s model, habituation to onsets appears to be controlled by their statistical occurrence, with stronger distraction attenuation where onsets are more likely compared with where they are less likely (Valsecchi & Turatto, 2022), and this even when the total number of onsets presented in the two conditions is the same (Turatto & Valsecchi, 2022), which gives strong support to the role of expectation in habituation.
In addition to habituation mechanisms, other mechanisms have been proposed to handle visual distraction (Chelazzi et al., 2019; Geng et al., 2019; van Moorselaar & Slagter, 2020). A common assumption is that filtering would occur because the peak of activation triggered by the salient distractor is suppressed, either reactively or proactively, at the level of the priority map or dimension-based maps (Liesefeld & Müller, 2019; Luck et al., 2021). However, none of the more recent theories of distractor filtering takes into account that distractors appear at a certain point in time, and that this information can be used to attenuate visual distraction.
By contrast, the time of occurrence of an irrelevant sensory input is a factor that is pivotal in the habituation model proposed by Sokolov, probably because it was essentially concerned with habituation emerging from a sequential series of stimuli (Sokolov, 1960, 1963). Consequently, the time of occurrence of the distractor should be one of the features incorporated in the stimulus neural model, which is used to make predictions about the upcoming event, and ultimately to filter the stimuli that match the prediction (Sokolov et al., 2002). In line with this possibility, Xu et al. (2021) provided preliminary (although inconsistent) evidence suggesting a role of time in the rejection of feature-singleton distractors.
In light of this scenario, we decided to test the hypothesis that habituation to onsets is governed by mechanisms that make predictions and filter the distractors also on the basis of their temporal characteristics. To this aim, we exposed participants for four blocks of trials to a “standard” distractor, appearing always at the same interval from the display presentation, and predicted habituation of the corresponding capture. Then, in the last two blocks of trials, we introduced a rare “delayed” distractor, consisting of the identical onset, which, however, appeared with a 1-second delay. If habituation of onsets capture is based also on onsets timing expectation, we foresee a recovery of capture from the “delayed” onset.
Methods
Sample-size justification
The main effect of interest in the present study was a difference in the amount of capture between the standard-distractor and the delayed-distractor conditions when the delayed distractor was introduced. Capture was defined as the response time (RT) difference between distractor-present and the distractor-absent trials. Since we will introduce the delayed distractor only from Block 3 onward, in that block capture should be larger than in the standard-distractor condition, where it should be already habituated. For this reason, we expect the effect to be medium-sized (d = .5). Therefore, the critical analysis will be a paired t test between the capture effect in the standard-distractor condition and the delayed-distractor condition. An a priori power analysis was conducted by using G*Power (Faul et al., 2007), which suggested a total N = 27, with d = .5 and α = .05 to achieve a power of 80% (one-tailed).
Participants
We recruited participants via Prolific (Prolific Academic Ltd, Oxford, UK), and 29 individuals successfully participated in the experiment (Mage = 24.5 years, with an effective range of 18–33 years, six females and 23 males). The criteria for participation were set as being fluent in English language, so that the instruction would be clear, have no literacy difficulties, have normal or corrected to normal vision, and to be naïve with regard to the experiment (individuals who participated in previous similar studies were not recruited). One participant was excluded from the analyses due to low overall accuracy (<75%).
Apparatus
The experiment was programmed in PsychoPy Software (Peirce, 2007) and run online via Pavlovia Platform (Open Science Tools Limited, Nottingham, UK). Only participants who performed the task via a personal computer were allowed to perform the experiment (no tablets or smartphones were allowed).
Stimuli and procedure
Since the experiment was conducted online, each participant performed the task on their own computer, with potentially different monitor’s sizes. Therefore, to make conditions more comparable across participants, the dimensions of the stimuli were programmed so that they were scaled according to the monitor’s sizes (the experiment was run in full-screen mode). Stimuli dimensions are reported as degrees of visual angle assuming a monitor height of 34 cm and a viewing distance of 60 cm.
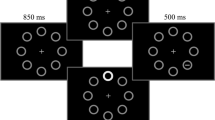
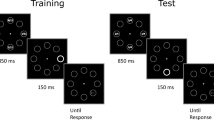
Each trial started with a white fixation cross positioned at the center of the screen (0.3° × 0.3°) and appearing over a black background. After 1,000 ms, a display appeared consisting of eight equidistant gray circles (3° diameter, 0.1° thick) positioned on an imaginary circumference with radius 10° centered on the fixation cross. In the standard-distractor condition, after that 1,000 ms were elapsed the target line (0.75° length, 0.1° thick) appeared inside one of four circles positioned along the oblique meridians. On distractor-present trials (50% rate), 150 ms before the target appearance one of the four circles positioned along the vertical or horizontal meridians became abruptly white and increased its thickness to 0.3° (see Fig. 1).

Schematic representation of the trial events in the experiment (see Methods for details). In the standard-distractor condition (100% of the trials in Blocks 1 and 2, and 90% of the trials in Blocks 3 and 4), the target appeared 1,000 ms after the initial display onset. In the delayed-distractor condition (10% of the trials in Blocks 3 and 4), the interval between the initial display and the target was increased to 2,000 ms. In both conditions, when present the distractor appeared 150 ms before the target. Hence, in Blocks 3 and 4, 90% of the distractors occurred at the expected time (850 ms), whereas 10% of the distractor occurred with an unexpected delay (1,850 ms). The participants’ task was to discriminate the orientation (vertical vs horizontal) of the target line
In the delayed-distractor condition, which was introduced only in Blocks 3 and 4 (10% of the total trials in those blocks, 5% distractor-present trials), the timing of the events was the same of the standard-distractor condition except that the target appeared 2,000 ms after the display onset, so that also the distractor onset was delayed by 1,000 ms as compared with the standard-distractor condition (850 ms vs. 1,850 ms; also see Fig. 1).
Participants were submitted to 400 trials divided into four blocks of 100 trials each, and their task was to report as quickly and as accurately as possible the orientation of the target line by pressing the down arrow of the keyboard if the line was vertical, or the right arrow if it was horizontal. The target remained on the screen for a maximum of 500 ms, but disappeared earlier for faster responses, and participants were given 1,500 ms for responding. If the response was incorrect or exceeded the time limit, a red message appeared on the screen (“Error” or “Try to be faster!”) for 800 ms. The intertrial interval was set to 500 ms.
All participants received detailed instructions concerning the task and were informed about the general aim of the experiment through the Prolific interface. They gave their consent by agreeing to be redirected to the experiment URL. Participants were paid 10 £/h for their participation, and the experiment lasted approximately 30 minutes. The experiment was carried out in accordance with the Declaration of Helsinki.
Results
The analyses were performed with custom-written scripts in MATLAB and JASP (Version 0.16.1). RTs outliers (2.2%) for correct trials (overall accuracy 94%) were identified and excluded using the procedure suggested by Cousineau and Chartier (2010). For null-hypothesis significance testing, when sphericity assumption was violated, Greenhouse–Geisser correction was applied to the degrees of freedom. Post hoc t tests were Bonferroni corrected for multiple comparisons. Bayes factors were also estimated quantifying how much more likely the data were under the alternative hypothesis than under the null hypothesis (BF10). For more than one predictor, we estimated the inclusion Bayes factor across matched models (BFincl; van den Bergh et al., 2020). Posterior odds were corrected for multiple comparisons.
As a first step, we assessed the impact of the distractor presence in the standard-distractor condition by entering RTs for correct responses into a repeated measures ANOVA with Block (1 to 4) and Distractor (present vs. absent) as within-subjects factors. The ANOVA revealed a main effect of Block, F(2.26, 61.1) = 11.6, p < .001, \({\upeta}_{\textrm{p}}^2\) = .300, BFincl = 9.00*103, a main effect of Distractor, F(1, 27) = 40.9, p < .001, \({\upeta}_{\textrm{p}}^2\) = .602, BFincl = 2.99*104, and a significant interaction, F(3, 81) = 13.4, p < .001, \({\upeta}_{\textrm{p}}^2\) = .332, BFincl = 2.54*104 (see Fig. 2a). The same analysis conducted on error rates yielded a significant effect of Block, F(1.37, 37.0) = 5.37, p = .017, \({\upeta}_{\textrm{p}}^2\) = .166, BFincl = 17.7, with errors decreasing across blocks, a significant effect of Distractor, F(1, 27) = 7.70, p = .010, \({\upeta}_{\textrm{p}}^2\) = .222, BFincl = 1.71, with distractors overall increasing the error rates (present: M = 7.0, SD = 4.2, absent: M = 5.4, SD = 3.4), but no interaction, p = .849, BFincl = 0.07.

Graphical representation of the results. a Absolute RTs as a function of distractor presence and condition (standard vs .delayed). b RT capture effect as a function of block and distractor timing. The continuous line (blue color) shows habituation of capture for the standard distractor across blocks; the dashed line (orange color) shows a recovery of capture triggered by the delayed distractor (c). Error rate capture effect as a function of block and distractor timing. d Dishabituation: capture triggered by a standard distractor in the three trials following a delayed distractor on trial n. Error bars represent SEM. (Color figure online)
An ANOVA conducted directly on the capture effect in the standard-distractor condition revealed a main effect of the factor Block (1 to 4), which resulted significant, F(3, 81) = 13.4, p < .001, \({\upeta}_{\textrm{p}}^2\) = .332, BF10 = 4.12*104. The results thus showed that in the standard-distractor condition onset capture was subject to habituation (see Fig. 2b, continuous or blue line), thus confirming previous findings with analogous paradigms (e.g., Turatto & Pascucci, 2016; Valsecchi & Turatto, 2022). The same analysis was conducted also on error rates and showed no significant effects (p = .849, BF10 = 0.07; see Fig. 2c, continuous or blue line).
The next crucial step was to test whether the introduction of the unexpected delayed distractor in Blocks 3 and 4, which was otherwise identical to the standard distractor, caused a new capture response. A repeated measures ANOVA on RTs with Distractor (present vs absent), Condition (standard vs. delayed) and Block (3 vs. 4) showed a main effect of Distractor, F(1, 27) = 35.71, p < .001, \({\upeta}_{\textrm{p}}^2\) = .569, BFincl = 8.34*103, a main effect of Condition, F(1, 27) = 4.87, p = .036, \({\upeta}_{\textrm{p}}^2\) = .153, BFincl = 1.83, a significant Distractor × Condition interaction, F(1, 27) = 7.39, p = .011, \({\upeta}_{\textrm{p}}^2\) = .215, BFincl = 4.93, but no other significant effect (Block, p = .598, BFincl = 0.271; Distractor × Block, p = .633, BFincl = 0.257; Condition × Block, p = .233, BFincl = 0.797, Distractor × Condition × Block, p = .133, BFincl = 0.952; see Fig. 2a). The same analysis on error rates revealed no significant effect (Distractor, p = .229, BFincl = 0.504; Condition, p = .059, BFincl = 0.758; Block, p = .386, BFincl = 0.258; Distractor × Condition, p = .694, BFincl = 0.306; Distractor × Block, p = .694, BFincl = 0.261; Block × Condition, p = .151, BFincl = 0.594; Distractor × Condition × Block, p = .598, BFincl = 0.248).
Again, an ANOVA on RTs conducted directly on the capture effect, that for the delayed distractor was calculated by using the corresponding distractor-absent trials, showed a main effect of Condition, F(1, 27) = 7.39, p = .011, \({\upeta}_{\textrm{p}}^2\) = .215, BFincl = 4.13, but neither an effect of Block, p = .633, BFincl = 0.289, nor a significant interaction, p = .133, BFincl = 0.845. Post hoc comparisons (t tests) confirmed that capture was significantly larger for the delayed distractor compared with the standard distractor in Block 3, t(27) = 3.10, p = .019, d = 0.707, BF10 = 9.04, but not in block 4 (p = 1, BF10 = 0.394). In addition, the amount of capture elicited by the delayed distractor in Block 3 was equivalent to that observed for the standard distractor in Block 1 (p = .843, BF10 = 0.204; see Fig. 2b, dashed or orange line). No significant results emerged from the analysis of error rates (Condition, p = .694, BFincl = 0.272; Block, p = .694, BFincl = 0.255; Condition × Block, p = .943, BFincl = 0.257; see Fig. 2c, dashed orange line).
Furthermore, we also addressed whether target processing per se was affected by our timing manipulation irrespective of any effect on distractors. Hence, RTs from distractor-absent trials only were entered into an ANOVA, with block (3 vs. 4) and target (standard vs delayed) showed no significant effects (block, p = .902, BFincl = 0.261; target, p = .704, BFincl = 0.302; Block × Target, p = .224, BFincl = 0.536; see Fig. 2a). No significant results emerged when the same analysis was performed on error rates (block, p = .742, BFincl = 0.257; target, p = .109, BFincl = 0.730; Block × Target, p = .258, BFincl = 0.563).
The results have clearly confirmed that the capture response triggered by a peripheral onset stimulus was subject to habituation. Crucially, however, we have also found that if the onset distractor was infrequently presented at an unexpected point in time, it triggered a new capture response, while the response to the standard distractor continued to remain habituated.
Test of dishabituation
One of the characteristics of habituation is the phenomenon of dishabituation—namely, the fact that the introduction of a new stimulus during an habituation phase results in a temporary recovery of the habituated response to the original stimulus when this is reintroduced (Rankin et al., 2009). In the current paradigm, the delayed distractor could have acted as a dishabituating stimulus, possibly causing a brief dishabituation of the capture response from the standard distractor, which may have been obscured by the mean block RT. To test for this possibility, we analyzed the capture response elicited by the standard distractor in the three trials following the occurrence a delayed distractor on a given trial n (because of the small number of observations available in our analysis we considered Blocks 3 and 4 pooled together). A paired-sample t test revealed that on trial n + 1 (i.e., the first standard distractor after a delayed distractor) the amount of capture elicited by the standard distractor was significantly larger than the corresponding mean capture in the pooled block, t(27) = 3.16, p = .004, d = 0.597, BF10 = 10.3, and did not differ from the mean capture of the delayed distractor in the pooled block (p = .808, BF10 = 0.206). However, this dishabituation effect was extremely short-lasting, as it vanished completely at trial n + 2 (p = .701, BF10 = 0.215), and n + 3 (p = .430, BF10 = 0.269; also see Fig. 2d).
General discussion
The results of the present experiment were clear cut and showed that although the capture triggered by a peripheral visual onset habituated, as shown previously (e.g., Pascucci & Turatto, 2015; Turatto, Bonetti, & Pascucci, 2018a; Turatto, Bonetti, Pascucci, & Chelazzi, 2018b), the capture response reemerged at full strength when, on a small proportion of trials, the onset was presented with an unexpected delay. We believe these findings have important implications both for models explaining how capture from salient distractors can be attenuated, and for the original Sokolov’s model and habituation mechanisms in general.
To begin with, on the assumption that onsets saliency is computed in the same fashion of feature-singleton distractors, the present findings show that the possibility to attenuate the onset capacity to attract attention is not only obtained by suppressing, on the basis of location or feature information, the saliency signal emerging from visual maps coding for local feature contrasts in the corresponding feature dimension (Ferrante et al., 2018; Gaspelin & Luck, 2018; Leber et al., 2016; Liesefeld & Müller, 2019; Luck et al., 2021; Stilwell et al., 2019; Wang & Theeuwes, 2018; Zelinsky & Bisley, 2015). At present, mechanisms based on this type of “static” saliency attenuation are incomplete because they do not take into account the time of occurrence of the salient element. Indeed, as we have shown, the possibility to learn the distractor temporal parameters, and therefore to generate a correct temporal expectation about its occurrence in time, is an important factor that allows to filter out the salient element and to mitigate the corresponding visual distraction. By contrast, Sokolov’s model of habituation specifically postulated that the temporal parameter of the irrelevant sensory input is a key component of the neural model of the irrelevant stimulation, which is used to attenuate the OR (Sokolov et al., 2002).
As far as the Sokolov model is concerned, it is often described as assuming that a novel stimulus evokes an OR because it does not match the prediction deriving from the model of the previous stimulations. When a new stimulus occurs, it will start to change the old model, which eventually would be more or less gradually substituted by one representing the new irrelevant iterative distractor, or the old model will be changed to also incorporate the new stimulus’ characteristics. However, as a matter of fact, our results attested that participants were captured by the delayed distractor while habituation to the standard distractor remained substantially unaltered, suggesting that to some extent the model of the standard distractor remained in place. How can this be explained?
Two scenarios are possible. The first is that more than one model of the irrelevant stimulation can be formed and held concurrently in memory, each one possibly generating more or less robust expectations depending on the corresponding distractor statistics (Turatto & Valsecchi, 2022). This possibility is also supported by the fact that different degrees of habituation have been observed for onset distractors appearing with different location probabilities (Valsecchi & Turatto, 2022). In addition, according to Sokolov the habituation model would be stored in short-term memory (Sokolov, 1960, 1963; Sokolov et al., 2002), whose capacity, at least for the relevant stimuli in the visual modality, is approximately of 4–5 elements (Luck & Vogel, 1997). This is not to say, however, that a new model is formed whenever there is a small variation in the habituating stimulus, because like any other learning process, habituation can be subject, to some extent, to generalization (Thompson, 2009). Also, the fact that multiple habituation models might be concurrently held in short-term memory does not necessarily imply a complete independence of such models, especially if some of them share some characteristics of the stimuli.
A second scenario is that in our experiment a robust model was formed only for the standard distractor, which was the only distractor appearing in the first two blocks of trials, and in 90% of cases in the last two blocks. Now, because with the exception of timing the standard distractor was identical to the delayed distractor, the rare occurrence of the latter may have been insufficient for the complete substitution of the old model with a new one. Instead, when the cognitive system detected a delayed distractor, this rare event was treated as initial evidence for a possible change in the characteristics of the standard distractor model, likely triggering a first tentative update of the model. However, given the extreme low frequency of the delayed distractor, the standard model was never really changed, and the delayed distractor continued to be an event that violated the expectation arising from the standard model, therefore capturing attention. The results of the dishabituation analysis appear in agreement with this scenario. Recall that the phenomenon of dishabituation consists in the fact that the presentation of a different stimulus generates a recovery of the habituated response to the original stimulus (Thompson, 2009). The reason would be that the presentation of a deviant stimulus alters or perturbates the previous habituating model (Steiner & Barry, 2011, 2014), so that the reoccurrence of the habituating stimulus is a bit less expected than before, which triggers again, at least partially, the OR. Our analysis showed a recovery of capture for the first standard distractor appearing after a delayed distractor, which can be seen as an instance of a very short-lasting dishabituation effect. At present it is difficult to understand which one of the two possibilities is more likely, but whatever it might be, what we have shown is that an onset occurring when expected is less distracting than an “equivalent” onset occurring when unexpected.
In our view, the recovery of capture for the delayed onset resulted from a violation of the habituation model, which generated a distractor expectation based on the standard time. However, an alternative possibility is that the capture recovery elicited by the delayed distractor was caused by the unexpected delayed target, as participants prepared an optimal attentional set for the target appearing at the standard time, and this attentional set also included the mechanisms to deal with the interference caused by the corresponding standard distractor. Hence, when the target did not appear when expected (as in Blocks 3 and 4), the attentional set was not properly configured, and consequently also the corresponding distractor filtering mechanisms. This idea differs from the one we have advocated, because we assume that the recovery of capture emerged from a violation of the standard distractor expectation per se, and not from a possible violation of the standard target set or template. However, the target attentional set hypothesis predicts a less efficient processing of the delayed target compared with the standard target, but as indicated by the analyses and the results depicted in Fig. 2a, we did not find evidence supporting this prediction. Indeed, RTs in the distractor-absent condition did not differ between the standard and delayed target. On the contrary, if anything we found a significant decrement in the error rates for the delayed target compared with the standard target when Blocks 3 and 4 were pooled together: t(27) = 2.19, p = .037, d = 0.414 (standard: M = 5.4, SD = 3.4, delayed: M = 2.9, SD = 6.6), and this while the distractor interference recovered almost completely, a pattern of findings that does not support the target-template hypothesis.
Results in agreement with those reported here have been reported by Remington et al. (1992), who showed that the amount of capture triggered by irrelevant onsets preceding the target was larger when the onsets appeared after a random interval from the blink of the fixation point compared with when, in a different experiment, the interval was fixed. More recently, Xu and colleagues have also reported evidence that in some cases the suppression of a feature-singleton distractor is more efficient when it appears at the expected point in time (Xu et al., 2021). These studies were aimed at addressing aspects of attentional capture control different from habituation to onsets, and although none of them showed a recovery of capture for a distractor appearing at an unexpected point in time, they converge on the idea that the possibility to predict the distractors time of occurrence allows to reduce the amount of capture, which is in line with our proposal.
In sum, our study documented that, likewise the correct allocation of attention in time leads to an enhanced target processing (Nobre, 2001; Nobre & van Ede, 2018), the mechanisms for distractors rejection are more efficient when they are tuned on the correct distractors timing (also, see Xu et al., 2021). If current theories of distractors filtering need to incorporate the time parameter to provide a more exhaustive picture of how distractors handling may take place, the temporal characteristics of the irrelevant sensory input are central in the model of habituation of the OR proposed by Sokolov more than 60 years ago.
Data availability
All data and codes for analyses have been made publicly available via OSF and can be accessed online (https://osf.io/n29ed/?view_only=032c68369f6848e8aa7f80990dc33512). The design and analysis plans for the experiment were not preregistered.
References
Breitmeyer, B. G., & Ganz, L. (1976). Implications of sustained and transient channels for theories of visual pattern masking, saccadic suppression, and information processing. Psychological Review, 83(1), 1–36.
Chelazzi, L., Marini, F., Pascucci, D., & Turatto, M. (2019). Getting rid of visual distractors: The why, when, how, and where. Current Opinion in Psychology, 29, 135–147.
Cousineau, D., & Chartier, S. (2010). Outliers detection and treatment: A review. International Journal of Psychological Research, 3(1), 58–67.
Dukewich, K. R. (2009). Reconceptualizing inhibition of return as habituation of the orienting response. Psychonomic Bulletin & Review, 16(2), 238–251.
Faul, F., Erdfelder, E., Lang, A.-G., & Buchner, A. (2007). G*Power 3: A flexible statistical power analysis program for the social, behavioral, and biomedical sciences. Journal of Materials and Environmental Science, 39(2), 175–191.
Ferrante, O., Patacca, A., Di Caro, V., Della Libera, C., Santandrea, E., & Chelazzi, L. (2018). Altering spatial priority maps via statistical learning of target selection and distractor filtering. Cortex, 102, 67–95.
Gaspelin, N., & Luck, S. J. (2018). The role of inhibition in avoiding distraction by salient stimuli. Trends in Cognitive Sciences, 22(1), 79–92.
Geng, J. J., Won, B.-Y., & Carlisle, N. B. (2019). Distractor ignoring: Strategies, learning, and passive filtering. Current Directions in Psychological Science, 28, 600–606.
Harris, J. D. (1943). Habituatory response decrement in the intact organism. Psychological Bulletin, 40(6), 385–422.
Jonides, J., & Yantis, S. (1988). Uniqueness of abrupt visual onset in capturing attention. Perception & Psychophysics, 43(4), 346–354.
Lamy, D., & Egeth, H. E. (2003). Attentional capture in singleton-detection and feature-search modes. Journal of Experimental Psychology: Human Perception and Performance, 29(5), 1003–1020.
Leber, A. B., Gwinn, R. E., Hong, Y., & O’Toole, R. J. (2016). Implicitly learned suppression of irrelevant spatial locations. Psychonomic Bulletin & Review, 23(6), 1873–1881.
Liesefeld, H. R., & Müller, H. J. (2019). Distractor handling via dimension weighting. Current Opinion in Psychology, 29, 160–167.
Luck, S. J., & Vogel, E. K. (1997). The capacity of visual working memory for features and conjunctions. Nature, 390(6657), 279–281.
Luck, S. J., Gaspelin, N., Folk, C. L., Remington, R. W., & Theeuwes, J. (2021). Progress toward resolving the attentional capture debate. Visual Cognition, 29(1), 1–21.
Nobre, A. C. (2001). Orienting attention to instants in time. Neuropsychologia, 39(12), 1317–1328.
Nobre, A. C., & van Ede, F. (2018). Anticipated moments: Temporal structure in attention. Nature Reviews Neuroscience, 19(1), 34–48.
Pascucci, D., & Turatto, M. (2015). The distracting impact of repeated visible and invisible onsets on focused attention. Journal of Experimental Psychology: Human Perception and Performance, 41(3), 879–892.
Pavlov, I. P. (1927). Conditioned reflexes: An investigation of the physiological activity of the cerebral cortex. Oxford University Press.
Peirce, J. W. (2007). PsychoPy—Psychophysics software in Python. Journal of Neuroscience Methods, 162(1/2), 8–13.
Rankin, C. H., Abrams, T., Barry, R. J., Bhatnagar, S., Clayton, D. F., Colombo, J., Coppola, G., Geyer, M. A., Glanzman, D. L., Marsland, S., McSweeney, F. K., Wilson, D. A., Wu, C. F., & Thompson, R. F. (2009). Habituation revisited: An updated and revised description of the behavioral characteristics of habituation. Neurobiology of Learning and Memory, 92(2), 135–138.
Remington, R. W., Johnston, J. C., & Yantis, S. (1992). Involuntary attentional capture by abrupt onsets. Perception & Psychophysics, 51(3), 279–290.
Shannon, C. E. (1948). A mathematical theory of communication. Bell System Technical Journal, 27, 379–423.
Sokolov, E. N. (1960). Neural model and the orienting influence. In M. A. Brazier (Ed.), The central nervous system and behavior. Macy Foundation.
Sokolov, E. N. (1963). Higher nervous functions: The orienting reflex. Annual Review of Physiology, 25(1), 545–580.
Sokolov, E. N., Spinks, J., Naatanen, R., & Lyytinen, H. (2002). The orienting response in information processing. Erlbaum. https://doi.org/10.4324/9781410601490
Steiner, G. Z., & Barry, R. J. (2011). Exploring the mechanism of dishabituation. Neurobiology of Learning and Memory, 95(4), 461–466.
Steiner, G. Z., & Barry, R. J. (2014). The mechanism of dishabituation. Frontiers in Integrative Neurosciences, 8, 1–8.
Stilwell, B. T., Bahle, B., & Vecera, S. P. (2019). Feature-based statistical regularities of distractors modulate attentional capture. Journal of Experimental Psychology: Human Perception and Performance, 45(3), 419–433.
Thompson, R. F. (2009). Habituation: A history. Neurobiology of Learning and Memory, 92(2), 127–134.
Turatto, M., & Pascucci, D. (2016). Short-term and long-term plasticity in the visual-attention system: Evidence from habituation of attentional capture. Neurobiology of Learning and Memory, 130, 159–169.
Turatto, M., & Valsecchi, M. (2022). Habituation to onsets is controlled by spatially selective distractor expectation. Journal of Experimental Psychology: Human Perception & Performance. https://doi.org/10.5281/zenodo.7115824
Turatto, M., Bonetti, F., & Pascucci, D. (2018a). Filtering visual onsets via habituation: A context-specific long-term memory of irrelevant stimuli. Psychonomic Bulletin & Review, 25(3), 1028–1034.
Turatto, M., Bonetti, F., Pascucci, D., & Chelazzi, L. (2018b). Desensitizing the attention system to distraction while idling: A new latent learning phenomenon in the visual attention domain. Journal of Experimental Psychology: General, 147(12), 1827–1850.
Valsecchi, M., & Turatto, M. (2022). Habituation to abrupt-onset distractors with different spatial occurrence probability. Attention, Perception, & Psychophysics. https://doi.org/10.3758/s13414-022-02531-1
van den Bergh, D., van Doorn, J., Marsman, M., Draws, T., van Kesteren, E.-J., Derks, K., Dablander, F., Gronau, Q. F., Kucharský, Š., Gupta, A. R. K. N., Sarafoglou, A., Voelkel, J. G., Stefan, A., Ly, A., Hinne, M., Matzke, D., & Wagenmakers, E.-J. (2020). A tutorial on conducting and interpreting a Bayesian ANOVA in JASP. L’Année Psychologique, 120(1), 73–96.
van Moorselaar, D., & Slagter, H. A. (2020). Inhibition in selective attention. Annals of the New York Academy of Sciences, 1464, 204–221.
Wang, B., & Theeuwes, J. (2018). Statistical regularities modulate attentional capture. Journal of Experimental Psychology: Human Perception and Performance, 44(1), 13–17.
Xu, Z., Los, S. A., & Theeuwes, J. (2021). Attentional suppression in time and space. Journal of Experimental Psychology: Human Perception and Performance, 47(8), 1056–1062.
Zelinsky, G. J., & Bisley, J. W. (2015). The what, where, and why of priority maps and their interactions with visual working memory. Annals of the New York Academy of Sciences, 1339(1), 154–164.
Code availability
Not applicable.
Funding
This study was supported by funds to Massimo Turatto from the Center for Mind/Brain sciences (CIMeC) of the University of Trento.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Ethics approval
Given that participants were adult people recruit online, and that our stimuli were motivational and emotional irrelevant stimuli consisting of a white line and white circles, we did not request any ethics approval. However, the experiment was carried out in accordance with the Declaration of Helsinki.
Consent to participate and for publication
Participants gave their consent by agreeing to be redirected to the experiment URL.
Conflicts of interest
The authors declare that they have no conflict of interest
Additional information
Open practices statement
All data and codes for analyses have been made publicly available via OSF and can be accessed online (https://osf.io/n29ed/?view_only=032c68369f6848e8aa7f80990dc33512).
The design and analysis plans for the experiment were not preregistered.
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Turatto, M., De Tommaso, M. Ignoring visual distractors: Habituation to onsets is driven by time-based expectation. Psychon Bull Rev 30, 1020–1027 (2023). https://doi.org/10.3758/s13423-022-02204-y
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.3758/s13423-022-02204-y