
Effects of Varicella-Zoster Virus Glycoprotein E Carboxyl-Terminal Mutation on mRNA Vaccine Efficacy


Institute of Medical Biology, Chinese Academy of Medical Sciences and Peking Union Medical College, Kunming 650118, China
*
Author to whom correspondence should be addressed.
†
Authors contributed equally to this work.
Vaccines 2021, 9(12), 1440; https://doi.org/10.3390/vaccines9121440
Submission received: 12 November 2021
/
Revised: 25 November 2021
/
Accepted: 3 December 2021
/
Published: 7 December 2021
(This article belongs to the Special Issue Varicella Zoster Virus: Recent Advances, Risks and Prevention in Infants and Adults in Particular (during Immunomodulating Therapy))
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Glycoprotein E (gE) is one of the most abundant glycoproteins in varicella-zoster virus and plays pivotal roles in virus replication and transmission between ganglia cells. Its extracellular domain has been successfully used as an antigen in subunit zoster vaccines. The intracellular C-terminal domain was reported to be decisive for gE trafficking between the endoplasmic reticulum, trans-Golgi network and endosomes and could influence virus spread and virus titers. Considering that the trafficking and distribution of mRNA vaccine-translated gE may be different from those of gE translated against the background of the viral genome (e.g., most gE in virus-infected cells exists as heterodimers with another glycoprotein, gI,), which may influence the immunogenicity of gE-based mRNA vaccines, we compared the humoral and cellular immunity induced by LNP-encapsulated mRNA sequences encoding the whole length of gE, the extracellular domain of gE and a C-terminal double mutant of gE (mutant Y569A with original motif AYRV, which targets gE to TGN, and mutants S593A, S595A, T596A and T598A with the original motif SSTT) that were reported to enhance virus spread and elevate virus titers. The results showed that while the humoral and cellular immunity induced by all of the mRNA vaccines was comparable to or better than that induced by the AS01B-adjuvanted subunit vaccines, the C-terminal double mutant of gE showed stable advantages in all of the indicators tested, including gE-specific IgG titers and T cell responses, and could be adopted as a candidate for both safer varicella vaccines and effective zoster vaccines.
1. Introduction
Ionizable lipid nanoparticle (LNP)-encapsulated mRNA vaccines have been approved by the FDA within one year of their development to prevent coronavirus disease 2019 (COVID-19), which is caused by severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) [1,2]. In addition to many other advantages, such as rapid development due to streamlined processes, low cost due to in vitro transcription and absence of antigen purification, SARS-CoV-2 mRNA vaccines have also shown a much better protection rate than other existing vaccine forms. In addition to the innate immune mobilization ability of mRNA itself for mRNA vaccines, its outstanding performance is also attributed substantially to the unique mechanism of intracellular translation of antigens: the protein translated from the mRNA that enters the cell of the vaccinated person has a high fidelity of posttranslational modifications, such as glycosylation, which is vital for the correct spatial structure of protein antigens; encoded and secreted protein antigens can be processed and presented by major histocompatibility complex (MHC) class II to stimulate CD4+ T helper cells to induce humoral immunity. Most importantly, cell-mediated immunity (CMI), which is less efficiently induced by subunit or inactivated forms of antigens and is highly dependent on the limited available adjuvants, could also be achieved incidentally by the self-adjuvant character of mRNA itself and the mechanism of mRNA vaccine antigen production: the protein antigens translated by mRNA in the cytoplasm could be fully processed into polypeptides and presented to MHC I as heterologous antigens produced by viral infection, which will activate CD8+ cytotoxic T lymphocytes that execute cellular immunity to selectively eliminate cells that express foreign antigens, such as virus-infected host cells [3].
CMI plays decisive roles in the efficacy of zoster vaccines [4,5,6,7,8,9]. As its name indicates, varicella-zoster virus (VZV) causes two different diseases: varicella/chicken pox upon primary infection and zoster/shingles when latent viruses in the sensory ganglia reactivate [10]. In fact, nearly every individual comes into contact with VZV before adulthood because of its high infectivity. While attenuated vaccines based on the Oka strain could effectively prevent infection with wild strains that cause varicella through humoral immunity, these vaccine strains could also lurk in the sensory ganglia and reactivate as wild strains do to cause zoster when the immune system is senescent (e.g., aging) or compromised (e.g., during infection by HIV or clinical immunosuppressive treatment) [11,12,13,14]. While pre-existing cellular immune responses upon primary VZV exposure or varicella vaccination could be boosted by the Oka strain at a dose as high as 19,400 plaque-forming units (PFU, compared with 1000–10,000 PFU in varicella vaccines), the attenuated zoster vaccine ZOSTVAXR (developed by Merk in 2005) showed a much lower protection rate than the VZV glycoprotein E (gE) and AS01B adjuvanted subunit vaccine ShingrixTM (developed by GSK in 2017) [15,16,17,18,19,20]. Considering the high price (approximately USD 150–200 per dose and two doses needed) of ShingrixTM, which is attributed mainly to the limited supply of its adjuvant component QS21, more economical zoster vaccines without a production limit are still needed, and mRNA vaccines with the advantages mentioned above are potential candidates [21,22].
In fact, a mRNA form of the zoster vaccine has been tested in nonhuman primates and showed comparable efficacy to that induced by VZV gE and AS01B adjuvanted subunit vaccines [23]. gE is one of the most abundant glycoproteins in VZV and plays pivotal roles in virus replication and transmission between ganglia cells [24,25,26,27]. As a highly glycosylated type I membrane protein, it frequently travels between the endoplasmic reticulum (ER), trans-Golgi network (TGN) and endosomes [28,29,30]. Although the extracellular domain was used as an antigen in ShingrixTM subunit vaccines, mainly to lower the difficulties of protein purification, the intracellular carboxyl terminus of gE was decisive for gE trafficking and should be evaluated in mRNA vaccines, which depend on antigen production in vivo [31,32,33]. Considering that the trafficking and distribution of mRNA vaccine-translated gE may be different from those translated against the background of the viral genome (e.g., most gE in virus-infected cells exists as heterodimers with another glycoprotein, gI), we compared the immunity induced by LNP-encapsulated mRNA vaccines encoding gE with several C-terminal domain mutations that were reported to affect the translocation of gE in different parts of cells, hoping to find a more suitable gE sequence that induces better CMI for zoster vaccines.
2. Materials and Methods
2.1. Preparation of Vaccines
DNA sequences encoding the whole length of gE (gE, 623 aa), the extracellular domain (1–538 aa) of gE (gE-E) and the C-terminal double mutant (mutant Y569A with original motif AYRV, which targets gE to TGN, and mutants S593A, S595A, T596A and T598A with original motif SSTT, which targets gE to the TGN or plasma membrane) of gE (gE-M) were codon-optimized and synthesized with 5′ and 3′ untranslated regions (Sangon Biotech Co., Ltd., Shanghai, China) [23,31,34,35]. After linearization, T7 polymerase-mediated transcription was performed using an mRNA synthesis kit (APEXBIO Technology, Houston, TX, USA), and the product was purified with MonarchR RNA purification columns (NEW ENGLAND BioLabs Inc., Ipswich, MA, USA).
LNP vaccines were prepared using a modified procedure previously described for mRNA vaccines [36,37,38]. Briefly, lipids (from AVT Pharmaceutical Technology Co., Ltd., Shanghai, China) were dissolved in ethanol at molar ratios of 50:10:37.5:2.5 (MC3: DSPC: cholesterol: DMG-PEG2000). The lipid mixtures were combined with 100 mM citrate buffer (pH 4.0) containing the above mRNA at a ratio of 3:1 with a microfluidic mixer (Precision Nanosystems, Inc., Vancouver, BC, Canada). Formulations were dialyzed against PBS, concentrated with a centrifugal filtration tube (Millipore, Tullagreen, Carrigtwohill, Co. Cork, Ireland), passed through a 0.22 μm syringe filter (PALL) and stored at 4 °C until use. Particle sizes were tested with a Zetasizer Nano ZS particle size analyzer (Malvern Panalytical, Malvern, UK). Loaded mRNA was detected with both a 1% denatured agarose gel and a Quant-iTTM RiboGreenR RNA Reagent Kit (Thermo Fisher, Eugene, OR, USA). The encapsulation efficiency was calculated as the amount of loaded nucleic acids detected compared with the amount of initial nucleic acids input in citrate buffer.
2.2. Animal Studies
Six-week-old female specific pathogen-free (SPF) C57BL/6N mice (15–18 g) were purchased from Vital River Laboratory Animal Technology Ltd. (Beijing, China), randomly divided into groups of 6 mice each (n = 6), maintained under SPF conditions and housed with free access to food and water at the Central Animal Services of the Institute of Medical Biology, Chinese Academy of Medical Sciences (IMB, CAMS). Polyinosinic–polycytidylic acid (Poly I:C, from InvivoGen, Inc., San Diego, CA, USA) and alum (Thermo Fisher, Eugene, OR, USA) mixtures or AS01B (from GSK, MD, USA)-adjuvanted extracellular domain of gE were used as controls, and PBS was used as a blank control. The mice were immunized intramuscularly in the thigh muscle twice with 50 μL of immunogen at 3-week intervals. After anesthetization by intraperitoneal injection of tribromoethanol, blood samples (via cardiac puncture) and spleens were collected 2 weeks after the final immunization for further analysis.
2.3. Enzyme-Linked Immunosorbent Assay (ELISA) of Antibody Titers
After clotting at 4 °C overnight, serum was collected after centrifugation at 3000 rpm for 10 min. gE-specific IgG titers were detected by ELISA, as previously described [39].
2.4. Enzyme-Linked Immunospot Assay (ELISPOT) of Splenocytes
Female spleens were dispersed with a 70 μm cell strainer (BD, USA). After red cell lysis by ammonium–chloride–potassium (ACK) lysis buffer at room temperature for 5 min, splenocytes were resuspended in Roswell Park Memorial Institute (RPMI) 1640 medium with 10% v/v fetal bovine serum (both from Biological Industries, Israel) and penicillin–streptomycin (Thermo Fisher) at a final concentration of 3 × 106 cells/mL. Then, 100 μL of cells was added to each well of a 96-well plate (Corning, USA) for further analysis with an ELISPOT assay kit (BD) according to the manufacturer’s protocol. The extracellular domain of gE expressed in Chinese hamster ovary cells (supplied by AtaGenix Laboratory Co., Ltd., Wuhan, China) at 20 μg/mL was added to stimulate gE-specific T cell responses by incubation overnight. Spots were counted with an ELISPOT reader system (Autoimmun Diagnostika GmbH, Germany) after immunoimaging.
2.5. Flow Cytometry
The splenocytes prepared as described above were also analyzed by flow cytometry to determine the proportion of activated or memory T cells [40]. Briefly, 10 μg/mL extracellular domain of gE was incubated for 2 h, and brefeldin A was added and incubated overnight to block cytokine release. After the addition of 5 μg/mL CD16/CD32 antibodies to block the nonspecific binding of Fc receptors by incubation at 4 °C for 10 min, PerCP-tagged anti-mouse CD4, FITC-tagged anti-mouse CD8a, Brilliant Violet 421-tagged anti-mouse CD62L and Brilliant Violet 510-tagged anti-mouse CD44 antibodies were added and incubated at 4 °C for another 30 min. After washing with permeabilization wash buffer, PE-tagged anti-mouse IFN-γ and APC-tagged anti-mouse IL-2 antibodies were added and incubated in the dark at room temperature for 30 min. Cells were gated (FSC/SSC), and samples with more than 20,000 events of CD4+ or CD8+ T cells were analyzed with a CytoFLEX flow cytometer (Beckman, Indianapolis, IN, USA) and FlowJo software (BD, USA).
2.6. Statistical Analysis
Data were analyzed with one-way analysis of variance (ANOVA) followed by Dunnett’s multiple comparisons test, with the gE-M mRNA vaccine group as the control. GraphPad Prism 8.0 (GraphPad Software Inc., La Jolla, CA. USA) was used for statistical analyses.
3. Results
3.1. LNPs Efficiently Encapsulated mRNA Antigens with Uniform Particle Sizes
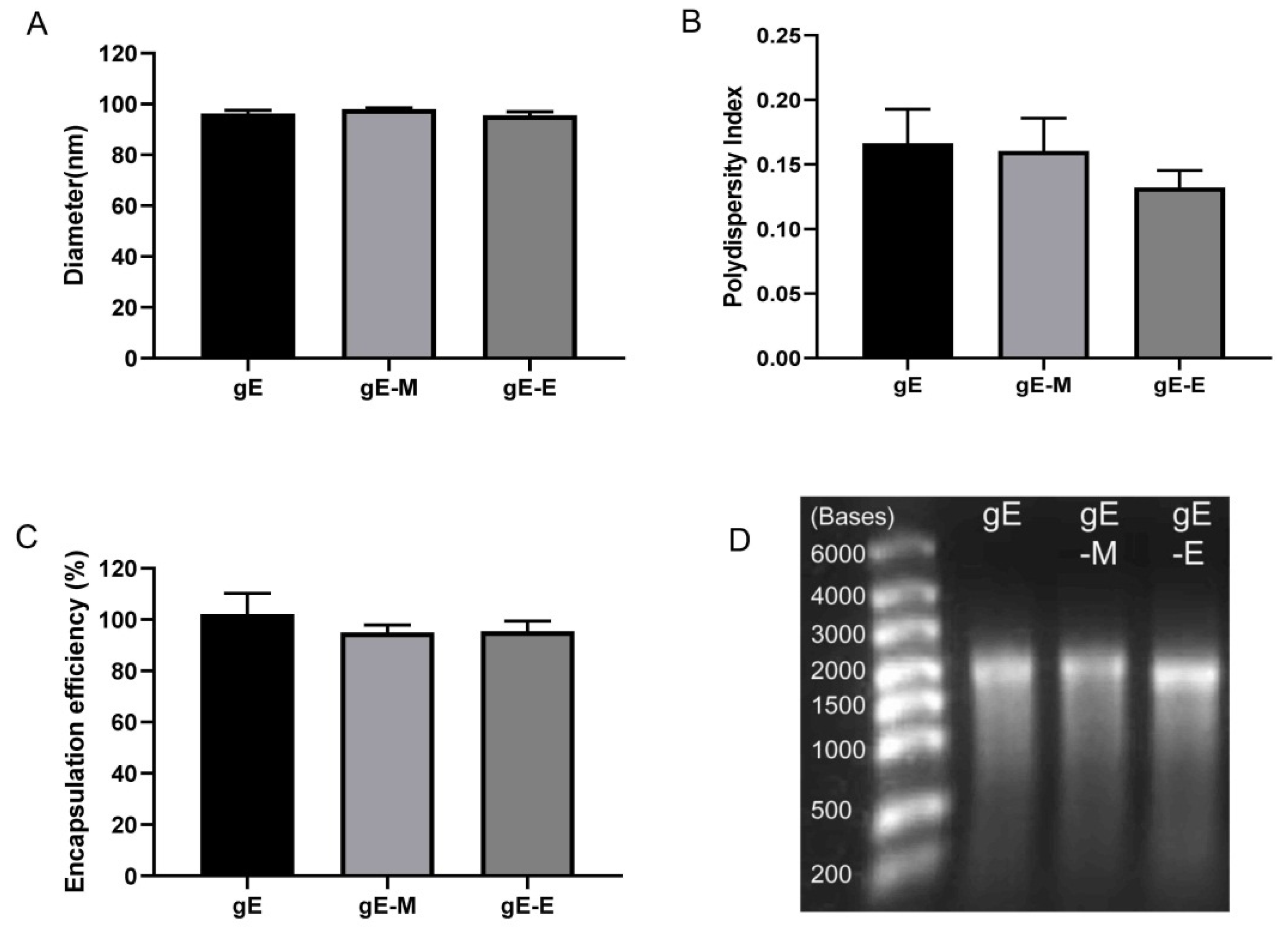
The diameters of LNPs encapsulating gE, gE-M and gE-E mRNA were 96.30 nm, 97.92 nm and 95.52 nm, respectively (Figure 1A). All of the polydispersity indices (PDIs), which are measures of the heterogeneity of a sample based on size, were lower than 0.2 (0.166 for gE, 0.160 for gE-M and 0.132 for gE-E, Figure 1B), which indicates good uniformity of the particle sizes. When 100 μg of mRNA was added to 1.5 mL of citrate buffer as the raw material for 12 doses of LNP vaccine, the encapsulation efficiencies were all above 94% (102.05% for gE, 94.91% for gE-M and 95.45% for gE-E, Figure 1C). All of these encapsulated mRNAs showed good integrity on denatured agarose gel as bands at approximately 2000 bases, with gE-E being slightly lower (Figure 1D).
3.2. mRNA Vaccines with C-terminal Mutations (gE-M) Induced the Most Potent Humoral Immune Responses
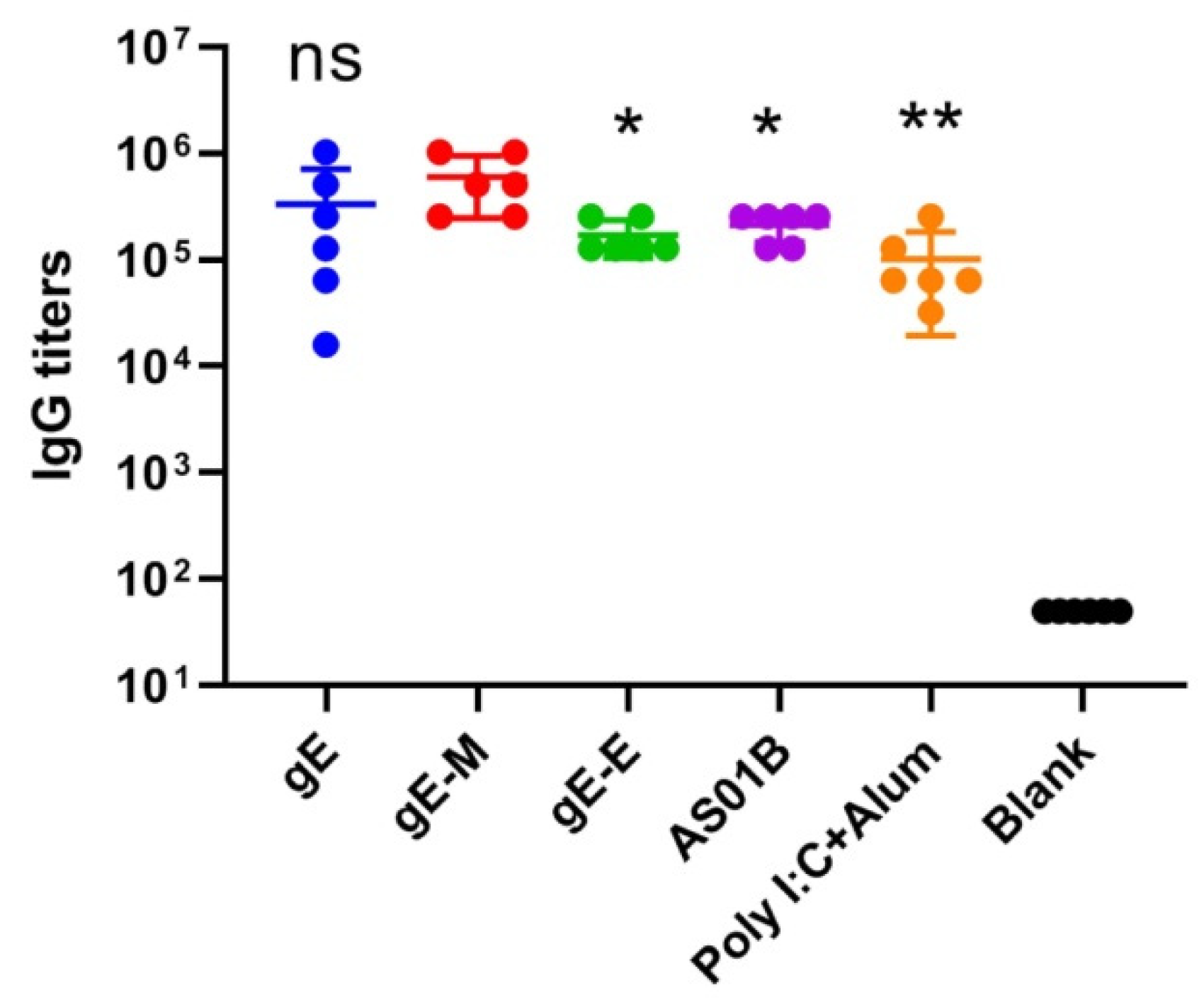
For the two control subunit vaccines, each dose of the AS01B group contained 5 μg of gE and 50 μL of AS01B adjuvants, which equals 1/10 dose of SingrixTM, and each dose of the Poly I:C+Alum group contained 10 μg of gE and 15 μg of Poly I:C. The gE-specific IgG titers of the AS01B group (213,333) were approximately two times those of the Poly I:C+Alum group (101,333) and higher than those of the gE-E group (170,667), but the mRNA vaccine with C-terminal mutations (gE-M group) showed the highest gE-specific IgG titers (597,333), which were approximately two times those of the whole gE group (gE, 333,333) and three times those of the AS01B group (Figure 2, p < 0.05).
3.3. C-Terminal Mutations (gE-M) Were Slightly Helpful in Inducing CMI in mRNA Vaccines
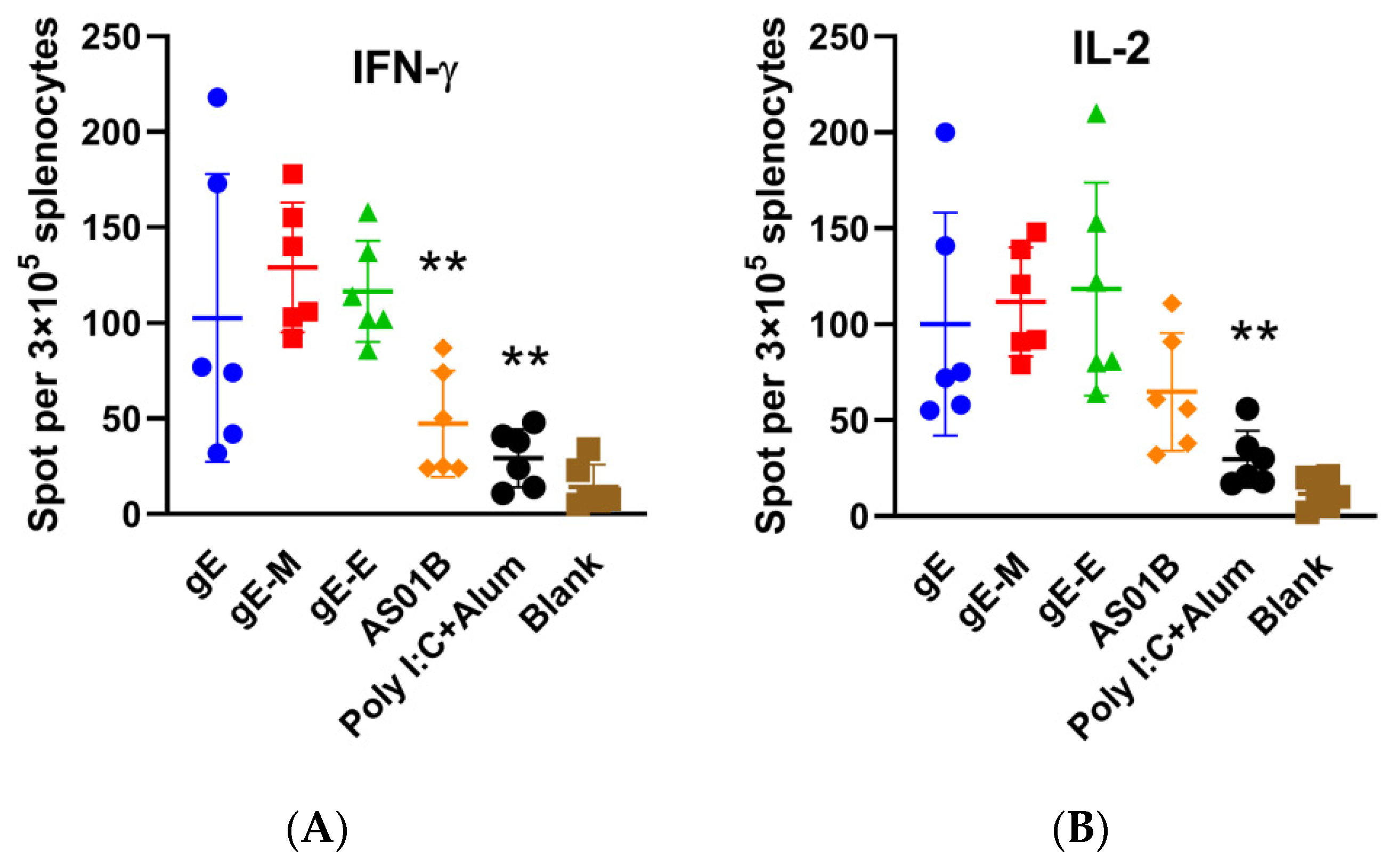
ELISPOT analysis showed that the mRNA vaccine with C-terminal mutations (gE-M group) showed the most IFN-γ-producing splenocytes (129 spots per 3 × 105 splenocytes in Figure 3A) among all three mRNA vaccines (116.5 for gE-E and 102.7 for gE) after gE stimulation, and the value was significantly higher than that of the AS01B group (47.3, p < 0.01). For gE-specific IL-2-producing splenocytes (Figure 3B), the gE-M group (spots of 111.7) had a slightly higher value than the gE group (spots of 100.2) but a slightly lower value than the gE-E group (spots of 118.3). Considering the higher outlier data in the gE-E group, we conclude that C-terminal mutations may still be the most potent in inducing CMI in mRNA vaccines.
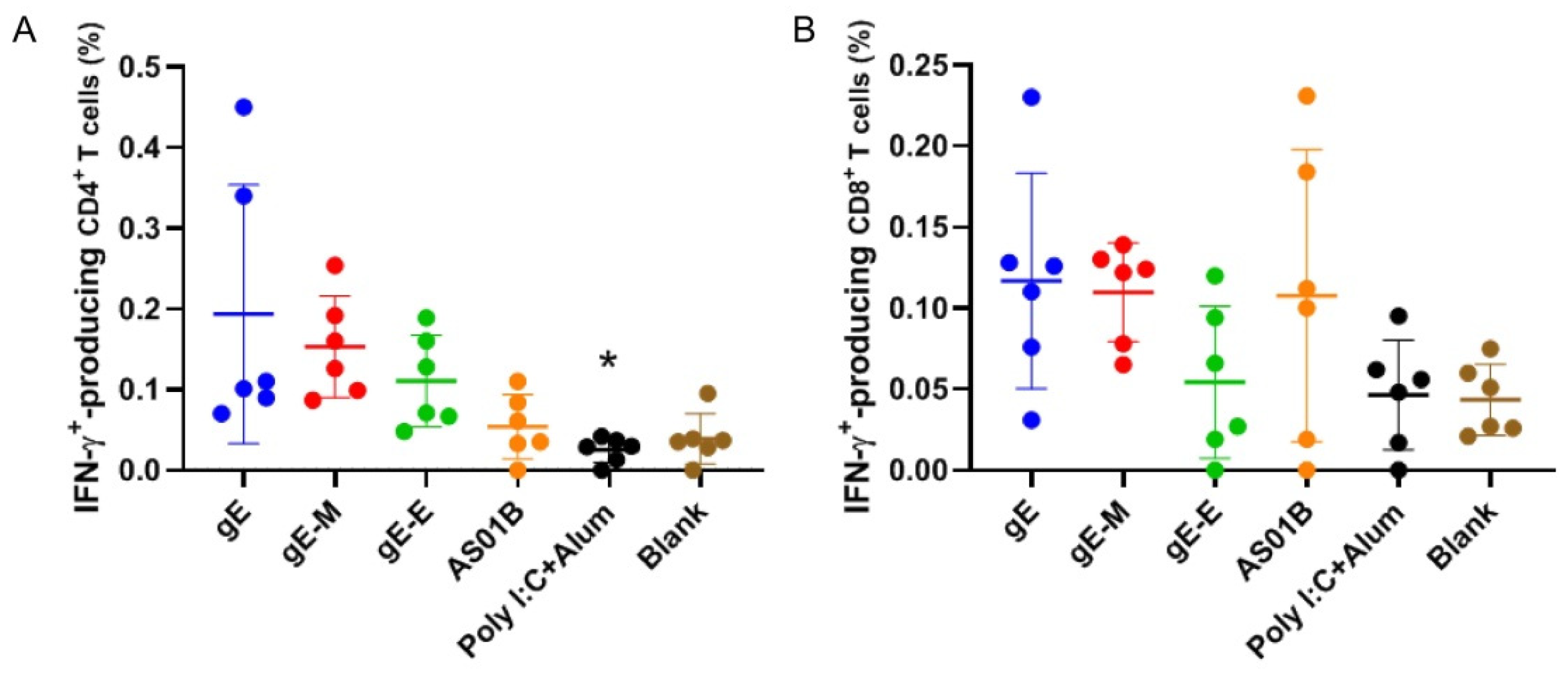
Similar conclusions could be drawn from the results of the flow cytometry analysis (Figure 4). While the gE-M group showed higher proportions of IFN-γ-producing CD4+ (Figure 4A) and IFN-γ-producing CD8+ T cells (Figure 4B) compared with the gE-E group, these values were slightly lower than those of the gE group concerning IFN-γ-producing CD4+ (0.153% versus 0.194%) and IFN-γ-producing CD8+ (0.110% versus 0.117%) T cells when the higher outlier data in the gE group were included.
3.4. C-Terminal Mutations (gE-M) Were Slightly Helpful to Elevate T Cell Memory in mRNA Vaccines
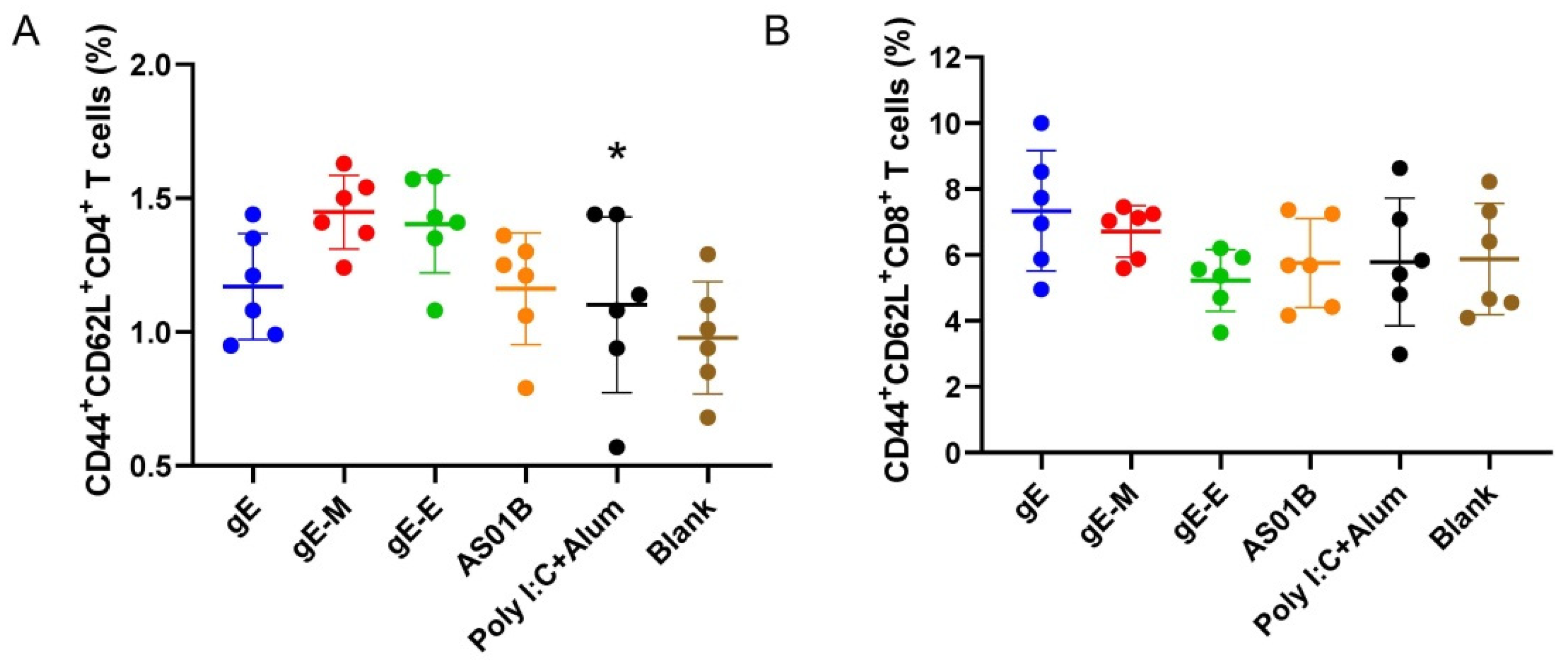
Antigen-experienced (CD44+) T cells responsible for central memory (CD62L+) were analyzed by flow cytometry [41,42]. Although the gE-M group showed the highest proportion of CD4+ central memory cells (1.45%) among all the vaccine groups (Figure 5A), a significant difference was detected only compared with the Poly I:C+Alum group (1.10%, p < 0.05). For CD8+ central memory cells, the gE group was the highest (7.34%), followed by the gE-M group (6.72%), but no significant difference existed between gE-M and any other group (Figure 5B).
4. Discussion
As one of the most abundant glycoproteins with conserved neutralization epitopes and T cell epitopes, gE is essential for VZV replication and transmission between ganglia cells [24,25,26,27,43]. As a subunit antigen, the extracellular domain has shown potential as a safe varicella vaccine that could avoid the latency of viruses from live-attenuated vaccines, which may reactivate as herpes zoster, and has potential as a zoster vaccine with suitable adjuvants that induce powerful CMI [9,36,44,45]. Although the subunit vaccine ShingrixTM with the extracellular domain of gE adjuvanted with the AS01B system showed an excellent protection rate against herpes zoster, it was not reported for use as a safer varicella vaccine, mainly because of the limited supply and infeasible synthesis of the key components of AS01B, i.e., the polysaccharide mixture QS21, which is extracted from the bark of Quillaja saponaria, which is distributed in the temperate regions of South America [21,22].
LNP-encapsulated mRNA vaccines could induce both humoral immunity and CMI by the in vivo translation of protein antigens and their self-adjuvant character [3]. Theoretically, the extracellular domain of VZV gE could be used as a corresponding mRNA vaccine. Notably, the intracellular carboxyl terminus of gE was found to be decisive for gE trafficking between the ER, TGN and endosomes [28,29,30]. Considering that the distribution of gE after translation may influence the final presentation pathway of gE as antigens, whether carboxyl-terminal mutations may influence the final immunity of mRNA vaccines should be evaluated in pursuit of better immunity [23].
When encapsulated in LNPs, all mRNA sequences, including the extracellular domain (1–538 aa) of gE, that were used in subunit vaccines to lower the difficulties of protein purification (gE-E), the whole sequence of gE (gE, 623 aa) and C-terminal double mutants (mutant Y569A and mutants S593A, S595A, T596A and T598A) of gE (gE-M), which were reported to enhance virus spread and increase virus titers against the background of the viral genome, showed uniform characteristics, including diameter, PDI and encapsulation efficiency (Figure 1). Two weeks after two intramuscular immunizations at 3-week intervals at a dose of approximately 7.5 μ/dose, gE-M induced the most potent humoral immune responses, showing the highest gE-specific IgG titers (Figure 2), which indicated that gE-M is usable as a varicella vaccine. gE-M also showed stable advantages in all of the indicators tested for CMI that play decisive roles in the efficacy of zoster vaccines, including gE-specific IFN-γ/IL-2-producing splenocyte numbers analyzed by ELISPOT (Figure 3) and the proportion of gE-specific IFN-γ-producing CD4+/CD8+ T cells among splenocytes analyzed by flow cytometry analysis (Figure 4) [4,5,6,7,8,9].
Although not statistically significant, the gE-specific IFN-γ produced CD4+ T cells, which are more frequently adopted than CD8+ T cells as good indicators for the potential of zoster vaccines in animal experiments and clinical trials, at approximately 2.84 times the quantity of the AS01B adjuvanted subunit vaccines, which was consistent with the result of a 573 aa carboxyl-terminal truncated and Y569A-mutated gE mRNA vaccine tested in nonhuman primates [17,23,44]. Because we adopted nearly identical nucleic acid sequences derived from the whole length of gE for each mRNA vaccine, the advantages of gE-M-induced immunity may be attributed mainly to the influence of carboxyl-terminal mutations and the distribution of translated protein antigens, as observed against the background of the viral genome, which ultimately manifested as rapid virus spread and higher virus titers [31]. Interestingly, gE-M also showed stable advantages in both CD4+ and CD8+ T cell memory indicators, as shown by a higher proportion of antigen-experienced (CD44+) central memory (CD62L+) T cells among splenocytes observed by flow cytometry analysis (Figure 5).
5. Conclusions
In conclusion, while mRNA vaccines encoding the whole length of gE, the extracellular domain of gE and a C-terminal double mutant sequence of gE all showed comparable or better humoral and cellular immunity to the AS01B adjuvanted subunit vaccines, the vaccines encoding double mutants at the C-terminus, which were reported to enhance virus spread and higher virus titers against the background of the viral genome, showed stable advantages in all of the indicators tested for humoral immunity and CMI and could be adopted as both safe varicella and effective zoster mRNA vaccine candidates.
Author Contributions
Conceptualization, C.L.; Data curation, H.C. and Y.W.; Investigation, H.C., Y.W., N.L. and K.L.; Project administration, C.L.; Supervision, C.L. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the CAMS Initiative for Innovative Medicine (grant number 2021-I2M-1-043), the Major Science and Technology Special Projects of Yunnan Province, China (grant number 202002AA100009), the China Health and Longevity Innovation Competition (2021-JKCS-012), the Special Biomedicine Projects of Yunnan Province (202102AA310035), the Basic Research Projects of Yunnan Province (202101AT070286), the Funds for the Training of High-Level Health Technical Personnel in Yunnan Province (grant number H-2019063) and the Funds for High-Level Scientific and Technological Talents Selection Special Project of Yunnan Province.
Institutional Review Board Statement
Animal experiments were approved by the Ethics Committee of Animal Care and Welfare of the Institute of Medical Biology, Chinese Academy of Medical Sciences and Peking Union Medical College (IMB, CAMS) (permit number SYXK (dian) K2019-00023), in accordance with the animal ethics guideline of the Chinese National Health and Medical Research Council and the Office of Laboratory Animal Management of Yunnan Province, China. All efforts were made to minimize animal suffering.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data used during the study are available from the corresponding author by request.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Baden, L.R.; El Sahly, H.M.; Essink, B.; Kotloff, K.; Frey, S.; Novak, R.; Diemert, D.; Spector, S.A.; Rouphael, N.; Creech, C.B.; et al. Efficacy and Safety of the mRNA-1273 SARS-CoV-2 Vaccine. N. Engl. J. Med. 2021, 384, 403–416. [Google Scholar] [CrossRef]
- Polack, F.P.; Thomas, S.J.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Perez, J.L.; Perez Marc, G.; Moreira, E.D.; Zerbini, C.; et al. Safety and Efficacy of the BNT162b2 mRNA Covid-19 Vaccine. N. Engl. J. Med. 2020, 383, 2603–2615. [Google Scholar] [CrossRef]
- Xu, S.; Yang, K.; Li, R.; Zhang, L. mRNA Vaccine Era-Mechanisms, Drug Platform and Clinical Prospection. Int. J. Mol. Sci. 2020, 21, 6582. [Google Scholar] [CrossRef]
- Asada, H. VZV-specific cell-mediated immunity, but not humoral immunity, correlates inversely with the incidence of herpes zoster and the severity of skin symptoms and zoster-associated pain: The SHEZ study. Vaccine 2019, 37, 6776–6781. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, P.B.; Gabriel, E.E.; Miao, X.; Li, X.; Su, S.C.; Parrino, J.; Chan, I.S. Fold rise in antibody titers by measured by glycoprotein-based enzyme-linked immunosorbent assay is an excellent correlate of protection for a herpes zoster vaccine, demonstrated via the vaccine efficacy curve. J. Infect. Dis. 2014, 210, 1573–1581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haberthur, K.; Engelmann, F.; Park, B.; Barron, A.; Legasse, A.; Dewane, J.; Fischer, M.; Kerns, A.; Brown, M.; Messaoudi, I. CD4 T cell immunity is critical for the control of simian varicella virus infection in a nonhuman primate model of VZV infection. PLoS Pathog. 2011, 7, e1002367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steain, M.; Sutherland, J.P.; Rodriguez, M.; Cunningham, A.L.; Slobedman, B.; Abendroth, A. Analysis of T cell responses during active varicella-zoster virus reactivation in human ganglia. J. Virol. 2014, 88, 2704–2716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weinberg, A.; Levin, M.J. VZV T cell-mediated immunity. Curr. Top. Microbiol. Immunol. 2010, 342, 341–357. [Google Scholar] [CrossRef] [PubMed]
- Wui, S.R.; Kim, K.S.; Ryu, J.I.; Ko, A.; Do, H.T.T.; Lee, Y.J.; Kim, H.J.; Lim, S.J.; Park, S.A.; Cho, Y.J.; et al. Efficient induction of cell-mediated immunity to varicella-zoster virus glycoprotein E co-lyophilized with a cationic liposome-based adjuvant in mice. Vaccine 2019, 37, 2131–2141. [Google Scholar] [CrossRef]
- Zerboni, L.; Sen, N.; Oliver, S.L.; Arvin, A.M. Molecular mechanisms of varicella zoster virus pathogenesis. Nat. Rev. Microbiol. 2014, 12, 197–210. [Google Scholar] [CrossRef] [Green Version]
- Arruti, M.; Pineiro, L.D.; Salicio, Y.; Cilla, G.; Goenaga, M.A.; Lopez de Munain, A. Incidence of varicella zoster virus infections of the central nervous system in the elderly: A large tertiary hospital-based series (2007–2014). J. Neurovirol. 2017, 23, 451–459. [Google Scholar] [CrossRef] [PubMed]
- Breuer, J. Molecular Genetic Insights into Varicella Zoster Virus (VZV), the vOka Vaccine Strain, and the Pathogenesis of Latency and Reactivation. J. Infect. Dis. 2018, 218, S75–S80. [Google Scholar] [CrossRef] [PubMed]
- Krause, P.R.; Klinman, D.M. Varicella vaccination: Evidence for frequent reactivation of the vaccine strain in healthy children. Nat. Med. 2000, 6, 451–454. [Google Scholar] [CrossRef] [PubMed]
- Levin, M.J.; Bresnitz, E.; Popmihajlov, Z.; Weinberg, A.; Liaw, K.L.; Willis, E.; Curtis, J.R. Studies with herpes zoster vaccines in immune compromised patients. Expert Rev. Vaccines 2017, 16, 1217–1230. [Google Scholar] [CrossRef]
- Chlibek, R.; Bayas, J.M.; Collins, H.; de la Pinta, M.L.; Ledent, E.; Mols, J.F.; Heineman, T.C. Safety and immunogenicity of an AS01-adjuvanted varicella-zoster virus subunit candidate vaccine against herpes zoster in adults ≥50 years of age. J. Infect. Dis. 2013, 208, 1953–1961. [Google Scholar] [CrossRef] [Green Version]
- Cohen, J.I. A new vaccine to prevent herpes zoster. N. Engl. J. Med. 2015, 372, 2149–2150. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, A.L.; Heineman, T.C.; Lal, H.; Godeaux, O.; Chlibek, R.; Hwang, S.J.; McElhaney, J.E.; Vesikari, T.; Andrews, C.; Choi, W.S.; et al. Immune Responses to a Recombinant Glycoprotein E Herpes Zoster Vaccine in Adults Aged 50 Years or Older. J. Infect. Dis. 2018, 217, 1750–1760. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, A.L.; Lal, H.; Kovac, M.; Chlibek, R.; Hwang, S.J.; Diez-Domingo, J.; Godeaux, O.; Levin, M.J.; McElhaney, J.E.; Puig-Barbera, J.; et al. Efficacy of the Herpes Zoster Subunit Vaccine in Adults 70 Years of Age or Older. N. Engl. J. Med. 2016, 375, 1019–1032. [Google Scholar] [CrossRef]
- Gilderman, L.I.; Lawless, J.F.; Nolen, T.M.; Sterling, T.; Rutledge, R.Z.; Fernsler, D.A.; Azrolan, N.; Sutradhar, S.C.; Wang, W.W.; Chan, I.S.; et al. A double-blind, randomized, controlled, multicenter safety and immunogenicity study of a refrigerator-stable formulation of Zostavax. Clin. Vaccine Immunol. 2008, 15, 314–319. [Google Scholar] [CrossRef] [Green Version]
- Oxman, M.N.; Levin, M.J.; Johnson, G.R.; Schmader, K.E.; Straus, S.E.; Gelb, L.D.; Arbeit, R.D.; Simberkoff, M.S.; Gershon, A.A.; Davis, L.E.; et al. A vaccine to prevent herpes zoster and postherpetic neuralgia in older adults. N. Engl. J. Med. 2005, 352, 2271–2284. [Google Scholar] [CrossRef] [Green Version]
- Upadhyay, S.; Jeena, G.S.; Shikha; Shukla, R.K. Recent advances in steroidal saponins biosynthesis and in vitro production. Planta 2018, 248, 519–544. [Google Scholar] [CrossRef] [PubMed]
- Wang, P. Natural and Synthetic Saponins as Vaccine Adjuvants. Vaccines 2021, 9, 222. [Google Scholar] [CrossRef]
- Monslow, M.A.; Elbashir, S.; Sullivan, N.L.; Thiriot, D.S.; Ahl, P.; Smith, J.; Miller, E.; Cook, J.; Cosmi, S.; Thoryk, E.; et al. Immunogenicity generated by mRNA vaccine encoding VZV gE antigen is comparable to adjuvanted subunit vaccine and better than live attenuated vaccine in nonhuman primates. Vaccine 2020, 38, 5793–5802. [Google Scholar] [CrossRef]
- Berarducci, B.; Ikoma, M.; Stamatis, S.; Sommer, M.; Grose, C.; Arvin, A.M. Essential functions of the unique N-terminal region of the varicella-zoster virus glycoprotein E ectodomain in viral replication and in the pathogenesis of skin infection. J. Virol. 2006, 80, 9481–9496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malavige, G.N.; Jones, L.; Black, A.P.; Ogg, G.S. Varicella zoster virus glycoprotein E-specific CD4+ T cells show evidence of recent activation and effector differentiation, consistent with frequent exposure to replicative cycle antigens in healthy immune donors. Clin. Exp. Immunol. 2008, 152, 522–531. [Google Scholar] [CrossRef] [PubMed]
- Mo, C.; Lee, J.; Sommer, M.; Grose, C.; Arvin, A.M. The requirement of varicella zoster virus glycoprotein E (gE) for viral replication and effects of glycoprotein I on gE in melanoma cells. Virology 2002, 304, 176–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mo, C.; Schneeberger, E.E.; Arvin, A.M. Glycoprotein E of varicella-zoster virus enhances cell-cell contact in polarized epithelial cells. J. Virol. 2000, 74, 11377–11387. [Google Scholar] [CrossRef] [Green Version]
- Gershon, A.A.; Sherman, D.L.; Zhu, Z.; Gabel, C.A.; Ambron, R.T.; Gershon, M.D. Intracellular transport of newly synthesized varicella-zoster virus: Final envelopment in the trans-Golgi network. J. Virol. 1994, 68, 6372–6390. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.H.; Gershon, M.D.; Lungu, O.; Zhu, Z.; Gershon, A.A. Trafficking of varicella-zoster virus glycoprotein gI: T(338)-dependent retention in the trans-Golgi network, secretion, and mannose 6-phosphate-inhibitable uptake of the ectodomain. J. Virol. 2000, 74, 6600–6613. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Z.; Hao, Y.; Gershon, M.D.; Ambron, R.T.; Gershon, A.A. Targeting of glycoprotein I (gE) of varicella-zoster virus to the trans-Golgi network by an AYRV sequence and an acidic amino acid-rich patch in the cytosolic domain of the molecule. J. Virol. 1996, 70, 6563–6575. [Google Scholar] [CrossRef] [Green Version]
- Moffat, J.; Mo, C.; Cheng, J.J.; Sommer, M.; Zerboni, L.; Stamatis, S.; Arvin, A.M. Functions of the C-terminal domain of varicella-zoster virus glycoprotein E in viral replication in vitro and skin and T-cell tropism in vivo. J. Virol. 2004, 78, 12406–12415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olson, J.K.; Grose, C. Endocytosis and recycling of varicella-zoster virus Fc receptor glycoprotein gE: Internalization mediated by a YXXL motif in the cytoplasmic tail. J. Virol. 1997, 71, 4042–4054. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.H.; Gershon, M.D.; Lungu, O.; Zhu, Z.; Mallory, S.; Arvin, A.M.; Gershon, A.A. Essential role played by the C-terminal domain of glycoprotein I in envelopment of varicella-zoster virus in the trans-Golgi network: Interactions of glycoproteins with tegument. J. Virol. 2001, 75, 323–340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bahl, K.; Senn, J.J.; Yuzhakov, O.; Bulychev, A.; Brito, L.A.; Hassett, K.J.; Laska, M.E.; Smith, M.; Almarsson, O.; Thompson, J.; et al. Preclinical and Clinical Demonstration of Immunogenicity by mRNA Vaccines against H10N8 and H7N9 Influenza Viruses. Mol. Ther. 2017, 25, 1316–1327. [Google Scholar] [CrossRef] [Green Version]
- Holtkamp, S.; Kreiter, S.; Selmi, A.; Simon, P.; Koslowski, M.; Huber, C.; Tureci, O.; Sahin, U. Modification of antigen-encoding RNA increases stability, translational efficacy, and T-cell stimulatory capacity of dendritic cells. Blood 2006, 108, 4009–4017. [Google Scholar] [CrossRef]
- Cao, H.; Wang, Y.; Luan, N.; Liu, C. Immunogenicity of Varicella-Zoster Virus Glycoprotein E Formulated with Lipid Nanoparticles and Nucleic Immunostimulators in Mice. Vaccines 2021, 9, 310. [Google Scholar] [CrossRef]
- Richner, J.M.; Himansu, S.; Dowd, K.A.; Butler, S.L.; Salazar, V.; Fox, J.M.; Julander, J.G.; Tang, W.W.; Shresta, S.; Pierson, T.C.; et al. Modified mRNA Vaccines Protect against Zika Virus Infection. Cell 2017, 169, 176. [Google Scholar] [CrossRef] [Green Version]
- Richner, J.M.; Jagger, B.W.; Shan, C.; Fontes, C.R.; Dowd, K.A.; Cao, B.; Himansu, S.; Caine, E.A.; Nunes, B.T.D.; Medeiros, D.B.A.; et al. Vaccine Mediated Protection Against Zika Virus-Induced Congenital Disease. Cell 2017, 170, 273–283. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Huang, P.; Zhao, D.; Xia, M.; Zhong, W.; Jiang, X.; Tan, M. Effects of rotavirus NSP4 protein on the immune response and protection of the SR69A-VP8 nanoparticle rotavirus vaccine. Vaccine 2021, 39, 263–271. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Chu, X.; Yan, M.; Qi, J.; Liu, H.; Gao, F.; Gao, R.; Ma, G.; Ma, Y. Encapsulation of Poly I:C and the natural phosphodiester CpG ODN enhanced the efficacy of a hyaluronic acid-modified cationic lipid-PLGA hybrid nanoparticle vaccine in TC-1-grafted tumors. Int. J. Pharm. 2018, 553, 327–337. [Google Scholar] [CrossRef]
- Roberts, A.D.; Ely, K.H.; Woodland, D.L. Differential contributions of central and effector memory T cells to recall responses. J. Exp. Med. 2005, 202, 123–133. [Google Scholar] [CrossRef] [PubMed]
- Sckisel, G.D.; Mirsoian, A.; Minnar, C.M.; Crittenden, M.; Curti, B.; Chen, J.Q.; Blazar, B.R.; Borowsky, A.D.; Monjazeb, A.M.; Murphy, W.J. Differential phenotypes of memory CD4 and CD8 T cells in the spleen and peripheral tissues following immunostimulatory therapy. J. Immunother. Cancer 2017, 5, 33. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Valcarcel, M.; Fowler, W.J.; Harper, D.R.; Jeffries, D.J.; Layton, G.T. Induction of neutralizing antibody and T-cell responses to varicella-zoster virus (VZV) using Ty-virus-like particles carrying fragments of glycoprotein E (gE). Vaccine 1997, 15, 709–719. [Google Scholar] [CrossRef]
- Dendouga, N.; Fochesato, M.; Lockman, L.; Mossman, S.; Giannini, S.L. Cell-mediated immune responses to a varicella-zoster virus glycoprotein E vaccine using both a TLR agonist and QS21 in mice. Vaccine 2012, 30, 3126–3135. [Google Scholar] [CrossRef]
- Wang, Y.; Qi, J.; Cao, H.; Liu, C. Immune Responses to Varicella-Zoster Virus Glycoprotein E Formulated with Poly(Lactic-co-Glycolic Acid) Nanoparticles and Nucleic Acid Adjuvants in Mice. Virol. Sin. 2020, 36, 122–132. [Google Scholar] [CrossRef]
Figure 1.
Characterization of LNP mRNA vaccines. (A) Diameters tested by size analyzer; (B) polydispersity index of LNPs; (C) mRNA encapsulation efficiency; (D) loaded mRNA detected with a 1% denatured agarose gel.
Figure 1.
Characterization of LNP mRNA vaccines. (A) Diameters tested by size analyzer; (B) polydispersity index of LNPs; (C) mRNA encapsulation efficiency; (D) loaded mRNA detected with a 1% denatured agarose gel.

Figure 2.
gE-specific IgG titers detected by enzyme-linked immunosorbent assay (ELISA). IgG titers were compared using one-way analysis of variance (ANOVA) followed by Dunnett’s multiple comparisons test, with the gE-M group as a control. * p < 0.05, ** p < 0.01. ns, no significant difference.
Figure 2.
gE-specific IgG titers detected by enzyme-linked immunosorbent assay (ELISA). IgG titers were compared using one-way analysis of variance (ANOVA) followed by Dunnett’s multiple comparisons test, with the gE-M group as a control. * p < 0.05, ** p < 0.01. ns, no significant difference.

Figure 3.
Enzyme-linked immunospot assay (ELISPOT) of splenocytes. (A) IFN-γ-producing splenocytes after gE stimulation; (B) IL-2-producing splenocytes after gE stimulation. ELISPOT numbers were compared using one-way ANOVA followed by Dunnett’s multiple comparisons test, with the gE-M group as a control. ** p < 0.01.
Figure 3.
Enzyme-linked immunospot assay (ELISPOT) of splenocytes. (A) IFN-γ-producing splenocytes after gE stimulation; (B) IL-2-producing splenocytes after gE stimulation. ELISPOT numbers were compared using one-way ANOVA followed by Dunnett’s multiple comparisons test, with the gE-M group as a control. ** p < 0.01.

Figure 4.
Flow cytometry assay for gE-specific IFN-γ-producing T cells. (A) Proportion of IFN-γ-producing CD4+ T cells among splenocytes after stimulation with gE; (B) proportion of IFN-γ-producing CD8+ T cells among splenocytes after stimulation with gE. Data were analyzed using one-way ANOVA followed by Dunnett’s multiple comparisons test, with the gE-M group as a control. * p < 0.05.
Figure 4.
Flow cytometry assay for gE-specific IFN-γ-producing T cells. (A) Proportion of IFN-γ-producing CD4+ T cells among splenocytes after stimulation with gE; (B) proportion of IFN-γ-producing CD8+ T cells among splenocytes after stimulation with gE. Data were analyzed using one-way ANOVA followed by Dunnett’s multiple comparisons test, with the gE-M group as a control. * p < 0.05.

Figure 5.
Flow cytometry assay for memory T cells. (A) Proportion of memory CD4+ T cells among splenocytes after stimulation with gE; (B) proportion of CD8+ memory T cells among splenocytes after stimulation with gE. Data were analyzed using one-way ANOVA followed by Dunnett’s multiple comparisons test, with the gE-M group as a control. * p < 0.05.
Figure 5.
Flow cytometry assay for memory T cells. (A) Proportion of memory CD4+ T cells among splenocytes after stimulation with gE; (B) proportion of CD8+ memory T cells among splenocytes after stimulation with gE. Data were analyzed using one-way ANOVA followed by Dunnett’s multiple comparisons test, with the gE-M group as a control. * p < 0.05.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Cao, H.; Wang, Y.; Luan, N.; Lin, K.; Liu, C. Effects of Varicella-Zoster Virus Glycoprotein E Carboxyl-Terminal Mutation on mRNA Vaccine Efficacy. Vaccines 2021, 9, 1440. https://doi.org/10.3390/vaccines9121440
AMA Style
Cao H, Wang Y, Luan N, Lin K, Liu C. Effects of Varicella-Zoster Virus Glycoprotein E Carboxyl-Terminal Mutation on mRNA Vaccine Efficacy. Vaccines. 2021; 9(12):1440. https://doi.org/10.3390/vaccines9121440
Chicago/Turabian StyleCao, Han, Yunfei Wang, Ning Luan, Kangyang Lin, and Cunbao Liu. 2021. "Effects of Varicella-Zoster Virus Glycoprotein E Carboxyl-Terminal Mutation on mRNA Vaccine Efficacy" Vaccines 9, no. 12: 1440. https://doi.org/10.3390/vaccines9121440
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.