
Immune Responses to Pandemic H1N1 Influenza Virus Infection in Pigs Vaccinated with a Conserved Hemagglutinin HA1 Peptide Adjuvanted with CAF®01 or CDA/αGalCerMPEG

 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Vaccine Antigens and Adjuvants
2.2. Cell Cultures and Virus
2.3. Experimental Design
2.4. Sampling
2.5. Clinical Signs and Pathological Assessment
2.6. Quantitative PCR RT-qPCR–Viral Load
2.7. Assessment of IFNγ Producing Cells
2.8. Humoral Immune Response Evaluation
2.9. Hemagglutination Inhibition and Neutralization Assays
2.10. Statistical Analysis
3. Results
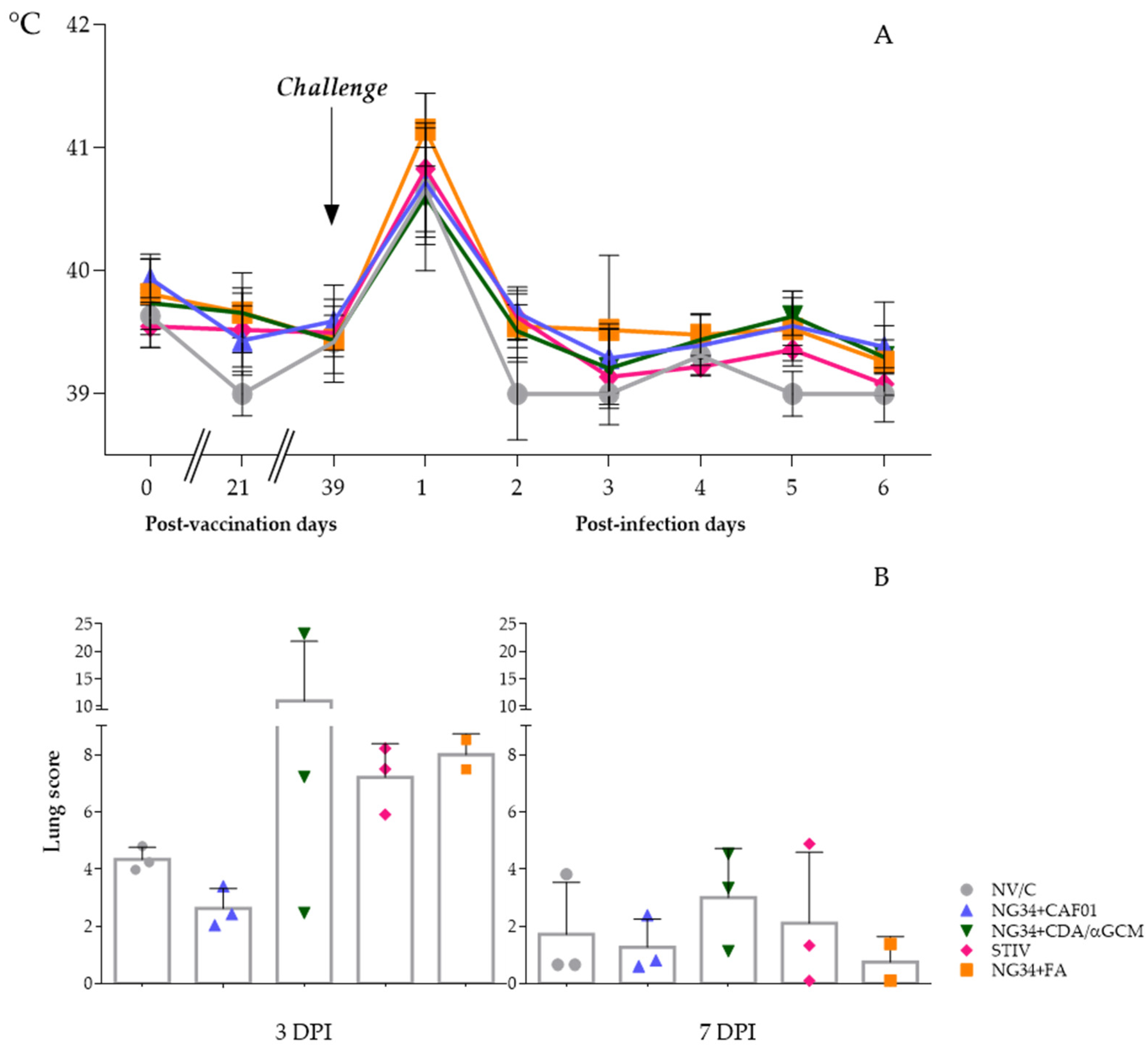
3.1. Clinical Signs and Pathology
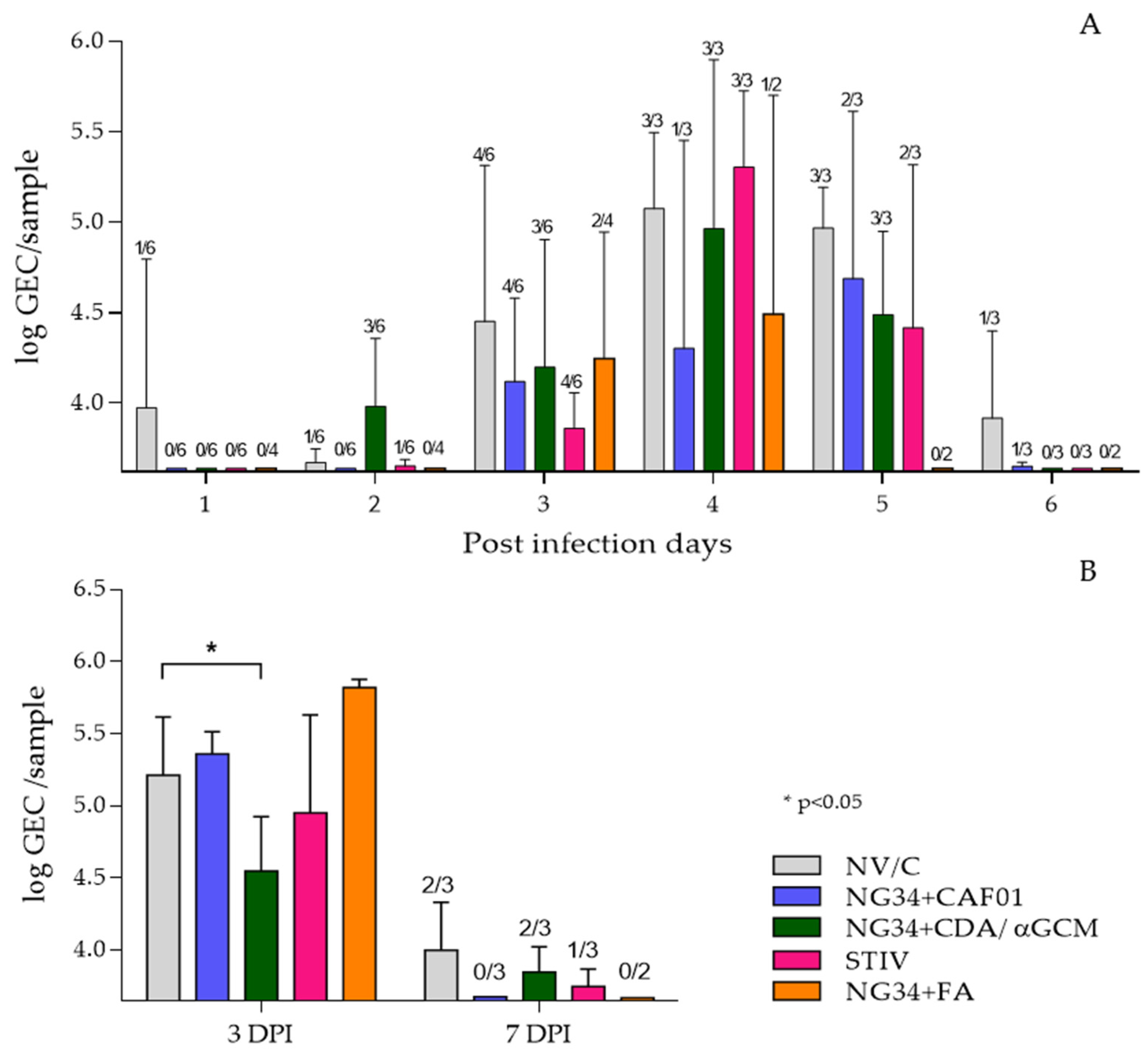
3.2. RT-qPCR Results–Viral Load
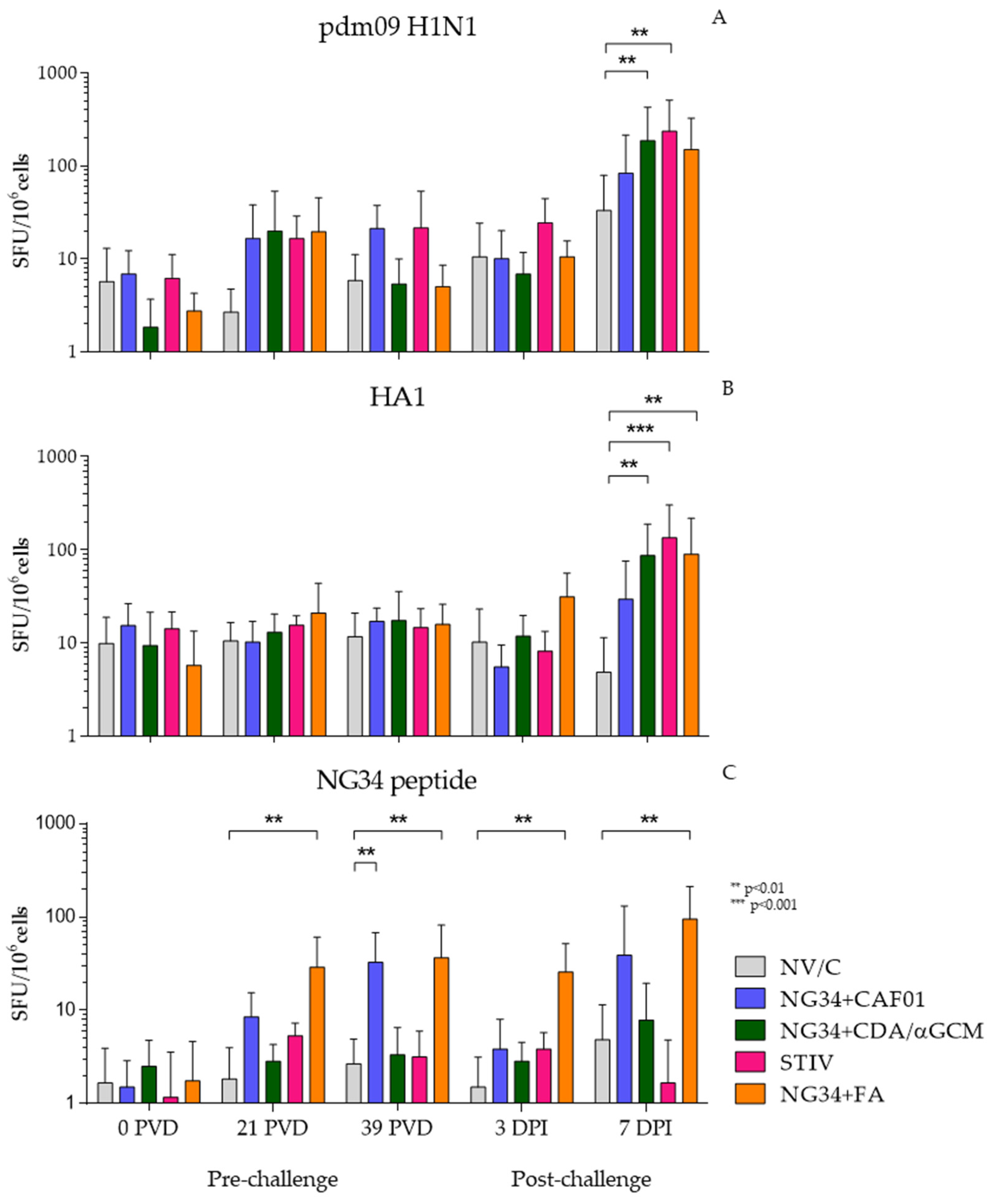
3.3. Cell Immune Response-ELISPOT
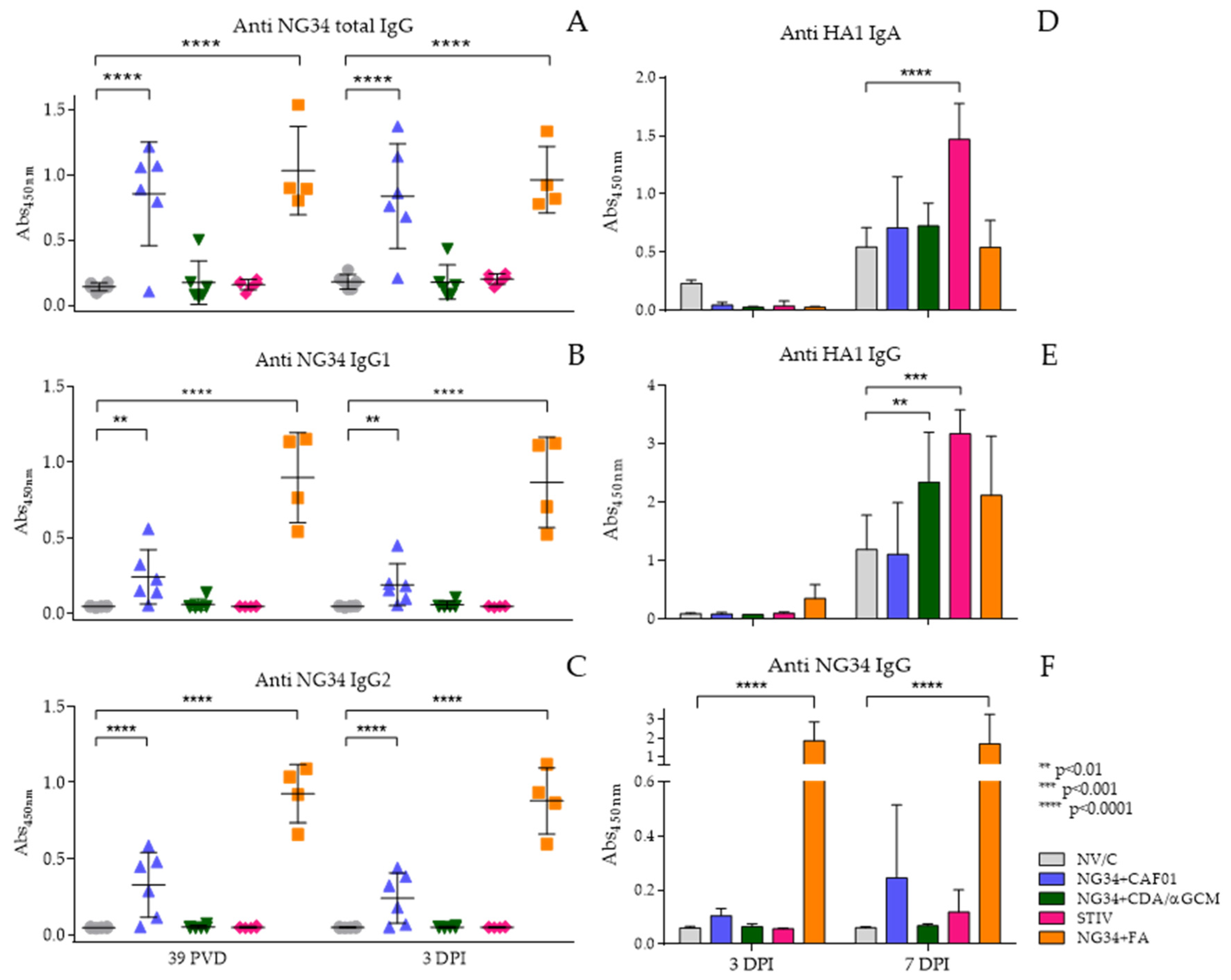
3.4. Antibody Response
3.5. Hemagglutination Inhibition and Neutralizing Antibodies
4. Discussion
5. Conclusions
6. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Long, J.S.; Mistry, B.; Haslam, S.M.; Barclay, W.S. Host and Viral Determinants of Influenza A Virus Species Specificity. Nat. Rev. Microbiol. 2019, 17, 67–81. [Google Scholar] [CrossRef]
- Wallensten, A. Influenza Virus in Wild Birds and Mammals Other than Man. Microb. Ecol. Health Dis. 2007, 19, 122–139. [Google Scholar] [CrossRef]
- Van Reeth, K.; Vincent, A.L. Chapter 36. Influenza Viruses. In Diseases of Swine; Wiley Online Books; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2019; ISBN 9781119350927. [Google Scholar]
- Rajao, D.S.; Vincent, A.L. Swine as a Model for Influenza Avirus Infection and Immunity. ILAR J. 2015, 56, 44–52. [Google Scholar] [CrossRef]
- Houser, K.; Subbarao, K. Influenza Vaccines: Challenges and Solutions. Cell Host Microbe 2015, 17, 295–300. [Google Scholar] [CrossRef] [Green Version]
- Yedidia, T.B.; Rudolph, W. Development of a Universal Influenza Vaccine. Bioprocess Int. 2011, 9, 46–49. [Google Scholar] [CrossRef] [Green Version]
- Sautto, G.A.; Kirchenbaum, G.A.; Ross, T.M. Towards a Universal Influenza Vaccine: Different Approaches for One Goal. Virol. J. 2018, 15, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keshavarz, M.; Mirzaei, H.; Salemi, M.; Momeni, F.; Mousavi, M.J.; Sadeghalvad, M.; Arjeini, Y.; Solaymani-Mohammadi, F.; Sadri Nahand, J.; Namdari, H.; et al. Influenza Vaccine: Where Are We and Where Do We Go? Rev. Med. Virol. 2019, 29, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Soema, P.C.; Kompier, R.; Amorij, J.P.; Kersten, G.F.A. Current and next Generation Influenza Vaccines: Formulation and Production Strategies. Eur. J. Pharm. Biopharm. 2015, 94, 251–263. [Google Scholar] [CrossRef] [Green Version]
- Veljkovic, V.; Niman, H.L.; Glisic, S.; Veljkovic, N.; Perovic, V.; Muller, C.P. Identification of Hemagglutinin Structural Domain and Polymorphisms Which May Modulate Swine H1N1 Interactions with Human Receptor. BMC Struct. Biol. 2009, 9, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vergara-Alert, J.; Argilaguet, J.M.; Busquets, N.; Ballester, M.; Martín-Valls, G.E.; Rivas, R.; López-Soria, S.; Solanes, D.; Majó, N.; Segalés, J.; et al. Conserved Synthetic Peptides from the Hemagglutinin of Influenza Viruses Induce Broad Humoral and T-Cell Responses in a Pig Model. PLoS ONE 2012, 7. [Google Scholar] [CrossRef]
- Sisteré-Oró, M.; Pedersen, G.K.; Córdoba, L.; López-Serrano, S.; Christensen, D.; Darji, A. Influenza NG-34 T Cell Conserved Epitope Adjuvanted with CAF01 as a Possible Influenza Vaccine Candidate. Vet. Res. 2020, 51, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Pedersen, G.K.; Andersen, P.; Christensen, D. Immunocorrelates of CAF Family Adjuvants. Semin. Immunol. 2018, 39, 4–13. [Google Scholar] [CrossRef]
- Martel, C.J.M.; Agger, E.M.; Poulsen, J.J.; Jensen, T.H.; Andresen, L.; Christensen, D.; Nielsen, L.P.; Blixenkrone-Møller, M.; Andersen, P.; Aasted, B. CAF01 Potentiates Immune Responses and Efficacy of an Inactivated Influenza Vaccine in Ferrets. PLoS ONE 2011, 6. [Google Scholar] [CrossRef]
- Christensen, D.; Christensen, J.P.; Korsholm, K.S.; Isling, L.K.; Erneholm, K.; Thomsen, A.R.; Andersen, P. Seasonal Influenza Split Vaccines Confer Partial Cross-Protection against Heterologous Influenza Virus in Ferrets When Combined with the CAF01 Adjuvant. Front. Immunol. 2018, 8, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Volckmar, J.; Knop, L.; Stegemann-Koniszewski, S.; Schulze, K.; Ebensen, T.; Guzmán, C.A.; Bruder, D. The STING Activator C-Di-AMP Exerts Superior Adjuvant Properties than the Formulation Poly(I:C)/CpG after Subcutaneous Vaccination with Soluble Protein Antigen or DEC-205-Mediated Antigen Targeting to Dendritic Cells. Vaccine 2019, 37, 4963–4974. [Google Scholar] [CrossRef] [PubMed]
- Ebensen, T.; Link, C.; Riese, P.; Schulze, K.; Morr, M.; Guzmán, C.A. A Pegylated Derivative of α-Galactosylceramide Exhibits Improved Biological Properties. J. Immunol. 2007, 179, 2065–2073. [Google Scholar] [CrossRef] [Green Version]
- Reed, L.J.; Muench, H. A simple method of estimating fifty per cent endpoints. Am. J. Epidemiol. 1938, 27, 493–497. [Google Scholar] [CrossRef]
- Spackman, E.; Senne, D.A.; Myers, T.J.; Bulaga, L.L.; Garber, L.P.; Perdue, M.L.; Lohman, K.; Daum, L.T.; Suarez, D.L. Development of a Real-Time Reverse Transcriptase PCR Assay for Type A Influenza Virus and the Avian H5 and H7 Hemagglutinin Subtypes. J. Clin. Microbiol. 2002, 40, 3256–3260. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Morante, B.; Segalés, J.; Fraile, L.; Pérez de Rozas, A.; Maiti, H.; Coll, T.; Sibila, M. Assessment of Mycoplasma Hyopneumoniae-Induced Pneumonia Using Different Lung Lesion Scoring Systems: A Comparative Review. J. Comp. Pathol. 2016, 154, 125–134. [Google Scholar] [CrossRef] [PubMed]
- Vergara-Alert, J.; Busquets, N.; Ballester, M.; Chaves, A.J.; Rivas, R.; Dolz, R.; Wang, Z.; Pleschka, S.; Majó, N.; Rodríguez, F.; et al. The NS Segment of H5N1 Avian Influenza Viruses (AIV) Enhances the Virulence of an H7N1 AIV in Chickens. Vet. Res. 2014, 45, 1–11. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization. Manual for the Laboratory Diagnosis and Virological Surveillance of Influenza; World Health Organization: Geneva, Switzerland, 2011; ISBN 978-92-4-154809-0. [Google Scholar]
- Organisation International des Épizooties (OIE). W.O. for A.H. Chapter 3. 9. 7. Influenza A Virus of Swine. In Manual of Diagnostic Tests and Vaccines for Terrestrial Animals 2019; OIE: Paris, France, 2019; pp. 1594–1607. ISBN 978-92-95108-18-9. [Google Scholar]
- Bastola, R.; Noh, G.; Keum, T.; Bashyal, S.; Seo, J.E.; Choi, J.; Oh, Y.; Cho, Y.S.; Lee, S. Vaccine Adjuvants: Smart Components to Boost the Immune System. Arch. Pharm. Res. 2017, 40, 1238–1248. [Google Scholar] [CrossRef] [PubMed]
- Donaldson, B.; Al-Barwani, F.; Young, V.; Scullion, S.; Ward, V.; Young, S. Virus-like Particles, a Versatile Subunit Vaccine Platform. Adv. Deliv. Sci. Technol. 2015, 2014, 159–180. [Google Scholar] [CrossRef]
- Van Roey, G.A.; Arias, M.A.; Tregoning, J.S.; Rowe, G.; Shattock, R.J. Thymic Stromal Lymphopoietin (TSLP) Acts as a Potent Mucosal Adjuvant for HIV-1 Gp140 Vaccination in Mice. Eur. J. Immunol. 2012, 42, 353–363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nouri-Shirazi, M.; Guinet, E. Exposure to Nicotine Adversely Affects the Dendritic Cell System and Compromises Host Response to Vaccination. J. Immunol. 2012, 188, 2359–2370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pellegrino, P.; Clementi, E.; Radice, S. On Vaccine’s Adjuvants and Autoimmunity: Current Evidence and Future Perspectives. Autoimmun. Rev. 2015, 14, 880–888. [Google Scholar] [CrossRef] [PubMed]
- Shaw, C.A.; Tomljenovic, L. Aluminum in the Central Nervous System (CNS): Toxicity in Humans and Animals, Vaccine Adjuvants, and Autoimmunity. Immunol. Res. 2013, 56, 304–316. [Google Scholar] [CrossRef] [PubMed]
- Agger, E.M.; Rosenkrands, I.; Hansen, J.; Brahimi, K.; Vandahl, B.S.; Aagaard, C.; Werninghaus, K.; Kirschning, C.; Lang, R.; Christensen, D.; et al. Cationic Liposomes Formulated with Synthetic Mycobacterial Cordfactor (CAF01): A Versatile Adjuvant for Vaccines with Different Immunological Requirements. PLoS ONE 2008, 3. [Google Scholar] [CrossRef] [Green Version]
- Wack, A.; Baudner, B.C.; Hilbert, A.K.; Manini, I.; Nuti, S.; Tavarini, S.; Scheffczik, H.; Ugozzoli, M.; Singh, M.; Kazzaz, J.; et al. Combination Adjuvants for the Induction of Potent, Long-Lasting Antibody and T-Cell Responses to Influenza Vaccine in Mice. Vaccine 2008, 26, 552–561. [Google Scholar] [CrossRef] [PubMed]
- Jegaskanda, S.; Job, E.R.; Kramski, M.; Laurie, K.; Isitman, G.; de Rose, R.; Winnall, W.R.; Stratov, I.; Brooks, A.G.; Reading, P.C.; et al. Cross-Reactive Influenza-Specific Antibody-Dependent Cellular Cytotoxicity Antibodies in the Absence of Neutralizing Antibodies. J. Immunol. 2013, 190, 1837–1848. [Google Scholar] [CrossRef] [Green Version]
- Gu, W.; Madrid, D.M.D.; Yang, G.; Artiaga, B.L.; Loeb, J.C.; Castleman, W.L.; Richt, J.A.; Lednicky, J.A.; Driver, J.P. Unaltered Influenza Disease Outcomes in Swine Prophylactically Treated with α-Galactosylceramide. Dev. Comp. Immunol. 2021, 114, 103843. [Google Scholar] [CrossRef]
- Ebensen, T.; Debarry, J.; Pedersen, G.K.; Blazejewska, P.; Weissmann, S.; Schulze, K.; McCullough, K.C.; Cox, R.J.; Guzmán, C.A. Mucosal Administration of Cycle-Di-Nucleotide-Adjuvanted Virosomes Efficiently Induces Protection against Influenza H5N1 in Mice. Front. Immunol. 2017, 8, 1–15. [Google Scholar] [CrossRef]
- Gauger, P.C.; Vincent, A.L.; Loving, C.L.; Lager, K.M.; Janke, B.H.; Kehrli, M.E.; Roth, J.A. Enhanced Pneumonia and Disease in Pigs Vaccinated with an Inactivated Human-like (δ-Cluster) H1N2 Vaccine and Challenged with Pandemic 2009 H1N1 Influenza Virus. Vaccine 2011, 29, 2712–2719. [Google Scholar] [CrossRef]
- Larsen, D.L.; Karasin, A.; Zuckermann, F.; Olsen, C.W. Systemic and Mucosal Immune Responses to H1N1 Influenza Virus Infection in Pigs. Vet. Microbiol. 2000, 74, 117–131. [Google Scholar] [CrossRef]
- Jelinek, I.; Leonard, J.N.; Price, G.; Brown, K.N.; Meyer-Manlapat, A.; Goldsmith, P.K.; Wang, Y.; Venzon, D.; Epstein, S.L.; Segal, D.M. Toll-Like Receptor 3-Specific DsRNA Oligonucleotide Adjuvants Induce Dendritic Cell Cross-Presentation, CTL Responses and Antiviral Protection. J. Immunol. 2011, 186, 2422–2429. [Google Scholar] [CrossRef] [PubMed]
- Zaks, K.; Jordan, M.; Guth, A.; Sellins, K.; Kedl, R.; Izzo, A.; Bosio, C.; Dow, S. Efficient Immunization and Cross-Priming by Vaccine Adjuvants Containing TLR3 or TLR9 Agonists Complexed to Cationic Liposomes. J. Immunol. 2006, 176, 7335–7345. [Google Scholar] [CrossRef] [Green Version]
- Le Bon, A.; Tough, D.F. Type I Interferon as a Stimulus for Cross-Priming. Cytokine Growth Factor Rev. 2008, 19, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Coffman, R.L.; Sher, A.; Seder, R.A. Vaccine Adjuvants: Putting Innate Immunity to Work. Immunity 2010, 33, 492–503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christensen, D.; Henriksen-Lacey, M.; Kamath, A.T.; Lindenstrøm, T.; Korsholm, K.S.; Christensen, J.P.; Rochat, A.F.; Lambert, P.H.; Andersen, P.; Siegrist, C.A.; et al. A Cationic Vaccine Adjuvant Based on a Saturated Quaternary Ammonium Lipid Have Different in Vivo Distribution Kinetics and Display a Distinct CD4 T Cell-Inducing Capacity Compared to Its Unsaturated Analog. J. Control. Release 2012, 160, 468–476. [Google Scholar] [CrossRef]
- Olafsdottir, T.A.; Lindqvist, M.; Nookaew, I.; Andersen, P.; Maertzdorf, J.; Persson, J.; Christensen, D.; Zhang, Y.; Anderson, J.; Khoomrung, S.; et al. Comparative Systems Analyses Reveal Molecular Signatures of Clinically Tested Vaccine Adjuvants. Sci. Rep. 2016, 6, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Canini, L.; Holzer, B.; Morgan, S.; Dinie Hemmink, J.; Clark, B.; Consortium, s.D.; Woolhouse, M.E.J.; Tchilian, E.; Charleston, B. Timelines of Infection and Transmission Dynamics of H1N1pdm09 in Swine. PLoS Pathog. 2020, 16, e1008628. [Google Scholar] [CrossRef]
- Kyriakis, C.S.; Gramer, M.R.; Barbé, F.; Van Doorsselaere, J.; Van Reeth, K. Efficacy of Commercial Swine Influenza Vaccines against Challenge with a Recent European H1N1 Field Isolate. Vet. Microbiol. 2010, 144, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Waffarn, E.E.; Baumgarth, N. Protective B Cell Responses to Flu--No Fluke! J. Immunol. 2011, 186, 3823–3829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vidaña, B.; Brookes, S.M.; Everett, H.E.; Garcon, F.; Nuñez, A.; Engelhardt, O.; Major, D.; Hoschler, K.; Brown, I.H.; Zambon, M. Inactivated Pandemic 2009 H1N1 Influenza A Virus Human Vaccines Have Different Efficacy after Homologous Challenge in the Ferret Model. Influenza Other Respir. Viruses 2021, 15, 142–153. [Google Scholar] [CrossRef] [PubMed]
- Malik Peiris, F.R.S.; Guan, Y. Options for the Control of Influenza VII. In Proceedings of the Immunogenicity and Protective Efficacy of Candidate Pandemic H1N1 Vaccines in the Ferret Model, Hong Kong, China, 3–7 September 2010; p. 409. [Google Scholar]
- Woodworth, J.S.; Christensen, D.; Cassidy, J.P.; Agger, E.M.; Mortensen, R.; Andersen, P. Mucosal Boosting of H56:CAF01 Immunization Promotes Lung-Localized T Cells and an Accelerated Pulmonary Response to Mycobacterium Tuberculosis Infection without Enhancing Vaccine Protection. Mucosal Immunol. 2019, 12, 816–826. [Google Scholar] [CrossRef] [PubMed]
- Lorenzen, E.; Follmann, F.; Bøje, S.; Erneholm, K.; Olsen, A.W.; Agerholm, J.S.; Jungersen, G.; Andersen, P. Intramuscular Priming and Intranasal Boosting Induce Strong Genital Immunity through Secretory IgA in Minipigs Infected with Chlamydia Trachomatis. Front. Immunol. 2015, 6, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Khatri, M.; Dwivedi, V.; Krakowka, S.; Manickam, C.; Ali, A.; Wang, L.; Qin, Z.; Renukaradhya, G.J.; Lee, C.W. Swine Influenza H1N1 Virus Induces Acute Inflammatory Immune Responses in Pig Lungs: A Potential Animal Model for Human H1N1 Influenza Virus. J. Virol. 2010, 84, 11210–11218. [Google Scholar] [CrossRef] [Green Version]
- Artiaga, B.L.; Yang, G.; Hackmann, T.J.; Liu, Q.; Richt, J.A.; Salek-Ardakani, S.; Castleman, W.L.; Lednicky, J.A.; Driver, J.P. α-Galactosylceramide Protects Swine against Influenza Infection When Administered as a Vaccine Adjuvant. Sci. Rep. 2016, 6, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Matos, M.N.; Cazorla, S.I.; Schulze, K.; Ebensen, T.; Guzmán, C.A.; Malchiodi, E.L. Immunization with Tc52 or Its Amino Terminal Domain Adjuvanted with C-Di-AMP Induces Th17+Th1 Specific Immune Responses and Confers Protection against Trypanosoma Cruzi. PLoS Negl. Trop. Dis. 2017, 11, 1–16. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experimental Group | N | Antigen | Adjuvant |
|---|---|---|---|
| 1. Unvaccinated/unchallenged (NV/NC) | 6 | PBS | None |
| 2. Unvaccinated/challenged (NV/C) | 6 | PBS | None |
| 3. NG34–CAF®01 | 6 | 50 µg of NG34 | 980 µL of CAF®01 |
| 4. NG34–CDA/αGalCerMPEG (CDA/αGCM) | 6 | 50 µg of NG34 * | 25 µg of CDA + 25 µg αGCM |
| 5. NG34–Freund’s Adjuvant (FA) | 4 | 50 µg of NG34 * | 600 µL of CFA/IFA 1 |
| 6. Seasonal Trivalent Influenza Vaccine (STIV) | 6 | 500 µL of Chiroflu® 2018–19 seasonal vaccine 2 | |
| Experimental Group | AUC Until 3 dpi (n = 6) | AUC Until 6 dpi (n = 3) | ||
|---|---|---|---|---|
| Mean | SD | Mean | SD | |
| Unvaccinated/Challenged (NV/C) | 0.6052 | ±0.6680 | 4.1520 | ±0.9719 |
| NG34 + CAF®01 | 0.2397 | ±0.2316 | 2.2317 | ±2.2538 |
| NG34 + CDA/αGCM | 0.6228 | ±0.6968 | 2.8888 | ±2.3776 |
| Seasonal Trivalent Influenza Vaccine (TIV) | 0.1255 | ±0.1180 | 2.5247 | ±0.4745 |
| NG34 + FA * | 0.3036 | ±0.3506 | 1.4695 | ±2.0782 |
| Experimental Group | HI Antibody Titer in Sera * | Neutralization Antibody Titer in BALF * |
|---|---|---|
| Unvaccinated/Challenged (NV/C) | 80 | 20 |
| 80 | 40 | |
| 80 | 30 | |
| NG34 + CAF®01 | 320 | 0 |
| 80 | 30 | |
| 160 | 120 | |
| NG34 + CDA/αGCM | 160 | 80 |
| 160 | 40 | |
| 80 | 20 | |
| Seasonal Trivalent Influenza Vaccine (STIV) | 160 | 120 |
| 640 | 30 | |
| 2560 | 100 | |
| NG34 + FA | 160 | 30 |
| 320 | 30 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
López-Serrano, S.; Cordoba, L.; Pérez-Maillo, M.; Pleguezuelos, P.; Remarque, E.J.; Ebensen, T.; Guzmán, C.A.; Christensen, D.; Segalés, J.; Darji, A. Immune Responses to Pandemic H1N1 Influenza Virus Infection in Pigs Vaccinated with a Conserved Hemagglutinin HA1 Peptide Adjuvanted with CAF®01 or CDA/αGalCerMPEG. Vaccines 2021, 9, 751. https://doi.org/10.3390/vaccines9070751
López-Serrano S, Cordoba L, Pérez-Maillo M, Pleguezuelos P, Remarque EJ, Ebensen T, Guzmán CA, Christensen D, Segalés J, Darji A. Immune Responses to Pandemic H1N1 Influenza Virus Infection in Pigs Vaccinated with a Conserved Hemagglutinin HA1 Peptide Adjuvanted with CAF®01 or CDA/αGalCerMPEG. Vaccines. 2021; 9(7):751. https://doi.org/10.3390/vaccines9070751
Chicago/Turabian StyleLópez-Serrano, Sergi, Lorena Cordoba, Mónica Pérez-Maillo, Patricia Pleguezuelos, Edmond J. Remarque, Thomas Ebensen, Carlos A. Guzmán, Dennis Christensen, Joaquim Segalés, and Ayub Darji. 2021. "Immune Responses to Pandemic H1N1 Influenza Virus Infection in Pigs Vaccinated with a Conserved Hemagglutinin HA1 Peptide Adjuvanted with CAF®01 or CDA/αGalCerMPEG" Vaccines 9, no. 7: 751. https://doi.org/10.3390/vaccines9070751