
The Function of the PRRSV–Host Interactions and Their Effects on Viral Replication and Propagation in Antiviral Strategies


State Key Laboratory of Agrobiotechnology and Beijing Advanced Innovation Center for Food Nutrition and Human Health, College of Biological Sciences, China Agricultural University, Beijing 100193, China
*
Author to whom correspondence should be addressed.
Vaccines 2021, 9(4), 364; https://doi.org/10.3390/vaccines9040364
Submission received: 4 March 2021
/
Revised: 6 April 2021
/
Accepted: 6 April 2021
/
Published: 9 April 2021
(This article belongs to the Special Issue PRRSV Vaccinology and Immunology)
Abstract
:Porcine reproductive and respiratory syndrome virus (PRRSV) affects the global swine industry and causes disastrous economic losses each year. The genome of PRRSV is an enveloped single-stranded positive-sense RNA of approximately 15 kb. The PRRSV replicates primarily in alveolar macrophages of pig lungs and lymphatic organs and causes reproductive problems in sows and respiratory symptoms in piglets. To date, studies on how PRRSV survives in the host, the host immune response against viral infections, and pathogenesis, have been reported. PRRSV vaccines have been developed, including inactive virus, modified live virus, attenuated live vaccine, DNA vaccine, and immune adjuvant vaccines. However, there are certain problems with the durability and effectiveness of the licensed vaccines. Moreover, the high variability and fast-evolving populations of this RNA virus challenge the design of PRRSV vaccines, and thus effective vaccines against PRRSV have not been developed successfully. As is well known, viruses interact with the host to escape the host’s immune response and then replicate and propagate in the host, which is the key to virus survival. Here, we review the complex network and the mechanism of PRRSV–host interactions in the processes of virus infection. It is critical to develop novel antiviral strategies against PRRSV by studying these host–virus interactions and structures to better understand the molecular mechanisms of PRRSV immune escape.
1. Introduction
Porcine reproductive and respiratory syndrome virus (PRRSV) has undoubtedly become a substantial financial issue that has affected pig production and caused substantial economic losses for the swine industry worldwide since its outbreak in the 1980s [1,2]. Annual economic losses in the US due to PRRSV manifested in reproductive problems in sows and respiratory symptoms in piglets were assessed as reaching USD 664 million in 2013 [3,4]. The latest economic estimate in Germany indicated the losses on farm profits due to the PRRS virus were −19.1% on average and −41% in the worst case [2]. As a global swine pathogen that has caused catastrophic economic losses, PRRSV is the cause for continuing and widespread concern [5].
As a member of the Nidovirales order in the Arteriviridae family, PRRSV is an enveloped virus with an average diameter of 55 nm; it is a positive-stranded RNA virus with an approximately 15 kb genome with a 5′ cap and a 3′ poly A tail [4,6]. Except for the 5′ and 3′ untranslated regions at both ends, the PRRSV genome contains at least 11 known open reading frames (ORFs) [7]. The first two ORFs occupy about 75% of the viral genomes coding for polyproteins, including pp1a and pp1ab, by ribosome shifting, and then the PRRSV proteases hydrolyze and cleave polyproteins into at least 16 distinct nonstructural proteins (nsps) [4,8]. The nsps that participate in viral genome replication and transcription are essential for the survival of the PRRSV [9]. PRRSV virions are composed of an N protein (nucleocapsid protein) and a lipid envelop (GP2, E, GP3, GP4, M, GP5, ORF5a) that envelops the membrane [10,11]. M and GP5 are the major components of the virus coat [12].
The PRRSV is mainly divided into two genotypes: type 1 (represented by the European strain Lelystad Virus) and type 2 (represented by the North American strain VR-2332), and both PRRSV genotypes have only 50–60% nucleotide identity [6,13]. In addition to the genotype differences between PRRSV-1 and PRRSV-2, the host immune responses have been shown to differ, sometimes considerably due to their biological differences including pathogenicity (Table 1). A large number of studies have found a general consensus that PRRSV-2 causes more severe respiratory disease than PRRSV-1 [14]. Therefore, it is necessary to emphasize the host immune response induced by different genotypes. Due to high inter-strain genetic exchange and rapid mutations of PRRSV, it has always been a substantial challenge to design effective vaccines and drugs [4].
2. The Process of the PRRSV Entry and Infection
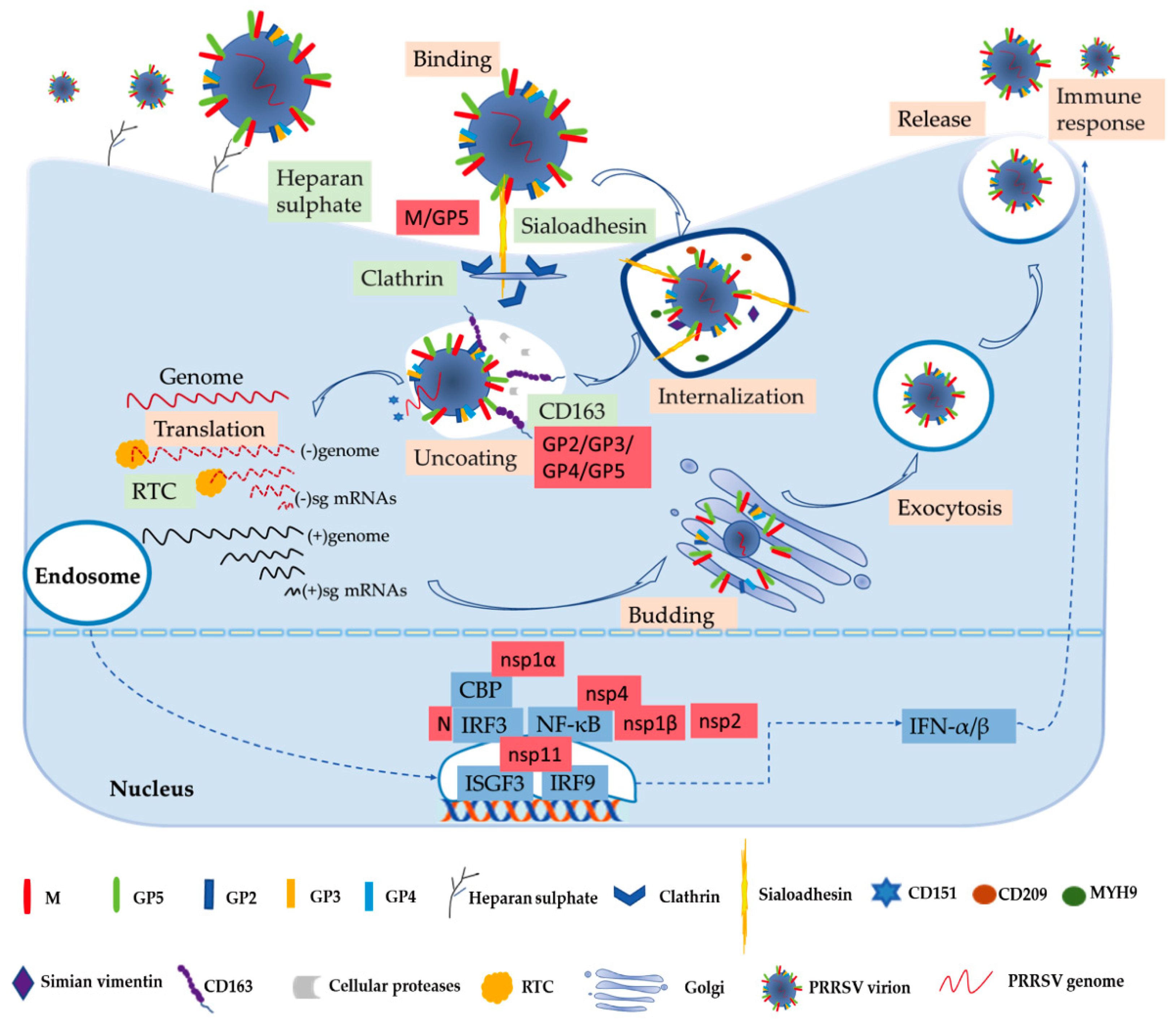
Numerous studies have been performed to learn more about the biochemistry of the PRRSV. The process of the virus entry into its host cell is the first crucial step in the infection and the focus of basic research [53]. PRRSV has a very narrow cell tropism, and the primary target cells are porcine alveolar macrophages (PAMs) [12,54]. Numerous studies have found that the existence of the specific entry mediators in the target cell leads to its restricted cell tropism [53,55]. These cellular factors, including heparan sulphate, CD163, porcine sialoadhesin (pSn), vimentin, CD151, CD209 (DC-SIGN), non-muscle myosin heavy chain 9 (MYH9), and others, are involved in virus binding, internalization, and genome release, as shown in Figure 1 [53,56].
In the early stages, PRRSV enters the macrophage via a receptor-mediated method [53,57]. PRRSV concentrates virions on the cell’s surface by interacting with heparan sulphate glycosaminoglycans that are present on the surface of mammalian cells, hence promoting a more efficient attachment for subsequent binding to one or more receptors involved in virus internalization; studies have found that PRRSV-1 and PRRSV-2 have different sensitivities to heparin [15,53,58]. Next, the integration with pSn gradually increases. The pSn, a PAMs-restricted type 1 transmembrane glycoprotein, is identified as a PRRSV attachment and internalization receptor via clathrin-mediated endocytosis [53,58]. The heparan sulphate enhances the interaction between the virus and sialoadhesin, but is not necessary, and the M/GP5 complex is involved in interacting with heparan and the N-terminal domain of pSn [15,16]. Upon internalization with the participation of pSn, the genome of the virus that is present in the early endosome is released into the cytoplasm [53]. The last stage is critically dependent on CD163, a scavenger receptor cysteine-rich (SRCR) family for hemoglobin clearance, which is the most specific and indispensable receptor for PRRSV entry and infection both in vitro and in vivo [8,12,59]. Additionally, the viral GP2 and GP4 glycoproteins bind to CD163 [13]. Moreover, a study has shown that CD163 interacts with GP3 and GP5 in addition to the known interactions with GP4 and GP2, and co-immunoprecipitation (co-IP) analysis indicated that the SRCR5-domain deletion of CD163 loses its interaction with viral GP2, GP3, and GP5, and thus blocks virus uncoating in the early endosomes [17]. A growing body of research has also indicated that CD163 plays an essential role in viral uncoating and genome release, and CD163 knockout pigs are resistant to PRRSV infection [13,59,60,61]. In addition to CD163, cellular proteases, such as the aspartic protease cathepsin E and trypsin-like serine proteases, have been implicated in the uncoating process and subsequent infection [62]. Moreover, there are some mediators involved in the entry process of PRRSV into the macrophage. CD151 interacting with the PRRSV-2 3′ untranslated region (UTR) RNA should cooperate in infection in vitro [18]. Simian vimentin bound to PRRSV-2 nucleocapsid protein is involved in mediating transportation of the virus in the cytosol [19]. CD209 (DC-SIGN) and MYH9 interacting with GP5 are essential factors in both PRRSV-1 and PRRSV-2 entry and infection [20,21,22].
The genome released by the virus was identified as the template used to translate into pp1a and pp1ab, and after that the proteases hydrolyze and cleave polyproteins into mature nonstructural proteins as described above [63]. The viral replication and transcription complex (RTC) is assembled by nsps, whose key components are nsp9 and nsp10 [64]. The RTC first engages in producing both full-length and subgenome (sg)-length minus strands using a mechanism of discontinuous transcription, the latter serving as templates for the synthesis of plus-strand sg mRNAs required to express the structural protein genes, which reside in the 3′-proximal quarter of the genome [8,65,66]. The last stage is assembling and releasing the virion. Novel generated RNA genomes are packaged into nucleocapsids that become enveloped virions by structural proteins through budding from smooth intracellular membranes, and the new virions are released from the cell in the form of exocytosis [8,65]. The entire process is illustrated in Figure 1.
3. The PRRSV–Host Interactions
The complex networks of virus–host interactions are essential in the overall process of PRRSV entry and infection. The viral structural proteins interact with host receptors to mediate viral entry as described above. The interactions of host and viral nonstructural proteins can exert an influence on the replication and transcription of viral genomes. The virus’s invasion causes a series of immune responses, and then virions escape from the host immune system to favor their own replication by interacting with the host. Only by better understanding the molecular mechanism of PRRSV immune evasion and modulation can we design more effective vaccines. Further studies of these host–virus interactions are critical to the development of novel antiviral strategies against PRRSV.
3.1. Interferons (IFNs)
Innate immunity is the first line of defense without memory and specificity, and interferons (IFNs) are one of the earliest cytokines and major elements of the host fighting with virus invasion, as shown in Figure 1 [13,67]. PRRSV-2 is sensitive to type I interferons (IFN-α/β), and has some immunosuppressive mechanisms of suppressing IFNs [13,68]. Activation of IFN regulatory factor 3 (IRF3) and nuclear factor-κB (NF-κB) plays an important role in activating the IFN-β promoter, and these factors bind to the IFN-β promoter to form an enhanceosome via the cAMP response element-binding (CREB)-binding protein (CBP) transcriptional co-activator [23]. This also suggests that the virus has evolved to assuage the host’s innate immunity [13]. The mechanism of suppressing IFNs by blocking the activation of the IRF3 or NF-κB is an important strategy to respond to the innate immunity of the host to the virus’s proteins, such as nsp1, nsp2, and nsp4 [27,28]. Some studies have revealed that IFNs are suppressed by the PRRSV-2′s nsp1, which is a potent IFN antagonist [24]. The zinc finger 1 motif of nsp1α is essential in suppressing IFNs by inducing CBP degradation to inhibit the recruitment of CBP for enhanceosome assembly, which is likely the key mechanism in IFN suppression [23,24]. Moreover, nsp1β inhibits both IRF3- and NF-κB-dependent gene induction via dsRNA and the Sendai virus, resulting in IFN suppression [25]. Similarly, the PRRSV-2 nucleocapsid (N) protein inhibits the phosphorylation and nuclear translocation of IRF3 to suppress IFNβ induction [26]. Although nsp11 endoribonuclease (NendoU) activity inhibits IFNβ by suppressing IRF3 and NF-κB activation, binding to IRF9 controls the formation and nuclear translocation of the IFN-stimulated gene factor 3 (ISGF3) and antagonizes type I IFN signaling in a NendoU activity-independent manner [29,30,31].
3.2. Interleukin (IL)
In highly pathogenic (HP) PRRSV-infected swine, the cytokines including interleukin (IL)-1, IL-6, IL-8, IL-10, and tumor necrosis factor (TNF)-α are significantly increased [69,70]. The PRRSV-2 envelope protein E remarkably increases the release of IL-1β from lipopolysaccharide (LPS)-primed PAMs [32]. Studies showed that nsp11 NendoU activity plays a key role in inhibiting the secretion of IL-1β [33]. IL-10 is a significant immunosuppression cytokine with anti-inflammatory properties that can counteract adaptive immunity, thereby preventing impairment to the host [71,72]. A previous study has demonstrated that the PRRSV-2 N protein can up-regulate IL-10 via NF-κB and p38 mitogen-activated protein kinase (MAPK) pathways in PAMs [73]. Additionally, in porcine monocyte-derived dendritic cells (MoDCs) and PAMs, the N protein can promote the expression of IL-10 [34]. The GP5 could significantly increase IL-10 production through p38 MAPK and signal transducer and activator of transcription-3 (STAT3) activation [36]. Similarly, nsp1 can increase the level of IL-10 as an inducer in vivo [35]. IL-17 is a proinflammatory cytokine associated with intense inflammation and is upregulated by HP-PRRSV of genotype 2 infection [13,37]. A study found that HP-PRRSV nsp11 could induce IL-17 production depending on the phosphatidylinositol 3-kinase (PI3K)-p38MAPK-C/EBPβ/CREB pathways [37].
3.3. Tripartite Motif (TRIM) Proteins
Tripartite motif (TRIM) proteins, as critical components of the innate immune system, play significant roles in fighting virus invasion for mammalian cells, and can regulate multiple cellular processes including transcription-dependent antiviral responses such as cytokine-mediated or autophagy-mediated antiviral modulation [40,74,75]. The N-terminal RING-type zinc finger domain of TRIM proteins confers E3 ubiquitin ligase activity [74]. Study has indicated that the PRRSV-2 N protein can antagonize the antiviral activity of TRIM25 and suppress innate immune responses of the host by competitively interacting with TRIM25, thereby interfering with TRIM25-mediated retinoic acid-inducible gene I (RIG-I) ubiquitination and inhibiting IFN-β production [38]. TRIM22 can interact with the PRRSV-2 N protein and reduce virus replication, and the SPla and the RYanodine Receptor (SPRY) domain and nuclear localization signal of TRIM22 are indispensable for this interaction [39]. Moreover, the N-terminal RING domain of TRIM59, which is an important antiviral component, can interact with the C-terminal NendoU domain of nsp11, thereby inhibiting PRRSV-2 infection [40].
3.4. MicroRNAs (miRNAs)
MicroRNAs (miRNAs) are small non-coding RNA molecules containing about 21 nucleotides in length, and they play key roles in the complex networks of PRRSV–host interactions [76,77]. More recently, miRNAs have been considered as crucial post-transcriptional gene regulators for viral replication and host immune responses in the process of PRRSV invasion [13,77,78]. This research had indicated that MicroRNA can target signaling pathways or host factors both related to PRRSV replication [5]. MiR-30c has a negative effect on the IFN-I response by targeting Janus kinase 1 (JAK1) to facilitate HP-PRRSV of genotype 2 infection [79]. On the contrary, miR-181 can suppress PRRSV-2 infection by targeting and down-regulating the PRRSV-2 receptor CD163 [41]. MiR-10a-5p can interact with 3′ UTR of pig SRP14 mRNA and reduce SRP14 expression through translational repression, thereby inhibiting PRRSV-1 replication [80]. Additionally, miRNAs also can target the PRRSV genome directly, and this is a new perspective on controlling PRRSV infection [81]. In addition to targeting the 3′ UTR of CD163 mRNA, MiR-181 can combine with ORF4, which is a highly conserved region of PRRSV-2 genomic RNA, and thus inhibits PRRSV-2 replication [41,42]. MiR-130 can directly target the HP-PRRSV 5′ UTR and exert antiviral activity. Further study has reported that miR-130 has an effect on inhibiting the replication of PRRSV-2 strains, but not the replication of classical PRRSV-1 strains [13,43]. MiR-23 and miR-378, which were identified as critical inhibitors of PRRSV-2 replication, can directly target PRRSV genomic ORF3 and ORF7, respectively [44]. Interestingly, miR-505, also as a critical inhibitor of PRRSV-2 replication, can directly target both ORF3 and ORF5.
3.5. Other Host Factors’ Interactions with PRRSV
PRRSV depends on host factors to complete genome replication and interacts with host molecules for its survival and reproduction. The host immune system will take some measures to suppress the virus replication during virus invasion. Besides the abovementioned host factors, other cellular components are also involved in interacting with PRRSV. The cellular protein nucleoporin 62 (Nup62) interacts with nsp1β, leading to inhibition of host antiviral protein expression, revealing a new strategy of immune escape [47]. In addition, research found that cellular retinoblastoma protein (pRb) interacts with the nsp9 of genotype 2 PRRSV, which will benefit the replication of PRRSV-2 [49]. Inversely, the leucine-rich repeats (LRR) domain of nucleotide-binding oligomerization domain-like receptor (NLR) X1 as a new host restriction factor interacts with the RdRp domain of PRRSV-2 nsp9, which restricts PRRSV-2 replication [50]. Similarly, the interaction of the zinc finger domain of ZAP, a zinc finger antiviral protein, and nsp9 mapped to amino acids 150 to 160, can repress PRRSV-2 replication [51]. Nsp1α can combine with the protein inhibitor of activated STAT1 (PIAS1) as the small ubiquitin-related modifier (SUMO) E3 ligase leading to the nsp1α sumoylation [45]. In virtue of the SUMO E3 ligase activity, nsp1α interacts with swine leukocyte antigen class I (SLA-I) to modulate degradation, in the same way facilitating the ubiquitinylation of CBP for degradation [45,46]. The nsp5 of PRRSV-1 and PRRSV-2 was shown to promote the degradation of signal transducer and activator of transcription 3 (STAT3), a pleiotropic signaling mediator of numerous cytokines, leading to interference with the JAK/STAT3 signaling and the host immune responses [48]. The PRRSV-2 N protein interacts with RNA-associated nuclear host proteins such as fibrillarin, nucleolin, and poly(A)-binding protein, but the specific function remains to be further clarified [52]. Furthermore, the PRRSV N protein appears to up-regulate NF-κB activation that is attributed to the N protein nuclear localization signal [45].
In conclusion, a complex network of PRRSV–host interactions exists throughout the virus cycle, including virus entry, replication, and infection, as shown in Table 1. The immune response caused by different virus genotypes is emphasized in Table 1. Although significant advancements have been made, the understanding of direct or indirect virus–host interaction networks remain limited. Moreover, some seemingly contradictory results are hard to explain. Study indicated that the NF-κB is stimulated by the N protein, which may up-regulate IFN [45]. However, the N protein inhibits the phosphorylation and nuclear translocation of IRF3 to suppress IFNβ induction [26]. The stimulation of NF-κB by the N protein may involve other cellular pathways, and the mechanism of cytokines’ regulation by the N protein needs to be further explored. The interaction of PRRSV nsp9, a critical component of the viral RTC, and some host proteins can inhibit virus replications such as NLRX1 and ZAP, and yet they can promote virus replications such as pRb, which has aroused much concern.
4. Current Antiviral Strategies and Prospects
Since the outbreak of PRRSV in the 1980s, efforts have been made vehemently to find effective antiviral strategies and vaccines to control its damage to the swine industry. However, for a variety of reasons, this has become a huge challenge. Modified-live virus (MLV) vaccines and inactivated vaccines against PRRSV are available and used worldwide [56]. Both non-structural and structural proteins are targets of PRRSV modified attenuation [82]. It was found that nsp2, nsp3, nsp10, GP2 and GP5 are major proteins involved in virus attenuation [82]. Particularly, nsp2 and GP5 are the most variable PRRSV proteins, and mutations generally occurred in the hypervariable regions of these two proteins [83]. For instance, HP-PRRSV MLV TJM-F92 has a unique 120-amino acid deletion located in the 628–747 residues of nsp2 [83,84]. The CH-1R MLV vaccines against classical PRRSV strains were developed from the North American strains CH-1a [85]. A total of 54 mutated amino acids were found in CH-1R; among them, 11 and three mutations are located in nsp2 and GP5, respectively [82]. PRRS-MLV can protect homologous viruses and mitigate lung damage but does not induce sufficient heterologous immunity [86]. Compared with PRRSV-1 MLV vaccines, PRRSV-2 MLV vaccines are more effective against both PRRSV-1 and PRRSV-2 infection [87,88]. In addition to the issue of immunity breadth to fight new outbreaks, MLV may revert to virulence, and safety issues have aroused concerns [89]. The safety of inactivated vaccines is sufficient, but their immunogenicity is poor [90]. The combination of immune adjuvant and inactivated vaccines can enhance vaccine efficacy [89]. It has been reported that intranasal delivery of a poly(lactic-co-glycolic acid) (PLGA) nanoparticle (NP)-entrapped inactivated PRRSV vaccine has the potential to induce a broadly cross-protective immune response [90]. In addition, the intranasal Th1-biased adjuvant of the recombinant B subunit of the Escherichia coli heat-labile enterotoxin rLTB can significantly strengthen cellular immune responses [89]. Whether subunit vaccines, DNA vaccines or virus vectored vaccines can replace MLV remains to be further studied [56]. Moreover, the rapid mutation of this RNA virus, which leads to a high degree of genetic and antigenic variation, challenges the design of PRRSV vaccine. Thus, there is an urgent need to improve upon the existing vaccines, which mainly refers to the enhancement of the breadth of immunity and creating cross-protective virus vaccines [91]. PRRSV nsp4- or nsp9-specific nanobodies can inhibit PRRSV replication by blocking these nsps, which are critical components of the viral RTC, therefore interfering with viral genome replication and transcription [92,93]. There are some relatively conserved nonstructural proteins, such as nsp1, nsp4, nsp9, and nsp11, that play important roles in the life cycle of viral infection according to our summary of the PRRSV–host interaction network, and nanobodies/inhibitors that target these proteins offer a novel perspective on antiviral strategies that may potentially resist PRRSV infection across genotypes. According to protein data bank (PDB) information, the structures of nsp1α (PDB code: 3IFU), nsp1β (PDB code: 3MTV), nsp10 (PDB code: 6JDU), and nsp11 (PDB code: 5EYI), provide structural basics for the study of biological function and action mechanism [94,95,96,97,98]. For example, the C-terminal extension of nsp1β binds to the putative substrate binding site of the papain-like cysteine protease domain, which illustrates the role of the substrate binding mode and provides a structural template to design nsp1 inhibitors with potential therapeutic value [96].
Classically attenuated viruses are created by passaging viruses in cultured cells and are effective for many viruses. However, many safety issues exist for these empiric attenuations [99]. Furthermore, one major drawback of the current live-attenuated PRRSV vaccines is that they cannot distinguish PRRSV-specific antibodies produced by natural infection with wild type (WT) viruses or by vaccination [100]. Therefore, a differentiating infected from vaccinated animals (DIVA) vaccine is extremely useful to survey and eradicate PRRSV, and has become an important aim of many current research efforts [101]. Epitope-M201, which is highly immunodominant and well-conserved among PRRSV-2 isolates, is located at the carboxyl terminus (residues 161–174) of the viral M protein, and provides a serologic marker for the development of live-attenuated DIVA vaccines against PRRSV-2 [100]. A2MC2-P90, which has a 181 residue deletion in nsp2 and 35 nucleotide mutations throughout the genome, could serve as an ideal and significant backbone for DIVA vaccine development due to its unique properties including the ability to induce IFN, a virulence in swine and its largest spontaneous deletion in nsp2 [56,102]. Advances in viral structures have provided new ways of controlling viral replication and virulence. These MLV vaccines may lead to a new generation of safer, more widely applicable PRRSV vaccines. As shown in Table 1, PRRSV structural envelope proteins play vital roles in the viral infection cycle. Unfortunately, the structures of these proteins are not available, making it difficult to understand the structural basis and molecular mechanisms of the virus–receptor interaction or antibody-mediated neutralization, so as to guide the optimization of vaccines and to improve immunogenicity [103]. However, a structure-based vaccine design approach can be used to improve the antigenicity and to create immunogens capable of eliciting robust neutralizing and protective immune responses at the atomic level. Furthermore, structure-based specific MLV design can be used to generate many kinds of MLVs without the risk of reverting to virulence.
Many viral proteins, such as nsp1α, nsp1β, nsp2, nsp4, nsp7, nsp10, and nsp11, play an essential role in suppressing the host innate immune response. These key proteins can be used as antiviral targets for attenuated vaccines. For example, the nsp4 D185N mutant exhibits a slower replication rate and a higher ability to induce IFN-I expression compared with WT PRRSV [104]. Additionally, based on the 3D structure of nsp2, mutated viruses lacking deubiquitinating enzyme activity exhibit WT replication kinetics, but strikingly enhance innate immune signaling [105]. The IFN-β mRNA levels are increased more than 10-fold compared with those in the WT virus-infected cells. These modified viruses display a significantly weakened ability to evade host immune responses, opening new possibilities for developing improved attenuated virus vaccines against economically important arteriviruses. Moreover, PRRSV nsp11 inhibits NF-κB signaling using its deubiquitinating activity [33]. Thus, selective mutations of the conserved sites C112 and Y219 of deubiquitinating activity will greatly increase host innate immune responses without affecting endoribonuclease activity. These mutations can be effectively applied to live attenuated PRRSV vaccines, thus providing the basis for a DIVA vaccine. Moreover, the combination of specific MLVs and immune adjuvants can further enhance vaccine efficacy.
Furthermore, the identification of miRNAs that target the PRRSV genome provides alternative targets for gene editing [5]. With the rapid development of CRISPR/Cas-based gene editing technology, the acquisition of gene-edited pigs is no longer a difficult task. In this context, the TRIM22 gene has been lost during evolution, and restoring TRIM22 through gene editing will be a potential antiviral strategy [39]. Furthermore, gene-editing of CD163, which has been determined to be the major receptor, generates pigs resisting PRRSV infection [61]. Notably, the crystal structure of the SRCR5 domain of CD163 (PDB code: 5JFB) consists of seven β-strands and two α-helices, which extends our understanding of the mechanism of PRRSV invasion [103]. Arg561, which is located in the long loop region, is an important residue for PRRSV invasion and may play a key role in the interaction between CD163 and PRRSV during viral infection, which provides a target for drug design and gene editing. However, the acceptance of genetically modified organisms as food challenges the development of these antiviral strategies, and thus biosafety should be increasingly emphasized in the development of new virus vaccines.
Subunit peptide vaccines reproducing the B and T epitopes are a novel antivirus strategy [106]. Cytotoxic T lymphocytes (CTLs) epitope vaccines have superiorities in terms of specificity, safety and clinical effect, and have been successfully used to fight viral infections, such as human immunodeficiency virus (HIV), human papilloma virus (HPV), and dengue virus [107,108,109]. The presentation of viral epitopes to CTLs by SLA-1 plays a key role in swine immunity [110]. The crystal structure of peptide-SLA-1*1502 (pSLA-1*1502) complexes with one peptide (nsp9-TMP9) (PDB code: 5YLX) provides the structural conformation basis of peptide presentation, and the overall structure is very similar to those of pSLA-1*0401 (PDB code: 3QQ3) and pSLA-3*hs0202 (PDB code: 5H94) [110,111,112]. The structural information displayed by the D pocket, as shown in Figure 2, plays a crucial part in determining and fixing the bound peptides, and provides a structural basis for designing effective peptide vaccines [111]. Study indicated that RdRp has the maximum number of conserved peptides by identifying SLA-1*1502-restricted potential epitopes peptides from whole genomes of different PRRSV strains; thus nsp9/10/11 (especially RdRp) may be the best targets for designing PRRSV vaccines to induce a CTL response to genetically heterologous strains [111]. Although the monomer structures of PRRSV nsp10 (PDB code: 6JDU) and nsp11 (PDB code: 5EYI) have been successfully solved, the structure and biological functional mechanisms of nsp9 remain elusive [95,98]. Table 1 shows the important protein structures and PDB codes that have been obtained in the PRRSV–host interaction network. Therefore, we are trying to explore the monomer or complex structure of nsp9, so as to provide a structural basis and new insight for treatments against PRRSV.
The review of the PRRSV–host interactions network helps us to understand the function of critical host factors involved in virus infection. More research is still needed to complete this complex network and provide the basis for the development of antiviral drugs and vaccines.
5. Conclusions
PRRSV is undoubtedly one of the most economically devastating swine pathogen, and has aroused sustained concern. In spite of constant efforts of the pathogenicity and immunology of PRRSV, it has always been a substantial challenge to design effective vaccines and drugs. MLV vaccines exist some safety issues. Compared with MLV vaccines, DIVA vaccines can distinguish PRRSV-specific antibodies produced by natural infection with WT viruses or by vaccination, and has become an important aim of many current research efforts. The acquisition of gene-edited pigs will be a potential antiviral strategy, but it’s a problem in acceptability of genetically modified foods. Structural information allows us to better understand the mechanisms of viral infection. A structure-based vaccine design approach may improve the antigenicity and create immunogens capable of eliciting robust neutralizing and protective immune responses.
The complex network of PRRSV–host interactions exists throughout the virus cycle. Only by studying the function of the PRRSV–host interactions in the overall process of PRRSV entry and infection, can we understand mechanisms of PRRSV replication and propagation, thereby providing the basis for antiviral strategies. Some host factors and viral proteins play important roles in the PRRSV–host network, but lack of structural information prevents us from understanding the mechanisms of infection thoroughly, Therefore, these host factors and viral proteins need to be explored to provide structural bases and new insights for treatments against PRRSV.
Author Contributions
Conceptualization and formal analysis, J.M. and Z.C.; writing—original draft preparation, J.M. and Z.C.; writing—review and editing, J.M., L.M., M.Y., W.W., W.F. and Z.C.; supervision, project administration, and funding acquisition, Z.C. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported financially by National Natural Science Foundation of China [32071210], National Key Research and Development Program of China [2018YFE0113100].
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Bloemraad, M.; de Kluijver, E.P.; Petersen, A.; Burkhardt, G.E.; Wensvoort, G. Porcine reproductive and respiratory syndrome: Temperature and pH stability of Lelystad virus and its survival in tissue specimens from viraemic pigs. Vet. Microbiol. 1994, 42, 361–371. [Google Scholar] [CrossRef]
- Renken, C.; Nathues, C.; Swam, H.; Fiebig, K.; Weiss, C.; Eddicks, M.; Ritzmann, M.; Nathues, H. Application of an economic calculator to determine the cost of porcine reproductive and respiratory syndrome at farm-level in 21 pig herds in Germany. Porc. Health Manag. 2021, 7, 12. [Google Scholar] [CrossRef]
- Holtkamp, D.J.; Kliebenstein, J.B.; Neumann, E.; Zimmerman, J.J.; Rotto, H.; Yoder, T.K.; Wang, C.; Yeske, P.; Mowrer, C.L.; Haley, C.A. Assessment of the economic impact of porcine reproductive and respiratory syndrome virus on United States pork producers. J. Swine Health Prod. 2013, 21, 72–84. [Google Scholar]
- Lalonde, C.; Provost, C.; Gagnona, C.A. Whole-Genome Sequencing of Porcine Reproductive and Respiratory Syndrome Virus from Field Clinical Samples Improves the Genomic Surveillance of the Virus. J. Clin. Microbiol. 2020, 58, e00097-20. [Google Scholar] [CrossRef]
- Du, T.; Nan, Y.; Xiao, S.; Zhao, Q.; Zhou, E.M. Antiviral Strategies against PRRSV Infection. Trends Microbiol. 2017, 25, 968–979. [Google Scholar] [CrossRef] [PubMed]
- Balka, G.; Wang, X.; Olasz, F.; Balint, A.; Kiss, I.; Banyai, K.; Rusvai, M.; Stadejek, T.; Marthaler, D.; Murtaugh, M.P.; et al. Full genome sequence analysis of a wild, non-MLV-related type 2 Hungarian PRRSV variant isolated in Europe. Virus Res. 2015, 200, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Johnson, C.R.; Griggs, T.F.; Gnanandarajah, J.; Murtaugh, M.P. Novel structural protein in porcine reproductive and respiratory syndrome virus encoded by an alternative ORF5 present in all arteriviruses. J. Gen. Virol. 2011, 92, 1107–1116. [Google Scholar] [CrossRef] [PubMed]
- Snijder, E.J.; Kikkert, M.; Fang, Y. Arterivirus molecular biology and pathogenesis. J. Gen. Virol. 2013, 94, 2141–2163. [Google Scholar] [CrossRef]
- Li, Y.; Tas, A.; Snijder, E.J.; Fang, Y. Identification of porcine reproductive and respiratory syndrome virus ORF1a-encoded non-structural proteins in virus-infected cells. J. Gen. Virol. 2012, 93, 829–839. [Google Scholar] [CrossRef]
- Wissink, E.H.J.; Kroese, M.V.; Maneschijn-Bonsing, J.G.; Meulenberg, J.J.M.; van Rijn, P.A.; Rijsewijk, F.A.M.; Rottier, P.J.M. Significance of the oligosaccharides of the porcine reproductive and respiratory syndrome virus glycoproteins GP2a and GP5 for infectious virus production. J. Gen. Virol. 2004, 85, 3715–3723. [Google Scholar] [CrossRef]
- Wissink, E.H.; Kroese, M.V.; van Wijk, H.A.; Rijsewijk, F.A.; Meulenberg, J.J.; Rottier, P.J. Envelope protein requirements for the assembly of infectious virions of porcine reproductive and respiratory syndrome virus. J. Virol. 2005, 79, 12495–12506. [Google Scholar] [CrossRef] [Green Version]
- Veit, M.; Matczuk, A.K.; Sinhadri, B.C.; Krause, E.; Thaa, B. Membrane proteins of arterivirus particles: Structure, topology, processing and function. Virus Res. 2014, 194, 16–36. [Google Scholar] [CrossRef] [PubMed]
- An, T.Q.; Li, J.N.; Su, C.M.; Yoo, D. Molecular and Cellular Mechanisms for PRRSV Pathogenesis and Host Response to Infection. Virus Res. 2020, 286, 197980. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Lobo, F.J.; Díez-Fuertes, F.; Segalés, J.; García-Artiga, C.; Simarro, I.; Castro, J.M.; Prieto, C. Comparative pathogenicity of type 1 and type 2 isolates of porcine reproductive and respiratory syndrome virus (PRRSV) in a young pig infection model. Vet. Microbiol. 2011, 154, 58–68. [Google Scholar] [CrossRef] [PubMed]
- Delputte, P.L.; Vanderheijden, N.; Nauwynck, H.J.; Pensaert, M.B. Involvement of the matrix protein in attachment of porcine reproductive and respiratory syndrome virus to a heparinlike receptor on porcine alveolar macrophages. J. Virol. 2002, 76, 4312–4320. [Google Scholar] [CrossRef] [Green Version]
- Van Breedam, W.; Van Gorp, H.; Zhang, J.Q.; Crocker, P.R.; Delputte, P.L.; Nauwynck, H.J. The M/GP(5) glycoprotein complex of porcine reproductive and respiratory syndrome virus binds the sialoadhesin receptor in a sialic acid-dependent manner. PLoS Pathog. 2010, 6, e1000730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, P.; Wei, R.; Dong, W.; Zhu, Z.; Zhang, X.; Chen, Y.; Liu, X.; Guo, C. CD163(DeltaSRCR5) MARC-145 Cells Resist PRRSV-2 Infection via Inhibiting Virus Uncoating, which Requires the Interaction of CD163 with Calpain 1. Front. Microbiol. 2019, 10, 3115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shanmukhappa, K.; Kim, J.K.; Kapil, S. Role of CD151, A tetraspanin, in porcine reproductive and respiratory syndrome virus infection. Virol. J. 2007, 4, 62. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.K.; Fahad, A.M.; Shanmukhappa, K.; Kapil, S. Defining the cellular target(s) of porcine reproductive and respiratory syndrome virus blocking monoclonal antibody 7G10. J. Virol. 2006, 80, 689–696. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.W.; Dryman, B.A.; Li, W.; Meng, X.J. Porcine DC-SIGN: Molecular cloning, gene structure, tissue distribution and binding characteristics. Dev. Comp. Immunol. 2009, 33, 464–480. [Google Scholar] [CrossRef]
- Gao, J.; Xiao, S.; Xiao, Y.; Wang, X.; Zhang, C.; Zhao, Q.; Nan, Y.; Huang, B.; Liu, H.; Liu, N.; et al. MYH9 Is an Essential Factor for Porcine Reproductive and Respiratory Syndrome Virus Infection. Sci. Rep. 2016, 6, 25120. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Xue, B.; Sun, W.; Gu, G.; Hou, G.; Zhang, L.; Wu, C.; Zhao, Q.; Zhang, Y.; Zhang, G.; et al. Recombinant MYH9 protein C-terminal domain blocks porcine reproductive and respiratory syndrome virus internalization by direct interaction with viral glycoprotein 5. Antivir. Res. 2018, 156, 10–20. [Google Scholar] [CrossRef]
- Kim, O.; Sun, Y.; Lai, F.W.; Song, C.; Yoo, D. Modulation of type I interferon induction by porcine reproductive and respiratory syndrome virus and degradation of CREB-binding protein by non-structural protein 1 in MARC-145 and HeLa cells. Virology 2010, 402, 315–326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, M.; Du, Y.; Song, C.; Yoo, D. Degradation of CREB-binding protein and modulation of type I interferon induction by the zinc finger motif of the porcine reproductive and respiratory syndrome virus nsp1α subunit. Virus Res. 2013, 172, 54–65. [Google Scholar] [CrossRef] [PubMed]
- Beura, L.K.; Sarkar, S.N.; Kwon, B.; Subramaniam, S.; Jones, C.; Pattnaik, A.K.; Osorio, F.A. Porcine reproductive and respiratory syndrome virus nonstructural protein 1beta modulates host innate immune response by antagonizing IRF3 activation. J. Virol. 2010, 84, 1574–1584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sagong, M.; Lee, C. Porcine reproductive and respiratory syndrome virus nucleocapsid protein modulates interferon-beta production by inhibiting IRF3 activation in immortalized porcine alveolar macrophages. Arch. Virol. 2011, 156, 2187–2195. [Google Scholar] [CrossRef]
- Li, H.; Zheng, Z.; Zhou, P.; Zhang, B.; Shi, Z.; Hu, Q.; Wang, H. The cysteine protease domain of porcine reproductive and respiratory syndrome virus non-structural protein 2 antagonizes interferon regulatory factor 3 activation. J. Gen. Virol. 2010, 91, 2947–2958. [Google Scholar] [CrossRef]
- Chen, Z.; Li, M.; He, Q.; Du, J.; Zhou, L.; Ge, X.; Guo, X.; Yang, H. The amino acid at residue 155 in nonstructural protein 4 of porcine reproductive and respiratory syndrome virus contributes to its inhibitory effect for interferon-beta transcription in vitro. Virus Res. 2014, 189, 226–234. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Ke, H.; Han, M.; Chen, N.; Fang, W.; Yoo, D. Nonstructural Protein 11 of Porcine Reproductive and Respiratory Syndrome Virus Suppresses Both MAVS and RIG-I Expression as One of the Mechanisms to Antagonize Type I Interferon Production. PLoS ONE 2016, 11, e0168314. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; Wang, L.; Li, X.; Zhang, G.; Guo, J.; Zhao, D.; Chai, S.; Deng, R. Endoribonuclease activities of porcine reproductive and respiratory syndrome virus nsp11 was essential for nsp11 to inhibit IFN-beta induction. Mol. Immunol. 2011, 48, 1568–1572. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Chen, J.; Yu, C.; Zhu, X.; Xu, S.; Fang, L.; Xiao, S. Porcine Reproductive and Respiratory Syndrome Virus nsp11 Antagonizes Type I Interferon Signaling by Targeting IRF9. J. Virol. 2019, 93, e00623-19. [Google Scholar] [CrossRef] [Green Version]
- Zhang, K.; Hou, Q.; Zhong, Z.; Li, X.; Chen, H.; Li, W.; Wen, J.; Wang, L.; Liu, W.; Zhong, F. Porcine reproductive and respiratory syndrome virus activates inflammasomes of porcine alveolar macrophages via its small envelope protein E. Virology 2013, 442, 156–162. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Shi, X.; Zhang, X.; Wang, A.; Wang, L.; Chen, J.; Deng, R.; Zhang, G. The Endoribonuclease Activity Essential for the Nonstructural Protein 11 of Porcine Reproductive and Respiratory Syndrome Virus to Inhibit NLRP3 Inflammasome-Mediated IL-1β Induction. DNA Cell Biol. 2015, 34, 728–735. [Google Scholar] [CrossRef] [PubMed]
- Wongyanin, P.; Buranapraditkul, S.; Yoo, D.; Thanawongnuwech, R.; Roth, J.A.; Suradhat, S. Role of porcine reproductive and respiratory syndrome virus nucleocapsid protein in induction of interleukin-10 and regulatory T-lymphocytes (Treg). J. Gen. Virol. 2012, 93, 1236–1246. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Bai, J.; Li, Y.; Wang, X.; Wang, X.; Jiang, P. Suppression of immune responses in pigs by nonstructural protein 1 of porcine reproductive and respiratory syndrome virus. Can. J. Vet. Res. 2012, 76, 255–260. [Google Scholar] [PubMed]
- Hou, J.; Wang, L.; Quan, R.; Fu, Y.; Zhang, H.; Feng, W.-h. Induction of interleukin-10 is dependent on p38 mitogen-activated protein kinase pathway in macrophages infected with porcine reproductive and respiratory syndrome virus. Virol. J. 2012, 9, 165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Du, L.; Liu, F.; Wei, Z.; Gao, L.; Feng, W.H. Highly Pathogenic Porcine Reproductive and Respiratory Syndrome Virus Induces Interleukin-17 Production via Activation of the IRAK1-PI3K-p38MAPK-C/EBPbeta/CREB Pathways. J. Virol. 2019, 93, e01100-19. [Google Scholar] [CrossRef] [Green Version]
- Zhao, K.; Li, L.W.; Jiang, Y.F.; Gao, F.; Zhang, Y.J.; Zhao, W.Y.; Li, G.X.; Yu, L.X.; Zhou, Y.J.; Tong, G.Z. Nucleocapsid protein of porcine reproductive and respiratory syndrome virus antagonizes the antiviral activity of TRIM25 by interfering with TRIM25-mediated RIG-I ubiquitination. Vet. Microbiol. 2019, 233, 140–146. [Google Scholar] [CrossRef]
- Jing, H.; Tao, R.; Dong, N.; Cao, S.; Sun, Y.; Ke, W.; Li, Y.; Wang, J.; Zhang, Y.; Huang, H.; et al. Nuclear localization signal in TRIM22 is essential for inhibition of type 2 porcine reproductive and respiratory syndrome virus replication in MARC-145 cells. Virus Genes 2019, 55, 660–672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jing, H.; Ke, W.; Tao, R.; Li, Y.; Zhao, Y.; Cao, S.; Sun, Y.; Wang, J.; Zhang, Y.; Dong, W.; et al. TRIM59 inhibits porcine reproductive and respiratory syndrome virus (PRRSV)-2 replication in vitro. Res. Vet. Sci. 2019, 127, 105–112. [Google Scholar] [CrossRef]
- Gao, L.; Guo, X.k.; Wang, L.; Zhang, Q.; Li, N.; Chen, X.x.; Wang, Y.; Feng, W.h. MicroRNA 181 Suppresses Porcine Reproductive and Respiratory Syndrome Virus (PRRSV) Infection by Targeting PRRSV Receptor CD163. J. Virol. 2013, 87, 8808–8812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, X.K.; Zhang, Q.; Gao, L.; Li, N.; Chen, X.X.; Feng, W.H. Increasing expression of microRNA 181 inhibits porcine reproductive and respiratory syndrome virus replication and has implications for controlling virus infection. J. Virol. 2013, 87, 1159–1171. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Gao, F.; Jiang, Y.; Yu, L.; Zhou, Y.; Zheng, H.; Tong, W.; Yang, S.; Xia, T.; Qu, Z.; et al. Cellular miR-130b inhibits replication of porcine reproductive and respiratory syndrome virus in vitro and in vivo. Sci. Rep. 2015, 5, 17010. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Guo, X.K.; Gao, L.; Huang, C.; Li, N.; Jia, X.; Liu, W.; Feng, W.H. MicroRNA-23 inhibits PRRSV replication by directly targeting PRRSV RNA and possibly by upregulating type I interferons. Virology 2014, 450–451, 182–195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Y.; Han, M.; Kim, C.; Calvert, J.G.; Yoo, D. Interplay between Interferon-Mediated Innate Immunity and Porcine Reproductive and Respiratory Syndrome Virus. Viruses 2012, 4, 424–446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, J.; Ge, X.; Liu, Y.; Jiang, P.; Wang, Z.; Zhang, R.; Zhou, L.; Guo, X.; Han, J.; Yang, H. Targeting Swine Leukocyte Antigen Class I Molecules for Proteasomal Degradation by the nsp1alpha Replicase Protein of the Chinese Highly Pathogenic Porcine Reproductive and Respiratory Syndrome Virus Strain JXwn06. J. Virol. 2016, 90, 682–693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ke, H.; Han, M.; Kim, J.; Gustin, K.E.; Yoo, D. Porcine Reproductive and Respiratory Syndrome Virus Nonstructural Protein 1 Beta Interacts with Nucleoporin 62 To Promote Viral Replication and Immune Evasion. J. Virol. 2019, 93, e00469-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, L.; Wang, R.; Ma, Z.; Xiao, Y.; Nan, Y.; Wang, Y.; Lin, S.; Zhang, Y.-J.; Lyles, D.S. Porcine Reproductive and Respiratory Syndrome Virus Antagonizes JAK/STAT3 Signaling via nsp5, Which Induces STAT3 Degradation. J. Virol. 2016, 91, e02087-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, J.; Zhang, N.; Ge, X.; Zhou, L.; Guo, X.; Yang, H. The interaction of nonstructural protein 9 with retinoblastoma protein benefits the replication of genotype 2 porcine reproductive and respiratory syndrome virus in vitro. Virology 2014, 464–465, 432–440. [Google Scholar] [CrossRef] [Green Version]
- Jing, H.; Song, T.; Cao, S.; Sun, Y.; Wang, J.; Dong, W.; Zhang, Y.; Ding, Z.; Wang, T.; Xing, Z.; et al. Nucleotide-binding oligomerization domain-like receptor X1 restricts porcine reproductive and respiratory syndrome virus-2 replication by interacting with viral Nsp9. Virus Res. 2019, 268, 18–26. [Google Scholar] [CrossRef]
- Zhao, Y.; Song, Z.; Bai, J.; Liu, X.; Nauwynck, H.; Jiang, P. ZAP, a CCCH-Type Zinc Finger Protein, Inhibits Porcine Reproductive and Respiratory Syndrome Virus Replication and Interacts with Viral Nsp9. J. Virol. 2019, 93, 23. [Google Scholar] [CrossRef] [Green Version]
- Yoo, D.; Wootton, S.K.; Li, G.; Song, C.; Rowland, R.R. Colocalization and Interaction of the Porcine Arterivirus Nucleocapsid Protein with the Small Nucleolar RNA-Associated Protein Fibrillarin. J. Virol. 2003, 77, 12173–12183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Breedam, W.; Delputte, P.L.; Van Gorp, H.; Misinzo, G.; Vanderheijden, N.; Duan, X.; Nauwynck, H.J. Porcine reproductive and respiratory syndrome virus entry into the porcine macrophage. J. Gen. Virol. 2010, 91, 1659–1667. [Google Scholar] [CrossRef] [PubMed]
- Duan, X.; Nauwynck, H.J.; Pensaert, M.B. Effects of origin and state of differentiation and activation of monocytes/macrophages on their susceptibility to porcine reproductive and respiratory syndrome virus (PRRSV). Arch. Virol. 1997, 142, 2483–2497. [Google Scholar] [CrossRef]
- Kreutz, L.C. Cellular membrane factors are the major determinants of porcine reproductive and respiratory syndrome virus tropism. Virus Res. 1998, 53, 121–128. [Google Scholar] [CrossRef]
- Nan, Y.; Wu, C.; Gu, G.; Sun, W.; Zhang, Y.J.; Zhou, E.M. Improved Vaccine against PRRSV: Current Progress and Future Perspective. Front. Microbiol. 2017, 8, 1635. [Google Scholar] [CrossRef]
- Nauwynck, H.J.; Duan, X.; Favoreel, H.W.; Van Oostveldt, P.; Pensaert, M.B. Entry of porcine reproductive and respiratory syndrome virus into porcine alveolar macrophages via receptor-mediated endocytosis. J. Gen. Virol. 1999, 80, 297–305. [Google Scholar] [CrossRef]
- Delputte, P.L.; Costers, S.; Nauwynck, H.J. Analysis of porcine reproductive and respiratory syndrome virus attachment and internalization: Distinctive roles for heparan sulphate and sialoadhesin. J. Gen. Virol. 2005, 86, 1441–1445. [Google Scholar] [CrossRef]
- Wang, J.; Liu, J.Y.; Shao, K.Y.; Han, Y.Q.; Li, G.L.; Ming, S.L.; Su, B.Q.; Du, Y.K.; Liu, Z.H.; Zhang, G.P.; et al. Porcine Reproductive and Respiratory Syndrome Virus Activates Lipophagy To Facilitate Viral Replication through Downregulation of NDRG1 Expression. J. Virol. 2019, 93, e00526-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Gorp, H.; Van Breedam, W.; Delputte, P.L.; Nauwynck, H.J. Sialoadhesin and CD163 join forces during entry of the porcine reproductive and respiratory syndrome virus. J. Gen. Virol. 2008, 89, 2943–2953. [Google Scholar] [CrossRef] [PubMed]
- Whitworth, K.M.; Rowland, R.R.; Ewen, C.L.; Trible, B.R.; Kerrigan, M.A.; Cino-Ozuna, A.G.; Samuel, M.S.; Lightner, J.E.; McLaren, D.G.; Mileham, A.J.; et al. Gene-edited pigs are protected from porcine reproductive and respiratory syndrome virus. Nat. Biotechnol. 2016, 34, 20–22. [Google Scholar] [CrossRef]
- Misinzo, G.M.; Delputte, P.L.; Nauwynck, H.J. Involvement of proteases in porcine reproductive and respiratory syndrome virus uncoating upon internalization in primary macrophages. Vet. Res. 2008, 39, 55. [Google Scholar] [CrossRef] [Green Version]
- Han, J.; Rutherford, M.S.; Faaberg, K.S. The porcine reproductive and respiratory syndrome virus nsp2 cysteine protease domain possesses both trans- and cis-cleavage activities. J. Virol. 2009, 83, 9449–9463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, P.; Chai, Y.; Song, J.; Liu, T.; Chen, P.; Zhou, L.; Ge, X.; Guo, X.; Han, J.; Yang, H. Reprogramming the unfolded protein response for replication by porcine reproductive and respiratory syndrome virus. PLoS Pathog. 2019, 15, e1008169. [Google Scholar] [CrossRef]
- Lunney, J.K.; Fang, Y.; Ladinig, A.; Chen, N.; Li, Y.; Rowland, B.; Renukaradhya, G.J. Porcine Reproductive and Respiratory Syndrome Virus (PRRSV): Pathogenesis and Interaction with the Immune System. Annu. Rev. Anim. Biosci. 2016, 4, 129–154. [Google Scholar] [CrossRef] [PubMed]
- Van Marle, G.; Dobbe, J.C.; Gultyaev, A.P.; Luytjes, W.; Spaan, W.J.; Snijder, E.J. Arterivirus discontinuous mRNA transcription is guided by base pairing between sense and antisense transcription-regulating sequences. Proc. Natl. Acad. Sci. USA 1999, 96, 12056–12061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pestka, S.; Krause, C.D.; Walter, M.R. Interferons, interferon-like cytokines, and their receptors. Immunol. Rev. 2004, 202, 8–32. [Google Scholar] [CrossRef] [PubMed]
- Song, Z.; Bai, J.; Liu, X.; Nauwynck, H.; Wu, J.; Liu, X.; Jiang, P. S100A9 regulates porcine reproductive and respiratory syndrome virus replication by interacting with the viral nucleocapsid protein. Vet. Microbiol. 2019, 239, 108498. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Wang, S.; Li, C.; Wang, C.; Liu, Y.; Wang, G.; He, X.; Hu, L.; Liu, Y.; Cui, M.; et al. Secondary Haemophilus parasuis infection enhances highly pathogenic porcine reproductive and respiratory syndrome virus (HP-PRRSV) infection-mediated inflammatory responses. Vet. Microbiol. 2017, 204, 35–42. [Google Scholar] [CrossRef]
- Thanawongnuwech, R.; Thacker, B.; Halbur, P.; Thacker, E.L. Increased production of proinflammatory cytokines following infection with porcine reproductive and respiratory syndrome virus and Mycoplasma hyopneumoniae. Clin. Diagn. Lab. Immunol. 2004, 11, 901–908. [Google Scholar] [CrossRef] [Green Version]
- Saraiva, M.; O’Garra, A. The regulation of IL-10 production by immune cells. Nat. Rev. Immunol. 2010, 10, 170–181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moore, K.W.; de Waal Malefyt, R.; Coffman, R.L.; O’Garra, A. Interleukin-10 and the interleukin-10 receptor. Annu. Rev. Immunol. 2001, 19, 683–765. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Liu, Y.; Zhang, Y.; Zhu, X.; Ren, S.; Guo, L.; Liu, X.; Sun, W.; Chen, Z.; Cong, X.; et al. The integrity of PRRSV nucleocapsid protein is necessary for up-regulation of optimal interleukin-10 through NF-kappaB and p38 MAPK pathways in porcine alveolar macrophages. Microb. Pathog. 2017, 109, 319–324. [Google Scholar] [CrossRef] [PubMed]
- Koepke, L.; Gack, M.U.; Sparrer, K.M. The antiviral activities of TRIM proteins. Curr. Opin. Microbiol. 2021, 59, 50–57. [Google Scholar] [CrossRef]
- Hatakeyama, S. TRIM Family Proteins: Roles in Autophagy, Immunity, and Carcinogenesis. Trends Biochem. Sci. 2017, 42, 297–311. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Feng, Y.; Yan, Y.; Zheng, Z.; Wang, W.; Zhang, Y.; Zhou, E.M.; Xiao, S. Cellular microRNA miR-c89 inhibits replication of porcine reproductive and respiratory syndrome virus by targeting the host factor porcine retinoid X receptor beta. J. Gen. Virol. 2019, 100, 1407–1416. [Google Scholar] [CrossRef]
- Krol, J.; Loedige, I.; Filipowicz, W. The widespread regulation of microRNA biogenesis, function and decay. Nat. Rev. Genet. 2010, 11, 597–610. [Google Scholar] [CrossRef]
- Dhorne-Pollet, S.; Crisci, E.; Mach, N.; Renson, P.; Jaffrézic, F.; Marot, G.; Maroilley, T.; Moroldo, M.; Lecardonnel, J.; Blanc, F.; et al. The miRNA-targeted transcriptome of porcine alveolar macrophages upon infection with Porcine Reproductive and Respiratory Syndrome Virus. Sci. Rep. 2019, 9, 3160. [Google Scholar] [CrossRef]
- Zhang, Q.; Huang, C.; Yang, Q.; Gao, L.; Liu, H.-C.; Tang, J.; Feng, W.-h. MicroRNA-30c Modulates Type I IFN Responses To Facilitate Porcine Reproductive and Respiratory Syndrome Virus Infection by Targeting JAK1. J. Immunol. 2016, 196, 2272–2282. [Google Scholar] [CrossRef] [Green Version]
- Zhao, G.; Hou, J.; Xu, G.; Xiang, A.; Kang, Y.; Yan, Y.; Zhang, X.; Yang, G.; Xiao, S.; Sun, S. Cellular microRNA miR-10a-5p inhibits replication of porcine reproductive and respiratory syndrome virus by targeting the host factor signal recognition particle 14. J. Gen. Virol. 2017, 98, 624–632. [Google Scholar] [CrossRef]
- Liu, F.; Du, Y.; Feng, W.-h. New perspective of host microRNAs in the control of PRRSV infection. Vet. Microbiol. 2017, 209, 48–56. [Google Scholar] [CrossRef] [PubMed]
- An, T.Q.; Tian, Z.J.; Zhou, Y.J.; Xiao, Y.; Peng, J.M.; Chen, J.; Jiang, Y.F.; Hao, X.F.; Tong, G.Z. Comparative genomic analysis of five pairs of virulent parental/attenuated vaccine strains of PRRSV. Vet. Microbiol. 2011, 149, 104–112. [Google Scholar] [CrossRef] [PubMed]
- Chen, N.; Li, X.; Xiao, Y.; Li, S.; Zhu, J. Characterization of four types of MLV-derived porcine reproductive and respiratory syndrome viruses isolated in unvaccinated pigs from 2016 to 2020. Res. Vet. Sci. 2021, 134, 102–111. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhou, L.; Ge, X.; Guo, X.; Han, J.; Yang, H. Evolutionary analysis of six isolates of porcine reproductive and respiratory syndrome virus from a single pig farm: MLV-evolved and recombinant viruses. Infect. Genet. Evol. 2018, 66, 111–119. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Yu, Y.; Zhang, C.; Tu, Y.; Tong, J.; Liu, Y.; Chang, Y.; Jiang, C.; Wang, S.; Zhou, E.M.; et al. Immune responses to modified live virus vaccines developed from classical or highly pathogenic PRRSV following challenge with a highly pathogenic PRRSV strain. Dev. Comp. Immunol. 2016, 62, 1–7. [Google Scholar] [CrossRef]
- Ouyang, K.; Hiremath, J.; Binjawadagi, B.; Shyu, D.-L.; Dhakal, S.; Arcos, J.; Schleappi, R.; Holman, L.; Roof, M.; Torrelles, J.B.; et al. Comparative analysis of routes of immunization of a live porcine reproductive and respiratory syndrome virus (PRRSV) vaccine in a heterologous virus challenge study. Vet. Res. 2016, 47, 11. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.; Oh, T.; Cho, H.; Chae, C. A comparison of commercial modified-live PRRSV-1 and PRRSV-2 vaccines against a dual heterologous PRRSV-1 and PRRSV-2 challenge in late term pregnancy gilts. Comp. Immunol. Microbiol. Infect. Dis. 2020, 69, 101423. [Google Scholar] [CrossRef]
- Oh, T.; Kim, H.; Park, K.H.; Yang, S.; Jeong, J.; Kim, S.; Kang, I.; Park, S.J.; Chae, C. Comparison of 4 commercial modified-live porcine reproductive and respiratory syndrome virus (PRRSV) vaccines against heterologous Korean PRRSV-1 and PRRSV-2 challenge. Can. J. Vet. Res. 2019, 83, 57–67. [Google Scholar]
- Su, F.; Xu, L.; Xue, Y.; Li, J.; Fu, Y.; Yu, B.; Wang, S.; Yuan, X. Th1-biased immunoadjuvant effect of the recombinant B subunit of an Escherichia coli heat-labile enterotoxin on an inactivated porcine reproductive and respiratory syndrome virus antigen via intranasal immunization in mice. J. Vet. Med. Sci. 2019, 81, 1475–1484. [Google Scholar] [CrossRef]
- Renukaradhya, G.; Binjawadagi, B.; Dwivedi, V.; Manickam, C.; Ouyang, K.; Wu, Y.; Lee, J.; Torrelles, J. Adjuvanted poly(lactic-co-glycolic) acid nanoparticle-entrapped inactivated porcine reproductive and respiratory syndrome virus vaccine elicits cross-protective immune response in pigs. Int. J. Nanomed. 2014, 9, 679–694. [Google Scholar] [CrossRef] [Green Version]
- Dhakal, S.; Renukaradhya, G.J. Nanoparticle-based vaccine development and evaluation against viral infections in pigs. Vet. Res. 2019, 50, 14. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Wang, Y.; Duan, H.; Zhang, A.; Liang, C.; Gao, J.; Zhang, C.; Huang, B.; Li, Q.; Li, N.; et al. An intracellularly expressed Nsp9-specific nanobody in MARC-145 cells inhibits porcine reproductive and respiratory syndrome virus replication. Vet. Microbiol. 2015, 181, 252–260. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Liang, C.; Duan, H.; Zhang, X.; Wang, X.; Xiao, S.; Zhou, E.M. Intracellularly expressed nanobodies against non-structural protein 4 of porcine reproductive and respiratory syndrome virus inhibit virus replication. Biotechnol. Lett. 2016, 38, 1081–1088. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Xue, F.; Guo, Y.; Ma, M.; Hao, N.; Zhang, X.C.; Lou, Z.; Li, X.; Rao, Z. Crystal structure of porcine reproductive and respiratory syndrome virus leader protease Nsp1alpha. J. Virol. 2009, 83, 10931–10940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, C.; Deng, Z.; Li, X.; Yang, M.; Tian, Z.; Chen, Z.; Wang, G.; Wu, W.; Feng, W.H.; Zhang, G.; et al. Helicase of Type 2 Porcine Reproductive and Respiratory Syndrome Virus Strain HV Reveals a Unique Structure. Viruses 2020, 12, 215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xue, F.; Sun, Y.; Yan, L.; Zhao, C.; Chen, J.; Bartlam, M.; Li, X.; Lou, Z.; Rao, Z. The crystal structure of porcine reproductive and respiratory syndrome virus nonstructural protein Nsp1beta reveals a novel metal-dependent nuclease. J. Virol. 2010, 84, 6461–6471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, Z.; Lehmann, K.C.; Li, X.; Feng, C.; Wang, G.; Zhang, Q.; Qi, X.; Yu, L.; Zhang, X.; Feng, W.; et al. Structural basis for the regulatory function of a complex zinc-binding domain in a replicative arterivirus helicase resembling a nonsense-mediated mRNA decay helicase. Nucleic Acids Res. 2014, 42, 3464–3477. [Google Scholar] [CrossRef]
- Zhang, M.; Li, X.; Deng, Z.; Chen, Z.; Liu, Y.; Gao, Y.; Wu, W.; Chen, Z. Structural Biology of the Arterivirus nsp11 Endoribonucleases. J. Virol. 2017, 91, e01309–e01316. [Google Scholar] [CrossRef] [Green Version]
- Renukaradhya, G.J.; Meng, X.J.; Calvert, J.G.; Roof, M.; Lager, K.M. Inactivated and subunit vaccines against porcine reproductive and respiratory syndrome: Current status and future direction. Vaccine 2015, 33, 3065–3072. [Google Scholar] [CrossRef] [PubMed]
- Vu, H.L.; Kwon, B.; de Lima, M.; Pattnaik, A.K.; Osorio, F.A. Characterization of a serologic marker candidate for development of a live-attenuated DIVA vaccine against porcine reproductive and respiratory syndrome virus. Vaccine 2013, 31, 4330–4337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Lima, M.; Kwon, B.; Ansari, I.H.; Pattnaik, A.K.; Flores, E.F.; Osorio, F.A. Development of a porcine reproductive and respiratory syndrome virus differentiable (DIVA) strain through deletion of specific immunodominant epitopes. Vaccine 2008, 26, 3594–3600. [Google Scholar] [CrossRef]
- Ma, Z.; Yu, Y.; Xiao, Y.; Opriessnig, T.; Wang, R.; Yang, L.; Nan, Y.; Samal, S.K.; Halbur, P.G.; Zhang, Y.J. Sustaining Interferon Induction by a High-Passage Atypical Porcine Reproductive and Respiratory Syndrome Virus Strain. Sci. Rep. 2016, 6, 36312. [Google Scholar] [CrossRef]
- Ma, H.; Jiang, L.; Qiao, S.; Zhi, Y.; Chen, X.X.; Yang, Y.; Huang, X.; Huang, M.; Li, R.; Zhang, G.P. The Crystal Structure of the Fifth Scavenger Receptor Cysteine-Rich Domain of Porcine CD163 Reveals an Important Residue Involved in Porcine Reproductive and Respiratory Syndrome Virus Infection. J. Virol. 2017, 91, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, Z.Y.; Liu, F.; Li, Y.; Wang, H.L.; Zhang, Z.D.; Chen, Z.Z.; Feng, W.H. Aspartic acid at residue 185 modulates the capacity of HP-PRRSV nsp4 to antagonize IFN-I expression. Virology 2020, 546, 79–87. [Google Scholar] [CrossRef]
- Van Kasteren, P.B.; Bailey-Elkin, B.A.; James, T.W.; Ninaber, D.K.; Beugeling, C.; Khajehpour, M.; Snijder, E.J.; Mark, B.L.; Kikkert, M. Deubiquitinase function of arterivirus papain-like protease 2 suppresses the innate immune response in infected host cells. Proc. Natl. Acad. Sci. USA 2013, 110, E838–E847. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blanco, E.; Guerra, B.; de la Torre, B.G.; Defaus, S.; Dekker, A.; Andreu, D.; Sobrino, F. Full protection of swine against foot-and-mouth disease by a bivalent B-cell epitope dendrimer peptide. Antivir. Res. 2016, 129, 74–80. [Google Scholar] [CrossRef] [PubMed]
- Muderspach, L.; Wilczynski, S.; Roman, L.; Bade, L.; Felix, J.; Small, L.A.; Kast, W.M.; Fascio, G.; Marty, V.; Weber, J. A phase I trial of a human papillomavirus (HPV) peptide vaccine for women with high-grade cervical and vulvar intraepithelial neoplasia who are HPV 16 positive. Clin. Cancer Res. 2000, 6, 3406–3416. [Google Scholar] [PubMed]
- Li, S.; Peng, L.; Zhao, W.; Zhong, H.; Zhang, F.; Yan, Z.; Cao, H. Synthetic peptides containing B- and T-cell epitope of dengue virus-2 E domain III provoked B- and T-cell responses. Vaccine 2011, 29, 3695–3702. [Google Scholar] [CrossRef] [PubMed]
- Spearman, P.; Kalams, S.; Elizaga, M.; Metch, B.; Chiu, Y.L.; Allen, M.; Weinhold, K.J.; Ferrari, G.; Parker, S.D.; McElrath, M.J.; et al. Safety and immunogenicity of a CTL multiepitope peptide vaccine for HIV with or without GM-CSF in a phase I trial. Vaccine 2009, 27, 243–249. [Google Scholar] [CrossRef] [Green Version]
- Zhang, N.; Qi, J.; Feng, S.; Gao, F.; Liu, J.; Pan, X.; Chen, R.; Li, Q.; Chen, Z.; Li, X.; et al. Crystal structure of swine major histocompatibility complex class I SLA-1 0401 and identification of 2009 pandemic swine-origin influenza A H1N1 virus cytotoxic T lymphocyte epitope peptides. J. Virol. 2011, 85, 11709–11724. [Google Scholar] [CrossRef] [Green Version]
- Pan, X.; Zhang, N.; Wei, X.; Jiang, Y.; Chen, R.; Li, Q.; Liang, R.; Zhang, L.; Ma, L.; Xia, C. Illumination of PRRSV Cytotoxic T Lymphocyte Epitopes by the Three-Dimensional Structure and Peptidome of Swine Lymphocyte Antigen Class I (SLA-I). Front. Immunol. 2019, 10, 2995. [Google Scholar] [CrossRef] [PubMed]
- Fan, S.; Wu, Y.; Wang, S.; Wang, Z.; Jiang, B.; Liu, Y.; Liang, R.; Zhou, W.; Zhang, N.; Xia, C. Structural and Biochemical Analyses of Swine Major Histocompatibility Complex Class I Complexes and Prediction of the Epitope Map of Important Influenza A Virus Strains. J. Virol. 2016, 90, 6625–6641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
PRRSV replication and infection cycle, including binding, internalization, uncoating, translation, assembling, releasing the virion, and immune response. The cytokine IFN-α/β stimulates host innate and adaptive immune response against PRRSV infection first. In this process, some important viral structural and non-structural proteins interact with host factors.
Figure 1.
PRRSV replication and infection cycle, including binding, internalization, uncoating, translation, assembling, releasing the virion, and immune response. The cytokine IFN-α/β stimulates host innate and adaptive immune response against PRRSV infection first. In this process, some important viral structural and non-structural proteins interact with host factors.

Figure 2.
D pocket structure of pSLA-1*1502 bound to nsp9-TMP9 peptide, and comparison of the peptide conformations of the three pSLA structures resolved. (a) Overall structure of pSLA-1*1502 complexes with one peptide (nsp9–TMP9) (Protein Data Bank (PDB) code: 5YLX) is represented by electrostatic surface (red, negatively charged; white, non-polar; and blue, positively charged). The nsp9–TMP9 is shown as a stick model (C, orange; N, blue; O, red). The D pocket is shown in particular; P3-PRO represents the third proline residue of the nsp9–TMP9 peptide, and Q155, M156, Y159 residues shown as stick models (C, green; N, blue; O, red) in complexes can interact with the P3-PRO by van der Waals forces. (b) The superposition of the pocket structures of pSLA-1*1502 (wheat, PDB code: 5YLX), pSLA-1*0401 (green, PDB code: 3QQ3), and pSLA-3*hs0202 (cyan, PDB code: 5H94) are shown as cartoons. There are slight differences in the conformation of peptides shown as stick models in the three structures.
Figure 2.
D pocket structure of pSLA-1*1502 bound to nsp9-TMP9 peptide, and comparison of the peptide conformations of the three pSLA structures resolved. (a) Overall structure of pSLA-1*1502 complexes with one peptide (nsp9–TMP9) (Protein Data Bank (PDB) code: 5YLX) is represented by electrostatic surface (red, negatively charged; white, non-polar; and blue, positively charged). The nsp9–TMP9 is shown as a stick model (C, orange; N, blue; O, red). The D pocket is shown in particular; P3-PRO represents the third proline residue of the nsp9–TMP9 peptide, and Q155, M156, Y159 residues shown as stick models (C, green; N, blue; O, red) in complexes can interact with the P3-PRO by van der Waals forces. (b) The superposition of the pocket structures of pSLA-1*1502 (wheat, PDB code: 5YLX), pSLA-1*0401 (green, PDB code: 3QQ3), and pSLA-3*hs0202 (cyan, PDB code: 5H94) are shown as cartoons. There are slight differences in the conformation of peptides shown as stick models in the three structures.


{kind=link}
{kind=link}
Table 1.
List of porcine reproductive and respiratory syndrome virus (PRRSV)–host interactions and their functions. Protein data bank (PDB) codes of structures are enclosed in the brackets.
Table 1.
List of porcine reproductive and respiratory syndrome virus (PRRSV)–host interactions and their functions. Protein data bank (PDB) codes of structures are enclosed in the brackets.
| Host | PRRSV | Function | Virus Genotypes | References |
|---|---|---|---|---|
| Heparan | M/GP5 | Concentrate virions on the cell surface | PRRSV-1/PRRSV-2 | [15] |
| pSn | M/GP5 | PRRSV attachment and internalization receptor via clathrin-mediated endocytosis | PRRSV-1 | [16] |
| CD163 (5JFB) | GP2/GP3/ GP4/GP5 | Uncoating and genome release | PRRSV-1/PRRSV-2 | [13,17] |
| CD151 | 3′ UTR RNA | Cooperate in infection | PRRSV-2 | [18] |
| Simian vimentin | N (1P65) | Mediate transportation of the virus in the cytosol | PRRSV-2 | [19] |
| CD209 | GP5 | Essential in PRRSV entry and infection | PRRSV-1/PRRSV-2 | [20] |
| MYH9 | GP5 | Essential in PRRSV entry and infection | PRRSV-1/PRRSV-2 | [21,22] |
| IFN-β | nsp1α (3IFU) | Suppress IFN by degrading CBP | PRRSV-2 | [23,24] |
| nsp1β (3MTV) | Suppress IFN by inhibiting both IRF-3 and NF-κB-dependent gene induction | PRRSV-2 | [25] | |
| N | Suppress IFN by inhibiting the phosphorylation and nuclear translocation of IRF3 | PRRSV-2 | [26] | |
| nsp2 | Suppress IFN by inhibiting the activation of the IRF-3 and NF-κB signaling | PRRSV-2 | [27] | |
| nsp4 (5Y4L) | Suppress IFN by blocking NF-κB activation | PRRSV-2 | [28] | |
| nsp11 (5EYI) | Suppress IFN by inhibiting the activation of the IRF-3 and NF-κB signaling, and inhibiting the formation and nuclear translocation of ISGF3 targeting IRF9 | PRRSV-2 | [29,30,31] | |
| IL-1β | E | Increase the release of IL-1β | PRRSV-2 | [32] |
| nsp11 | Inhibit the secretion of IL-1β | PRRSV-2 | [33] | |
| IL-10 | N | Up-regulate IL-10 via NF-κB and p38 MAPK pathways in PAMs | PRRSV-2 | [34] |
| nsp1 | Up-regulate IL-10 | PRRSV-2 | [35] | |
| Gp5 | Up-regulate IL-10 through p38 MAPK and signal transducer and activator of transcription-3 (STAT3) activation | PRRSV-2 | [36] | |
| IL-17 | nsp11 | Induced IL-17 production depending on the PI3K-p38MAPK-C/EBPβ/CREB pathways | PRRSV-2 | [37] |
| TRIM25 | N | Competitively interact with TRIM25, thereby interfering with TRIM25-mediated RIG-I ubiquitination | PRRSV-2 | [38] |
| TRIM22 | N | Interact with TRIM22 thereby reducing virus replication | PRRSV-2 | [39] |
| TRIM59 | nsp11 | Interact with TRIM59 thereby reducing virus replication | PRRSV-2 | [40] |
| MiR-181 | ORF4 | Inhibit PRRSV replication | PRRSV-2 | [41,42] |
| MiR-130 | 5′ UTR | PRRSV-2 | [43] | |
| MiR-23 | ORF3 | PRRSV-2 | [44] | |
| MiR-378 | ORF7 | PRRSV-2 | [44] | |
| MiR-505 | ORF3/ORF5 | PRRSV-2 | [44] | |
| PIAS1 SLA-I(5YLX) | nsp1α | Modulate degradation via SUMO E3 ligase activity | PRRSV-2 PRRSV-2 | [45] [45,46] |
| Nup62 | nsp1β | Inhibit host antiviral protein expression | PRRSV-1/PRRSV-2 | [47] |
| STAT3 | nsp5 | Promote the degradation of STAT3 and interference with the JAK/STAT3 signaling | PRRSV-1/PRRSV-2 | [48] |
| pRb | nsp9 | Benefit the replication of PRRSV | PRRSV-2 | [49] |
| NLRX1 | nsp9 | Restrict PRRSV replication | PRRSV-2 | [50] |
| ZAP | nsp9 | Repress PRRSV replication. | PRRSV-2 | [51] |
| Fibrillarin Nucleolin Poly(A)-binding | N | Function remains to be further clarified | PRRSV-2 | [52] |
PRRSV: porcine reproductive and respiratory syndrome virus; UTR: untranslated region; ORF: open reading frame; N: nucleocapsid; IFN: interferon; CBP: cAMP response element-binding (CREB)-binding protein; NF-κB: nuclear factor-κB; IRF: IFN regulatory factor; ISGF3: IFN-stimulated gene factor 3; IL: interleukin; p38 MAPK: p38 mitogen-activated protein kinase; STAT3: signal transducer and activator of transcription-3; TRIM: tripartite motif; RIG-I: retinoic acid–inducible gene I; SUMO: small ubiquitin-related modifier; JAK: Janus kinase; PAMs: porcine alveolar macrophages; pSn: porcine sialoadhesin; MYH9: non-muscle myosin heavy chain 9; MiR: microRNA; PIAS1: protein inhibitor of activated STAT1; SLA-I: swine leukocyte antigen class I; Nup62: nucleoporin 62; pRb: retinoblastoma protein; NLRX1: nucleotide-binding oligomerization domain-like receptor X1; ZAP: zinc finger antiviral protein.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Ma, J.; Ma, L.; Yang, M.; Wu, W.; Feng, W.; Chen, Z. The Function of the PRRSV–Host Interactions and Their Effects on Viral Replication and Propagation in Antiviral Strategies. Vaccines 2021, 9, 364. https://doi.org/10.3390/vaccines9040364
AMA Style
Ma J, Ma L, Yang M, Wu W, Feng W, Chen Z. The Function of the PRRSV–Host Interactions and Their Effects on Viral Replication and Propagation in Antiviral Strategies. Vaccines. 2021; 9(4):364. https://doi.org/10.3390/vaccines9040364
Chicago/Turabian StyleMa, Jun, Lulu Ma, Meiting Yang, Wei Wu, Wenhai Feng, and Zhongzhou Chen. 2021. "The Function of the PRRSV–Host Interactions and Their Effects on Viral Replication and Propagation in Antiviral Strategies" Vaccines 9, no. 4: 364. https://doi.org/10.3390/vaccines9040364
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.