Recombinant Costimulatory Fusion Proteins as Functional Immunomodulators Enhance Antitumor Activity in Murine B16F10 Melanoma

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Lines and Animals
2.2. Construction of Expression Vectors Construction
2.3. Protein Expression and Purification
2.4. Costimulatory Fusion Proteins Treatments
2.5. Ex vivo Stimulation
2.6. Preventive and Therapeutic Tumor Models
2.7. Enzyme-Linked Immunosorbent Assay (ELISA) for Antibody Titers, Interferon-γ (IFN-γ), Interleukin-6 (IL-6) and Interleukin-8 (IL-8) Secretion
2.8. Treg/Teff Ratio and Cytokines Expression in TME
2.9. Serum Transfer Models
2.10. Quantification of the Liver Injury
2.11. Histology and Immunohistochemistry
2.12. Statistical Analysis
3. Results
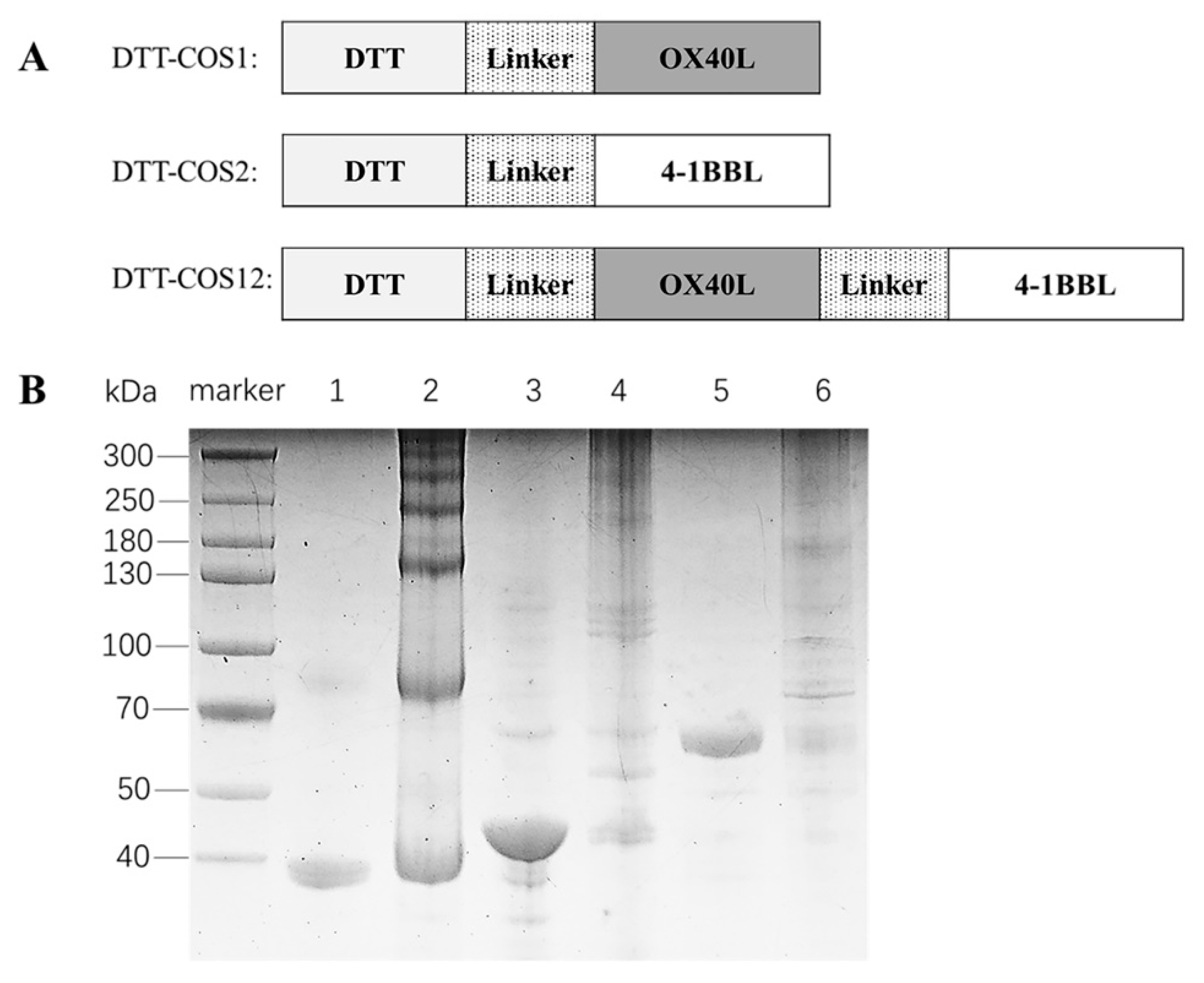
3.1. Design and Biological Activities of Costimulatory Fusion Proteins
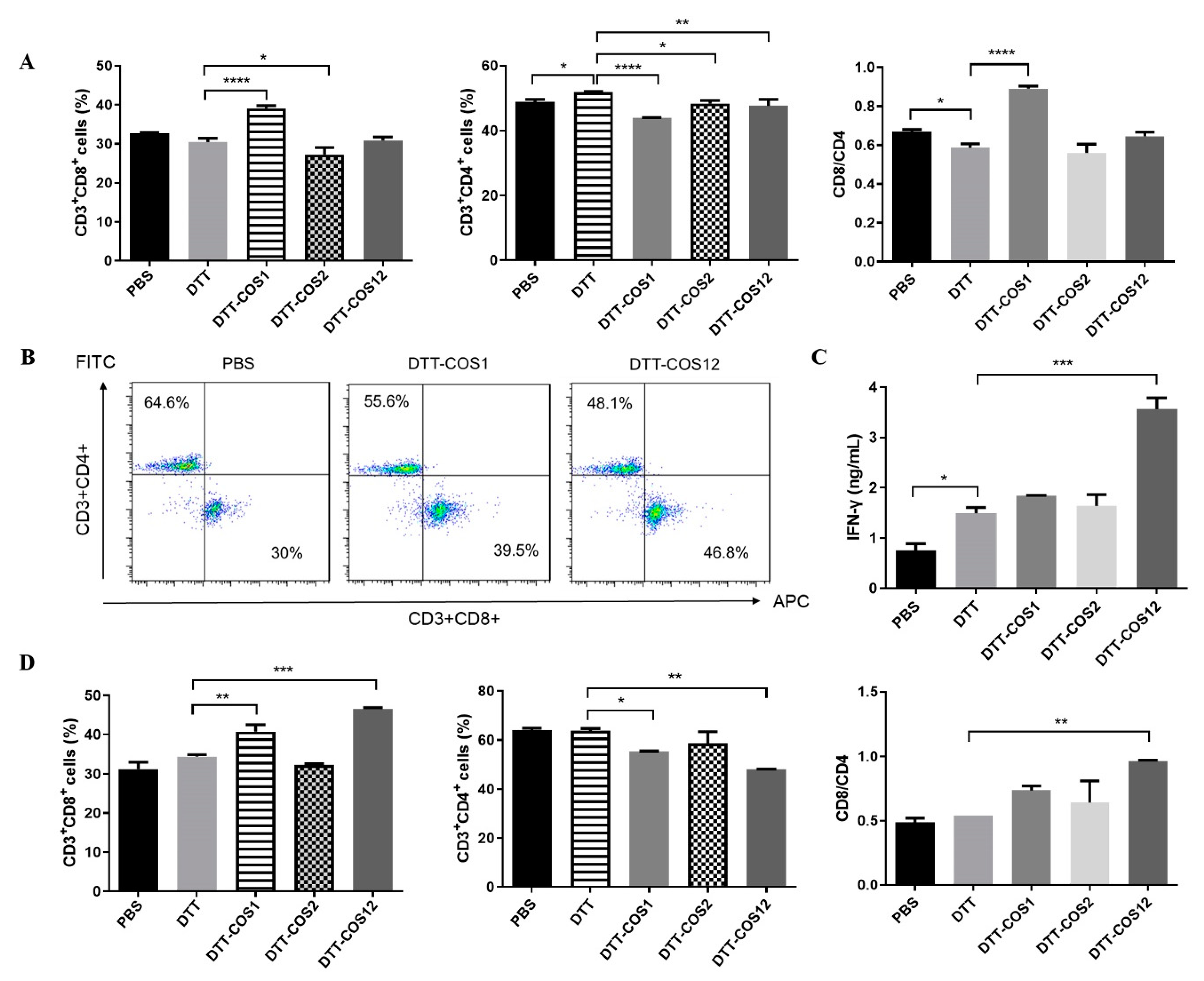
3.2. DTT-COS1 and DTT-COS12 Protect Mice against Tumor Challenge in the Prophylactic Model and Therapeutic Tumor Models In Vivo
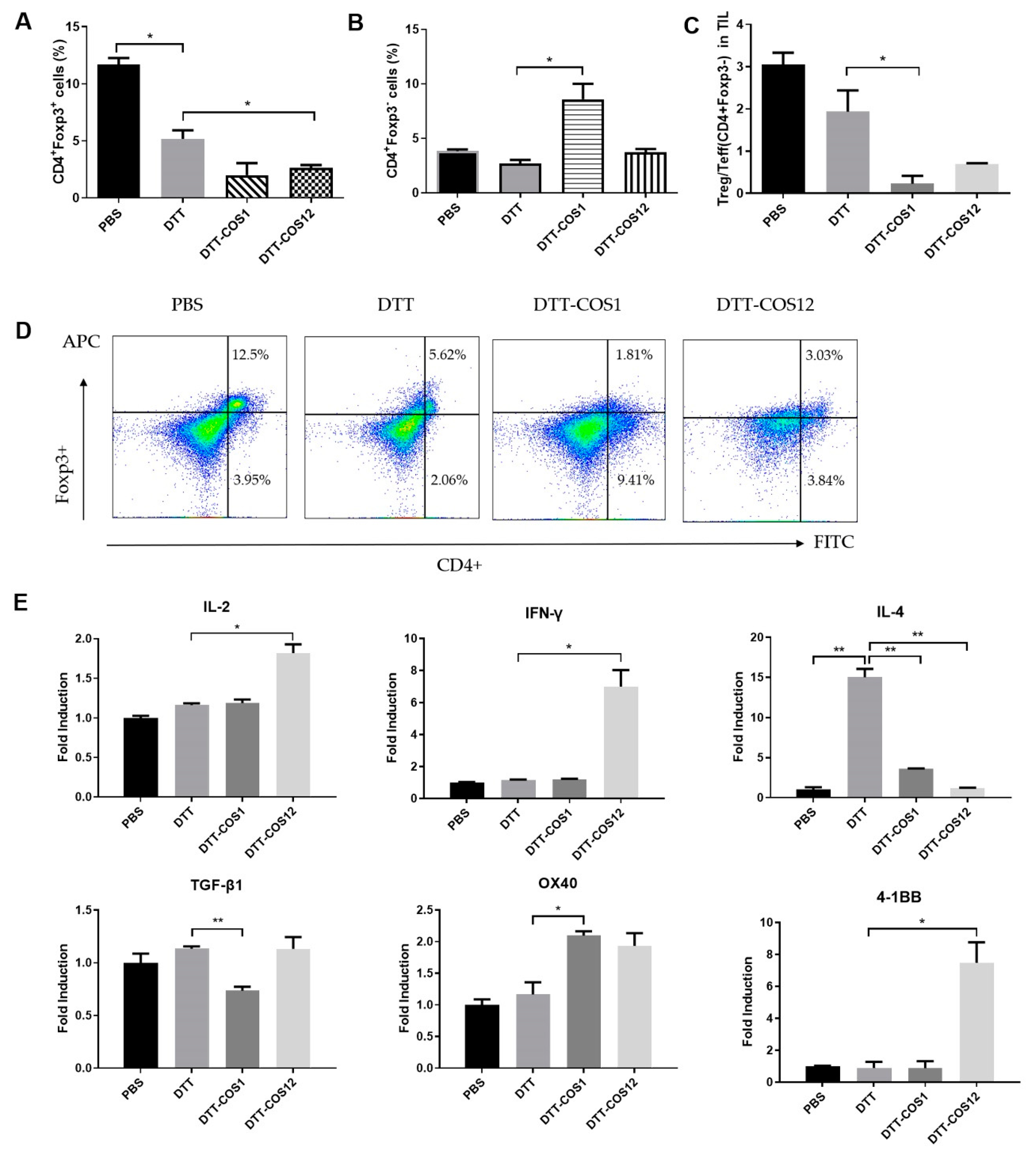
3.3. DTT-COS1 and DTT-COS12 Modulate the Proportion of Treg/Teff Cells and Expression of Cytokines in TME Following Therapeutic Models
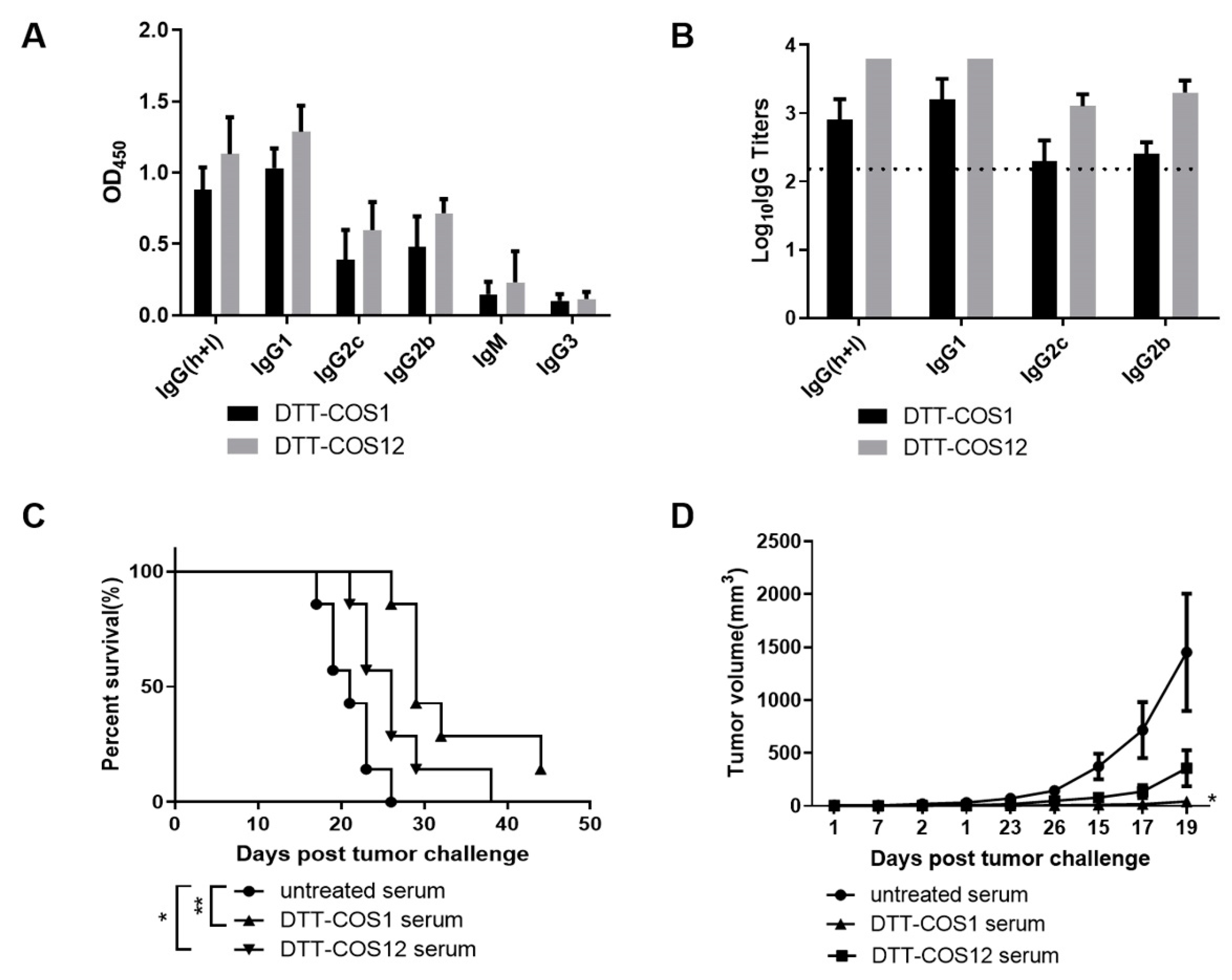
3.4. DTT-COS1 and DTT-COS12 Can Generate Endogenous Antibodies Which May Contribute to Protective Effect against Tumors
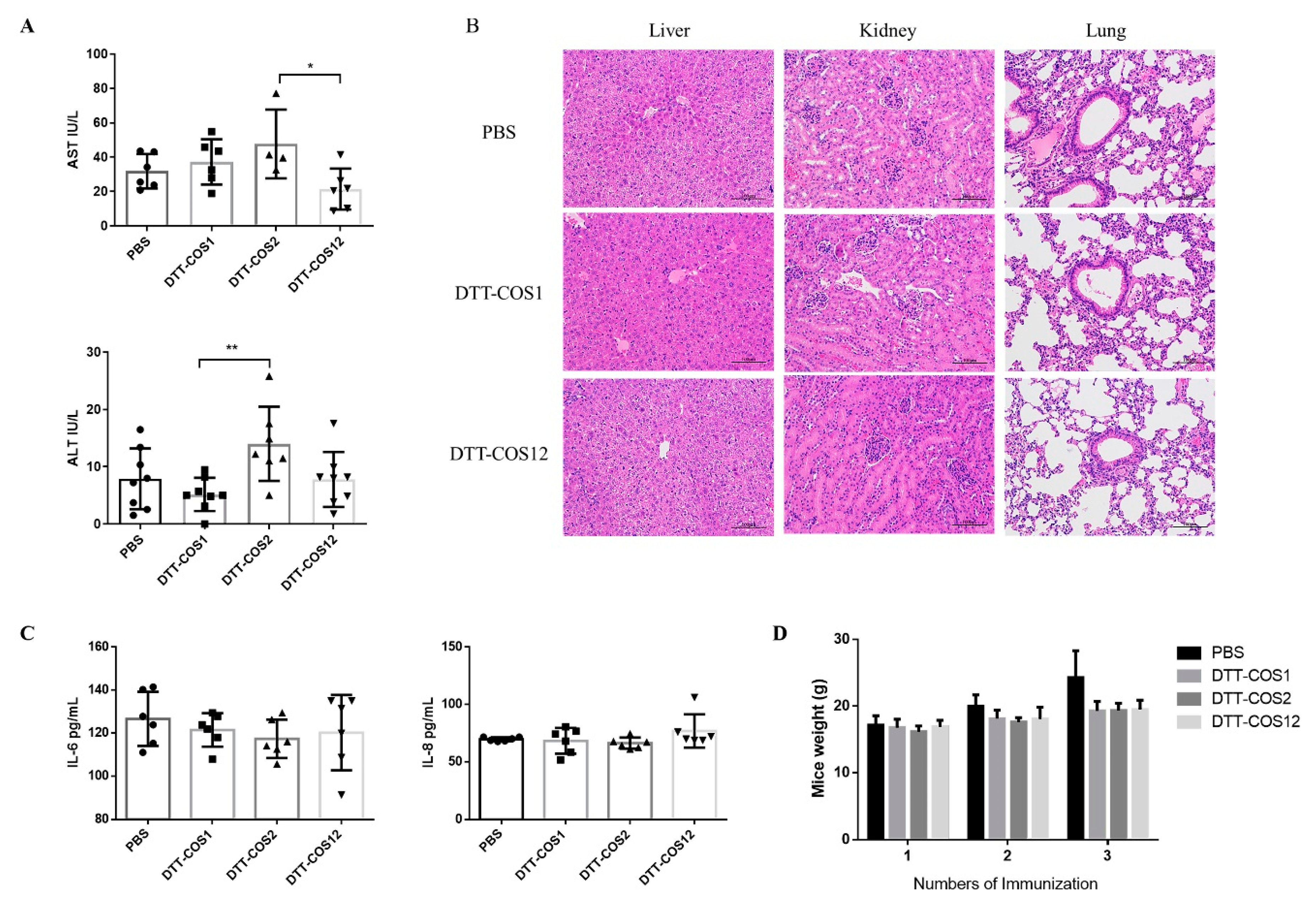
3.5. DTT-COS1 and DTT-COS12 as Safe Formulations in Mice
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Chen, L.; Flies, D.B. Molecular mechanisms of T cell co-stimulation and co-inhibition. Nat. Rev. Immunol. 2013, 13, 227–242. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.; Hu-Lieskovan, S.; Wargo, J.A.; Ribas, A. Primary, Adaptive, and Acquired Resistance to Cancer Immunotherapy. Cell 2017, 168, 707–723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahoney, K.M.; Rennert, P.D.; Freeman, G.J. Combination cancer immunotherapy and new immunomodulatory targets. Nat. Rev. Drug Discov. 2015, 14, 561–584. [Google Scholar] [CrossRef] [PubMed]
- Gough, M.J.; Crittenden, M.R.; Sarff, M.; Pang, P.; Seung, S.K.; Vetto, J.T.; Hu, H.M.; Redmond, W.L.; Holland, J.; Weinberg, A.D. Adjuvant therapy with agonistic antibodies to CD134 (OX40) increases local control after surgical or radiation therapy of cancer in mice. J. Immunother. 2010, 33, 798–809. [Google Scholar] [CrossRef] [Green Version]
- Pan, P.Y.; Zang, Y.; Weber, K.; Meseck, M.L.; Chen, S.H. OX40 ligation enhances primary and memory cytotoxic T lymphocyte responses in an immunotherapy for hepatic colon metastases. Mol. Ther. 2002, 6, 528–536. [Google Scholar] [CrossRef]
- Andrew, D.; Weinberg, K.W.W.; Castle, F.; Ruth, H.W. Blocking OX-40/OX-40 Ligand Interaction In Vitro and In Vivo Leads to Decreased T Cell Function and Amelioration of Experimental Allergic Encephalomyelitis. J. Immunol. 1999, 162, 1818–1826. [Google Scholar]
- hen, A.I.; McAdam, A.J.; Buhlmann, J.E.; Scott, S.; Lupher, M.L., Jr.; Greenfield, E.A.; Baum, P.R.; Fanslow, W.C.; Calderhead, D.M.; Freeman, G.J. Ox40-Ligand Has a Critical Costimulatory Role in Dendritic Cell:T Cell Interactions. Immunity 1999, 11, 689–698. [Google Scholar]
- Myers, L.M.; Vella, A.T. Interfacing T-cell effector and regulatory function through CD137 (4-1BB) co-stimulation. Trends Immunol. 2005, 26, 440–446. [Google Scholar] [CrossRef]
- Frigault, M.J.; Lee, J.; Basil, M.C.; Carpenito, C.; Motohashi, S.; Scholler, J.; Kawalekar, O.U.; Guedan, S.; McGettigan, S.E.; Posey, A.D., Jr.; et al. Identification of chimeric antigen receptors that mediate constitutive or inducible proliferation of T cells. Cancer Immunol. Res. 2015, 3, 356–367. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Lee, L.F.; Fisher, T.S.; Jessen, B.; Elliott, M.; Evering, W.; Logronio, K.; Tu, G.H.; Tsaparikos, K.; Li, X.; et al. Combination of 4-1BB agonist and PD-1 antagonist promotes antitumor effector/memory CD8 T cells in a poorly immunogenic tumor model. Cancer Immunol. Res. 2015, 3, 149–160. [Google Scholar] [CrossRef] [Green Version]
- Bartkowiak, T.; Curran, M.A. 4-1BB Agonists: Multi-Potent Potentiators of Tumor Immunity. Front. Oncol. 2015, 5, 117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nam, K.O.; Kang, H.; Shin, S.M.; Cho, K.H.; Kwon, B.; Kwon, B.S.; Kim, S.J.; Lee, H.W. Cross-Linking of 4-1BB Activates TCR-Signaling Pathways in CD8+ T Lymphocytes. J. Immunol. 2005, 174, 1898–1905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chu, N.R.D.M.; Stiernholm, B.J.; Barber, B.H.; Watts, T.H. Role of IL-12 and 4-1BB ligand in cytokine production by CD28+ and CD28- T cells. J. Immunol. 1997, 158, 3081–3089. [Google Scholar] [PubMed]
- Ignacio Melero, N.B.; Karl Erik, H.; Alejandro, A.; Robert, S.; Mittler Chen, A.L. Amplification of tumor immunity by gene transfer of the co-stimulatory 4-1BB ligand: Synergy with the CD28 co-stimulatory pathway. Eur. J. Immunol. 1998, 28, 1116–1121. [Google Scholar] [CrossRef]
- Barbara-ann Guinn, M.A.D.; Tania, H.W.; Neil, L. Berinstein. 4-1BBL Cooperates with B7-1 and B7-2 in Converting a B Cell Lymphoma Cell Line into a Long-Lasting Antitumor Vaccine. J. Immunol. 1999, 162, 5003–5010. [Google Scholar]
- Young-June Kim, S.H.K.; Patricia, M.; Byoung, S.K. Human 4-1BB regulates CD28 co-stimulation to promote Th1 cell responses. Eur. J. Immunol. 1998, 28, 881–890. [Google Scholar]
- Segal, N.H.; Logan, T.F.; Hodi, F.S.; McDermott, D.; Melero, I.; Hamid, O.; Schmidt, H.; Robert, C.; Chiarion-Sileni, V.; Ascierto, P.A.; et al. Results from an Integrated Safety Analysis of Urelumab, an Agonist Anti-CD137 Monoclonal Antibody. Clin. Cancer Res. 2017, 23, 1929–1936. [Google Scholar] [CrossRef] [Green Version]
- Yonezawa, A.; Dutt, S.; Chester, C.; Kim, J.; Kohrt, H.E. Boosting Cancer Immunotherapy with Anti-CD137 Antibody Therapy. Clin. Cancer Res. 2015, 21, 3113–3120. [Google Scholar] [CrossRef] [Green Version]
- Srivastava, A.K.; Dinc, G.; Sharma, R.K.; Yolcu, E.S.; Zhao, H.; Shirwan, H. SA-4-1BBL and monophosphoryl lipid A constitute an efficacious combination adjuvant for cancer vaccines. Cancer Res. 2014, 74, 6441–6451. [Google Scholar] [CrossRef] [Green Version]
- Malamas, A.S.; Hammond, S.A.; Schlom, J.; Hodge, J.W. Combination therapy with an OX40L fusion protein and a vaccine targeting the transcription factor twist inhibits metastasis in a murine model of breast cancer. Oncotarget 2017, 8, 90825–90841. [Google Scholar] [CrossRef] [Green Version]
- Murphy, K.A.; Lechner, M.G.; Popescu, F.E.; Bedi, J.; Decker, S.A.; Hu, P.; Erickson, J.R.; O’Sullivan, M.G.; Swier, L.; Salazar, A.M.; et al. An in vivo immunotherapy screen of costimulatory molecules identifies Fc-OX40L as a potent reagent for the treatment of established murine gliomas. Clin. Cancer Res. 2012, 18, 4657–4668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wuguang, L.; Lingchong, Q.; Zhanpeng, Y.; Zhibing, L.; Meng, C. Cytotoxic T cell responses are enhanced by antigen design involving the presentation of MUC1 peptide on cholera toxin B subunit. Oncotarget 2015, 6, 34537. [Google Scholar]
- Elpek, K.G.; Yolcu, E.S.; Franke, D.D.H.; Lacelle, C.; Schabowsky, R.H.; Shirwan, H. Ex Vivo Expansion of CD4+CD25+FoxP3+ T Regulatory Cells Based on Synergy between IL-2 and 4-1BB Signaling. J. Immunol. 2007, 179, 7295–7304. [Google Scholar] [CrossRef] [Green Version]
- Xu, A.; Zhang, L.; Chen, Y.; Lin, Z.; Li, R. Immunogenicity and efficacy of a rationally designed vaccine against vascular endothelial growth factor in mouse solid tumor models. Cancer Immunol. Immunother. 2017, 66, 181–192. [Google Scholar] [CrossRef] [PubMed]
- Moynihan, K.D.; Opel, C.F.; Szeto, G.L.; Tzeng, A.; Zhu, E.F.; Engreitz, J.M.; Williams, R.T.; Rakhra, K.; Zhang, M.H.; Rothschilds, A.M.; et al. Eradication of large established tumors in mice by combination immunotherapy that engages innate and adaptive immune responses. Nat. Med. 2016, 22, 1402–1410. [Google Scholar] [CrossRef]
- McNeela, E.A.; O’Connor, D.; Jabbal-Gill, I.; Illum, L.; Davis, S.S.; Pizza, M.; Peppoloni, S.; Rappuoli, R.; Mills, K.H.G. A mucosal vaccine against diphtheria: Formulation of cross reacting material (CRM197) of diphtheria toxin with chitosan enhances local and systemic antibody and Th2 responses following nasal delivery. Vaccine 2000, 19, 1188–1198. [Google Scholar] [CrossRef]
- Schrama, D.; Reisfeld, R.A.; Becker, J.C. Antibody targeted drugs as cancer therapeutics. Nat. Rev. Drug Discov. 2006, 5, 147–159. [Google Scholar] [CrossRef]
- Dekkers, G.; Bentlage, A.E.H.; Stegmann, T.C.; Howie, H.L.; Lissenberg-Thunnissen, S.; Zimring, J.; Rispens, T.; Vidarsson, G. Affinity of human IgG subclasses to mouse Fc gamma receptors. MAbs 2017, 9, 767–773. [Google Scholar] [CrossRef]
- Nishimoto, T.; Okazaki, Y.; Numajiri, M.; Kuwana, M. Mouse immune thrombocytopenia is associated with Th1 bias and expression of activating Fcgamma receptors. Int. J. Hematol. 2017, 105, 598–605. [Google Scholar] [CrossRef]
- Kao, D.; Lux, A.; Schaffert, A.; Lang, R.; Altmann, F.; Nimmerjahn, F. IgG subclass and vaccination stimulus determine changes in antigen specific antibody glycosylation in mice. Eur. J. Immunol. 2017, 47, 2070–2079. [Google Scholar] [CrossRef] [Green Version]
- Pomwised, R.; Intamaso, U.; Teintze, M.; Young, M.; Pincus, S.H. Coupling Peptide Antigens to Virus-Like Particles or to Protein Carriers Influences the Th1/Th2 Polarity of the Resulting Immune Response. Vaccines 2016, 4, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ozer, J.; Ratner, M.; Shaw, M.; Bailey, W.; Schomaker, S. The current state of serum biomarkers of hepatotoxicity. Toxicology 2008, 245, 194–205. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Yao, Y.; Zhang, Q.; Li, S.; Tang, L. Inflammatory responses induced by Helicobacter pylori on the carcinogenesis of gastric epithelial GES-1 cells. Int. J. Oncol. 2019, 54, 2200–2210. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.; He, D.; Ma, C.; Gao, Z.; Chen, Y.; Wang, S. Bifunctional Fusion Proteins Derived from Tumstatin and 4-1BBL for Targeted Cancer Therapy. Mol. Pharm. 2018, 16, 867–876. [Google Scholar] [CrossRef] [PubMed]
- Rabu, C.; Quemener, A.; Jacques, Y.; Echasserieau, K.; Vusio, P.; Lang, F. Production of recombinant human trimeric CD137L (4-1BBL). Cross-linking is essential to its T cell co-stimulation activity. J. Biol. Chem. 2005, 280, 41472–41481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saoulli, K.; Lee, S.Y.; Cannons, J.L.; Yeh, W.C.; Santana, A.; Goldstein, M.D.; Bangia, N.; DeBenedette, M.A.; Mak, T.W.; Choi, Y.; et al. CD28-independent, TRAF2-dependent costimulation of resting T cells by 4-1BB ligand. J. Exp. Med. 1998, 187, 1849–1862. [Google Scholar] [CrossRef]
- Guo, H.; Jiang, W.; Liu, W.; Gao, Y.; Yang, M.; Zhou, Y.; Wang, J.; Qi, J.; Cheng, X.; Zhu, Z.; et al. Extracellular domain of 4-1BBL enhanced the antitumoral efficacy of peripheral blood lymphocytes mediated by anti-CD3 x anti-Pgp bispecific diabody against human multidrug-resistant leukemia. Cell Immunol. 2008, 251, 102–108. [Google Scholar] [CrossRef]
- Morris, N.P.; Peters, C.; Montler, R.; Hu, H.M.; Curti, B.D.; Urba, W.J.; Weinberg, A.D. Development and characterization of recombinant human Fc:OX40L fusion protein linked via a coiled-coil trimerization domain. Mol. Immunol. 2007, 44, 3112–3121. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Wang, J.; Xu, A.; Zhong, C.; Lu, W.; Deng, L.; Li, R. A Rationally Designed TNF-alpha Epitope-Scaffold Immunogen Induces Sustained Antibody Response and Alleviates Collagen-Induced Arthritis in Mice. PLoS ONE 2016, 11, e0163080. [Google Scholar]
- Zhong, C.; Zhang, L.; Chen, L.; Deng, L.; Li, R. Coagulation factor XI vaccination: An alternative strategy to prevent thrombosis. J. Thromb. Haemost. 2017, 15, 122–130. [Google Scholar] [CrossRef] [Green Version]
- Aminian, M.; Sivam, S.; Lee, C.W.; Halperin, S.A.; Lee, S.F. Expression and purification of a trivalent pertussis toxin-diphtheria toxin-tetanus toxin fusion protein in Escherichia coli. Protein Expr. Purif. 2007, 51, 170–178. [Google Scholar] [CrossRef] [PubMed]
- Chenal, A.; Nizard, P.; Forge, V.; Pugniere, M.; Roy, M.O. Does fusion of domains from unrelated proteins affect their folding pathways and the structural changes involved in their function? A case study with the diphtheria toxin T domain. Protein Eng. 2002, 15, 383–391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frankel, A.E.; Rossi, P.; Kuzel, T.M.; Foss, F. Diphtheria Fusion Protein Therapy of Chemoresistant Malignancies. Curr. Cancer Drug Targets 2002, 2, 19–36. [Google Scholar] [CrossRef] [PubMed]
- Frankel, A.E.; Ramage, J.; Melanie, K.; Alexander, R.; Kucera, G.; Miller, M.S. Characterization of diphtheria fusion proteins targeted to the human interleukin-3 receptor. Protein Eng. 2000, 13, 575–581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shuford, W.W.; Klussman, K.; Tritchler, D.D.; Loo, D.T.; Chalupny, J.; Siadak, A.W.; Brown, T.J.; Emswiler, J.; Raecho, H.; Larsen, C.P. 4-1BB Costimulatory Signals Preferentially Induce CD8+ T Cell Proliferation and Lead to the Amplification In Vivo of Cytotoxic T Cell Responses. J. Exp. Med. 1997, 186, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.W.; Park, S.J.; Choi, B.K.; Kim, H.H.; Nam, K.O.; Kwon, B.S. 4-1BB Promotes the Survival of CD8+ T Lymphocytes by Increasing Expression of Bcl-xL and Bfl-1. J. Immunol. 2002, 169, 4882–4888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cannons, J.L.; Lau, P.; Ghumman, B.; Debenedette, M.A.; Yagita, H.; Okumura, K.; Watts, T.H. 4-1BB Ligand Induces Cell Division, Sustains Survival, and Enhances Effector Function of CD4 and CD8 T Cells with Similar Efficacy. J. Immunol. 2001, 167, 1313–1324. [Google Scholar] [CrossRef] [Green Version]
- Bertram, E.M.; Lau, P.; Watts, T.H. Temporal Segregation of 4-1BB Versus CD28-Mediated Costimulation: 4-1BB Ligand Influences T Cell Numbers Late in the Primary Response and Regulates the Size of the T Cell Memory Response Following Influenza Infection. J. Immunol. 2002, 168, 3777–3785. [Google Scholar] [CrossRef] [Green Version]
- Debenedette, M.A.; Wen, T.; Bachmann, M.F.; Ohashi, P.S.; Barber, B.H.; Stocking, K.L.; Peschon, J.J.; Watts, T.H. Analysis of 4-1BB Ligand (4-1BBL)-Deficient Mice and of Mice Lacking Both 4-1BBL and CD28 Reveals a Role for 4-1BBL in Skin Allograft Rejection and in the Cytotoxic T Cell Response to Influenza Virus1. J. Immunol. 1999, 163, 4833–4841. [Google Scholar]
- Sagiv-Barfi, I.; Czerwinski, D.K.; Levy, S.; Alam, I.S.; Mayer, A.T.; Gambhir, S.S.; Levy, R. Eradication of spontaneous malignancy by local immunotherapy. Sci. Transl. Med. 2018, 10, eaan4488. [Google Scholar] [CrossRef] [Green Version]
- Barsoumian, H.B.; Batra, L.; Shrestha, P.; Bowen, W.S.; Zhao, H.; Egilmez, N.K.; Gomez-Gutierrez, J.G.; Yolcu, E.S.; Shirwan, H. A Novel Form of 4-1BBL Prevents Cancer Development via Nonspecific Activation of CD4(+) T and Natural Killer Cells. Cancer Res. 2019, 79, 783–794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belal, C.; Eyad, E. Regulatory T Cells in the Tumor Microenvironment and Cancer Progression: Role and Therapeutic Targeting. Vaccines 2016, 4, 28. [Google Scholar]
- Vu, M.D.; Xiao, X.; Gao, W.; Degauque, N.; Chen, M.; Kroemer, A.; Killeen, N.; Ishii, N.; Li, X.C. OX40 costimulation turns off Foxp3+ Tregs. Blood 2007, 110, 2501–2510. [Google Scholar] [CrossRef] [Green Version]
- Bulliard, Y.; Jolicoeur, R.; Zhang, J.; Dranoff, G.; Wilson, N.S.; Brogdon, J.L. OX40 engagement depletes intratumoral Tregs via activating FcgammaRs, leading to antitumor efficacy. Immunol. Cell Biol. 2014, 92, 475–480. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.L.; Pittet, M.J.; Gorelik, L.; Flavell, R.A.; Weissleder, R.; von Boehmer, H.; Khazaie. K. Regulatory T cells suppress tumor-specific CD8 T cell cytotoxicity through TGF-beta signals in vivo. Proc. Natl. Acad. Sci. USA 2005, 102, 419–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riley, J.L.; June, C.H. The CD28 family: A T-cell rheostat for therapeutic control of T-cell activation. Blood 2005, 105, 13–21. [Google Scholar] [CrossRef] [Green Version]
- Tan, T.T.; Coussens, L.M. Humoral immunity, inflammation and cancer. Curr. Opin. Immunol. 2007, 19, 209–216. [Google Scholar] [CrossRef]
- Schaer, D.A.; Hirschhorn-Cymerman, D.; Wolchok, J.D. Targeting tumor-necrosis factor receptor pathways for tumor immunotherapy. J. Immunother. Cancer 2014, 2, 7. [Google Scholar] [CrossRef] [Green Version]
- Van der Lee, S.; Sanders, E.A.M.; Berbers, G.A.M.; Buisman, A.M. Whole-cell or acellular pertussis vaccination in infancy determines IgG subclass profiles to DTaP booster vaccination. Vaccine 2018, 36, 220–226. [Google Scholar] [CrossRef]
- Zhang, Z.; Goldschmidt, T.; Salter, H. Possible allelic structure of IgG2a and IgG2c in mice. Mol. Immunol. 2012, 50, 169–171. [Google Scholar] [CrossRef]
- Klaus, T.; Bereta, J. CH2 Domain of Mouse IgG3 Governs Antibody Oligomerization, Increases Functional Affinity to Multivalent Antigens and Enhances Hemagglutination. Front. Immunol. 2018, 9, 1096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palma, E.; Cho, M.J. Improved systemic pharmacokinetics, biodistribution, and antitumor activity of CpG oligodeoxynucleotides complexed to endogenous antibodies in vivo. J. Control Release 2007, 120, 95–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cai, H.; Wang, W.; Lin, Z.; Zhang, Y.; Wu, B.; Wan, Y.; Li, R. Recombinant Costimulatory Fusion Proteins as Functional Immunomodulators Enhance Antitumor Activity in Murine B16F10 Melanoma. Vaccines 2020, 8, 223. https://doi.org/10.3390/vaccines8020223
Cai H, Wang W, Lin Z, Zhang Y, Wu B, Wan Y, Li R. Recombinant Costimulatory Fusion Proteins as Functional Immunomodulators Enhance Antitumor Activity in Murine B16F10 Melanoma. Vaccines. 2020; 8(2):223. https://doi.org/10.3390/vaccines8020223
Chicago/Turabian StyleCai, Huaman, Wenfang Wang, Zhibing Lin, Yan Zhang, Bing Wu, Yuhua Wan, and Rongxiu Li. 2020. "Recombinant Costimulatory Fusion Proteins as Functional Immunomodulators Enhance Antitumor Activity in Murine B16F10 Melanoma" Vaccines 8, no. 2: 223. https://doi.org/10.3390/vaccines8020223