
Antibody-Dependent Respiratory Burst against Plasmodium falciparum Merozoites in Individuals Living in an Area with Declining Malaria Transmission
 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area and Population
2.2. Parasite Detection by Microscopy
2.3. Parasite Detection by Real-Time PCR
2.4. Plasmodium falciparum Parasite Preparation
2.5. Neutrophils Preparation
2.6. ADRB Assay
2.7. Data Analysis
2.8. Ethical Statement
3. Results
3.1. Demographic Characteristics
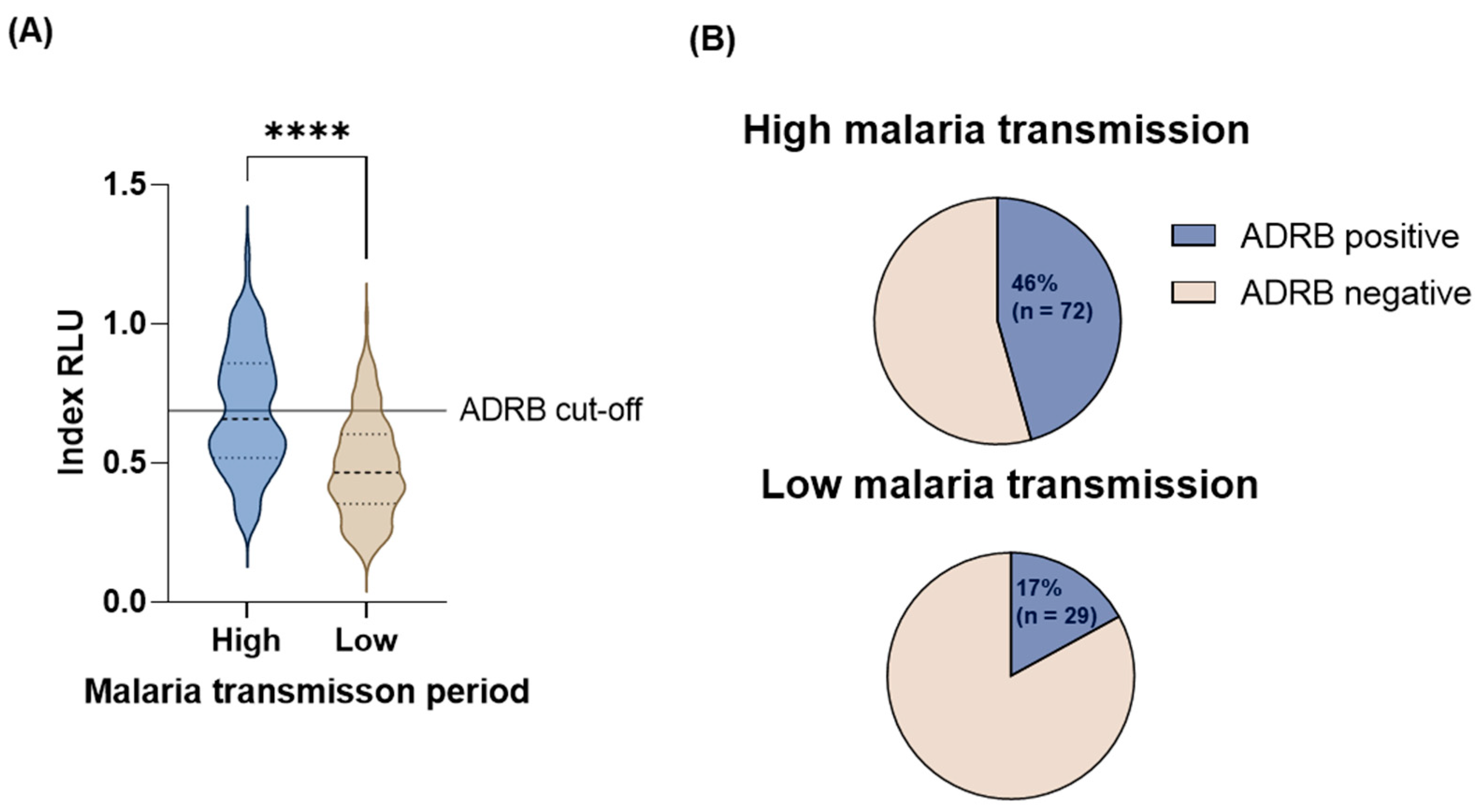
3.2. Malaria Transmission
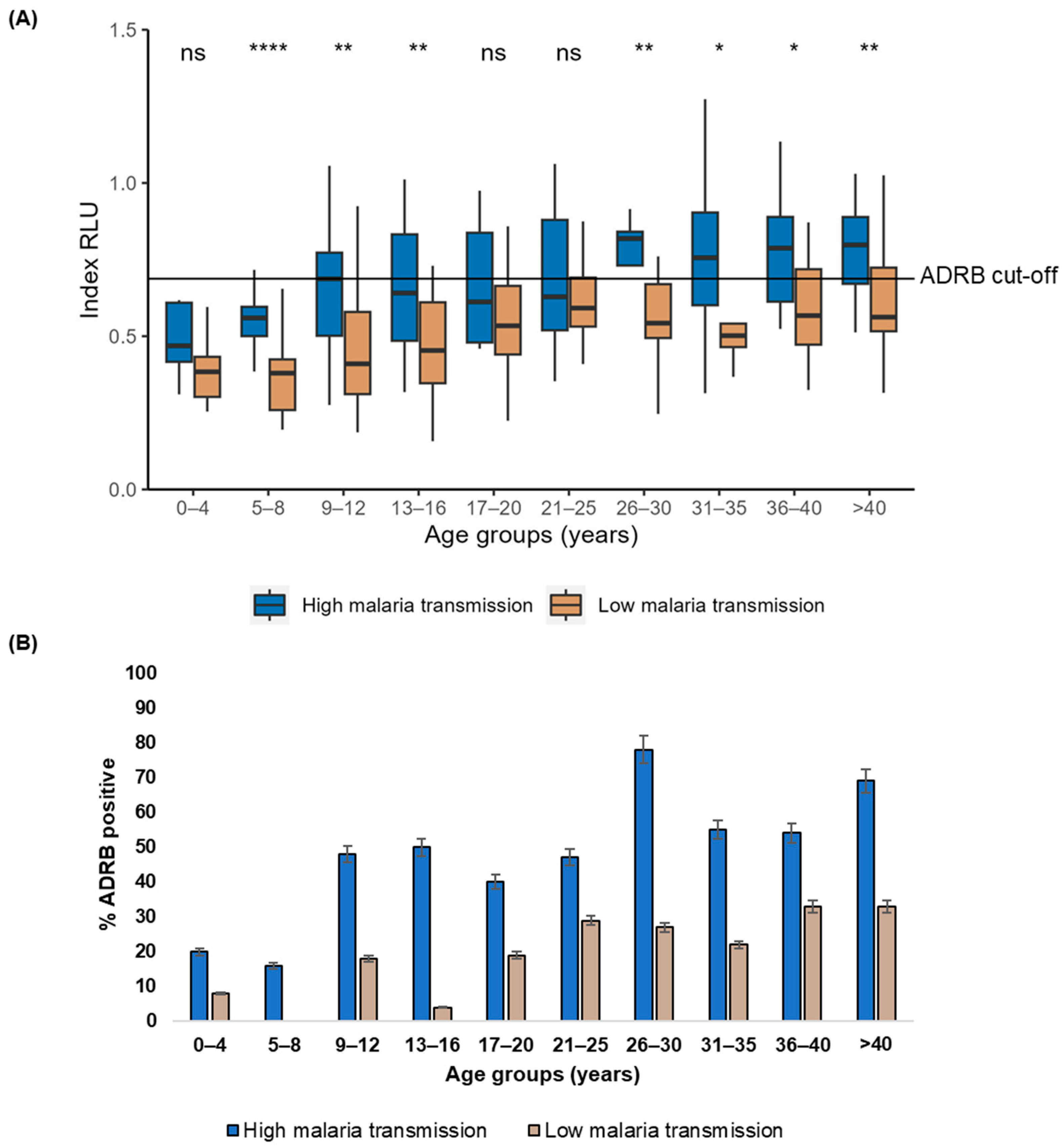
3.3. Age and Malaria Transmission Intensity Influence ADRB Activity
3.4. Age, Parasitemia, and Symptomatic Malaria Are Associated with ADRB Activity
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WHO. World Malaria Report 2023; World Health Organisation: Geneva, Switzerland, 2023. [Google Scholar]
- Seidlein, L.V.; Hanboonkunupakarn, B.; Jittmala, P. RTS, S/AS01, a vaccine targeting pre-erythrocytic stages of Plasmodium falciparum. Emerg. Top Life Sci. 2017, 1, 533–537. [Google Scholar]
- White, M.T.; Bejon, P.; Olotu, A.; Griffin, J.T.; Bojang, K.; Lusingu, J.; Salim, N.; Abdulla, S.; Otsyula, N.; Agnandji, S.T.; et al. A combined analysis of immunogenicity, antibody kinetics and vaccine efficacy from phase 2 trials of the RTS, S malaria vaccine. BMC Med. 2014, 12, 117. [Google Scholar] [CrossRef]
- Collins, K.A.; Snaith, R.; Cottingham, M.G.; Gilbert, S.C.; Hill, A.V.S. Enhancing protective immunity to malaria with a highly immunogenic virus-like particle vaccine. Sci. Rep. 2017, 7, 46621. [Google Scholar] [CrossRef] [PubMed]
- WHO. Recommends R21/Matrix-M Vaccine for Malaria Prevention in Updated Advice on Immunization. Available online: https://www.who.int/news/item/02-10-2023-who-recommends-r21-matrix-m-vaccine-for-malaria-prevention-in-updated-advice-on-immunization (accessed on 5 January 2024).
- Datoo, M.S.; Dicko, A.; Tinto, H.; Ouédraogo, J.; Hamaluba, M.; Olotu, A.; Beaumont, E.; Lopez, F.R.; Natama, H.M.; Weston, S.; et al. Safety and efficacy of malaria vaccine candidate R21/Matrix-M in African children: A multicentre, double-blind, randomised, phase 3 trial. Lancet. 2024, 403, 533–544. [Google Scholar] [CrossRef]
- Quagliata, M.; Papini, A.M.; Rovero, P. Malaria vaccines. Expert Opin. Ther. Pat. 2023, 33, 169–178. [Google Scholar] [CrossRef]
- Cohen, S.; McGregor, I.A.; Carrington, S. Gamma-globulin and acquired immunity to human Malaria. Nature 1961, 192, 733–734. [Google Scholar] [CrossRef]
- Edozien, J.C.; Gilles, H.M.; Udeozo, I.O.K. Adult and cord-blood gamma-globulin and immunity to malaria in Nigerians. Lancet 1962, 280, 951–955. [Google Scholar] [CrossRef]
- Tijani, M.K.; Lugaajju, A.; Persson, K.E.M. Naturally acquired antibodies against Plasmodium falciparum: Friend or foe? Pathogens 2021, 10, 832. [Google Scholar] [CrossRef]
- Duncan, C.J.A.; Hill, A.V.S.; Ellis, R.D. Can growth inhibition assays (GIA) predict blood-stage malaria vaccine efficacy? Hum. Vaccines Immunother. 2012, 8, 706–714. [Google Scholar] [CrossRef] [PubMed]
- Nauseef, W.M. Neutrophils, from cradle to grave and beyond. Immunol. Rev. 2016, 273, 5–10. [Google Scholar] [CrossRef] [PubMed]
- Liew, P.X.; Kubes, P. During Health and Disease. Physiol. Rev. 2019, 99, 1223–1248. [Google Scholar] [CrossRef]
- Greve, B.; Lehman, L.G.; Lell, B.; Luckner, D.; Schmidt-Ott, R.; Kremsner, P.G. High Oxygen Radical Production Is Associated with Fast Parasite Clearance in Children with Plasmodium falciparum Malaria. J. Infect. Dis. 1999, 179, 1584–1586. [Google Scholar] [CrossRef]
- Llewellyn, D.; Miura, K.; Fay, M.P.; Williams, A.R.; Murungi, L.M.; Shi, J.; Hodgson, S.H.; Douglas, A.D.; Osier, F.H.; Fairhurst, R.M.; et al. Standardization of the antibody-dependent respiratory burst assay with human neutrophils and Plasmodium falciparum malaria. Sci. Rep. 2015, 5, 14081. [Google Scholar] [CrossRef] [PubMed]
- Mansourou, A.; Joos, C.; Niass, O.; Diouf, B.; Tall, A.; Perraut, R.; Niang, M.; Toure-Balde, A. Improvement of the antibody-dependent respiratory burst assay for assessing protective immune responses to malaria. Open Biol. 2022, 12, 210288. [Google Scholar] [CrossRef]
- Joos, C.; Marrama, L.; Polson, H.E.; Corre, S.; Diatta, A.M.; Diouf, B.; Trape, J.F.; Tall, A.; Longacre, S.; Perraut, R. Clinical protection from falciparum malaria correlates with neutrophil respiratory bursts induced by merozoites opsonized with human serum antibodies. PLoS ONE 2010, 5, e9871. [Google Scholar] [CrossRef] [PubMed]
- Langhorne, J.; Ndungu, F.M.; Sponaas, A.M.; Marsh, K. Immunity to malaria: More questions than answers. Nat. Immunol. 2008, 9, 725–732. [Google Scholar] [CrossRef]
- Ataíde, R.; Powell, R.; Moore, K.; McLean, A.; Phyo, A.P.; Nair, S.; White, M.; Anderson, T.J.; Beeson, J.G.; Simpson, J.A.; et al. Declining Transmission and Immunity to Malaria and Emerging Artemisinin Resistance in Thailand: A Longitudinal Study. J. Infect. Dis. 2017, 216, 723–731. [Google Scholar] [CrossRef]
- Mugyenyi, C.K.; Elliott, S.R.; Yap, X.Z.; Feng, G.; Boeuf, P.; Fegan, G.; Osier, F.F.; Fowkes, F.J.; Avril, M.; Williams, T.N.; et al. Declining Malaria Transmission Differentially Impacts the Maintenance of Humoral Immunity to Plasmodium falciparum in Children. J. Infect. Dis. 2017, 216, 879–898. [Google Scholar] [CrossRef]
- Raghavan, M.; Kalantar, K.L.; Duarte, E.; Teyssier, N.; Takahashi, S.; Kung, A.F.; Rajan, J.V.; Rek, J.; Tetteh, K.K.; Drakeley, C.; et al. Proteome-wide antigenic profiling in Ugandan cohorts identifies associations between age, exposure intensity, and responses to repeat-containing antigens in Plasmodium falciparum. Elife 2023, 12, e81402. [Google Scholar]
- Kurtovic, L.; Agius, P.A.; Feng, G.; Drew, D.R.; Ubillos, I.; Sacarlal, J.; Aponte, J.J.; Fowkes, F.J.I.; Dobaño, C.; Beeson, J.G. Induction and decay of functional complement-fixing antibodies by the RTS, S malaria vaccine in children, and a negative impact of malaria exposure. BMC Med. 2019, 17, 45. [Google Scholar] [CrossRef]
- Feng, G.; Kurtovic, L.; Agius, P.A.; Aitken, E.H.; Sacarlal, J.; Wines, B.D.; Hogarth, P.M.; Rogerson, S.J.; Fowkes, F.J.I.; Dobaño, C.; et al. Induction, decay, and determinants of functional antibodies following vaccination with the RTS, S malaria vaccine in young children. BMC Med. 2022, 20, 289. [Google Scholar] [CrossRef]
- Dent, A.E.; Chelimo, K.; Sumba, P.O.; Spring, M.D.; Crabb, B.S.; Moormann, A.M.; Tisch, D.J.; Kazura, J.W. Temporal stability of naturally acquired immunity to merozoite surface protein-1 in Kenyan adults. Malar. J. 2009, 8, 162. [Google Scholar] [CrossRef]
- Crompton, P.D.; Kayala, M.A.; Traore, B.; Kayentao, K.; Ongoiba, A.; Weiss, G.E. A prospective analysis of the Ab response to Plasmodium falciparum before and after a malaria season by protein microarray. Proc. Natl. Acad. Sci. USA 2010, 107, 2–7. [Google Scholar] [CrossRef]
- Osier, F.H.; Feng, G.; Boyle, M.J.; Langer, C.; Zhou, J.; Richards, J.S.; McCallum, F.J.; Reiling, L.; Jaworowski, A.; Anders, R.F.; et al. Opsonic phagocytosis of Plasmodium falciparum merozoites: Mechanism in human immunity and a correlate of protection against malaria. BMC Med. 2014, 12, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Färnert, A.; Yman, V.; Vafa Homann, M.; Wandell, G.; Mhoja, L.; Johansson, M.; Jesaja, S.; Sandlund, J.; Tanabe, K.; Hammar, U.; et al. Epidemiology of malaria in a village in the Rufiji River Delta, Tanzania: Declining transmission over 25 years revealed by different parasitological metrics. Malar. J. 2014, 13, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Maqbool, M.; Vidyadaran, S.; George, E.; Ramasamy, R. Optimisation of Laboratory Procedures for Isolating Human Peripheral Blood-Derived Neutrophils. Med. J. Malays. 2011, 66, 296–299. [Google Scholar]
- Posit Team. R Studio: Integrated Development Environment for R; Posit Software, PBC: Boston, MA, USA, 2023. [Google Scholar]
- Doolan, D.L.; Dobaño, C.; Baird, J.K. Acquired immunity to Malaria. Clin. Microbiol. Rev. 2009, 22, 13–36. [Google Scholar] [CrossRef] [PubMed]
- Freya, F. Immunity to malaria in an era of declining malaria transmission. Parasitology 2016, 143, 139–153. [Google Scholar]
- Weiss, G.E.; Traore, B.; Kayentao, K.; Ongoiba, A.; Doumbo, S.; Doumtabe, D.; Kone, Y.; Dia, S.; Guindo, A.; Traore, A.; et al. The Plasmodium falciparum-specific human memory b cell compartment expands gradually with repeated malaria infections. PLoS Pathog. 2010, 6, e1000912. [Google Scholar] [CrossRef]
- Hill, D.L.; Eriksson, E.M.; Li Wai Suen, C.S.; Chiu, C.Y.; Ryg-Cornejo, V.; Robinson, L.J.; Siba, P.M.; Mueller, I.; Hansen, D.S.; Schofield, L. Opsonising Antibodies to P. falciparum Merozoites Associated with Immunity to Clinical Malaria. PLoS ONE 2013, 8, e7462. [Google Scholar] [CrossRef]
- Kurtovic, L.; Behet, M.C.; Feng, G.; Reiling, L.; Chelimo, K.; Dent, A.E.; Mueller, I.; Kazura, J.W.; Sauerwein, R.W.; Fowkes, F.J.I.; et al. Human antibodies activate complement against Plasmodium falciparum sporozoites and are associated with protection against malaria in children. BMC Med. 2018, 16, 61. [Google Scholar] [CrossRef]
- Bhatt, S.; Weiss, D.J.; Cameron, E.; Bisanzio, D.; Mappin, B.; Dalrymple, U.; Battle, K.E.; Moyes, C.L.; Henry, A.; Eckhoff, P.A.; et al. The effect of malaria control on Plasmodium falciparum in Africa between 2000 and 2015. Nature 2015, 526, 207–211. [Google Scholar] [CrossRef] [PubMed]
- White, M.T.; Griffin, J.T.; Akpogheneta, O.; Conway, D.J.; Koram, K.A.; Riley, E.M.; Ghani, A.C. Dynamics of the antibody response to Plasmodium falciparum infection in African children. J. Infect. Dis. 2014, 210, 1115–1122. [Google Scholar] [CrossRef] [PubMed]
- Akpogheneta, O.J.; Dunyo, S.; Pinder, M.; Conway, D.J. Boosting antibody responses to Plasmodium falciparum merozoite antigens in children with highly seasonal exposure to infection. Parasite Immunol. 2010, 32, 296–304. [Google Scholar] [CrossRef] [PubMed]
- Odera, D.O.; Tuju, J.; Mwai, K.; Nkumama, I.N.; Fürle, K.; Chege, T.; Kimathi, R.; Diehl, S.; Musasia, F.K.; Rosenkranz, M.; et al. Anti-merozoite antibodies induce natural killer cell effector function and are associated with immunity against malaria. Sci. Transl. Med. 2023, 15, eabn59. [Google Scholar] [CrossRef]
- Shabihkhani, M.; Lucey, G.M.; Wei, B.; Mareninov, S.; Lou, J.J.; Vinters, H.V.; Singer, E.J.; Cloughesy, T.F.; Yong, W.H. The procurement, storage, and quality assurance of frozen blood and tissue biospecimens in pathology, biorepository, and biobank settings. Clin. Biochem. 2014, 47, 258–266. [Google Scholar] [CrossRef] [PubMed]
- Cherif, M.K.; Ouédraogo, O.; Sanou, G.S.; Diarra, A.; Ouédraogo, A.; Tiono, A.; Cavanagh, D.R.; Michael, T.; Konaté, A.T.; Watson, N.L.; et al. Antibody responses to P. falciparum blood stage antigens and incidence of clinical malaria in children living in an endemic area in Burkina Faso. BMC Res. Notes 2017, 10, 472. [Google Scholar] [CrossRef]
- Dobaño, C.; Santano, R.; Vidal, M.; Jiménez, A.; Jairoce, C.; Ubillos, I.; Dosoo, D.; Aguilar, R.; Williams, N.A.; Díez-Padrisa, N.; et al. Differential patterns of IgG subclass responses to plasmodium falciparum antigens in relation to malaria protection and RTS, S vaccination. Front. Immunol. 2019, 10, 439. [Google Scholar] [CrossRef]
- Hamre, K.E.; Ondigo, B.N.; Hodges, J.S.; Dutta, S.; Theisen, M.; Ayodo, G.; John, C.C. Malaria transmission and immunity in the Kenyan highlands antibody correlates of protection from clinical Plasmodium falciparum malaria in an area of low and unstable malaria transmission. Am. J. Trop. Med. Hyg. 2020, 103, 2174–2182. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| High Malaria Transmission Period | Low Malaria Transmission Period | ||
|---|---|---|---|
| Survey years | 1993, 1994, 1995 | 2016, 2018, 2019 | |
| Individuals, n | 158 | 171 | |
| Age, years | Median (IQR) | 16 (10–31) | 16 (9–29) |
| Age groups, n | 0–4 | 10 | 13 |
| 5–8 | 25 | 27 | |
| 9–12 | 23 | 28 | |
| 13–16 | 22 | 23 | |
| 17–20 | 10 | 16 | |
| 21–25 | 19 | 14 | |
| 26–30 | 9 | 11 | |
| 31–35 | 11 | 9 | |
| 36–40 | 13 | 12 | |
| >40 | 16 | 18 | |
| Sex, n (% [95% CI]) | Female | 88 (55.7 [47.9–63.2]) | 104 (60.8 [53.3–67.8]) |
| Male | 70 (44.3 [0.37–0.52]) | 67 (39.2 [0.32–0.47]) | |
| Fever at sampling, n (% [95% CI]) | 8 (5.1 [2.6–9.7]) | 41 (24 [18.2–30.9]) | |
| Microscopy positive, n (% [95% CI]) | All | 60 (37.9 [30.8–45.7]) | 9 (5.3 [2.8–9.7]) |
| Asymptomatic | 52 (32.9 [26.2–40.6]) | 4 (2.3 [0.9–5.9]) | |
| Symptomatic | 8 (5.1 [2.6–9.7]) | 5 (2.9 [1.3–6.7]) | |
| P. falciparum parasite densities, p/µL (IQR) by microscopy | All | 360 (80–800) | 760 (560–3160) |
| Asymptomatic | 280 (80–560) | 1520 * | |
| Symptomatic | 2760 (1940–7220) | 3160 (560–15,360) | |
| Real-time PCR positive, n (% [95% CI]) | All | 120 (75.9 [68.7–81.9]) | 21 (12.3 [81.7–18.0]) |
| Real-time PCR positive P. falciparum, n (% [95% CI]) | All | 113 (71.5 [64–78]) | 17 (9.9 [6.3–15.3]) |
| Asymptomatic | 106 (67.1 [59.4–73.9]) | 8 (4.7 [2.4–9]) | |
| Symptomatic | 7(4.4 [2.2–8.9]) | 9(5.3 [2.8–9.7]) | |
| Variables | OR | 95% CI | p Value | |
|---|---|---|---|---|
| High malaria transmission period | Age | 1.109 | 1.052–1.186 | <0.001 |
| Sex, Female | 0.927 | 0.339–2.492 | 0.957 | |
| Parasite-positive | 1.033 | 0.315–3.219 | 0.956 | |
| Febrile | 1.059 | 0.112–10.073 | 0.957 | |
| Low malaria transmission period | Age | 1.071 | 1.038–1.110 | <0.001 |
| Sex, Female | 0.694 | 0.258–1.858 | 0.464 | |
| Parasite-positive | 8.273 | 1.851–40.596 | 0.006 | |
| Febrile | 0.676 | 0.191–2.112 | 0.518 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mutemi, D.D.; Tuju, J.; Ogwang, R.; Nyamako, L.; Wambui, K.M.; Cruz, I.R.; Villner, P.; Yman, V.; Kinyanjui, S.M.; Rooth, I.; et al. Antibody-Dependent Respiratory Burst against Plasmodium falciparum Merozoites in Individuals Living in an Area with Declining Malaria Transmission. Vaccines 2024, 12, 203. https://doi.org/10.3390/vaccines12020203
Mutemi DD, Tuju J, Ogwang R, Nyamako L, Wambui KM, Cruz IR, Villner P, Yman V, Kinyanjui SM, Rooth I, et al. Antibody-Dependent Respiratory Burst against Plasmodium falciparum Merozoites in Individuals Living in an Area with Declining Malaria Transmission. Vaccines. 2024; 12(2):203. https://doi.org/10.3390/vaccines12020203
Chicago/Turabian StyleMutemi, Doreen D., James Tuju, Rodney Ogwang, Lydia Nyamako, Kennedy M. Wambui, Ivette R. Cruz, Pär Villner, Victor Yman, Samson M. Kinyanjui, Ingegerd Rooth, and et al. 2024. "Antibody-Dependent Respiratory Burst against Plasmodium falciparum Merozoites in Individuals Living in an Area with Declining Malaria Transmission" Vaccines 12, no. 2: 203. https://doi.org/10.3390/vaccines12020203