
A Polysaccharide-RBD-Fc-Conjugated COVID-19 Vaccine, SCTV01A, Showed High Immunogenicity and Low Toxicity in Animal Models
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Reagents
2.3. Plasmid Construction and Expression of RBD-Fc
2.4. Preparation of Pneumococcal Capsular Polysaccharides
2.5. Conjugation of RBD-Fc to PPS14
2.6. Reduced Polyacrylamide Gel Electrophoresis
2.7. Size Exclusion Chromatography Coupled with Multi Angle Light Scattering (SEC-MALS)
2.8. Transmission Electron Microscopy (TEM)
2.9. Dynamic Light Scattering (DLS)
2.10. Receptor Binding of SCTV01A to Human ACE2 by BLI
2.11. Immunogenicity Analysis of SCTV01A in Mice
2.12. RBD-Specific Antibody ELISA Assay
2.13. SARS-CoV-2 Pseudovirus-Based Neutralization Assay
2.14. Enzyme-Linked Immune Absorbent Spot (ELISpot) Assay
2.15. Opsonophagocytic Assay
2.16. Challenge Assay of Rhesus Macaques
2.17. Micro-Neutralization Assay
2.18. Sample Collection and Viral Nucleic Acid Extraction
2.19. qRT-PCR
2.20. X-ray
2.21. Hematoxylin and Eosin (H&E) Staining
2.22. Safety Evaluation of SCTV01A in Nonhuman Primates
2.23. Statistical Analysis
3. Results
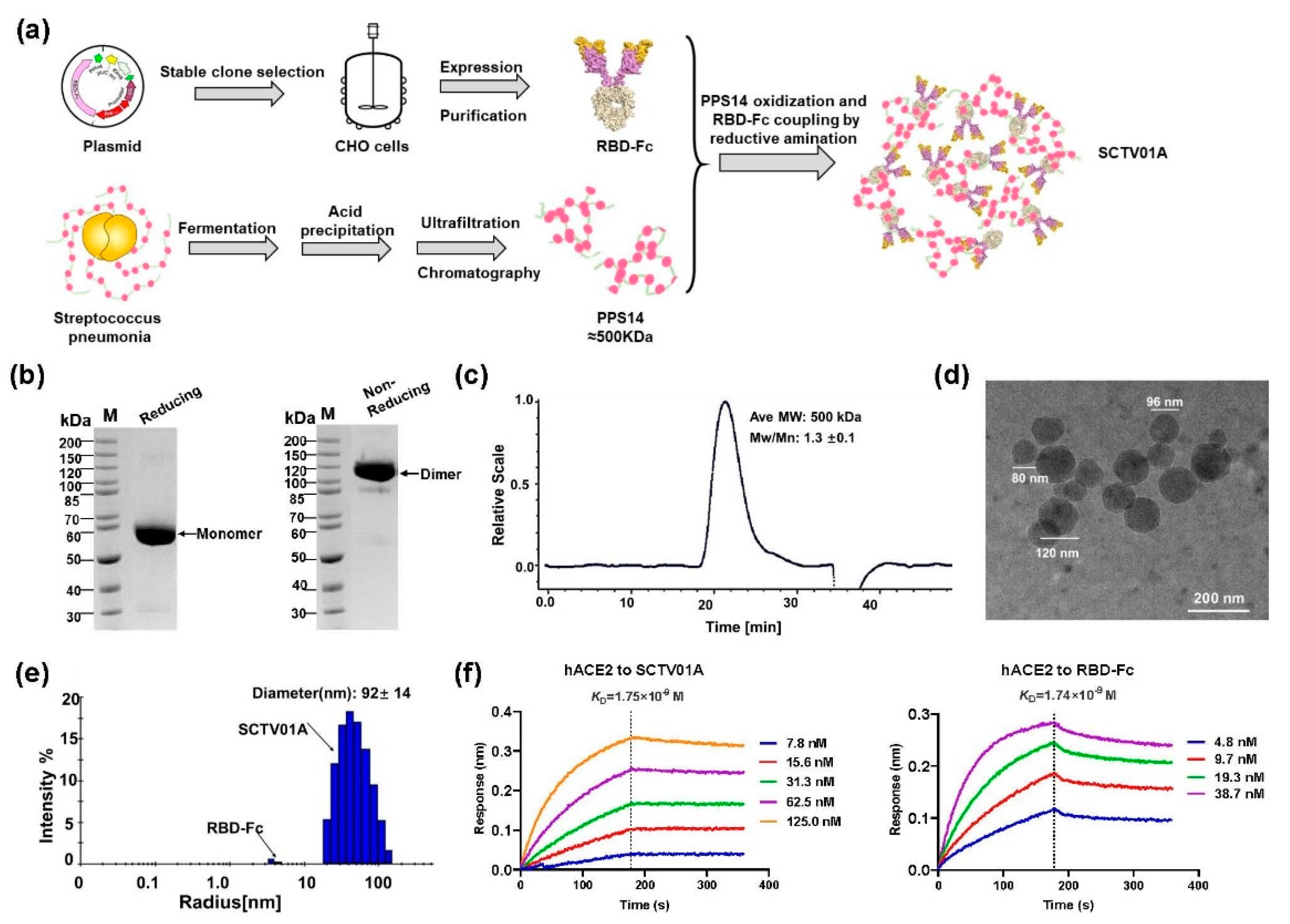
3.1. Design, Production and Characterization of SCTV01A
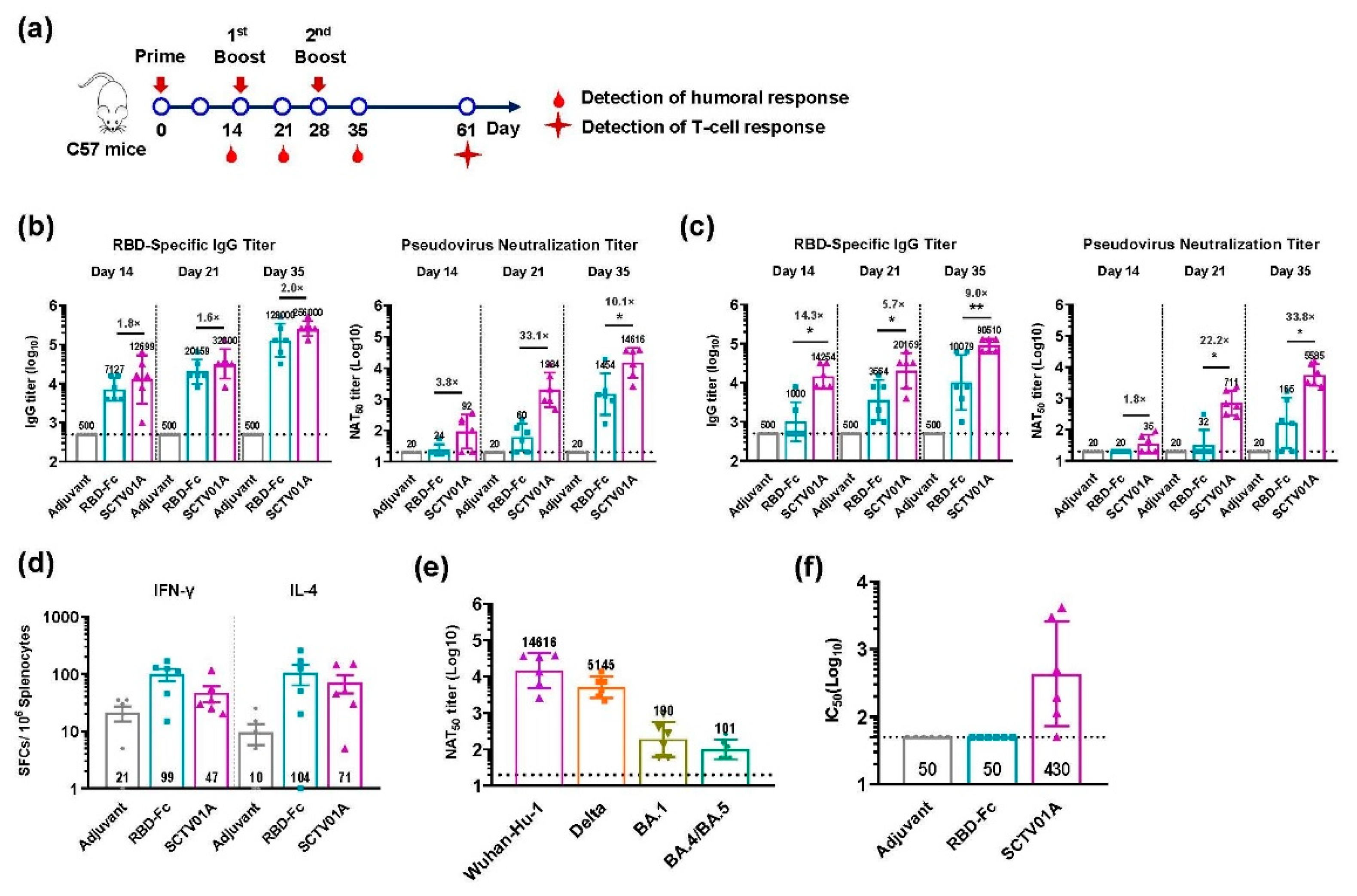
3.2. SCTV01A Augments RBD-Specific Antigenicity in Mice
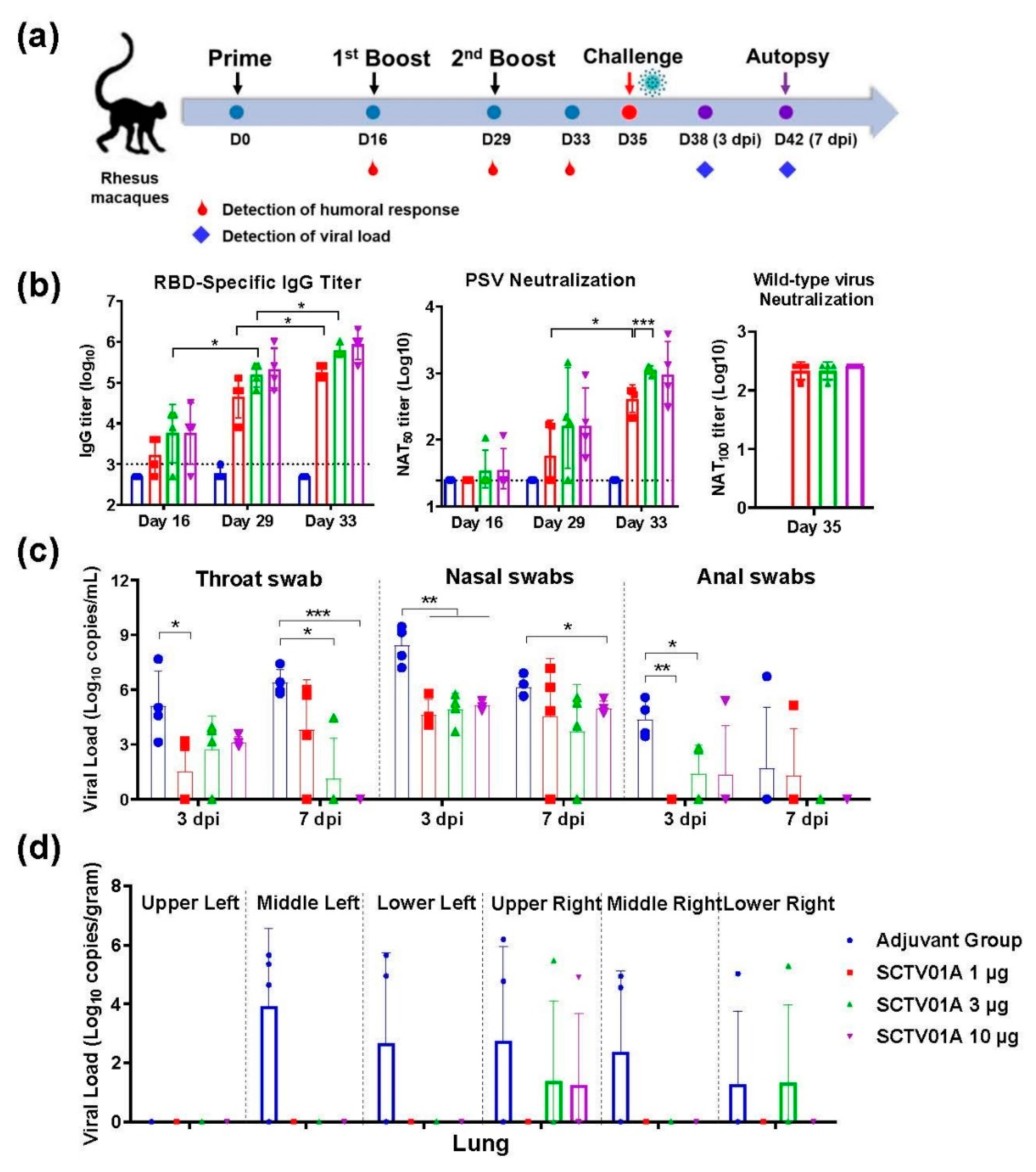
3.3. SCTV01A Induced a Protective Immune Response in Nonhuman Primates
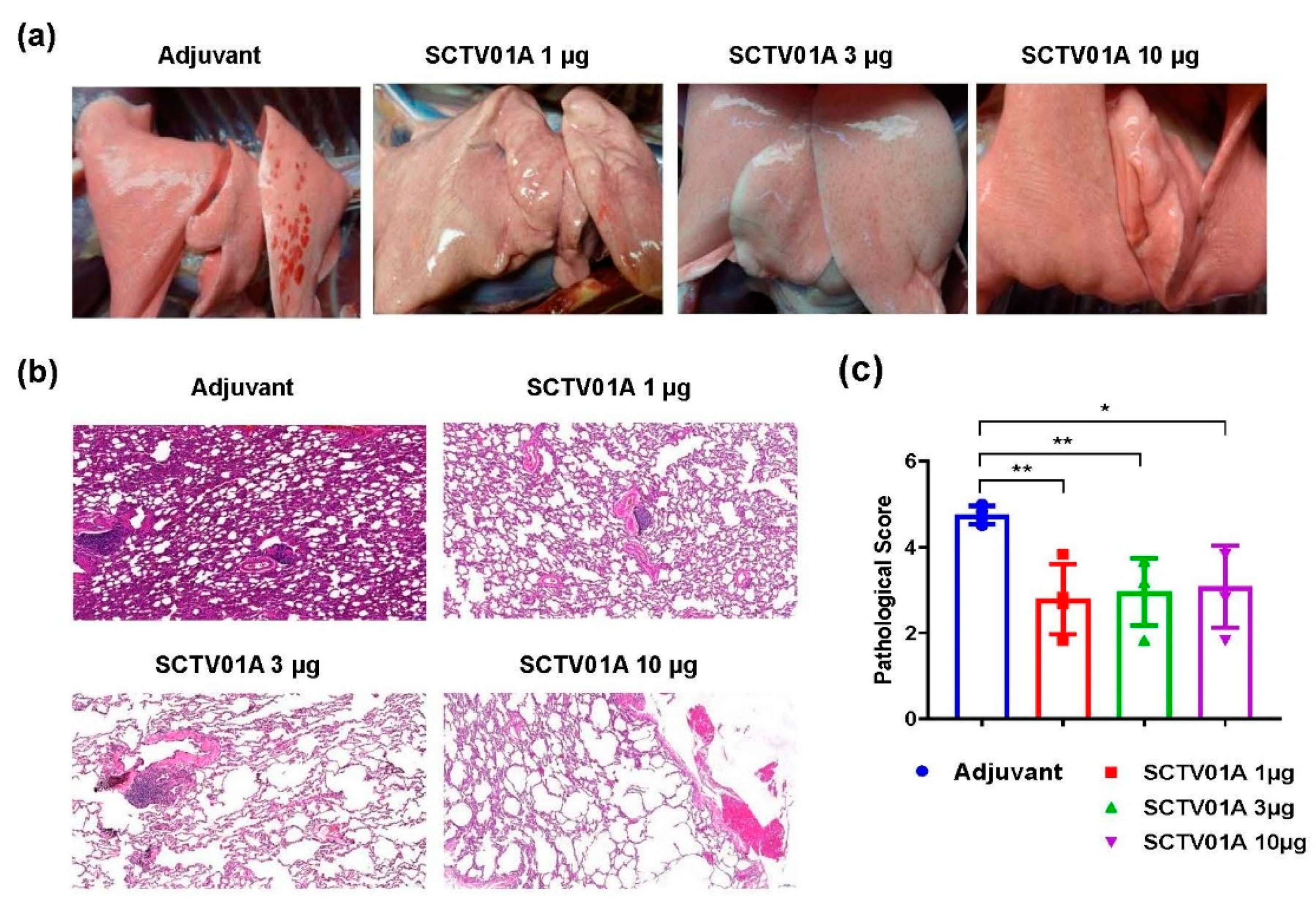
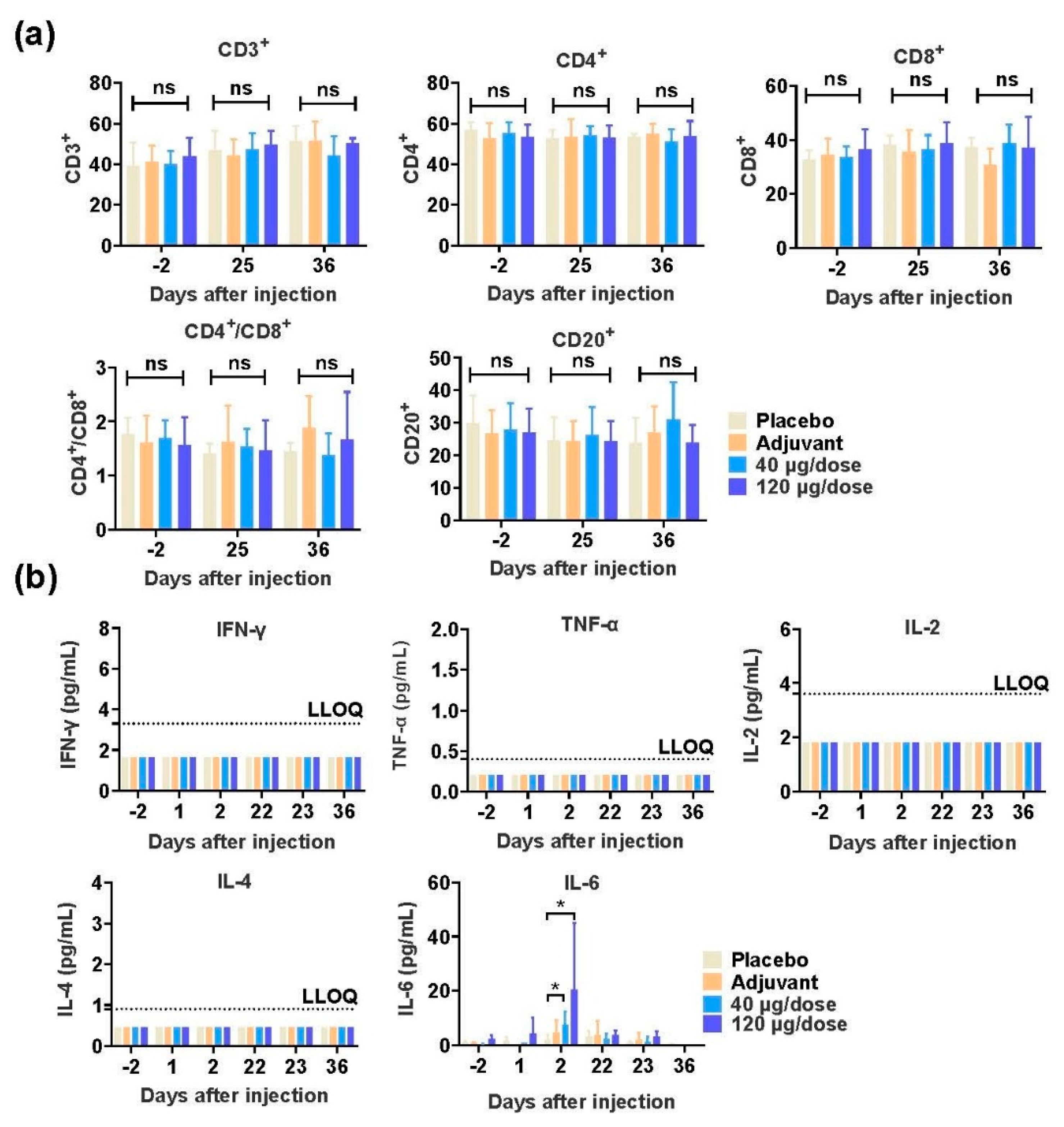
3.4. SCTV01A Is Safe in Rhesus Macaques
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kudlay, D.; Svistunov, A. COVID-19 Vaccines: An Overview of Different Platforms. Bioengineering 2022, 9, 72. [Google Scholar] [CrossRef] [PubMed]
- WHO. COVID-19 Vaccine Tracker and Landscape. Available online: https://www.who.int/publications/m/item/draft-landscape-of-covid-19-candidate-vaccines (accessed on 7 October 2022).
- Zhou, P.; Yang, X.L.; Wang, X.G.; Hu, B.; Zhang, L.; Zhang, W.; Si, H.R.; Zhu, Y.; Li, B.; Huang, C.L.; et al. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature 2020, 579, 270–273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoffmann, M.; Kleine-Weber, H.; Schroeder, S.; Krüger, N.; Herrler, T.; Erichsen, S.; Schiergens, T.S.; Herrler, G.; Wu, N.-H.; Nitsche, A. SARS-CoV-2 cell entry depends on ACE2 and TMPRSS2 and is blocked by a clinically proven protease inhibitor. Cell 2020, 181, 271–280.e8. [Google Scholar] [CrossRef]
- To, K.K.; Tsang, O.T.; Leung, W.S.; Tam, A.R.; Wu, T.C.; Lung, D.C.; Yip, C.C.; Cai, J.P.; Chan, J.M.; Chik, T.S.; et al. Temporal profiles of viral load in posterior oropharyngeal saliva samples and serum antibody responses during infection by SARS-CoV-2: An observational cohort study. Lancet Infect. Dis. 2020, 20, 565–574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walls, A.C.; Fiala, B.; Schäfer, A.; Wrenn, S.; Pham, M.N.; Murphy, M.; Tse, L.V.; Shehata, L.; O’Connor, M.A.; Chen, C.; et al. Elicitation of Potent Neutralizing Antibody Responses by Designed Protein Nanoparticle Vaccines for SARS-CoV-2. Cell 2020, 183, 1367–1382.e1317. [Google Scholar] [CrossRef]
- Liang, Y.; Zhang, J.; Yuan, R.Y.; Wang, M.Y.; He, P.; Su, J.G.; Han, Z.B.; Jin, Y.Q.; Hou, J.W.; Zhang, H. Design of a mutation-integrated trimeric RBD with broad protection against SARS-CoV-2. Cell Discov. 2022, 8, 17. [Google Scholar] [CrossRef]
- Xu, K.; Gao, P.; Liu, S.; Lu, S.; Lei, W.; Zheng, T.; Liu, X.; Xie, Y.; Zhao, Z.; Guo, S.; et al. Protective prototype-Beta and Delta-Omicron chimeric RBD-dimer vaccines against SARS-CoV-2. Cell 2022, 185, 2265–2278. [Google Scholar] [CrossRef]
- Baum, A.; Ajithdoss, D. REGN-COV2 antibodies prevent and treat SARS-CoV-2 infection in rhesus macaques and hamsters. Science 2020, 370, 1110–1115. [Google Scholar] [CrossRef]
- Zhu, L.; Deng, Y.-Q.; Zhang, R.-R.; Cui, Z.; Sun, C.-Y.; Fan, C.-F.; Xing, X.; Huang, W.; Chen, Q.; Zhang, N.-N.; et al. Double lock of a potent human therapeutic monoclonal antibody against SARS-CoV-2. Natl. Sci. Rev. 2020, 8, nwaa297. [Google Scholar] [CrossRef]
- Sun, C.; Chi, H.; Yuan, F.; Li, J.; Yang, J.; Zheng, A.; Wang, F.; Sun, L.; Zhang, Y.; Hu, P. An antibody cocktail with broadened mutational resistance and effective protection against SARS-CoV-2. Sci. China Life Sci. 2022, 66, 165–179. [Google Scholar] [CrossRef]
- Pinto, D.; Park, Y.J.; Beltramello, M.; Walls, A.C.; Tortorici, M.A. Cross-neutralization of SARS-CoV-2 by a human monoclonal SARS-CoV antibody. Nature 2020, 583, 290–295. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Jian, F.; Zhang, Z.; Yisimayi, A.; Hao, X.; Bao, L.; Yuan, F.; Yu, Y.; Du, S.; Wang, J.; et al. Rational identification of potent and broad sarbecovirus-neutralizing antibody cocktails from SARS convalescents. Cell Rep. 2022, 41, 111845. [Google Scholar] [CrossRef] [PubMed]
- Nimmerjahn, F.; Ravetch, J.V. Fcγ receptors as regulators of immune responses. Nat. Rev. Immunol. 2008, 8, 34–47. [Google Scholar] [CrossRef]
- Bournazos, S.; Ravetch, J.V. Fcγ receptor function and the design of vaccination strategies. Immunity 2017, 47, 224–233. [Google Scholar] [CrossRef] [PubMed]
- Regnault, A.; Lankar, D.; Lacabanne, V.; Rodriguez, A.; Théry, C.; Rescigno, M.; Saito, T.; Verbeek, S.; Bonnerot, C.; Ricciardi-Castagnoli, P. Fcγ receptor–mediated induction of dendritic cell maturation and major histocompatibility complex class I–restricted antigen presentation after immune complex internalization. J. Exp. Med. 1999, 189, 371–380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Czajkowsky, D.M.; Hu, J.; Shao, Z.; Pleass, R.J. Fc-fusion proteins: New developments and future perspectives. EMBO Mol. Med. 2012, 4, 1015–1028. [Google Scholar] [CrossRef]
- Li, Y.; Li, R.; Wang, M.; Liu, Y.; Yin, Y.; Zai, X.; Song, X.; Chen, Y.; Xu, J.; Chen, W. Fc-based recombinant henipavirus vaccines elicit broad neutralizing antibody responses in mice. Viruses 2020, 12, 480. [Google Scholar] [CrossRef] [Green Version]
- Loureiro, S.; Robinson, E.; Chen, H.; Phapugrangkul, P.; Colaco, C.; Jones, I.M. Virus Glycoproteins Tagged with the Human Fc Domain as Second Generation Vaccine Candidates. In Innovation in Vaccinology; Baschieri, S., Ed.; Springer: Dordrecht, The Netherlands, 2012; pp. 45–63. [Google Scholar]
- Jann, K.; Jann, B. Capsular polysaccharides. In Molecular Medical Microbiology; Academic Press: Cambridge, MA, USA, 2002; pp. 47–77. [Google Scholar]
- Lee, C.-J.; Lee, L.H.; Lu, C.-s.; Wu, A. Bacterial polysaccharides as vaccines—Immunity and chemical characterization. In The Molecular Immunology of Complex Carbohydrates—2; Springer: New York, NY, USA, 2001; pp. 453–471. [Google Scholar]
- Adams, E.W.; Ratner, D.M.; Seeberger, P.H.; Hacohen, N. Carbohydrate-mediated targeting of antigen to dendritic cells leads to enhanced presentation of antigen to T cells. Chembiochem 2008, 9, 294–303. [Google Scholar] [CrossRef] [Green Version]
- Balin, S.J.; Platt, J.L.; Cascalho, M. New insights into the functions of B cells. Pediatr. Transplant. 2008, 12, 510–515. [Google Scholar] [CrossRef] [Green Version]
- Zamze, S.; Martinez-Pomares, L.; Jones, H.; Taylor, P.R.; Stillion, R.J.; Gordon, S.; Wong, S.Y. Recognition of bacterial capsular polysaccharides and lipopolysaccharides by the macrophage mannose receptor. J. Biol. Chem. 2002, 277, 41613–41623. [Google Scholar] [CrossRef] [Green Version]
- Koppel, E.A.; Saeland, E.; De Cooker, D.J.; Van Kooyk, Y.; Geijtenbeek, T.B. DC-SIGN specifically recognizes Streptococcus pneumoniae serotypes 3 and 14. Immunobiology 2005, 210, 203–210. [Google Scholar] [CrossRef] [PubMed]
- Miyasaka, T.; Akahori, Y.; Toyama, M.; Miyamura, N.; Ishii, K.; Saijo, S.; Iwakura, Y.; Kinjo, Y.; Miyazaki, Y.; Oishi, K. Dectin-2-dependent NKT cell activation and serotype-specific antibody production in mice immunized with pneumococcal polysaccharide vaccine. PLoS ONE 2013, 8, e78611. [Google Scholar] [CrossRef] [PubMed]
- Meltzer, U.; Goldblatt, D. Pneumococcal polysaccharides interact with human dendritic cells. Infect. Immun. 2006, 74, 1890–1895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, Y.; Li, J.; Sun, C.; Chi, H.; Luo, D.; Wang, R.; Qiu, H.; Zhang, Y.; Wu, M.; Zhang, X. Rational Development of a Polysaccharide–Protein-Conjugated Nanoparticle Vaccine Against SARS-CoV-2 Variants and Streptococcus pneumoniae. Adv. Mater. 2022, 34, 2200443. [Google Scholar] [CrossRef]
- Carmenate, T.; Canaán, L.; Alvarez, A.; Delgado, M.; González, S.; Menéndez, T.; Rodés, L.; Guillén, G. Effect of conjugation methodology on the immunogenicity and protective efficacy of meningococcal group C polysaccharide-P64k protein conjugates. FEMS Immunol. Med. Microbiol. 2004, 40, 193–199. [Google Scholar] [CrossRef] [Green Version]
- Xie, T.; Lu, S.; He, Z.; Liu, H.; Wang, J.; Tang, C.; Yang, T.; Yu, W.; Li, H.; Yang, Y. Three doses of prototypic SARS-CoV-2 inactivated vaccine induce cross-protection against its variants of concern. Signal Transduct. Target. Ther. 2022, 7, 61. [Google Scholar] [CrossRef]
- Liu, Z.; Xu, W.; Xia, S.; Gu, C.; Wang, X.; Wang, Q.; Zhou, J.; Wu, Y.; Cai, X.; Qu, D. RBD-Fc-based COVID-19 vaccine candidate induces highly potent SARS-CoV-2 neutralizing antibody response. Signal Transduct. Target. Ther. 2020, 5, 282. [Google Scholar] [CrossRef]
- Liu, X.; Drelich, A.; Li, W.; Chen, C.; Sun, Z.; Shi, M.; Adams, C.; Mellors, J.W.; Tseng, C.-T.; Dimitrov, D.S. Enhanced elicitation of potent neutralizing antibodies by the SARS-CoV-2 spike receptor binding domain Fc fusion protein in mice. Vaccine 2020, 38, 7205–7212. [Google Scholar] [CrossRef]
- Sun, S.; He, L.; Zhao, Z.; Gu, H.; Fang, X.; Wang, T.; Yang, X.; Chen, S.; Deng, Y.; Li, J. Recombinant vaccine containing an RBD-Fc fusion induced protection against SARS-CoV-2 in nonhuman primates and mice. Cell. Mol. Immunol. 2021, 18, 1070–1073. [Google Scholar] [CrossRef]
- Song, J.Y.; Moseley, M.A.; Burton, R.L.; Nahm, M.H. Pneumococcal vaccine and opsonic pneumococcal antibody. J. Infect. Chemother. 2013, 19, 412–425. [Google Scholar] [CrossRef] [Green Version]
- Xie, J.; Zhang, Y.; Caro-Aguilar, I.; Indrawati, L.; Smith, W.J.; Giovarelli, C.; Winters, M.A.; MacNair, J.; He, J.; Abeygunawardana, C. Immunogenicity comparison of a next generation pneumococcal conjugate vaccine in animal models and human infants. Pediatr. Infect. Dis. J. 2020, 39, 70–77. [Google Scholar] [CrossRef]
- Sun, Y.-S.; Zhou, J.-J.; Zhu, H.-P.; Xu, F.; Zhao, W.-B.; Lu, H.-J.; Wang, Z.; Chen, S.-Q.; Yao, P.-P.; Jiang, J.-M. Development of a recombinant RBD subunit vaccine for SARS-CoV-2. Viruses 2021, 13, 1936. [Google Scholar] [CrossRef]
- Sun, S.; Cai, Y.; Song, T.-Z.; Pu, Y.; Cheng, L.; Xu, H.; Sun, J.; Meng, C.; Lin, Y.; Huang, H. Interferon-armed RBD dimer enhances the immunogenicity of RBD for sterilizing immunity against SARS-CoV-2. Cell Res. 2021, 31, 1011–1023. [Google Scholar] [CrossRef]
- Wilhelm, A.; Widera, M.; Grikscheit, K.; Toptan, T.; Schenk, B.; Pallas, C.; Metzler, M.; Kohmer, N.; Hoehl, S.; Helfritz, F.A. Reduced Neutralization of SARS-CoV-2 Omicron Variant by Vaccine Sera and Monoclonal Antibodies. 2021. Available online: https://www.medrxiv.org/content/10.1101/2021.12.07.21267432v4 (accessed on 13 December 2021).
- Roessler, A.; Riepler, L.; Bante, D.; von Laer, D.; Kimpel, J. SARS-CoV-2 B. 1.1. 529 Variant (Omicron) Evades Neutralization by Sera from Vaccinated and Convalescent Individuals. 2021. Available online: https://www.medrxiv.org/content/10.1101/2021.12.08.21267491v1 (accessed on 11 December 2021).
- Kurhade, C.; Zou, J.; Xia, H.; Liu, M.; Chang, H.C.; Ren, P.; Xie, X.; Shi, P.Y. Low neutralization of SARS-CoV-2 Omicron BA. 2.75. 2, BQ. 1.1, and XBB. 1 by parental mRNA vaccine or a BA. 5-bivalent booster. Nat. Med. 2022, 29, 344–347. [Google Scholar] [CrossRef]
- Cao, Y.; Yisimayi, A.; Jian, F.; Song, W.; Xiao, T.; Wang, L.; Du, S.; Wang, J.; Li, Q.; Chen, X.; et al. BA.2.12.1, BA.4 and BA.5 escape antibodies elicited by Omicron infection. Nature 2022, 608, 593–602. [Google Scholar] [CrossRef]
- Variant-Specific Immune Responses Following Primary and Boosted Vaccination. 2021. Available online: https://ir.novavax.com/download/2021-12-22-6-month-w-Omicron-P19-FINAL.pdf (accessed on 22 December 2021).
- Wang, Q.; Guo, Y.; Iketani, S.; Nair, M.S.; Li, Z.; Mohri, H.; Wang, M.; Yu, J.; Bowen, A.D.; Chang, J.Y.; et al. Antibody evasion by SARS-CoV-2 Omicron subvariants BA.2.12.1, BA.4, and BA.5. Nature 2022, 608, 603–608. [Google Scholar] [CrossRef]
- Garcia-Beltran, W.F.; St. Denis, K.J.; Hoelzemer, A.; Lam, E.C.; Nitido, A.D.; Sheehan, M.L.; Berrios, C.; Ofoman, O.; Chang, C.C.; Hauser, B.M.; et al. mRNA-based COVID-19 vaccine boosters induce neutralizing immunity against SARS-CoV-2 Omicron variant. Cell 2021, 185, 457–466. [Google Scholar] [CrossRef]
- Tuekprakhon, A.; Nutalai, R.; Dijokaite-Guraliuc, A.; Zhou, D.; Ginn, H.M.; Selvaraj, M.; Liu, C.; Mentzer, A.J.; Supasa, P.; Duyvesteyn, H.M.E.; et al. Antibody escape of SARS-CoV-2 Omicron BA.4 and BA.5 from vaccine and BA.1 serum. Cell 2022, 185, 2422–2433.e13. [Google Scholar] [CrossRef]
- Deliyannis, G.; Gherardin, N.A.; Wong, C.Y.; Grimley, S.L.; Cooney, J.P.; Redmond, S.; Ellenberg, P.; Davidson, K.; Mordant, F.L.; Smith, T.; et al. Broad Immunity to SARS-CoV-2 Variants of Concern Mediated by a SARS-CoV-2 Receptor-Binding Domain Protein Vaccine. medRxiv 2022. [Google Scholar] [CrossRef]
- Liang, J.G.; Su, D.; Song, T.-Z.; Zeng, Y.; Huang, W.; Wu, J.; Xu, R.; Luo, P.; Yang, X.; Zhang, X. S-Trimer, a COVID-19 subunit vaccine candidate, induces protective immunity in nonhuman primates. Nat. Commun. 2021, 12, 1346. [Google Scholar] [CrossRef]
- Sridhar, S.; Joaquin, A.; Bonaparte, M.I.; Bueso, A.; Chabanon, A.-L.; Chen, A.; Chicz, R.M.; Diemert, D.; Essink, B.J.; Fu, B. Safety and immunogenicity of an AS03-adjuvanted SARS-CoV-2 recombinant protein vaccine (CoV2 preS dTM) in healthy adults: Interim findings from a phase 2, randomised, dose-finding, multicentre study. Lancet Infect. Dis. 2022, 22, 636–648. [Google Scholar] [CrossRef] [PubMed]
- Goepfert, P.A.; Fu, B.; Chabanon, A.L.; Bonaparte, M.I.; Davis, M.G.; Essink, B.J.; Frank, I.; Haney, O.; Janosczyk, H.; Keefer, M.C.; et al. Safety and immunogenicity of SARS-CoV-2 recombinant protein vaccine formulations in healthy adults: Interim results of a randomised, placebo-controlled, phase 1-2, dose-ranging study. Lancet Infect. Dis. 2021, 21, 1257–1270. [Google Scholar] [CrossRef] [PubMed]
- Chappell, K.J.; Mordant, F.L.; Li, Z.; Wijesundara, D.K.; Ellenberg, P.; Lackenby, J.; Cheung, S.; Modhiran, N.; Avumegah, M.S.; Henderson, C.L. First Report of a Phase 1 Randomised Trial of Molecular Clamp-Stabilised Spike Protein-Based and MF59-Adjuvanted Vaccine for SARS-CoV-2. 2021. Available online: https://papers.ssrn.com/sol3/papers.cfm?abstract_id=3769210 (accessed on 1 February 2021).
- Rivera-Hernandez, T.; Carnathan, D.G.; Moyle, P.M.; Toth, I.; West, N.P.; Young, P.L.; Silvestri, G.; Walker, M.J. The contribution of non-human primate models to the development of human vaccines. Discov. Med. 2014, 18, 313. [Google Scholar] [PubMed]
- Kennedy, R.C.; Shearer, M.H.; Hildebrand, W. Nonhuman primate models to evaluate vaccine safety and immunogenicity. Vaccine 1997, 15, 903–908. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, C.; Kong, D.; Guo, E.; Zhao, J.; Jia, J.; Wang, R.; Ma, J.; Chen, M.; Lu, J.; Yu, C.; et al. A Polysaccharide-RBD-Fc-Conjugated COVID-19 Vaccine, SCTV01A, Showed High Immunogenicity and Low Toxicity in Animal Models. Vaccines 2023, 11, 526. https://doi.org/10.3390/vaccines11030526
Sun C, Kong D, Guo E, Zhao J, Jia J, Wang R, Ma J, Chen M, Lu J, Yu C, et al. A Polysaccharide-RBD-Fc-Conjugated COVID-19 Vaccine, SCTV01A, Showed High Immunogenicity and Low Toxicity in Animal Models. Vaccines. 2023; 11(3):526. https://doi.org/10.3390/vaccines11030526
Chicago/Turabian StyleSun, Chunyun, Desheng Kong, Erhong Guo, Jun Zhao, Jilei Jia, Rui Wang, Juan Ma, Meng Chen, Jianbo Lu, Chulin Yu, and et al. 2023. "A Polysaccharide-RBD-Fc-Conjugated COVID-19 Vaccine, SCTV01A, Showed High Immunogenicity and Low Toxicity in Animal Models" Vaccines 11, no. 3: 526. https://doi.org/10.3390/vaccines11030526