
Antiviral Activities of HIV-1-Specific Human Broadly Neutralizing Antibodies Are Isotype-Dependent

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells
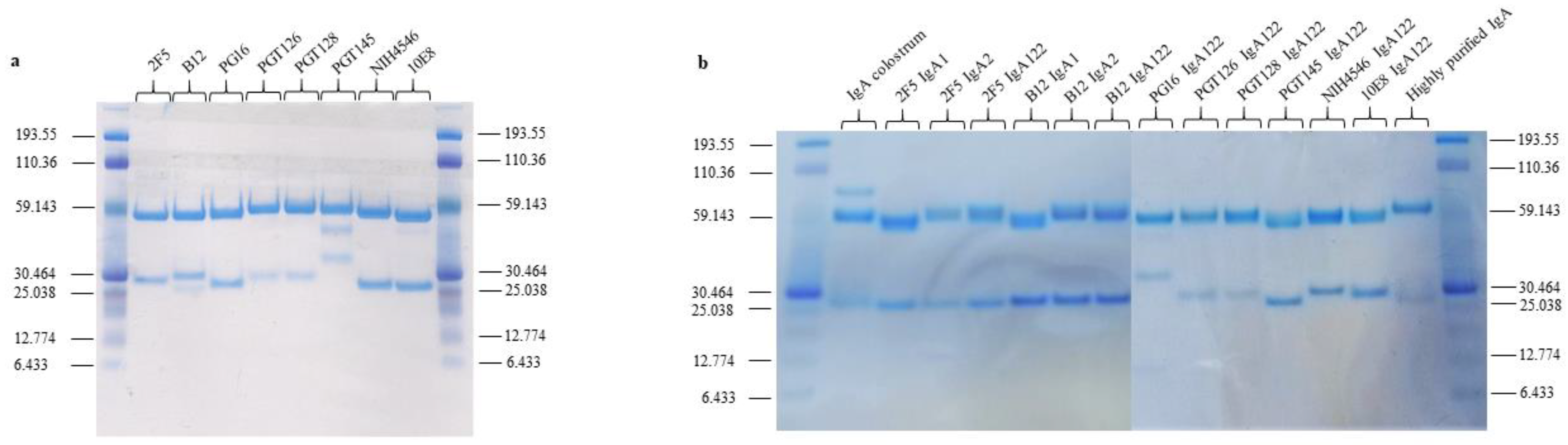
2.2. Construction and Production of Recombinant IgA1, IgA2, IgA122, and IgG
2.3. Analysis of the Specificity of the Different bNAb Isotypes
2.4. HIV-1-Specific Neutralization Assays
2.5. TZMbl/Pseudovirus Assay
2.6. PBMC/Primary Isolate Assay
2.7. HIV-1 Env-Dependent Antibody-Dependent Cellular Cytotoxicity (ADCC)-like Assay
2.8. Statistical Analysis
3. Results
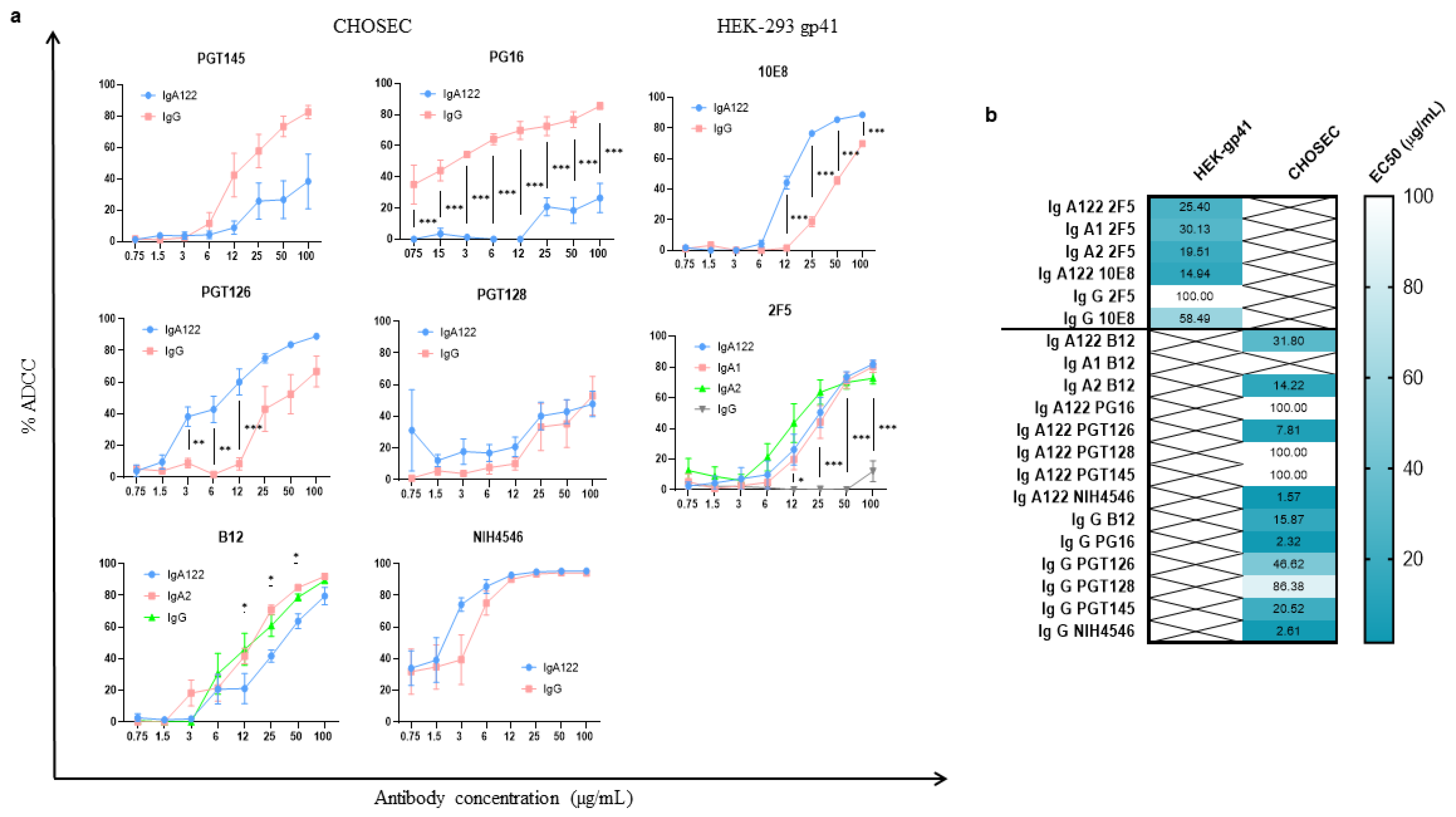
3.1. Development of a New Highly Potent Cellular Assay to Measure the ADCC-like Activity
3.2. ADCC-like Activities of bNAbs Depend on Their Isotypes and Their Specificity
3.2.1. Higher ADCC-like Activity of gp41-Specific IgA Isotypes
3.2.2. Heterogenous gp120-Specific ADCC-like Activity of IgG1 and IgA Isotypes
3.3. Development of a New Highly Potent Cellular Assay to Measure the ADCC-like Activity
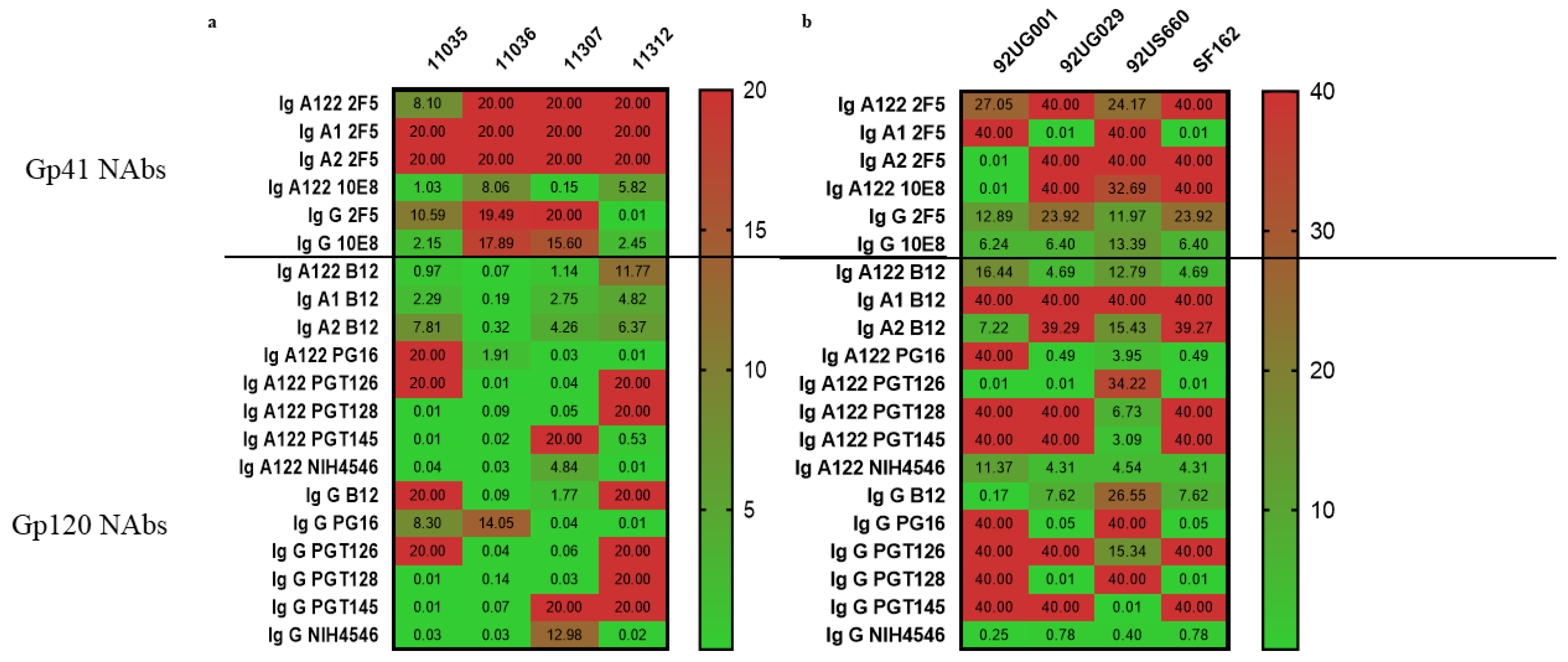
3.3.1. Gp41-Specific IgG1 Has More Potent Neutralizing Activity
3.3.2. Gp120-Specific IgG1 and IgAs Have Similar Neutralization Effect
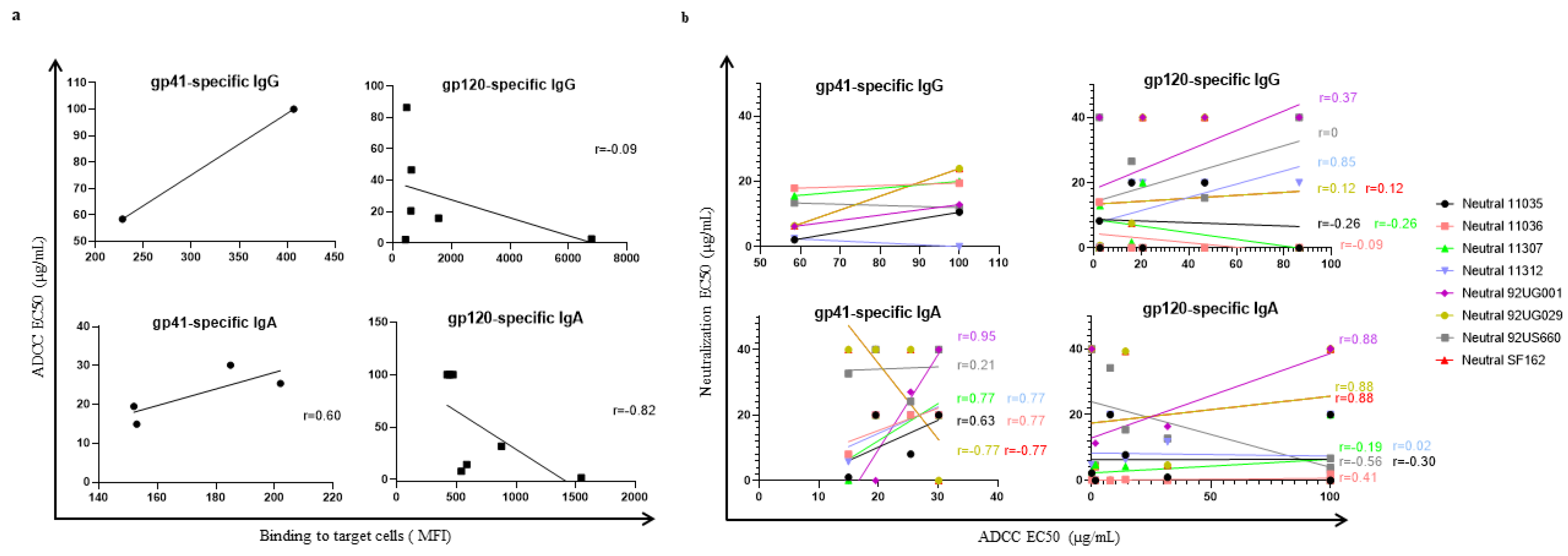
3.4. Correlation between ADCC-like Activity and Virus Neutralization
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fouda, G.G.; Eudailey, J.; Kunz, E.L.; Amos, J.D.; Liebl, B.E.; Himes, J.; Boakye-Agyeman, F.; Beck, K.; Michaels, A.J.; Cohen-Wolkowiez, M.; et al. Systemic Administration of an HIV-1 Broadly Neutralizing Dimeric IgA Yields Mucosal Secretory IgA and Virus Neutralization. Mucosal Immunol. 2017, 10, 228–237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watkins, J.D.; Sholukh, A.M.; Mukhtar, M.M.; Siddappa, N.B.; Lakhashe, S.K.; Kim, M.; Reinherz, E.L.; Gupta, S.; Forthal, D.N.; Sattentau, Q.J.; et al. Anti-HIV IgA Isotypes: Differential Virion Capture and Inhibition of Transcytosis Are Linked to Prevention of Mucosal R5 SHIV Transmission. AIDS 2013, 27, F13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torres, M.; Casadevall, A. The Immunoglobulin Constant Region Contributes to Affinity and Specificity. Trends Immunol. 2008, 29, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Tudor, D.; Yu, H.; Maupetit, J.; Drillet, A.S.; Bouceba, T.; Schwartz-Cornil, I.; Lopalco, L.; Tuffery, P.; Bomsel, M. Isotype Modulates Epitope Specificity, Affinity, and Antiviral Activities of Anti-HIV-1 Human Broadly Neutralizing 2F5 Antibody. Proc. Natl. Acad. Sci. USA 2012, 109, 12680–12685. [Google Scholar] [CrossRef] [Green Version]
- Astronomo, R.D.; Santra, S.; Ballweber-Fleming, L.; Westerberg, K.G.; Mach, L.; Hensley-McBain, T.; Sutherland, L.; Mildenberg, B.; Morton, G.; Yates, N.L.; et al. Neutralization Takes Precedence Over IgG or IgA Isotype-Related Functions in Mucosal HIV-1 Antibody-Mediated Protection. EBioMedicine 2016, 14, 97–111. [Google Scholar] [CrossRef] [Green Version]
- Wolbank, S.; Kunert, R.; Stiegler, G.; Katinger, H. Characterization of Human Class-Switched Polymeric (Immunoglobulin M [IgM] and IgA) Anti-Human Immunodeficiency Virus Type 1 Antibodies 2F5 and 2G12. J. Virol. 2003, 77, 4095–4103. [Google Scholar] [CrossRef] [Green Version]
- Shen, R.; Drelichman, E.R.; Bimczok, D.; Ochsenbauer, C.; Kappes, J.C.; Cannon, J.A.; Tudor, D.; Bomsel, M.; Smythies, L.E.; Smith, P.D. GP41-Specific Antibody Blocks Cell-Free HIV Type 1 Transcytosis through Human Rectal Mucosa and Model Colonic Epithelium. J. Immunol. 2010, 184, 3648–3655. [Google Scholar] [CrossRef] [Green Version]
- Mantis, N.J.; Palaia, J.; Hessell, A.J.; Mehta, S.; Zhu, Z.; Corthésy, B.; Neutra, M.R.; Burton, D.R.; Janoff, E.N. Inhibition of HIV-1 Infectivity and Epithelial Cell Transfer by Human Monoclonal IgG and IgA Antibodies Carrying the B12 V Region. J. Immunol. 2007, 179, 3144–3152. [Google Scholar] [CrossRef] [Green Version]
- Liu, F.; Bergami, P.L.; Duval, M.; Kuhrt, D.; Posner, M.; Cavacini, L. Expression and Functional Activity of Isotype and Subclass Switched Human Monoclonal Antibody Reactive with the Base of the V3 Loop of HIV-1 Gp120. AIDS Res. Hum. Retrovir. 2003, 19, 597–607. [Google Scholar] [CrossRef]
- Richardson, S.I.; Lambson, B.E.; Crowley, A.R.; Bashirova, A.; Scheepers, C.; Garrett, N.; Karim, S.A.; Mkhize, N.N.; Carrington, M.; Ackerman, M.E.; et al. IgG3 Enhances Neutralization Potency and Fc Effector Function of an HIV V2-Specific Broadly Neutralizing Antibody. PLoS Pathog. 2019, 15, e1008064. [Google Scholar] [CrossRef] [Green Version]
- Bournazos, S.; Klein, F.; Pietzsch, J.; Seaman, M.S.; Nussenzweig, M.C.; Ravetch, J.V. Broadly Neutralizing Anti-HIV-1 Antibodies Require Fc Effector Functions for in Vivo Activity. Cell 2014, 158, 1243–1253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mayr, L.; Su, B.; Moog, C. Role of Nonneutralizing Antibodies in Vaccines and/or HIV Infected Individuals. Curr. Opin. HIV AIDS 2017, 12, 209–215. [Google Scholar] [CrossRef] [PubMed]
- Bomsel, M.; Tudor, D.; Drillet, A.S.; Alfsen, A.; Ganor, Y.; Roger, M.G.; Mouz, N.; Amacker, M.; Chalifour, A.; Diomede, L.; et al. Immunization with HIV-1 Gp41 Subunit Virosomes Induces Mucosal Antibodies Protecting Nonhuman Primates against Vaginal SHIV Challenges. Immunity 2011, 34, 269–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chu, T.H.; Crowley, A.R.; Backes, I.; Chang, C.; Tay, M.; Broge, T.; Tuyishime, M.; Ferrari, G.; Seaman, M.S.; Richardson, S.I.; et al. Hinge Length Contributes to the Phagocytic Activity of HIV-Specific IgG1 and IgG3 Antibodies. PLoS Pathog. 2020, 16, e1008083. [Google Scholar] [CrossRef] [Green Version]
- Duchemin, M.; Khamassi, M.; Xu, L.; Tudor, D.; Bomsel, M. IgA Targeting Human Immunodeficiency Virus-1 Envelope Gp41 Triggers Antibody-Dependent Cellular Cytotoxicity Cross-Clade and Cooperates with Gp41-Specific IgG to Increase Cell Lysis. Front. Immunol. 2018, 9, 244. [Google Scholar] [CrossRef] [Green Version]
- Duchemin, M.; Tudor, D.; Cottignies-Calamarte, A.; Bomsel, M. Antibody-Dependent Cellular Phagocytosis of HIV-1-Infected Cells Is Efficiently Triggered by IgA Targeting HIV-1 Envelope Subunit Gp41. Front. Immunol. 2020, 11, 1141. [Google Scholar] [CrossRef]
- Jia, M.; Liberatore, R.A.; Guo, Y.; Chan, K.W.; Pan, R.; Lu, H.; Waltari, E.; Mittler, E.; Chandran, K.; Finzi, A.; et al. VSV-Displayed HIV-1 Envelope Identifies Broadly Neutralizing Antibodies Class-Switched to IgG and IgA. Cell Host Microbe 2020, 27, 963–975.e5. [Google Scholar] [CrossRef]
- Tomaras, G.D.; Plotkin, S.A. Complex Immune Correlates of Protection in HIV-1 Vaccine Efficacy Trials. Immunol. Rev. 2017, 275, 245–261. [Google Scholar] [CrossRef] [Green Version]
- Dawood, R.; Benjelloun, F.; Pin, J.J.; Kone, A.; Chanut, B.; Jospin, F.; Lucht, F.; Verrier, B.; Moog, C.; Genin, C.; et al. Generation of HIV-1 Potent and Broad Neutralizing Antibodies by Immunization with Postfusion HR1/HR2 Complex. AIDS 2013, 27, 717–730. [Google Scholar] [CrossRef]
- Montefiori, D.C. Evaluating Neutralizing Antibodies against HIV, SIV, and SHIV in Luciferase Reporter Gene Assays. Curr. Protoc. Immunol. 2004, 64, 125–131. [Google Scholar] [CrossRef]
- Weiss, C.D.; White, J.M. Characterization of Stable Chinese Hamster Ovary Cells Expressing Wild-Type, Secreted, and Glycosylphosphatidylinositol-Anchored Human Immunodeficiency Virus Type 1 Envelope Glycoprotein. J. Virol. 1993, 67, 7060–7066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Girard, A.; Jelicic, K.; Van Ryk, D.; Rochereau, N.; Cicala, C.; Arthos, J.; Noailly, B.; Genin, C.; Verrier, B.; Laurant, S.; et al. Neutralizing and Targeting Properties of a New Set of A4β7-Specific Antibodies Are Influenced by Their Isotype. J. Acquir. Immune Defic. Syndr. 2017, 75, 118–127. [Google Scholar] [CrossRef] [PubMed]
- Mascola, J.R.; Haynes, B.F. HIV-1 Neutralizing Antibodies: Understanding Nature’s Pathways. Immunol. Rev. 2013, 254, 225–244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, M.; Ruprecht, R.M. Are Anti-HIV IgAs Good Guys or Bad Guys? Retrovirology 2014, 11, 109. [Google Scholar] [CrossRef] [PubMed]
- Pancera, M.; Shahzad-Ul-Hussan, S.; Doria-Rose, N.A.; McLellan, J.S.; Bailer, R.T.; Dai, K.; Loesgen, S.; Louder, M.K.; Staupe, R.P.; Yang, Y.; et al. Structural Basis for Diverse N-Glycan Recognition by HIV-1-Neutralizing V1-V2-Directed Antibody PG16. Nat. Struct. Mol. Biol. 2013, 20, 804–813. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Ofek, G.; Laub, L.; Louder, M.K.; Doria-Rose, N.A.; Longo, N.S.; Imamichi, H.; Bailer, R.T.; Chakrabarti, B.; Sharma, S.K.; et al. Broad and Potent Neutralization of HIV-1 by a Gp41-Specific Human Antibody. Nature 2012, 491, 406–412. [Google Scholar] [CrossRef] [Green Version]
- Shibuya, A.; Sakamoto, N.; Shimizu, Y.; Shibuya, K.; Osawa, M.; Hiroyama, T.; Eyre, H.J.; Sutherland, G.R.; Endo, Y.; Fujita, T.; et al. Fcα/μ Receptor Mediates Endocytosis of IgM-Coated Microbes. Nat. Immunol. 2000, 1, 441–446. [Google Scholar] [CrossRef] [Green Version]
- Bruel, T.; Guivel-Benhassine, F.; Amraoui, S.; Malbec, M.; Richard, L.; Bourdic, K.; Donahue, D.A.; Lorin, V.; Casartelli, N.; Noël, N.; et al. Elimination of HIV-1-Infected Cells by Broadly Neutralizing Antibodies. Nat. Commun. 2016, 7, 10844. [Google Scholar] [CrossRef] [Green Version]
- von Bredow, B.; Arias, J.F.; Heyer, L.N.; Moldt, B.; Le, K.; Robinson, J.E.; Zolla-Pazner, S.; Burton, D.R.; Evans, D.T. Virus Neutralization by HIV-1 Env-Specific Monoclonal Antibodies. J. Virol. 2016, 90, 6127–6139. [Google Scholar] [CrossRef] [Green Version]
- Frey, G.; Peng, H.; Rits-Volloch, S.; Morelli, M.; Cheng, Y.; Chen, B. A Fusion-Intermediate State of HIV-1 Gp41 Targeted by Broadly Neutralizing Antibodies. Proc. Natl. Acad. Sci. USA 2008, 105, 3739–3744. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Frey, G.; Peng, H.; Rits-Volloch, S.; Garrity, J.; Seaman, M.S.; Chen, B. Mechanism of HIV-1 Neutralization by Antibodies Targeting a Membrane-Proximal Region of Gp41. J. Virol. 2014, 88, 1249–1258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cardoso, R.M.F.; Zwick, M.B.; Stanfield, R.L.; Kunert, R.; Binley, J.M.; Katinger, H.; Burton, D.R.; Wilson, I.A. Broadly Neutralizing Anti-HIV Antibody 4E10 Recognizes a Helical Conformation of a Highly Conserved Fusion-Associated Motif in Gp41. Immunity 2005, 22, 163–173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Irimia, A.; Sarkar, A.; Stanfield, R.L.; Wilson, I.A. Crystallographic Identification of Lipid as an Integral Component of the Epitope of HIV Broadly Neutralizing Antibody 4E10. Immunity 2016, 44, 21–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Von Bredow, B.; Andrabi, R.; Grunst, M.; Grandea, A.G., III; Le, K.; Song, G.; Berndsen, Z.T.; Porter, K.; Pallesen, J.; Ward, A.B.; et al. Differences in the Binding Affinity of an HIV-1 V2 Apex- Specific Antibody for the SIV Smm / Mac Envelope Glycoprotein Uncouple Antibody-Dependent Cellular Cytotoxicity From. MBio 2019, 10, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Tomaras, G.D.; Ferrari, G.; Shen, X.; Alam, S.M.; Liao, H.X.; Pollara, J.; Bonsignori, M.; Moody, M.A.; Fong, Y.; Chen, X.; et al. Vaccine-Induced Plasma IgA Specific for the C1 Region of the HIV-1 Envelope Blocks Binding and Effector Function of IgG. Proc. Natl. Acad. Sci. USA 2013, 110, 9019–9024. [Google Scholar] [CrossRef] [Green Version]
- Khamassi, M.; Xu, L.; Rey, J.; Duchemin, M.; Bouceba, T.; Tuffery, P.; Tudor, D.; Bomsel, M. The CH1α Domain of Mucosal Gp41 IgA Contributes to Antibody Specificity and Antiviral Functions in HIV-1 Highly Exposed Sero-Negative Individuals. PLoS Pathog. 2020, 16, e1009103. [Google Scholar] [CrossRef]
- Scheepers, C.; Bekker, V.; Anthony, C.; Richardson, S.I.; Oosthuysen, B.; Moyo, T.; Kgagudi, P.; Kitchin, D.; Nonyane, M.; York, T.; et al. Antibody Isotype Switching as a Mechanism to Counter HIV Neutralization Escape. Cell Rep. 2020, 33, 108430. [Google Scholar] [CrossRef]
- Sawyer, L.S.; Wrin, M.T.; Crawford-Miksza, L.; Potts, B.; Wu, Y.; Weber, P.A.; Alfonso, R.D.; Hanson, C. V Neutralization Sensitivity of Human Immunodeficiency Virus Type 1 Is Determined in Part by the Cell in Which the Virus Is Propagated. J. Virol. 1994, 68, 1342–1349. [Google Scholar] [CrossRef] [Green Version]
- Wrin, T.; Loh, T.P.; Vennari, J.C.; Schuitemaker, H.; Nunberg, J.H. Adaptation to Persistent Growth in the H9 Cell Line Renders a Primary Isolate of Human Immunodeficiency Virus Type 1 Sensitive to Neutralization by Vaccine Sera. J. Virol. 1995, 69, 39–48. [Google Scholar] [CrossRef] [Green Version]
- Hammonds, J.; Chen, X.; Ding, L.; Fouts, T.; De Vico, A.; Zur Megede, J.; Barnett, S.; Spearman, P. Gp120 Stability on HIV-1 Virions and Gag-Env Pseudovirions Is Enhanced by an Uncleaved Gag Core. Virology 2003, 314, 636–649. [Google Scholar] [CrossRef] [Green Version]
- Pugach, P.; Kuhmann, S.E.; Taylor, J.; Marozsan, A.J.; Snyder, A.; Ketas, T.; Wolinsky, S.M.; Korber, B.T.; Moore, J.P. The Prolonged Culture of Human Immunodeficiency Virus Type 1 in Primary Lymphocytes Increases Its Sensitivity to Neutralization by Soluble CD4. Virology 2004, 321, 8–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Protein | Epitope * | Antibody | Isotype | MFI | % of Positivity |
|---|---|---|---|---|---|
| gp41 (HEK-gp41) | MPER | 2F5 | IgG1 | 407 | 93.5% |
| IgA1 | 185 | 55.0% | |||
| IgA2 | 152 | 45.5% | |||
| IgA122 | 202 | 64.5% | |||
| 10E8 | IgG1 | 228 | 41.6% | ||
| IgA122 | 153 | 8.76% | |||
| gp140 (CHOSEC) | CD4 binding site | B12 | IgG1 | 1566 | 99.6% |
| IgA1 | 886 | 98.4% | |||
| IgA2 | 586 | 73.5% | |||
| IgA122 | 878 | 92.3% | |||
| NIH4546 | IgG1 | 6784 | 100% | ||
| IgA122 | 1548 | 99.4% | |||
| V2 glycan | PG16 | IgG1 | 439 | 0.049% | |
| IgA122 | 422 | 0.23% | |||
| PGT145 | IgG1 | 623 | 41.4% | ||
| IgA122 | 471 | 4.34% | |||
| V3 glycan | PGT126 | IgG1 | 638 | 47.6% | |
| IgA122 | 540 | 27.4% | |||
| PGT128 | IgG1 | 476 | 13.2% | ||
| IgA122 | 442 | 1.62% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Noailly, B.; Yaugel-Novoa, M.; Werquin, J.; Jospin, F.; Drocourt, D.; Bourlet, T.; Rochereau, N.; Paul, S. Antiviral Activities of HIV-1-Specific Human Broadly Neutralizing Antibodies Are Isotype-Dependent. Vaccines 2022, 10, 903. https://doi.org/10.3390/vaccines10060903
Noailly B, Yaugel-Novoa M, Werquin J, Jospin F, Drocourt D, Bourlet T, Rochereau N, Paul S. Antiviral Activities of HIV-1-Specific Human Broadly Neutralizing Antibodies Are Isotype-Dependent. Vaccines. 2022; 10(6):903. https://doi.org/10.3390/vaccines10060903
Chicago/Turabian StyleNoailly, Blandine, Melyssa Yaugel-Novoa, Justine Werquin, Fabienne Jospin, Daniel Drocourt, Thomas Bourlet, Nicolas Rochereau, and Stéphane Paul. 2022. "Antiviral Activities of HIV-1-Specific Human Broadly Neutralizing Antibodies Are Isotype-Dependent" Vaccines 10, no. 6: 903. https://doi.org/10.3390/vaccines10060903