
A Potential of an Anti-HTLV-I gp46 Neutralizing Monoclonal Antibody (LAT-27) for Passive Immunization against Both Horizontal and Mother-to-Child Vertical Infection with Human T Cell Leukemia Virus Type-I

Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Cell Culture and Syncytium Inhibition Assay
2.3. ELISA
2.4. Animal Experiments
2.5. Isolation of Human T-Cells from Mouse Spleen
2.6. Genomic DNA Extraction and Quantification of HTLV-I Proviral Load
2.7. Flow Cytometry
3. Results
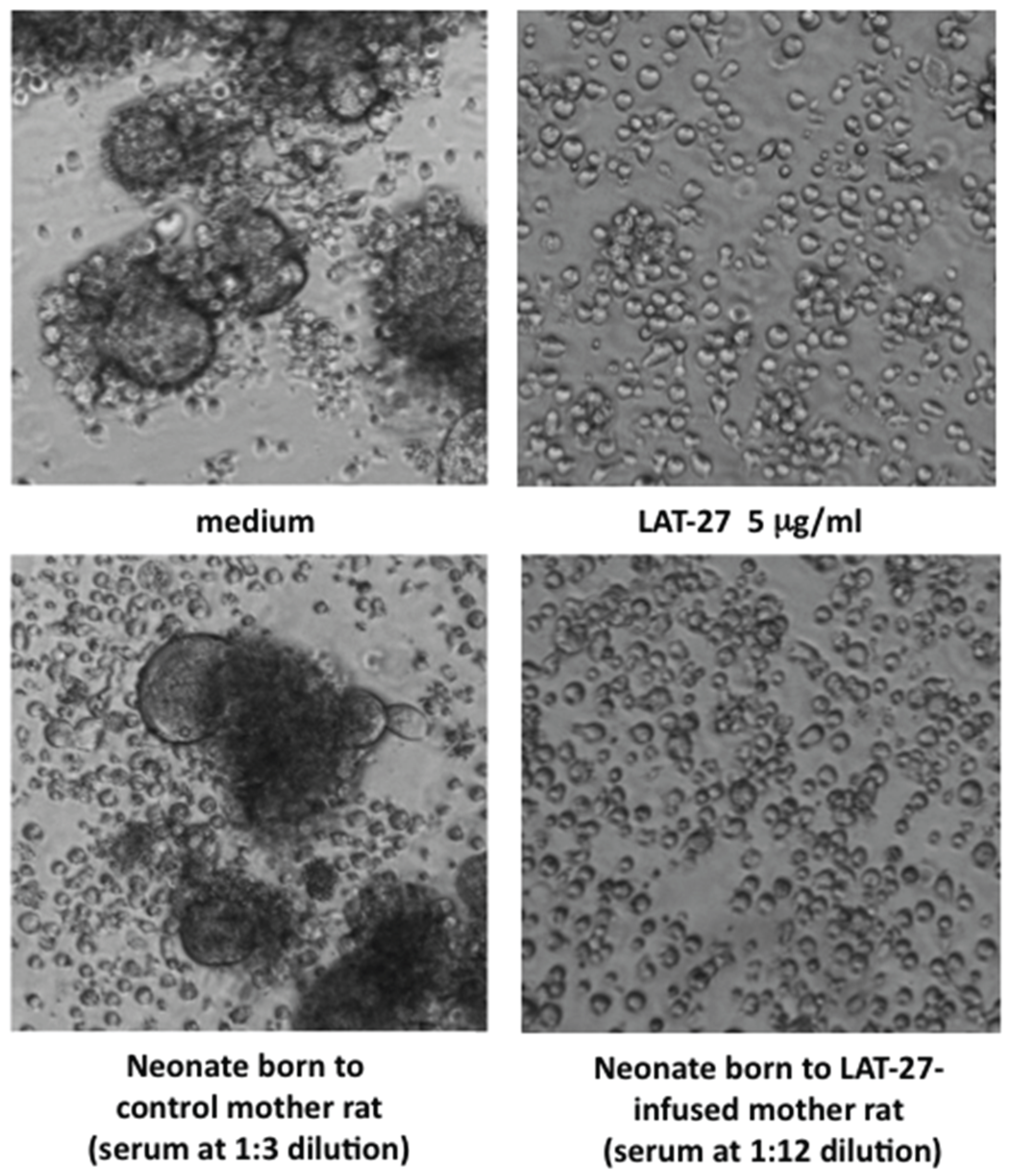
3.1. Mother-to-Child Transfer of Passively Immunized LAT-27

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pregnant Rat #1 | Serum LAT-27 conc. (μg/mL) | Pregnant Rat #2 | Serum LAT-27 conc. (μg/mL) | |
|---|---|---|---|---|
| Mother | 102 | Mother | 51 | |
| Offspring-1 | 204 | Offspring-1 | 102 | |
| Offspring-2 | 204 | Offspring-2 | 51 | |
| Offspring-3 | 102 | Offspring-3 | 102 | |
| Offspring-4 | 102 | Offspring-4 | 102 | |
| Offspring-5 | 204 | Offspring-5 | 51 | |
| Offspring-6 | 102 | Offspring-6 | 51 | |
| Offspring-7 | 204 | Offspring-7 | 51 | |
| Offspring-8 | 204 | Offspring-8 | 102 | |
| Offspring-9 | 102 | Offspring-9 | 102 | |
| Offspring-10 | 51 |

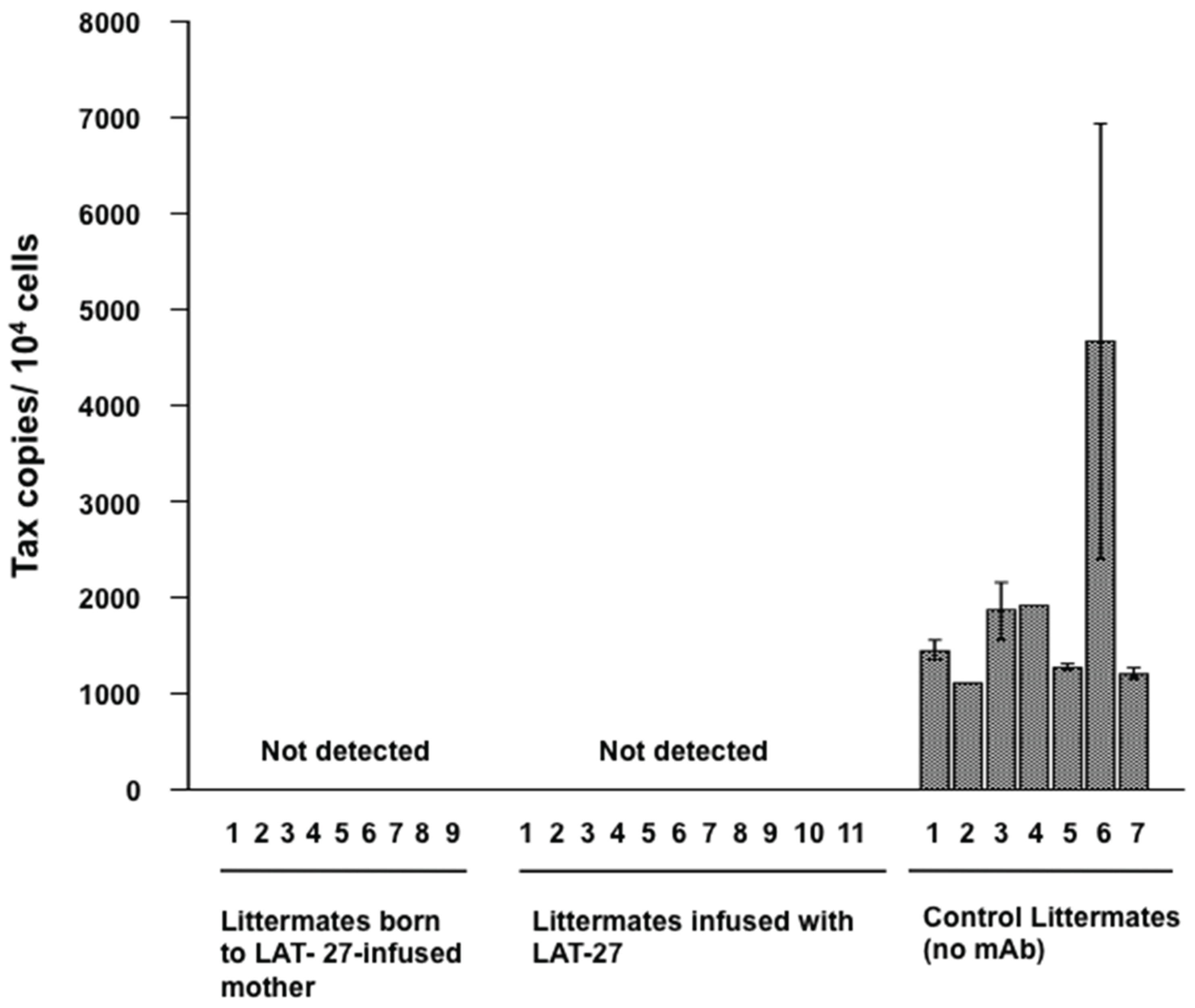
3.2. LAT-27 from Mothers Protects Newborns against HTLV-I Infection

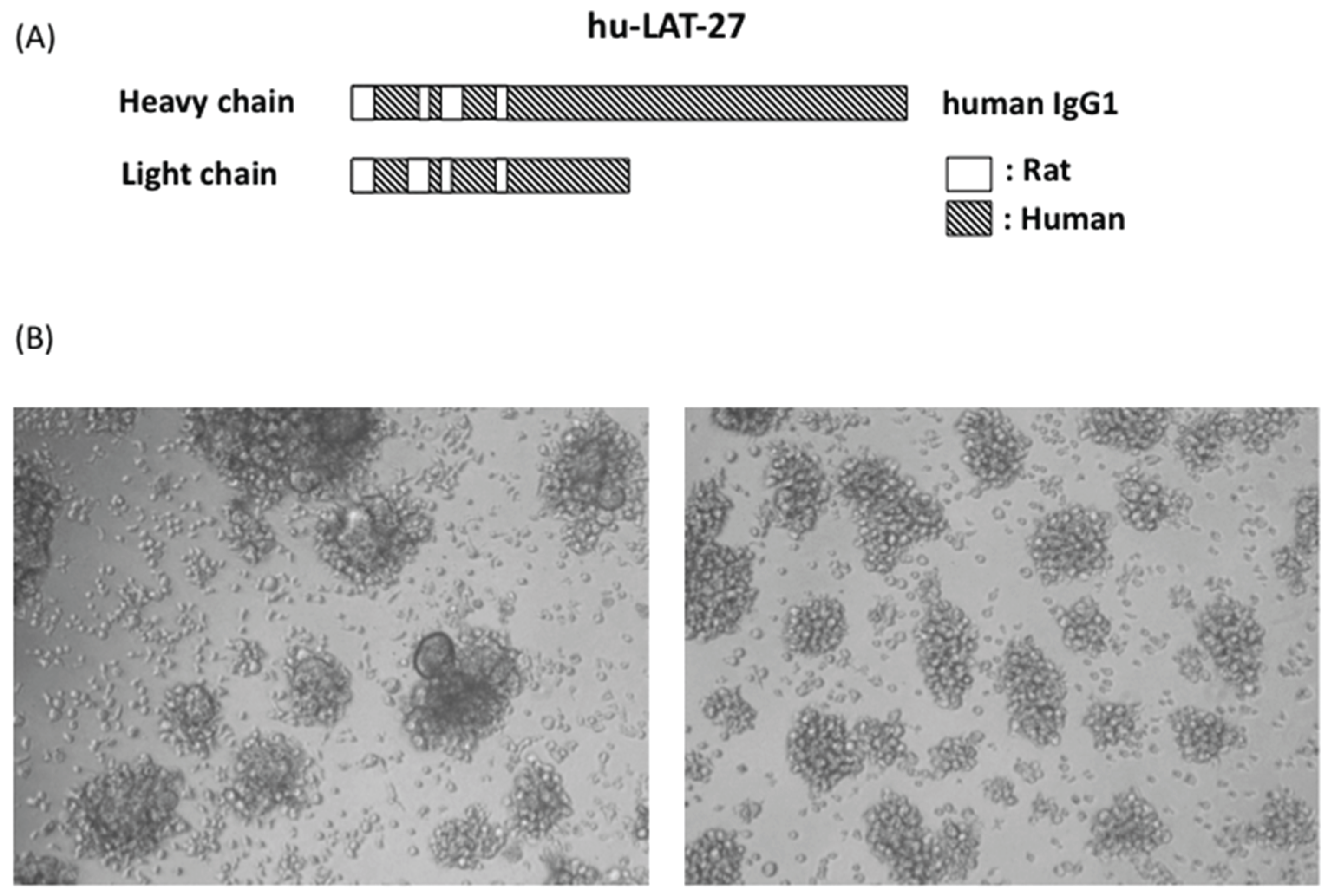
3.3. Retention of Humanized LAT-27 in NOG Mice

| Days after Injection | hu-LAT-27 Concentration in Serum (μg/mL) | ||
|---|---|---|---|
| NOG#1 | NOG#2 | NOG#3 | |
| 1 | 268 | 249 | 277 |
| 2 | 114 | 155 | 124 |
| 3 | 79 | 88 | 80 |
| 4 | 57 | 86 | 61 |
| 5 | 49 | 47 | 48 |
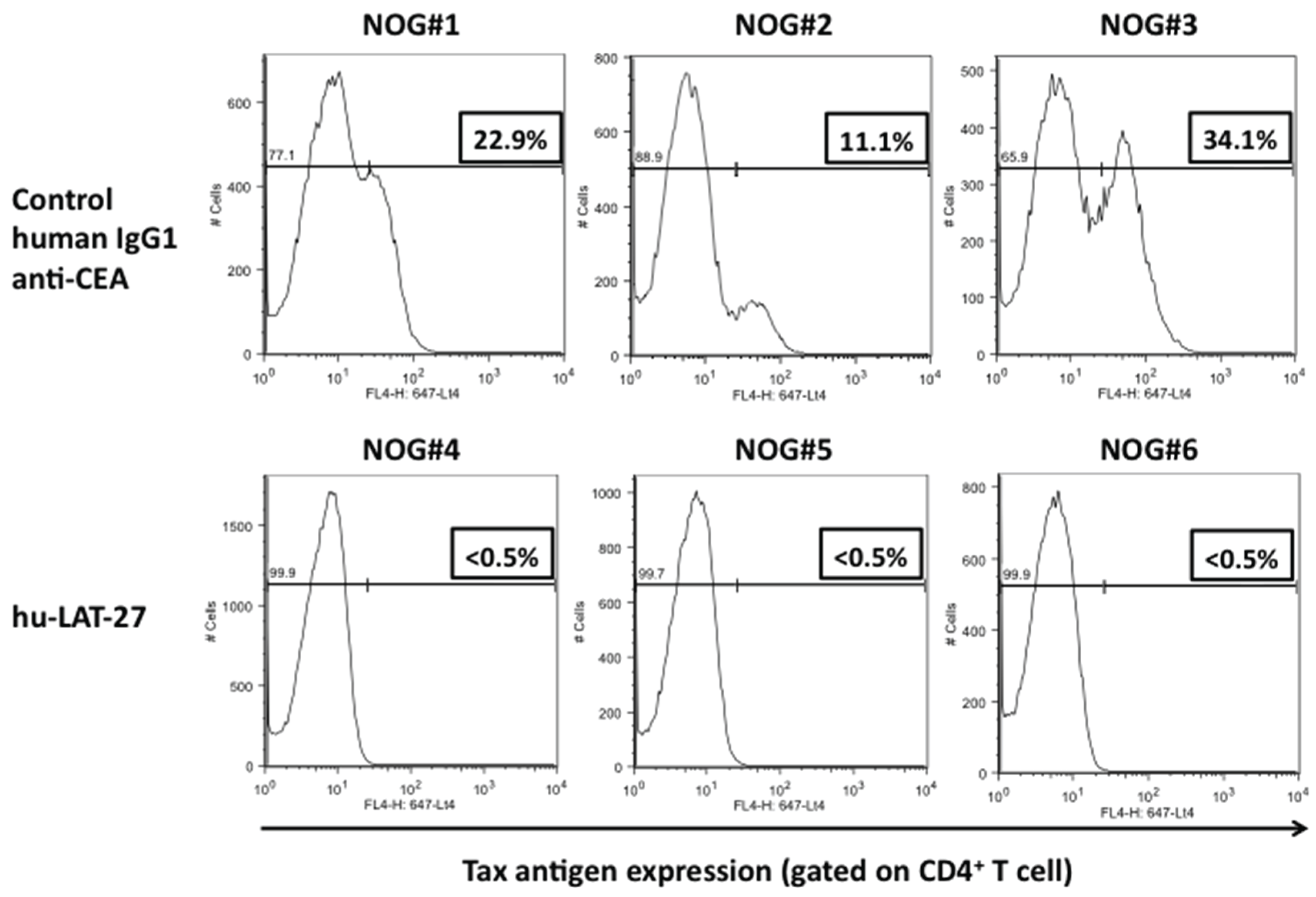
3.4. Protection of hu-PBL-NOG Mice against HTLV-I Infection by hu-LAT-27

4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Poiesz, B.J.; Ruscetti, F.W.; Gazdar, A.F.; Bunn, P.A.; Minna, J.D.; Gallo, R.C. Detection and isolation of type C retrovirus particles from fresh and cultured lymphocytes of a patient with cutaneous T-cell lymphoma. Proc. Natl. Acad. Sci. USA 1980, 77, 7415–7419. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, M.; Miyoshi, I.; Hinuma, Y. Isolation and characterization of retrovirus from cell lines of human adult T-cell leukemia and its implication in the disease. Proc. Natl. Acad. Sci. USA 1982, 79, 2031–2035. [Google Scholar] [CrossRef] [PubMed]
- Hinuma, Y.; Nagata, K.; Hanaoka, M.; Nakai, M.; Matsumoto, T.; Kinoshita, K.I.; Shirakawa, S.; Miyoshi, I. Adult T-cell leukemia: Antigen in an ATL cell line and detection of antibodies to the antigen in human sera. Proc. Natl. Acad. Sci. USA 1981, 78, 6476–6480. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, M.; Seiki, M.; Yamaguchi, K.; Takatsuki, K. Monoclonal integration of human T-cell leukemia provirus in all primary tumors of adult T-cell leukemia suggests causative role of human T-cell leukemia virus in the disease. Proc. Natl. Acad. Sci. USA 1984, 81, 2534–2537. [Google Scholar] [CrossRef] [PubMed]
- Gessain, A.; Barin, F.; Vernant, J.C.; Gout, O.; Maurs, L.; Calender, A.; de The, G. Antibodies to human T-lymphotropic virus type-I in patients with tropical spastic paraparesis. Lancet 1985, 2, 407–410. [Google Scholar] [CrossRef]
- Osame, M.; Usuku, K.; Izumo, S.; Ijichi, N.; Amitani, H.; Igata, A.; Matsumoto, M.; Tara, M. HTLV-I associated myelopathy, a new clinical entity. Lancet 1986, 1, 1031–1032. [Google Scholar] [CrossRef]
- Proietti, F.A.; Carneiro-Proietti, A.B.; Catalan-Soares, B.C.; Murphy, E.L. Global epidemiology of HTLV-I infection and associated diseases. Oncogene 2005, 24, 6058–6068. [Google Scholar] [CrossRef] [PubMed]
- Igakura, T.; Stinchcombe, J.C.; Goon, P.K.; Taylor, G.P.; Weber, J.N.; Griffiths, G.M.; Tanaka, Y.; Osame, M.; Bangham, C.R. Spread of HTLV-I between lymphocytes by virus-induced polarization of the cytoskeleton. Science 2003, 299, 1713–1716. [Google Scholar] [CrossRef] [PubMed]
- Kobe, B.; Center, R.J.; Kemp, B.E.; Poumbourios, P. Crystal structure of human T cell leukemia virus type 1 gp21 ectodomain crystallized as a maltose-binding protein chimera reveals structural evolution of retroviral transmembrane proteins. Proc. Natl. Acad. Sci. USA 1999, 96, 4319–4324. [Google Scholar] [CrossRef] [PubMed]
- Jones, K.S.; Lambert, S.; Bouttier, M.; Bénit, L.; Ruscetti, F.W.; Hermine, O.; Pique, C. Molecular aspects of HTLV-1 entry: Functional domains of the HTLV-1 surface subunit (SU) and their relationships to the entry receptors. Viruses 2011, 3, 794–810. [Google Scholar] [CrossRef] [PubMed]
- Gessain, A.; Gallo, R.C.; Franchini, G. Low degree of human T-cell leukemia/lymphoma virus type I genetic drift in vivo as a means of monitoring viral transmission and movement of ancient human populations. J. Virol. 1992, 66, 2288–2295. [Google Scholar] [PubMed]
- Tanaka, Y.; Zeng, L.; Shiraki, H.; Shida, H.; Tozawa, H. Identification of a neutralization epitope on the envelope gp46 antigen of human T cell leukemia virus type I and induction of neutralizing antibody by peptide immunization. J. Immunol. 1991, 147, 354–360. [Google Scholar] [PubMed]
- Tanaka, Y.; Tanaka, R.; Terada, E.; Koyanagi, Y.; Miyano-Kurosaki, N.; Yamamoto, N.; Baba, E.; Nakamura, M.; Shida, H. Induction of antibody responses that neutralize human T-cell leukemia virus type I infection in vitro and in vivo by peptide immunization. J. Virol. 1994, 68, 6323–6331. [Google Scholar] [PubMed]
- Baba, E.; Nakamura, M.; Tanaka, Y.; Kuroki, M.; Itoyama, Y.; Nakano, S.; Niho, Y. Multiple neutralizing B-cell epitopes of human T-cell leukemia virus type 1 (HTLV-1) identified by human monoclonal antibodies. A basis for the design of an HTLV-1 peptide vaccine. J. Immunol. 1993, 151, 1013–1024. [Google Scholar] [PubMed]
- Baba, E.; Nakamura, M.; Ohkuma, K.; Kira, J.; Tanaka, Y.; Nakano, S.; Niho, Y. A peptide-based human T cell leukemia virus type I vaccine containing T and B cell epitopes that induces high titers of neutralizing antibodies. J. Immunol. 1995, 154, 399–412. [Google Scholar] [PubMed]
- Goncalves, D.U.; Proietti, F.A.; Ribas, J.G.; Araújo, M.G.; Pinheiro, S.R.; Guedes, A.C.; Carneiro-Proietti, A.B. Epidemiology, treatment, and prevention of human T-cell leukemia virus type 1-associated diseases. Clin. Microbiol. Rev. 2010, 23, 577–589. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, Y.; Takahashi, Y.; Tanaka, R.; Kodama, A.; Fujii, H.; Hasegawa, A.; Kannagi, M.; Ansari, A.A.; Saito, M. Elimination of human T cell leukemia virus type-1 (HTLV-1)-infected cells by neutralizing and ADCC-inducing antibodies against HTLV-1 envelope gp46. AIDS Res. Hum. Retrovir. 2014, 30, 542–552. [Google Scholar] [CrossRef] [PubMed]
- Saito, M.; Tanaka, R.; Fujii, H.; Kodama, A.; Takahashi, Y.; Matsuzaki, T.; Takashima, H.; Tanaka, Y. The neutralizing function of the anti-HTLV-1 antibody is essential in preventing in vivo transmission of HTLV-1 to human T cells in NOD-SCID/γc null (NOG) mice. Retrovirology 2014, 11. [Google Scholar] [CrossRef]
- Tanaka, Y.; Yasumoto, M.; Nyunoya, H.; Ogura, T.; Kikuchi, M.; Shimotohno, K.; Shiraki, H.; Kuroda, N.; Shida, H.; Tozawa, H. Generation and characterization of monoclonal antibodies against multiple epitopes on the C-terminal half of envelope gp46 of human T-cell leukemia virus type-I (HTLV-I). Int. J. Cancer 1990, 46, 675–681. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, A.; Tanaka, R.; Murakami, T.; Takahashi, Y.; Koyanagi, Y.; Nakamura, M.; Ito, M.; Yamamoto, N.; Tanaka, Y. Induction of protective immune responses against R5 human immunodeficiency virus type 1 (HIV-1) infection in hu-PBL-SCID mice by intrasplenic immunization with HIV-1-pulsed dendritic cells: Possible involvement of a novel factor of human CD4+ T-cell origin. J. Virol. 2003, 77, 8719–8728. [Google Scholar] [CrossRef] [PubMed]
- Nagai, M.; Usuku, K.; Matsumoto, W.; Kodama, D.; Takenouchi, N.; Moritoyo, T.; Hashiguchi, S.; Ichinose, M.; Bangham, C.R.; Izumo, S.; et al. Analysis of HTLV-I proviral load in 202 HAM/TSP patients and 243 asymptomatic HTLV-I carriers: High proviral load strongly predisposes to HAM/TSP. J. Neurovirol. 1998, 4, 586–593. [Google Scholar] [CrossRef] [PubMed]
- Grindstaff, J.L.; Brodie, E.D., 3rd; Ketterson, E.D. Immune function across generations: Integrating mechanism and evolutionary process in maternal antibody transmission. Proc. Biol. Sci. 2003, 270, 2309–2319. [Google Scholar] [CrossRef] [PubMed]
- Komori, K.; Hasegawa, A.; Kurihara, K.; Honda, T.; Yokozeki, H.; Masuda, T.; Kannagi, M. Reduction of human T-cell leukemia virus type 1 (HTLV-1) proviral loads in rats orally infected with HTLV-1 by reimmunization with HTLV-1-infected cells. J. Virol. 2006, 80, 7375–7381. [Google Scholar] [CrossRef] [PubMed]
- Lobo, E.D.; Hansen, R.J.; Balthasar, J.P. Antibody pharmacokinetics and pharmacodynamics. J. Pharm. Sci. 2004, 93, 2645–2668. [Google Scholar] [CrossRef] [PubMed]
- Miyoshi, I.; Takehara, N.; Sawada, T.; Iwahara, Y.; Kataoka, R.; Yang, D.; Hoshino, H. Immunoglobulin prophylaxis against HTLV-I in a rabbit model. Leukemia 1992, 6, S24–S26. [Google Scholar]
- Sawada, T.; Iwahara, Y.; Ishii, K.; Taguchi, H.; Hoshino, H.; Miyoshi, I. Immunoglobulin prophylaxis against milkborne transmission of human T cell leukemia virus type I in rabbits. J. Infect. Dis. 1991, 164, 1193–1196. [Google Scholar] [CrossRef] [PubMed]
- Murata, N.; Hakoda, E.; Machida, H.; Ikezoe, T.; Sawada, T.; Hoshino, H.; Miyoshi, I. Prevention of human T cell lymphotropic virus type I infection in Japanese macaques by passive immunization. Leukemia 1996, 10, 1971–1974. [Google Scholar] [PubMed]
- Kuroki, M.; Nakamura, M.; Itoyama, Y.; Tanaka, Y.; Shiraki, H.; Baba, E.; Esaki, T.; Tatsumoto, T.; Nagafuchi, S.; Nakano, S.; et al. Identification of new epitopes recognized by human monoclonal antibodies with neutralizing and antibody-dependent cellular cytotoxicity activities specific for human T cell leukemia virus type 1. J. Immunol. 1992, 149, 940–948. [Google Scholar] [PubMed]
- Gövert, F.; Krumbholz, A.; Witt, K.; Hopfner, F.; Feldkamp, T.; Korn, K.; Knöll, A.; Jansen, O.; Deuschl, G.; Fickenscher, H. HTLV-1 associated myelopathy after renal transplantation. J. Clin. Virol. 2015, 72, 102–105. [Google Scholar] [CrossRef] [PubMed]
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fujii, H.; Shimizu, M.; Miyagi, T.; Kunihiro, M.; Tanaka, R.; Takahashi, Y.; Tanaka, Y. A Potential of an Anti-HTLV-I gp46 Neutralizing Monoclonal Antibody (LAT-27) for Passive Immunization against Both Horizontal and Mother-to-Child Vertical Infection with Human T Cell Leukemia Virus Type-I. Viruses 2016, 8, 41. https://doi.org/10.3390/v8020041
Fujii H, Shimizu M, Miyagi T, Kunihiro M, Tanaka R, Takahashi Y, Tanaka Y. A Potential of an Anti-HTLV-I gp46 Neutralizing Monoclonal Antibody (LAT-27) for Passive Immunization against Both Horizontal and Mother-to-Child Vertical Infection with Human T Cell Leukemia Virus Type-I. Viruses. 2016; 8(2):41. https://doi.org/10.3390/v8020041
Chicago/Turabian StyleFujii, Hideki, Mamoru Shimizu, Takuya Miyagi, Marie Kunihiro, Reiko Tanaka, Yoshiaki Takahashi, and Yuetsu Tanaka. 2016. "A Potential of an Anti-HTLV-I gp46 Neutralizing Monoclonal Antibody (LAT-27) for Passive Immunization against Both Horizontal and Mother-to-Child Vertical Infection with Human T Cell Leukemia Virus Type-I" Viruses 8, no. 2: 41. https://doi.org/10.3390/v8020041