
Lavandula austroapennina: Assessment of the Antiviral Activity of Lipophilic Extracts from Its Organs

 , , , , , , and
, , , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Extraction
2.2. UHPLC-ESI-QqTOF-MS/MS Analyses
2.3. Cell Lines and Cytotoxicity Test
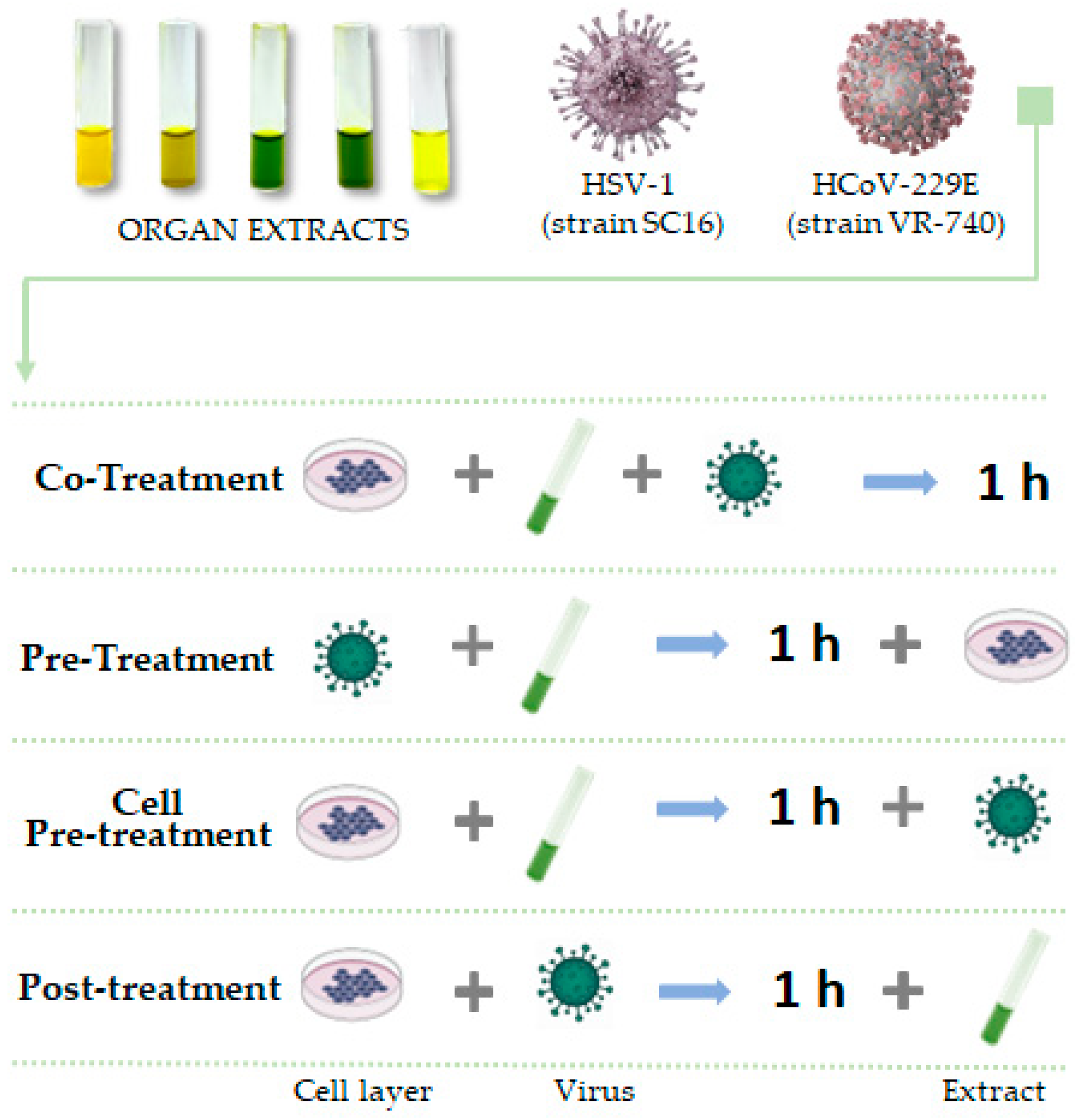
2.4. Virus and Antiviral Activity
- Co-treatment assay: the monolayer was treated and infected simultaneously with extracts and viruses for 1 h;
- Pre-treatment assay: Extracts were previously incubated for 1 h with viruses at 0.1 MOI at 37 °C. Cells were then infected with dilutions of the viral mixture;
- Cell pre-treatment: cell monolayer was pre-treated with compounds for 1 h and then infected with virus;
- Post-treatment: cell monolayer was first infected with virus, then treated with extracts.
2.5. Statistical Analysis
3. Results
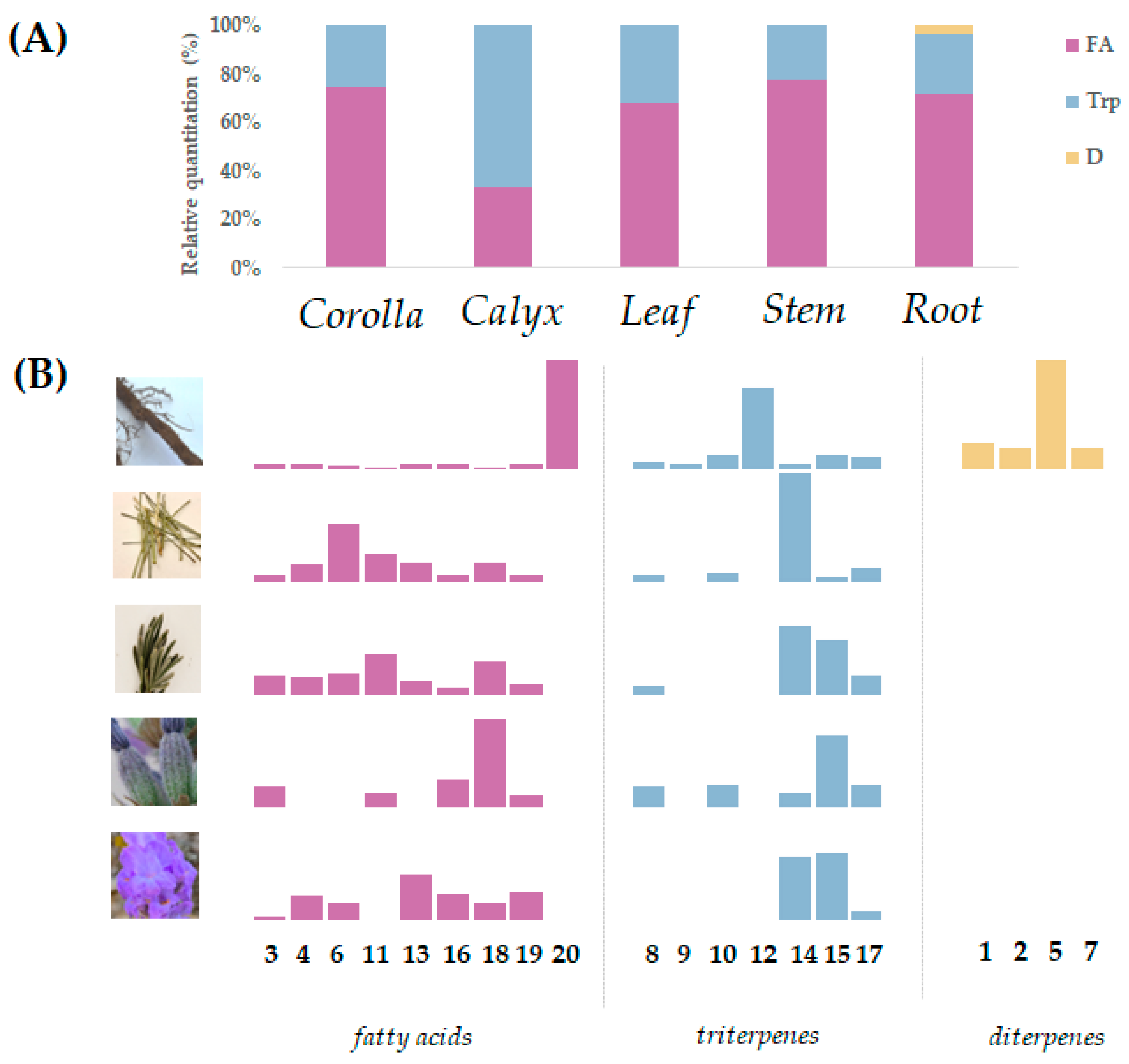
3.1. Lipid Profile of the Different L. austroapennina Organs
3.2. Cytotoxicity Screening of Apolar Extracts from L. austroapennina Organs
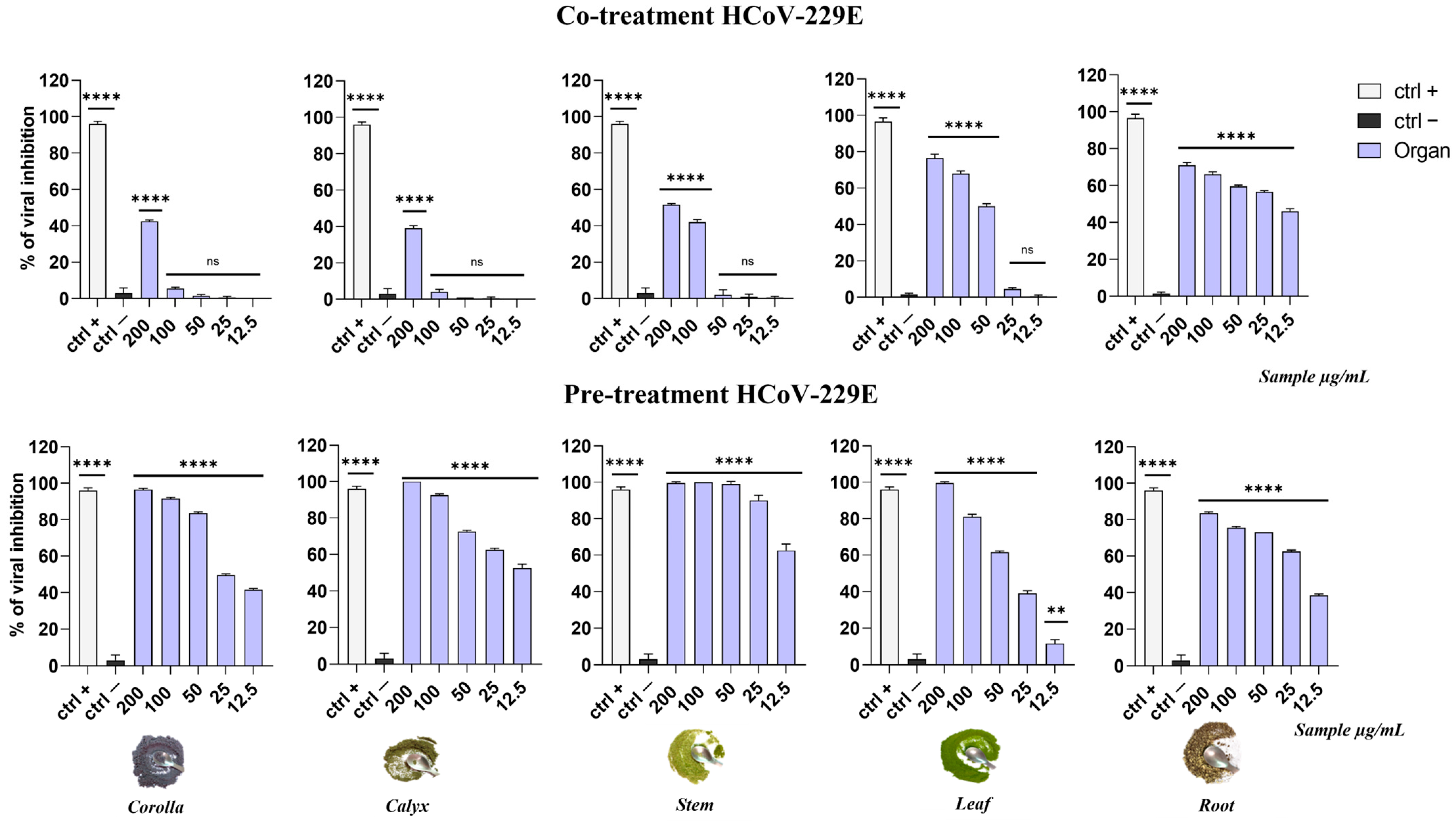
3.3. Antiviral Activity of Apolar Extracts from L. austroapennina Organs against HSV-1 and HCoV-229E
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Johnson, W.E. Origins and evolutionary consequences of ancient endogenous retroviruses. Nat. Rev. Microbiol. 2019, 17, 355–370. [Google Scholar] [CrossRef] [PubMed]
- Bachar, S.C.; Mazumder, K.; Bachar, R.; Aktar, A.; Al Mahtab, M. A Review of Medicinal Plants with Antiviral Activity Available in Bangladesh and Mechanistic Insight Into Their Bioactive Metabolites on SARS-CoV-2, HIV and HBV. Front. Pharmacol. 2021, 12, 732891. [Google Scholar] [CrossRef] [PubMed]
- Petrillo, F.; Chianese, A.; De Bernardo, M.; Zannella, C.; Galdiero, M.; Reibaldi, M.; Avitabile, T.; Boccia, G.; Galdiero, M.; Rosa, N.; et al. Inhibitory Effect of Ophthalmic Solutions against SARS-CoV-2: A Preventive Action to Block the Viral Transmission? Microorganisms 2021, 9, 1550. [Google Scholar] [CrossRef]
- Zannella, C.; Chianese, A.; Palomba, L.; Marcocci, M.E.; Bellavita, R.; Merlino, F.; Grieco, P.; Folliero, V.; De Filippis, A.; Mangoni, M.; et al. Broad-Spectrum Antiviral Activity of the Amphibian Antimicrobial Peptide Temporin L and Its Analogs. Int. J. Mol. Sci. 2022, 23, 2060. [Google Scholar] [CrossRef] [PubMed]
- Stelitano, D.; Franci, G.; Chianese, A.; Galdiero, S.; Morelli, G.; Galdiero, M. HSV membrane glycoproteins, their function in viral entry and their use in vaccine studies. Amino Acids Pept. Proteins 2019, 43, 14–43. [Google Scholar] [CrossRef]
- Kausar, S.; Khan, F.S.; Rehman, M.I.M.U.; Akram, M.; Riaz, M.; Rasool, G.; Khan, A.H.; Saleem, I.; Shamim, S.; Malik, A. A review: Mechanism of action of antiviral drugs. Int. J. Immunopathol. Pharmacol. 2021, 35. [Google Scholar] [CrossRef] [PubMed]
- Mahajan, S.; Choudhary, S.; Kumar, P.; Tomar, S. Antiviral strategies targeting host factors and mechanisms obliging +ssRNA viral pathogens. Bioorganic Med. Chem. 2021, 46, 116356. [Google Scholar] [CrossRef]
- Pagano, E. Phytocompounds and COVID-19: Two years of knowledge. Phytotherapy Res. 2022, 36, 2267–2271. [Google Scholar] [CrossRef]
- Raihan, T.; Rabbee, M.F.; Roy, P.; Choudhury, S.; Baek, K.-H.; Azad, A.K. Microbial Metabolites: The Emerging Hotspot of Antiviral Compounds as Potential Candidates to Avert Viral Pandemic Alike COVID-19. Front. Mol. Biosci. 2021, 8, 732256. [Google Scholar] [CrossRef]
- Ponticelli, M.; Bellone, M.L.; Parisi, V.; Iannuzzi, A.; Braca, A.; de Tommasi, N.; Russo, D.; Sileo, A.; Quaranta, P.; Freer, G.; et al. Specialized metabolites from plants as a source of new multi-target antiviral drugs: A systematic review. Phytochem. Rev. 2023, 22, 615–693. [Google Scholar] [CrossRef]
- Fiorentino, M.; Piccolella, S.; Gravina, C.; Stinca, A.; Esposito, A.; Catauro, M.; Pacifico, S. Encapsulating Calendula arvensis (Vaill.) L. Florets: UHPLC-HRMS Insights into Bioactive Compounds Preservation and Oral Bioaccessibility. Molecules 2022, 28, 199. [Google Scholar] [CrossRef]
- World Health Organization. WHO Traditional Medicine Strategy: 2014–2023; World Health Organization: Geneva, Switzerland, 2013; ISBN 978-92-4-150609-0. [Google Scholar]
- Ali, S.I.; Sheikh, W.M.; Rather, M.A.; Venkatesalu, V.; Bashir, S.M.; Nabi, S.U. Medicinal plants: Treasure for antiviral drug discovery. Phytother. Res. 2021, 35, 3447–3483. [Google Scholar] [CrossRef] [PubMed]
- Algieri, F.; Rodriguez-Nogales, A.; Vezza, T.; Garrido-Mesa, J.; Garrido-Mesa, N.; Utrilla, M.P.; González-Tejero, M.R.; Casares-Porcel, M.; Molero-Mesa, J.; Contreras, M.d.M.; et al. Anti-inflammatory activity of hydroalcoholic extracts of Lavandula dentata L. and Lavandula stoechas L. J. Ethnopharmacol. 2016, 190, 142–158. [Google Scholar] [CrossRef] [PubMed]
- Dobros, N.; Zawada, K.D.; Paradowska, K. Phytochemical Profiling, Antioxidant and Anti-Inflammatory Activity of Plants Belonging to the Lavandula Genus. Molecules 2022, 28, 256. [Google Scholar] [CrossRef] [PubMed]
- Conforti, F.; Perri, M.R.; Guerrini, A.; Sacchetti, G.; Statti, G. Lavandula austroapennina and Lavandula angustifolia essential oils and bioactive components: In vitro anti-denaturation effect of lavender from the Pollino massif (Southern Italy). Plant Biosyst. Int. J. Deal. all Asp. Plant Biol. 2023, 157, 339–345. [Google Scholar] [CrossRef]
- Baker, D.H.A.; Amarowicz, R.; Kandeil, A.; Ali, M.A.; Ibrahim, E.A. Antiviral activity of Lavandula angustifolia L. and Salvia officinalis L. essential oils against avian influenza H5N1 virus. J. Agric. Food Res. 2021, 4, 100135. [Google Scholar] [CrossRef] [PubMed]
- Hashem-Dabaghian, F.; Azimi, S.A.; Bahrami, M.; Latifi, S.-A.; Enayati, A.; Qaraaty, M. Effect of Lavender (Lavandula angustifolia L.) syrup on olfactory dysfunction in COVID-19 infection: A pilot controlled clinical trial. Avicenna J. Phytomedicine 2022, 12, 1–7. [Google Scholar] [CrossRef]
- Gravina, C.; Fiorentino, M.; Formato, M.; Pecoraro, M.T.; Piccolella, S.; Stinca, A.; Pacifico, S.; Esposito, A. LC-HR/MS Analysis of Lipophilic Extracts from Calendula arvensis (Vaill.) L. Organs: An Unexplored Source in Cosmeceuticals. Molecules 2022, 27, 8905. [Google Scholar] [CrossRef]
- Gravina, C.; Formato, M.; Piccolella, S.; Fiorentino, M.; Stinca, A.; Pacifico, S.; Esposito, A. Lavandula austroapennina (Lamiaceae): Getting Insights into Bioactive Polyphenols of a Rare Italian Endemic Vascular Plant. Int. J. Mol. Sci. 2023, 24, 8038. [Google Scholar] [CrossRef]
- Upson, T.; Andrews, S.; Royal Botanic Gardens, K. The Genus Lavandula; Royal Botanic Gardens: Kew, UK, 2004. [Google Scholar]
- Passalacqua, N.G.; Tundis, R.; Upson, T.M. A new species of Lavandula sect. Lavandula (Lamiaceae) and review of species boundaries in Lavandula angustifolia. Phytotaxa 2017, 292, 161–170. [Google Scholar] [CrossRef]
- Despinasse, Y.; Moja, S.; Soler, C.; Jullien, F.; Pasquier, B.; Bessière, J.-M.; Baudino, S.; Nicolè, F. Structure of the Chemical and Genetic Diversity of the True Lavender over Its Natural Range. Plants 2020, 9, 1640. [Google Scholar] [CrossRef] [PubMed]
- Pignatti, S.; Guarino, R.; La Rosa, M. Flora d’Italia; Edagricole: Milano, Italy, 2018; Volume 3, ISBN 978-88-506-5244-0. [Google Scholar]
- Pignatti, S.; Guarino, R.; La Rosa, M. Flora d’Italia; Edagricole: Milano, Italy, 2019; Volume 4, ISBN 978-88-506-5245-7. [Google Scholar]
- Politi, M.; De Tommasi, N.; Pescitelli, G.; Di Bari, L.; Morelli, I.; Braca, A. Structure and absolute configuration of new diterpenes from Lavandula multifida. J. Nat. Prod. 2002, 65, 1742–1745. [Google Scholar] [CrossRef] [PubMed]
- Johnson, S.R.; Bhat, W.W.; Bibik, J.; Turmo, A.; Hamberger, B.; Evolutionary Mint Genomics, C.; Hamberger, B. A database-driven approach identifies additional diterpene synthase activities in the mint family (Lamiaceae). J. Biol. Chem. 2019, 294, 1349–1362. [Google Scholar] [CrossRef] [Green Version]
- Shafaghat, A.; Salimi, F.; Amani-Hooshyar, V. Phytochemical and antimicrobial activities of Lavandula officinalis leaves and stems against some pathogenic microorganisms. J. Med. Plants Res. 2012, 6, 455–460. [Google Scholar] [CrossRef] [Green Version]
- Park, C.H.; Yeo, H.J.; Park, Y.E.; Kim, Y.J.; Park, C.; Kim, J.K.; Park, S.U. Integrated Analysis of Transcriptome and Metabolome and Evaluation of Antioxidant Activities in Lavandulapubescens. Antioxidants 2021, 10, 1027. [Google Scholar] [CrossRef]
- Ciocarlan, A.; Lupascu, L.; Aricu, A.; Dragalin, I.; Popescu, V.; Geana, E.I.; Ionete, R.E.; Vornicu, N.; Duliu, O.G.; Hristozova, G.; et al. Chemical Composition and Assessment of Antimicrobial Activity of Lavender Essential Oil and Some By-Products. Plants 2021, 10, 1829. [Google Scholar] [CrossRef]
- Hatem, N.A.; Najah, Z.M. Isolation and elucidation of some chemical constituents of Lavandula officinalis. J. Chem. Pharm. Res. 2016, 8, 304–401. [Google Scholar]
- Gimenez-Rota, C.; Loran, S.; Mainar, A.M.; Hernaiz, M.J.; Rota, C. Supercritical Carbon Dioxide Antisolvent Fractionation for the Sustainable Concentration of Lavandula luisieri (Rozeira) Riv.-Mart Antimicrobial and Antioxidant Compounds and Comparison with Its Conventional Extracts. Plants 2019, 8, 455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papanov, G.; Bozov, P.; Malakov, P. Triterpenoids from lavandula spica. Phytochemistry 1992, 31, 1424–1426. [Google Scholar] [CrossRef]
- Elsbaey, M.; Mwakalukwa, R.; Shimizu, K.; Miyamoto, T. Pentacylic triterpenes from Lavandula coronopifolia: Structure related inhibitory activity on α-glucosidase. Nat. Prod. Res. 2021, 35, 1436–1444. [Google Scholar] [CrossRef]
- Falev, D.I.; Ul’yanovskii, N.V.; Ovchinnikov, D.V.; Faleva, A.V.; Kosyakov, D.S. Screening and semi-quantitative determination of pentacyclic triterpenoids in plants by liquid chromatography-tandem mass spectrometry in precursor ion scan mode. Phytochem. Anal. 2021, 32, 252–261. [Google Scholar] [CrossRef] [PubMed]
- Topcu, G.; Ayral, M.N.; Aydin, A.; Goren, A.C.; Chai, H.B.; Pezzuto, J.M. Triterpenoids of the roots of Lavandula stoechas ssp. stoechas. Pharmazie 2001, 56, 892–895. [Google Scholar] [PubMed]
- Barrett, P.N.; Mundt, W.; Kistner, O.; Howard, M.K. Vero cell platform in vaccine production: Moving towards cell culture-based viral vaccines. Expert. Rev. Vaccines 2009, 8, 607–618. [Google Scholar] [CrossRef] [PubMed]
- Osada, N.; Kohara, A.; Yamaji, T.; Hirayama, N.; Kasai, F.; Sekizuka, T.; Kuroda, M.; Hanada, K. The genome landscape of the african green monkey kidney-derived vero cell line. DNA Res. 2014, 21, 673–683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giugliano, R.; Buonocore, C.; Zannella, C.; Chianese, A.; Palma Esposito, F.; Tedesco, P.; De Filippis, A.; Galdiero, M.; Franci, G.; de Pascale, D. Antiviral Activity of the Rhamnolipids Mixture from the Antarctic Bacterium Pseudomonas gessardii M15 against Herpes Simplex Viruses and Coronaviruses. Pharmaceutics 2021, 13, 2121. [Google Scholar] [CrossRef]
- Ansari, M.; Sharififar, F.; Arabzadeh, A.M.; Mehni, F.; Mirtadzadini, M.; Iranmanesh, Z.; Nikpour, N. In vitro evaluation of anti-herpes simplex-1 activity of three standardized medicinal plants from Lamiaceae. Anc. Sci. Life 2014, 34, 33–38. [Google Scholar] [CrossRef]
- Denaro, M.; Smeriglio, A.; Barreca, D.; De Francesco, C.; Occhiuto, C.; Milano, G.; Trombetta, D. Antiviral activity of plants and their isolated bioactive compounds: An update. Phytother. Res. 2020, 34, 742–768. [Google Scholar] [CrossRef]
- Chaughule, R.S.; Barve, R.S. Role of herbal medicines in the treatment of infectious diseases. Vegetos 2023. online ahead of print. [Google Scholar]
- Yucharoen, R.; Chansakaow, S.; Tragoolpua, Y. Inhibitory effect of aromatic herbs, lavender, sage and chamomile against herpes simplex virus infection. Afr. J. Biotechnol. 2011, 10, 15394–15401. [Google Scholar] [CrossRef]
- Nelson, E.O.; Ruiz, G.G.; Kozin, A.F.; Turner, T.C.; Langland, E.V.; Langland, J.O. Resolution of Recurrent Oro-facial Herpes Simplex Using a Topical Botanical Gel: A Case Report. Yale J. Biol. Med. 2020, 93, 277–281. [Google Scholar]
- Jonsdottir, H.R.; Dijkman, R. Coronaviruses and the human airway: A universal system for virus-host interaction studies. Virol. J. 2016, 13, 24. [Google Scholar] [CrossRef] [Green Version]
- Guarner, J. Three Emerging Coronaviruses in Two Decades. Am. J. Clin. Pathol. 2020, 153, 420–421. [Google Scholar] [CrossRef] [PubMed]
- Kaushik, N.K.; Bhartiya, P.; Kaushik, N.; Shin, Y.; Nguyen, L.N.; Park, J.S.; Kim, D.; Choi, E.H. Nitric-oxide enriched plasma-activated water inactivates 229E coronavirus and alters antiviral response genes in human lung host cells. Bioact. Mater. 2023, 19, 569–580. [Google Scholar] [CrossRef] [PubMed]
- Maccioni, R.; Serra, M.; Marongiu, J.; Cottiglia, F.; Maccioni, E.; Bassareo, V.; Morelli, M.; Kasture, S.B.; Acquas, E. Effects of docosanyl ferulate, a constituent of Withania somnifera, on ethanol- and morphine-elicited conditioned place preference and ERK phosphorylation in the accumbens shell of CD1 mice. Psychopharmacology 2022, 239, 795–806. [Google Scholar] [CrossRef] [PubMed]
- Biltekin, S.N.; Karadag, A.E.; Demirci, B.; Demirci, F. ACE2 and LOX Enzyme Inhibitions of Different Lavender Essential Oils and Major Components Linalool and Camphor. ACS Omega 2022, 7, 36561–36566. [Google Scholar] [CrossRef]
- Prall, S.; Bowles, E.J.; Bennett, K.; Cooke, C.G.; Agnew, T.; Steel, A.; Hausser, T. Effects of essential oils on symptoms and course (duration and severity) of viral respiratory infections in humans: A rapid review. Adv. Integr. Med. 2020, 7, 218–221. [Google Scholar] [CrossRef]
- Thormar, H.; Isaacs, C.E.; Brown, H.R.; Barshatzky, M.R.; Pessolano, T. Inactivation of enveloped viruses and killing of cells by fatty acids and monoglycerides. Antimicrob. Agents Chemother. 1987, 31, 27–31. [Google Scholar] [CrossRef] [Green Version]
- Kohn, A.; Gitelman, J.; Inbar, M. Unsaturated free fatty acids inactivate animal enveloped viruses. Arch. Virol. 1980, 66, 301–307. [Google Scholar] [CrossRef]
- Conzelmann, C.; Zou, M.; Gross, R.; Harms, M.; Rocker, A.; Riedel, C.U.; Munch, J.; Muller, J.A. Storage-Dependent Generation of Potent Anti-ZIKV Activity in Human Breast Milk. Viruses 2019, 11, 591. [Google Scholar] [CrossRef] [Green Version]
- Yoon, B.K.; Jackman, J.A.; Valle-Gonzalez, E.R.; Cho, N.J. Antibacterial Free Fatty Acids and Monoglycerides: Biological Activities, Experimental Testing, and Therapeutic Applications. Int. J. Mol. Sci. 2018, 19, 1114. [Google Scholar] [CrossRef] [Green Version]
- Hilmarsson, H.; Kristmundsdottir, T.; Thormar, H. Virucidal activities of medium- and long-chain fatty alcohols, fatty acids and monoglycerides against herpes simplex virus types 1 and 2: Comparison at different pH levels. APMIS 2005, 113, 58–65. [Google Scholar] [CrossRef]
- Ozcelik, B.; Aslan, M.; Orhan, I.; Karaoglu, T. Antibacterial, antifungal, and antiviral activities of the lipophylic extracts of Pistacia vera. Microbiol. Res. 2005, 160, 159–164. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Yang, L.; Wang, H.; Xiong, Y. Recent Advances in Antiviral Activities of Triterpenoids. Pharmaceuticals 2022, 15, 1169. [Google Scholar] [CrossRef] [PubMed]
- Shan, T.; Ye, J.; Jia, J.; Wang, Z.; Jiang, Y.; Wang, Y.; Wang, Y.; Zheng, K.; Ren, Z. Viral UL8 Is Involved in the Antiviral Activity of Oleanolic Acid Against HSV-1 Infection. Front. Microbiol. 2021, 12, 689607. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Jia, J.; Jiang, Y.; Li, F.; Wang, Y.; Song, X.; Qin, S.; Wang, Y.; Zheng, K.; Hu, B.; et al. Oleanolic Acid Derivative AXX-18 Exerts Antiviral Activity by Inhibiting the Expression of HSV-1 Viral Genes UL8 and UL52. Viruses 2022, 14, 1287. [Google Scholar] [CrossRef]
- Kumar, A.; Choudhir, G.; Shukla, S.K.; Sharma, M.; Tyagi, P.; Bhushan, A.; Rathore, M. Identification of phytochemical inhibitors against main protease of COVID-19 using molecular modeling approaches. J. Biomol. Struct. Dyn. 2021, 39, 3760–3770. [Google Scholar] [CrossRef]
- Bag, P.; Chattopadhyay, D.; Mukherjee, H.; Ojha, D.; Mandal, N.; Sarkar, M.C.; Chatterjee, T.; Das, G.; Chakraborti, S. Anti-herpes virus activities of bioactive fraction and isolated pure constituent of Mallotus peltatus: An ethnomedicine from Andaman Islands. Virol. J. 2012, 9, 98. [Google Scholar] [CrossRef] [Green Version]
- Al-Kuraishy, H.M.; Al-Gareeb, A.I.; El-Saber Batiha, G. The possible role of ursolic acid in COVID-19: A real game changer. Clin. Nutr. ESPEN 2022, 47, 414–417. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak n. | RT (min) | Tentative Assignment | Formula | [M-H]− Found (m/z) | Error (ppm) | RDB | MS/MS Fragment Ions (m/z) |
|---|---|---|---|---|---|---|---|
| 1 | 0.672 | Diterpene (e.g., rosmaridiphenol) | C20H28O3 | 315.1961 | −1.5 | 7 | 315.1962; 283.1694; 227.1076 |
| 2 | 0.682 | Diterpene (e.g., rosmanol) | C20H26O5 | 345.1707 | −1.0 | 8 | 345.1695; 330.1474; 300.1360 |
| 3 | 1.140 | dihydroxy-octadecadienoic acid | C18H32O4 | 311.2225 | −0.9 | 3 | 311.2217; 293.2119; 275.1970 |
| 4 | 1.569 | 9-oxooctadeca-10,12-dienoic acid | C18H30O3 | 293.2117 | 0.6 | 4 | 293.2117; 275.2007; 235.1689; 183.1388; 171.1024 |
| 5 | 1.728 | Diterpene derivative (e.g., rosmaquinone B) | C21H26O5 | 357.1712 | 1.3 | 9 | 357.1709; 311.1646; 296.1413; 241.0861 |
| 6 | 1.782 | 9-oxooctadeca-10,12,15-trienoic acid | C18H28O3 | 291.1966 | 1.8 | 5 | 291.1966; 273.1854; 247.2058; 181.1254 |
| 7 | 1.877 | Diterpene (e.g., carnosic acid) | C20H28O4 | 331.1919 | 1.3 | 7 | 331.1912; 303.1952 |
| 8 | 1.993 | trihydroxy urs-12-en-28-oic acid | C30H48O5 | 487.3435 | 1.2 | 7 | 487.3442; 469.3331; 443.3543; 425.3430; 409.3113 |
| 9 | 2.535 | dioxo-hydroxyurs-12-ene-28-oic acid | C30H44O5 | 483.3117 | 0.2 | 7 | 483.3118; 465.3017; 439.3200; 421.3106; 379.2992 |
| 10 | 2.903 | 3-oxo-hydroxyurs-12-en-28-oic acid | C30H46O4 | 469.3330 | 1.4 | 8 | 469.3344; 451.3223; 407.3311 |
| 11 | 3.281 | hydroxyoctadec-12-enoic acid | C18H34O3 | 297.2446 | 3.6 | 2 | 297.2454; 279.2341; 155.1085 |
| 12 | 4.544 | hydroxy-3-oxo-ursa-1,12-dien-28-oic acid | C30H44O4 | 467.3175 | 1.7 | 9 | 467.3184; 449.3077 |
| 13 | 5.166 | linolenic acid | C18H30O2 | 277.2178 | 1.7 | 9 | 277.2176 |
| 14 | 5.438 | oleanolic acid | C30H48O3 | 455.3539 | 1.8 | 7 | 455.3538 |
| 15 | 5.671 | ursolic acid | C30H48O3 | 455.3536 | 1.2 | 7 | 455.3543 |
| 16 | 5.909 | linoleic acid | C18H32O2 | 279.2334 | 1.6 | 3 | 279.2335 |
| 17 | 6.076 | dehydroursolic acid | C30H46O3 | 453.3381 | 1.5 | 8 | 453.3392; 407.3312 |
| 18 | 6.471 | palmitic acid | C16H32O2 | 255.2334 | 1.4 | 1 | 255.2331; 219.8351 |
| 19 | 6.723 | oleic acid | C18H34O2 | 281.2489 | 1.1 | 2 | 281.2491 |
| 20 | 7.889 | docosyl ferulate | C32H54O4 | 501.3962 | 2.5 | 6 | 501.3962; 486.3723; 193.0508; 177.0196; 133.0299 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chianese, A.; Gravina, C.; Morone, M.V.; Ambrosino, A.; Formato, M.; Palma, F.; Foglia, F.; Nastri, B.M.; Zannella, C.; Esposito, A.; et al. Lavandula austroapennina: Assessment of the Antiviral Activity of Lipophilic Extracts from Its Organs. Viruses 2023, 15, 1648. https://doi.org/10.3390/v15081648
Chianese A, Gravina C, Morone MV, Ambrosino A, Formato M, Palma F, Foglia F, Nastri BM, Zannella C, Esposito A, et al. Lavandula austroapennina: Assessment of the Antiviral Activity of Lipophilic Extracts from Its Organs. Viruses. 2023; 15(8):1648. https://doi.org/10.3390/v15081648
Chicago/Turabian StyleChianese, Annalisa, Claudia Gravina, Maria Vittoria Morone, Annalisa Ambrosino, Marialuisa Formato, Francesca Palma, Francesco Foglia, Bianca Maria Nastri, Carla Zannella, Assunta Esposito, and et al. 2023. "Lavandula austroapennina: Assessment of the Antiviral Activity of Lipophilic Extracts from Its Organs" Viruses 15, no. 8: 1648. https://doi.org/10.3390/v15081648