
Genomic Characterization and gE/gI-Deleted Strain Construction of Novel PRV Variants Isolated in Central China
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells, Viruses and Antibodies
2.2. Virus Isolation and Identification
2.3. Multistep Growth Analysis
2.4. Plaque Size Analysis
2.5. Genomic Sequencing
2.6. Phylogenetic Analysis and Sequence Alignment
2.7. Construction of a gE/gI-Deleted Virus
2.8. Experiments in Mice
2.8.1. Pathogenicity Test
2.8.2. Immunization and Challenge Test
2.9. Quantitative PCR (qPCR)
2.10. Histopathological Examinations
2.11. Statistical Analyses
3. Results
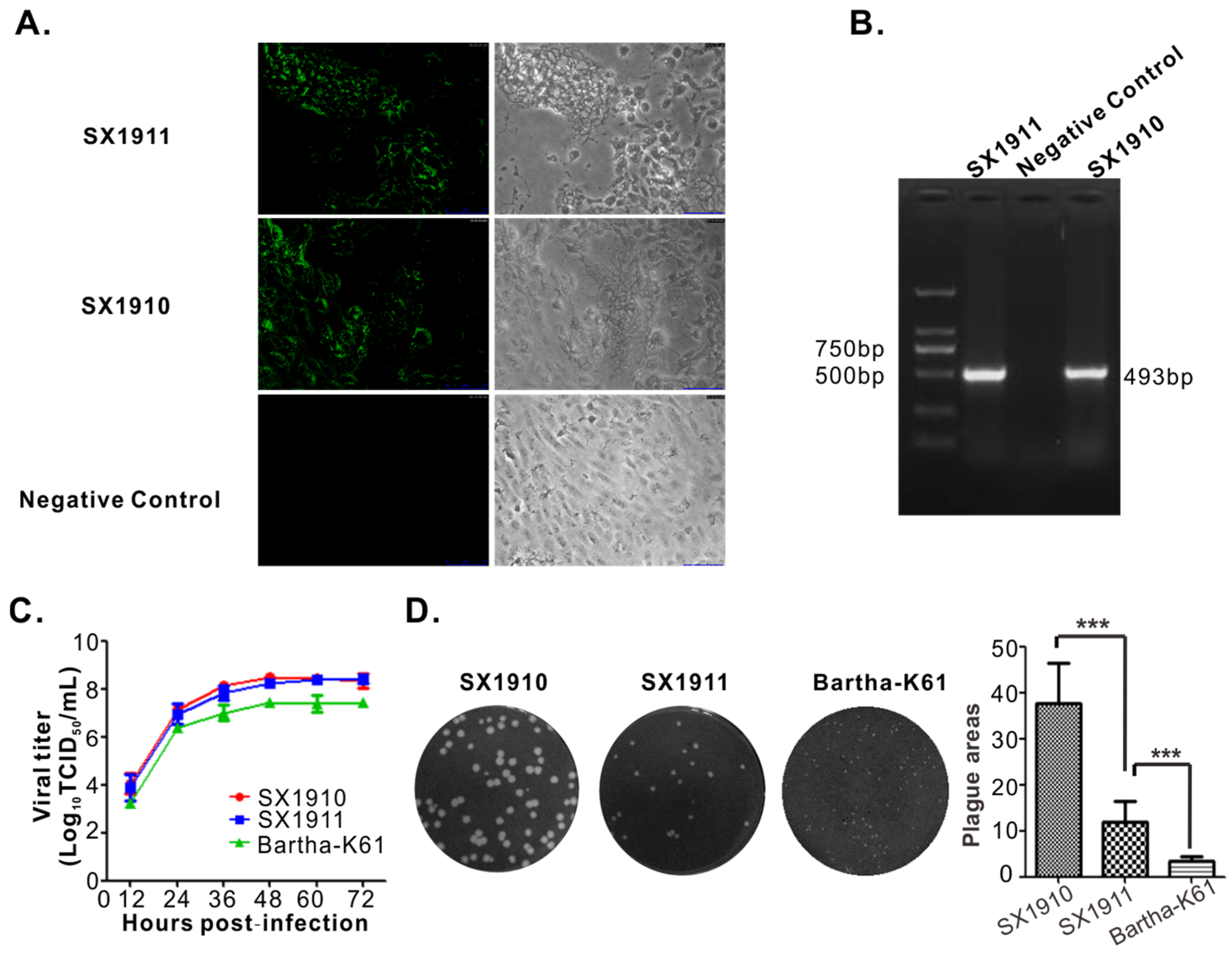
3.1. Isolation and Identification of Virus
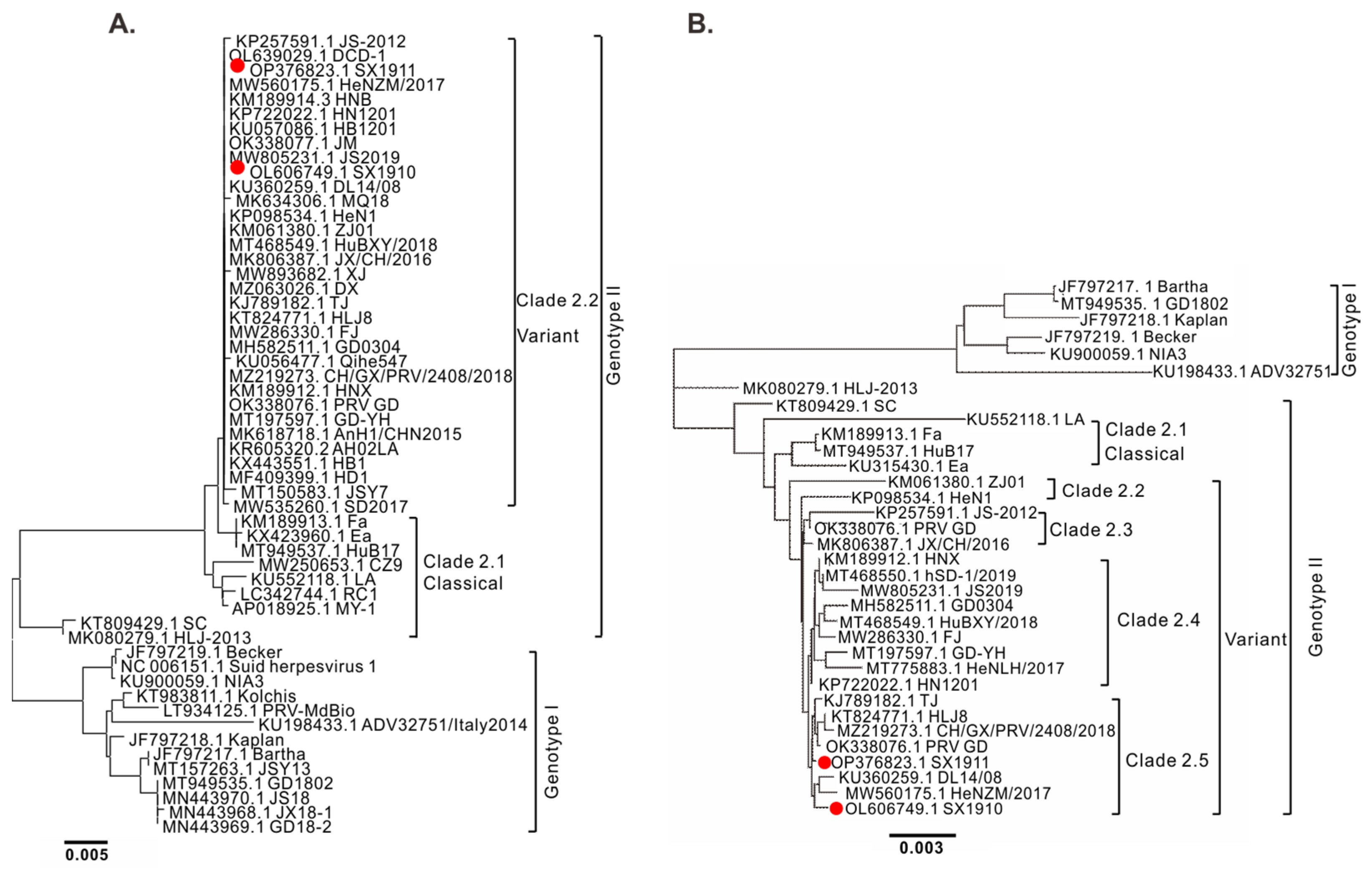
3.2. Genomic Sequencing and Phylogenetic Analysis
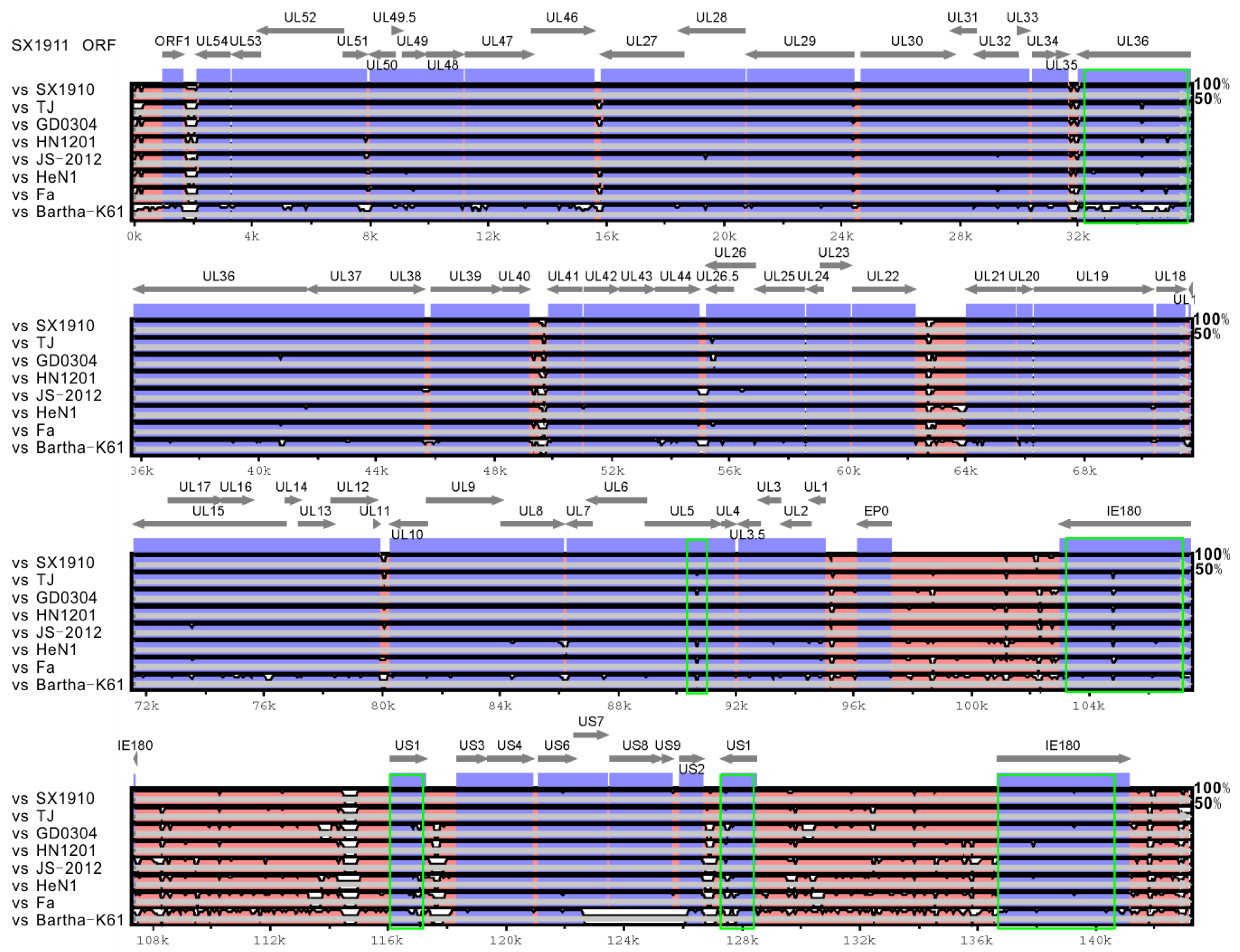
3.3. The Extensive Variations That the Two Isolates Exhibit in the Protein-Coding Sequences UL5, UL36, US1 and IE180
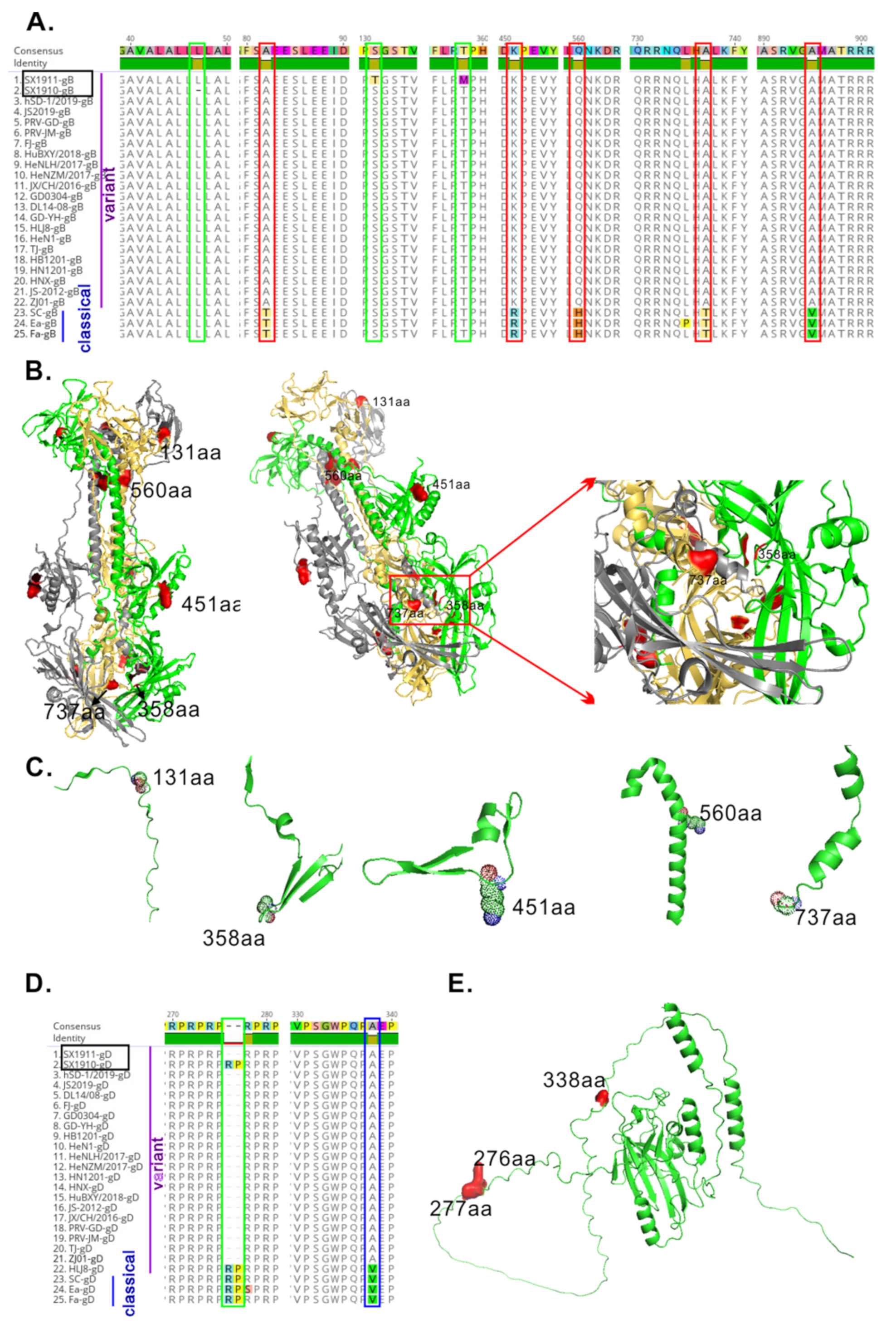
3.4. The Glycoproteins gB and gD of the Two Isolates Have New Amino Acid Mutations
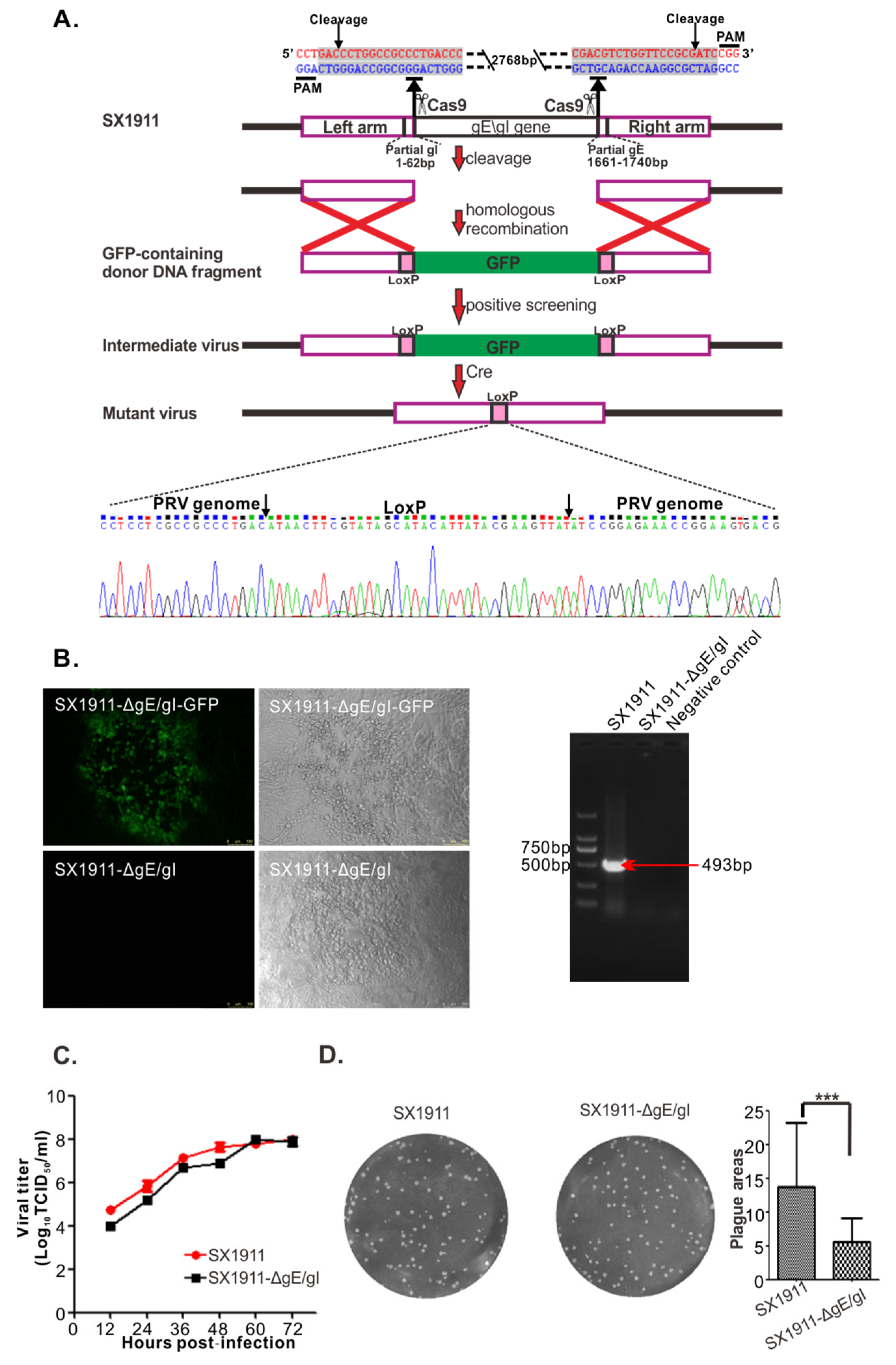
3.5. Construction and Biological Characterization of gE/gI-Deleted Virus of SX1911
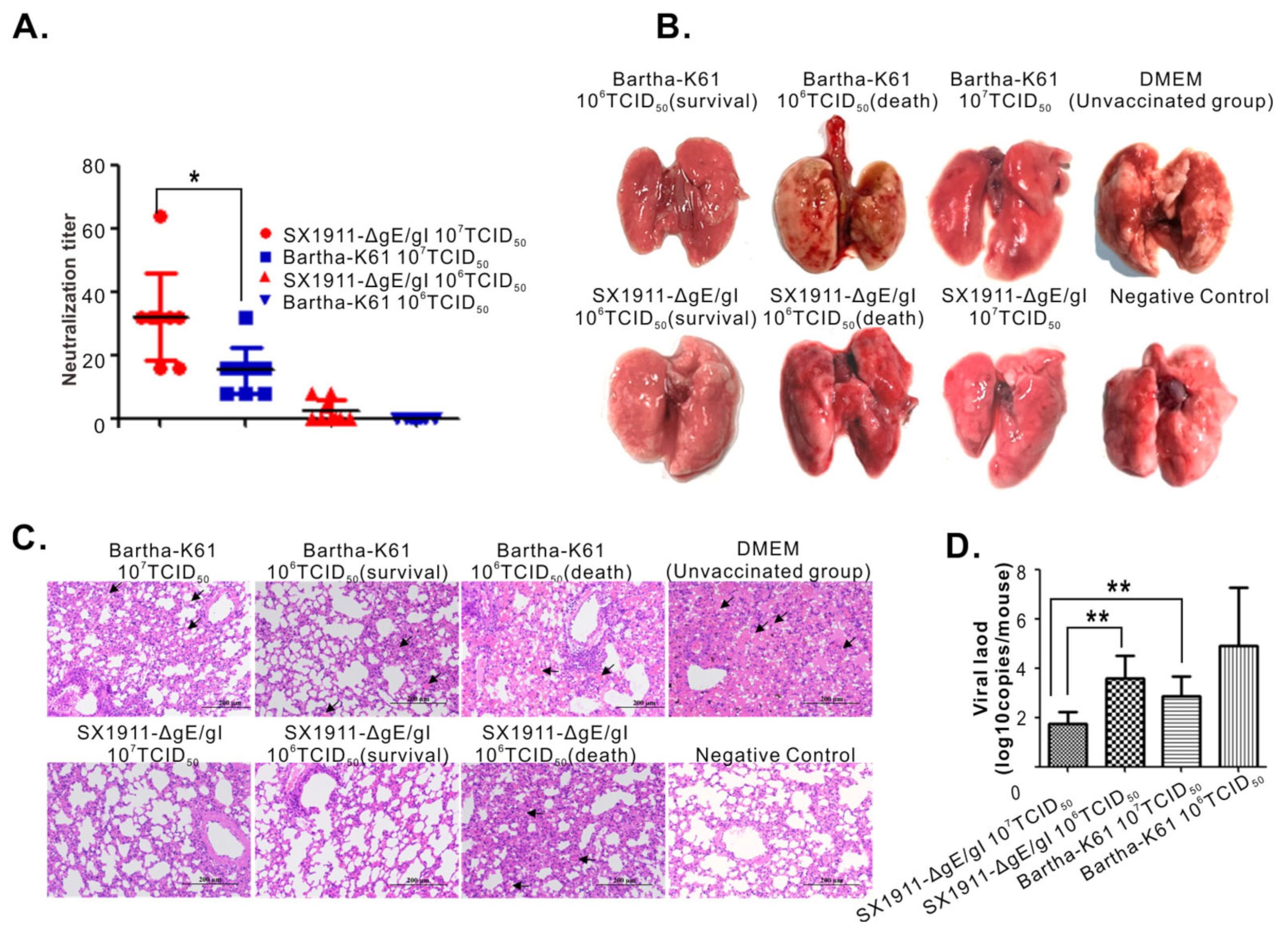
3.6. Bartha-K61 Provides a Comparable Protection Range to SX1911-ΔgE/gI
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mettenleiter, T.C. Aujeszky’s disease (pseudorabies) virus: The virus and molecular pathogenesis—state of the art, June 1999. Vet. Res. 2000, 31, 99–115. [Google Scholar] [CrossRef] [PubMed]
- Pomeranz, L.E.; Reynolds, A.E.; Hengartner, C.J. Molecular biology of pseudorabies virus: Impact on neurovirology and veterinary medicine. Microbiol. Mol. Biol. Rev. MMBR 2005, 69, 462–500. [Google Scholar] [CrossRef] [PubMed]
- Muller, T.; Hahn, E.C.; Tottewitz, F.; Kramer, M.; Klupp, B.G.; Mettenleiter, T.C.; Freuling, C. Pseudorabies virus in wild swine: A global perspective. Arch. Virol. 2011, 156, 1691–1705. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.Y.; Wilson, M.R. A review of pseudorabies (Aujeszky’s disease) in pigs. Can. Vet. J. 1979, 20, 65–69. [Google Scholar]
- Sun, Y.; Luo, Y.; Wang, C.H.; Yuan, J.; Li, N.; Song, K.; Qiu, H.J. Control of swine pseudorabies in China: Opportunities and limitations. Vet. Microbiol. 2016, 183, 119–124. [Google Scholar] [CrossRef]
- Freuling, C.M.; Muller, T.F.; Mettenleiter, T.C. Vaccines against pseudorabies virus (PrV). Vet. Microbiol. 2017, 206, 3–9. [Google Scholar] [CrossRef]
- Mettenleiter, T.C. Aujeszky’s Disease and the Development of the Marker/DIVA Vaccination Concept. Pathogens 2020, 9, 563. [Google Scholar] [CrossRef]
- Laval, K.; Enquist, L.W. The Neuropathic Itch Caused by Pseudorabies Virus. Pathogens 2020, 9, 254. [Google Scholar] [CrossRef]
- Zheng, H.H.; Fu, P.F.; Chen, H.Y.; Wang, Z.Y. Pseudorabies Virus: From Pathogenesis to Prevention Strategies. Viruses 2022, 14, 1638. [Google Scholar] [CrossRef]
- Delva, J.L.; Nauwynck, H.J.; Mettenleiter, T.C.; Favoreel, H.W. The Attenuated Pseudorabies Virus Vaccine Strain Bartha K61: A Brief Review on the Knowledge Gathered during 60 Years of Research. Pathogens 2020, 9, 897. [Google Scholar] [CrossRef]
- Pannett, G.R.; Motha, M.X.; MacDiarmid, S.C. Eradication of Aujeszky’s disease from New Zealand pig herds 1976–1997. Vet. Rec. 1999, 144, 365–369. [Google Scholar] [CrossRef] [PubMed]
- Yang, H. Epidemic status and characteristics of swine pseudorabies [in Chinese]. Swine Indus. Sci. 2016, 33, 38. [Google Scholar]
- An, T.Q.; Peng, J.M.; Tian, Z.J.; Zhao, H.Y.; Li, N.; Liu, Y.M.; Chen, J.Z.; Leng, C.L.; Sun, Y.; Chang, D.; et al. Pseudorabies virus variant in Bartha-K61-vaccinated pigs, China, 2012. Emerg. Infect. Dis. 2013, 19, 1749–1755. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Zhou, Z.; Hu, D.; Zhang, Q.; Han, T.; Li, X.; Gu, X.; Yuan, L.; Zhang, S.; Wang, B.; et al. Pathogenic pseudorabies virus, China, 2012. Emerg. Infect. Dis. 2014, 20, 102–104. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Li, N.; Cong, X.; Wang, C.H.; Du, M.; Li, L.; Zhao, B.; Yuan, J.; Liu, D.D.; Li, S.; et al. Pathogenicity and genomic characterization of a pseudorabies virus variant isolated from Bartha-K61-vaccinated swine population in China. Vet. Microbiol. 2014, 174, 107–115. [Google Scholar] [CrossRef]
- Tong, W.; Liu, F.; Zheng, H.; Liang, C.; Zhou, Y.J.; Jiang, Y.F.; Shan, T.L.; Gao, F.; Li, G.X.; Tong, G.Z. Emergence of a Pseudorabies virus variant with increased virulence to piglets. Vet. Microbiol. 2015, 181, 236–240. [Google Scholar] [CrossRef]
- Yang, Q.Y.; Sun, Z.; Tan, F.F.; Guo, L.H.; Wang, Y.Z.; Wang, J.; Wang, Z.Y.; Wang, L.L.; Li, X.D.; Xiao, Y.; et al. Pathogenicity of a currently circulating Chinese variant pseudorabies virus in pigs. World J. Virol. 2016, 5, 23–30. [Google Scholar] [CrossRef]
- Wang, C.H.; Yuan, J.; Qin, H.Y.; Luo, Y.; Cong, X.; Li, Y.; Chen, J.; Li, S.; Sun, Y.; Qiu, H.J. A novel gE-deleted pseudorabies virus (PRV) provides rapid and complete protection from lethal challenge with the PRV variant emerging in Bartha-K61-vaccinated swine population in China. Vaccine 2014, 32, 3379–3385. [Google Scholar] [CrossRef]
- Gu, Z.; Dong, J.; Wang, J.; Hou, C.; Sun, H.; Yang, W.; Bai, J.; Jiang, P. A novel inactivated gE/gI deleted pseudorabies virus (PRV) vaccine completely protects pigs from an emerged variant PRV challenge. Virus Res. 2015, 195, 57–63. [Google Scholar] [CrossRef]
- Hu, R.M.; Zhou, Q.; Song, W.B.; Sun, E.C.; Zhang, M.M.; He, Q.G.; Chen, H.C.; Wu, B.; Liu, Z.F. Novel pseudorabies virus variant with defects in TK, gE and gI protects growing pigs against lethal challenge. Vaccine 2015, 33, 5733–5740. [Google Scholar] [CrossRef]
- Ren, J.; Wang, H.; Zhou, L.; Ge, X.; Guo, X.; Han, J.; Yang, H. Glycoproteins C and D of PRV Strain HB1201 Contribute Individually to the Escape From Bartha-K61 Vaccine-Induced Immunity. Front. Microbiol. 2020, 11, 323. [Google Scholar] [CrossRef]
- Ye, C.; Zhang, Q.Z.; Tian, Z.J.; Zheng, H.; Zhao, K.; Liu, F.; Guo, J.C.; Tong, W.; Jiang, C.G.; Wang, S.J.; et al. Genomic characterization of emergent pseudorabies virus in China reveals marked sequence divergence: Evidence for the existence of two major genotypes. Virology 2015, 483, 32–43. [Google Scholar] [CrossRef] [PubMed]
- Ye, C.; Guo, J.C.; Gao, J.C.; Wang, T.Y.; Zhao, K.; Chang, X.B.; Wang, Q.; Peng, J.M.; Tian, Z.J.; Cai, X.H.; et al. Genomic analyses reveal that partial sequence of an earlier pseudorabies virus in China is originated from a Bartha-vaccine-like strain. Virology 2016, 491, 56–63. [Google Scholar] [CrossRef] [PubMed]
- Dong, J.; Gu, Z.; Jin, L.; Lv, L.; Wang, J.; Sun, T.; Bai, J.; Sun, H.; Wang, X.; Jiang, P. Polymorphisms affecting the gE and gI proteins partly contribute to the virulence of a newly-emergent highly virulent Chinese pseudorabies virus. Virology 2018, 519, 42–52. [Google Scholar] [CrossRef]
- Zhai, X.; Zhao, W.; Li, K.; Zhang, C.; Wang, C.; Su, S.; Zhou, J.; Lei, J.; Xing, G.; Sun, H.; et al. Genome Characteristics and Evolution of Pseudorabies Virus Strains in Eastern China from 2017 to 2019. Virol. Sin. 2019, 34, 601–609. [Google Scholar] [CrossRef]
- Liu, J.; Chen, C.; Li, X. Novel Chinese pseudorabies virus variants undergo extensive recombination and rapid interspecies transmission. Transbound. Emerg. Dis. 2020, 67, 2274–2276. [Google Scholar] [CrossRef] [PubMed]
- Hu, R.; Wang, L.; Liu, Q.; Hua, L.; Huang, X.; Zhang, Y.; Fan, J.; Chen, H.; Song, W.; Liang, W.; et al. Whole-Genome Sequence Analysis of Pseudorabies Virus Clinical Isolates from Pigs in China between 2012 and 2017 in China. Viruses 2021, 13, 1322. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.; Shao, Y.; Tan, C.; Shen, Y.; Zhang, X.; Xiao, J.; Wu, Y.; He, L.; Shao, G.; Han, M.; et al. Commercial vaccine against pseudorabies virus: A hidden health risk for dogs. Vet. Microbiol. 2019, 233, 102–112. [Google Scholar] [CrossRef]
- Fan, S.; Yuan, H.; Liu, L.; Li, H.; Wang, S.; Zhao, W.; Wu, Y.; Wang, P.; Hu, Y.; Han, J.; et al. Pseudorabies virus encephalitis in humans: A case series study. J. Neurovirol. 2020, 26, 556–564. [Google Scholar] [CrossRef]
- Ou, J.; Cai, S.; Zheng, F.; Lu, G.; Zhang, G. Human pseudorabies virus infection: A new threat in China. J. Infect. 2020, 80, 578–606. [Google Scholar] [CrossRef]
- Tu, L.; Lian, J.; Pang, Y.; Liu, C.; Cui, S.; Lin, W. Retrospective detection and phylogenetic analysis of pseudorabies virus in dogs in China. Arch. Virol. 2021, 166, 91–100. [Google Scholar] [CrossRef] [PubMed]
- Reed, L.J.; Muench, H. A simple method of estimating fifty per cent endpoints. Am. J. Epidemiol. 1938, 27, 493–497. [Google Scholar] [CrossRef]
- Szpara, M.L.; Tafuri, Y.R.; Enquist, L.W. Preparation of viral DNA from nucleocapsids. J. Vis. Exp. JoVE 2011, 54, e3151. [Google Scholar]
- Hirt, B. Selective extraction of polyoma DNA from infected mouse cell cultures. J. Mol. Biol. 1967, 26, 365–369. [Google Scholar] [CrossRef] [PubMed]
- Tan, L.; Yao, J.; Yang, Y.; Luo, W.; Yuan, X.; Yang, L.; Wang, A. Current Status and Challenge of Pseudorabies Virus Infection in China. Virol. Sin. 2021, 36, 588–607. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.H.; Bai, Y.L.; Xu, T.; Zheng, L.L.; Li, X.S.; Chen, H.Y.; Wang, Z.Y. Isolation and Phylogenetic Analysis of Reemerging Pseudorabies Virus Within Pig Populations in Central China During 2012 to 2019. Front. Vet. Sci. 2021, 8, 764982. [Google Scholar] [CrossRef]
- Bo, Z.; Li, X. A Review of Pseudorabies Virus Variants: Genomics, Vaccination, Transmission, and Zoonotic Potential. Viruses 2022, 14, 1003. [Google Scholar] [CrossRef]
- Liu, Q.; Kuang, Y.; Li, Y.; Guo, H.; Zhou, C.; Guo, S.; Tan, C.; Wu, B.; Chen, H.; Wang, X. The Epidemiology and Variation in Pseudorabies Virus: A Continuing Challenge to Pigs and Humans. Viruses 2022, 14, 1463. [Google Scholar] [CrossRef]
- Liu, H.; Shi, Z.; Liu, C.; Wang, P.; Wang, M.; Wang, S.; Liu, Z.; Wei, L.; Sun, Z.; He, X.; et al. Implication of the Identification of an Earlier Pseudorabies Virus (PRV) Strain HLJ-2013 to the Evolution of Chinese PRVs. Front. Microbiol. 2020, 11, 612474. [Google Scholar] [CrossRef]
- Rice, S.A.; Davido, D.J. HSV-1 ICP22: Hijacking host nuclear functions to enhance viral infection. Future Microbiol. 2013, 8, 311–321. [Google Scholar] [CrossRef]
- Takacs, I.F.; Tombacz, D.; Berta, B.; Prazsak, I.; Poka, N.; Boldogkoi, Z. The ICP22 protein selectively modifies the transcription of different kinetic classes of pseudorabies virus genes. BMC Mol. Biol. 2013, 14, 2. [Google Scholar] [CrossRef] [PubMed]
- Zaichick, S.V.; Bohannon, K.P.; Hughes, A.; Sollars, P.J.; Pickard, G.E.; Smith, G.A. The herpesvirus VP1/2 protein is an effector of dynein-mediated capsid transport and neuroinvasion. Cell Host Microbe 2013, 13, 193–203. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Liu, S.; Jiang, H.; Deng, H.; Dong, C.; Shen, W.; Chen, H.; Gao, C.; Xiao, S.; Liu, Z.F.; et al. G(2)-quadruplex in the 3’UTR of IE180 regulates Pseudorabies virus replication by enhancing gene expression. RNA Biol. 2020, 17, 816–827. [Google Scholar] [CrossRef] [PubMed]
- Brandt, C.R.; Kolb, A.W.; Shah, D.D.; Pumfery, A.M.; Kintner, R.L.; Jaehnig, E.; Van Gompel, J.J. Multiple determinants contribute to the virulence of HSV ocular and CNS infection and identification of serine 34 of the US1 gene as an ocular disease determinant. Investig. Ophthalmol. Vis. Sci. 2003, 44, 2657–2668. [Google Scholar] [CrossRef]
- O’Toole, J.M.; Aubert, M.; Kotsakis, A.; Blaho, J.A. Mutation of the protein tyrosine kinase consensus site in the herpes simplex virus 1 alpha22 gene alters ICP22 posttranslational modification. Virology 2003, 305, 153–167. [Google Scholar] [CrossRef]
- Mostafa, H.H.; Davido, D.J. Herpes simplex virus 1 ICP22 but not US 1.5 is required for efficient acute replication in mice and VICE domain formation. J. Virol. 2013, 87, 13510–13519. [Google Scholar] [CrossRef]
- Wu, Y.; Tan, S.; He, Q.; Wang, M.; Chen, S.; Jia, R.; Yang, Q.; Zhu, D.; Liu, M.; Zhao, X.; et al. Deletion of Double Copies of the US1 Gene Reduces the Infectivity of Recombinant Duck Plague Virus In Vitro and In Vivo. Microbiol. Spectr. 2022, 10, e0114022. [Google Scholar] [CrossRef]
- Yu, Z.Q.; Tong, W.; Zheng, H.; Li, L.W.; Li, G.X.; Gao, F.; Wang, T.; Liang, C.; Ye, C.; Wu, J.Q.; et al. Variations in glycoprotein B contribute to immunogenic difference between PRV variant JS-2012 and Bartha-K61. Vet. Microbiol. 2017, 208, 97–105. [Google Scholar] [CrossRef]
- Zhang, C.; Liu, Y.; Chen, S.; Qiao, Y.; Guo, M.; Zheng, Y.; Xu, M.; Wang, Z.; Hou, J.; Wang, J. A gD&gC-substituted pseudorabies virus vaccine strain provides complete clinical protection and is helpful to prevent virus shedding against challenge by a Chinese pseudorabies variant. BMC Vet. Res. 2019, 15, 2. [Google Scholar]
- Zhou, M.; Wu, X.; Jiang, D.; Sui, C.; Chen, L.; Cong, X.; Xin, X.; Wang, G.; Li, Y.; Tian, F.; et al. Characterization of a moderately pathogenic pseudorabies virus variant isolated in China, 2014. Infect. Genet. Evol. 2019, 68, 161–171. [Google Scholar] [CrossRef]
- Wang, J.; Cui, X.; Wang, X.; Wang, W.; Gao, S.; Liu, X.; Kai, Y.; Chen, C. Efficacy of the Bartha-K61 vaccine and a gE−/gI−/TK− prototype vaccine against variant porcine pseudorabies virus (vPRV) in piglets with sublethal challenge of vPRV. Res. Vet. Sci. 2020, 128, 16–23. [Google Scholar] [CrossRef] [PubMed]
- Cong, X.; Lei, J.L.; Xia, S.L.; Wang, Y.M.; Li, Y.; Li, S.; Luo, Y.; Sun, Y.; Qiu, H.J. Pathogenicity and immunogenicity of a gE/gI/TK gene-deleted pseudorabies virus variant in susceptible animals. Vet. Microbiol. 2016, 182, 170–177. [Google Scholar] [CrossRef] [PubMed]
- Tong, W.; Li, G.; Liang, C.; Liu, F.; Tian, Q.; Cao, Y.; Li, L.; Zheng, X.; Zheng, H.; Tong, G. A live, attenuated pseudorabies virus strain JS-2012 deleted for gE/gI protects against both classical and emerging strains. Antivir. Res. 2016, 130, 110–117. [Google Scholar] [CrossRef] [PubMed]
- Dong, J.; Bai, J.; Sun, T.; Gu, Z.; Wang, J.; Sun, H.; Jiang, P. Comparative pathogenicity and immunogenicity of triple and double gene-deletion pseudorabies virus vaccine candidates. Res. Vet. Sci. 2017, 115, 17–23. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Symptoms | Clinical Scores a |
|---|---|
| Normal | 0 |
| Ruffled hair | 1 |
| Depression | 1 |
| Anorexia | 1 |
| Moderately labored breathing | 1 |
| Urgent breathing | 2 |
| Itching | 2 |
| Skin biting | 3 |
| Paralysis | 3 |
| Death | 3 |
| Total scores | 17 |
| Groups | Amounts | Doses (TCID50/mL) | Clinical Signs a | Mortality (Mean Days to Death) | LD50 |
|---|---|---|---|---|---|
| SX1910 | 10 | 106 | +++ | 10/10 (2.98) | 103.84 |
| 10 | 105 | +++ | 8/10 (3.51) | ||
| 10 | 104 | ++ | 6/10 (4.08) | ||
| 10 | 103 | + | 2/10 (4.25) | ||
| SX1911 | 10 | 106 | +++ | 10/10 (3.25) | 104.42 |
| 10 | 105 | +++ | 7/10 (3.99) | ||
| 10 | 104 | ++ | 3/10 (4.16) | ||
| 10 | 103 | + | 1/10 (4.25) | ||
| SX1911-ΔgE/gI | 10 | 106 | ++ | 5/10 (4.07) | 106.00 |
| 10 | 105 | + | 2/10 (6.68) | ||
| 10 | 104 | / | 0/10 | ||
| 10 | 103 | / | 0/10 | ||
| DMEM | 10 | 0.1 mL | / | 0/10 |
| Protein Name a | Amino Acid Residues Found in PRV SX1911, which Differ from PRV SX1910 b |
|---|---|
| gK (UL53) | F124L |
| gN (UL49.5) | W42R |
| gB (UL27) | 45 (+L), S131T, T358M |
| ICP18.5 (UL28) | G255A, 256 (+G) |
| ICP8 (UL29) | M178T |
| VP1/2 (UL36) | E404G, L536R, S1464P |
| UL37 | D844G |
| RR1 (UL39) | G589V |
| UL43 | G206D |
| Scaffold (UL26.5) | V269P |
| VP24 (UL26) | V515M |
| OBP (UL9) | L565, W723R |
| UL8 | V5A, V293A, A532E |
| UL5 | 74–79 (PGGPAG > Δ) |
| ICP0 (EP0) | V345A |
| ICP4 (IE180) | Q98Δ, 869–880 (STKSSSSTKSSS > Δ), 448 (+S) |
| ICP22 (US1) | 352–371(EEEEDEEEEDEEEEDEEED > Δ) |
| gD (US6) | 280–281 (RP > Δ) |
| gE (US8) | V386A, G510S |
| Groups | Amounts | Immunization Doses (TCID50/mL) | Clinical Signs a | Mortality (Mean Days to Death) |
|---|---|---|---|---|
| SX1911-ΔgE/gI | 9 | 106 | + | 3/9 (4.00) |
| SX1911-ΔgE/gI | 9 | 107 | / | 0/9 |
| Bartha-K61 | 9 | 106 | + | 3/9 (3.66) |
| Bartha-K61 | 9 | 107 | + | 0/9 |
| DMEM | 9 | 0.2 mL DMEM | +++ | 9/9 (4.44) |
| Negative control | 9 | 0.2 mL DMEM | / | 0/9 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ren, J.; Tan, S.; Chen, X.; Yao, J.; Niu, Z.; Wang, Y.; Ma, L.; Gao, X.; Niu, S.; Liang, L.; et al. Genomic Characterization and gE/gI-Deleted Strain Construction of Novel PRV Variants Isolated in Central China. Viruses 2023, 15, 1237. https://doi.org/10.3390/v15061237
Ren J, Tan S, Chen X, Yao J, Niu Z, Wang Y, Ma L, Gao X, Niu S, Liang L, et al. Genomic Characterization and gE/gI-Deleted Strain Construction of Novel PRV Variants Isolated in Central China. Viruses. 2023; 15(6):1237. https://doi.org/10.3390/v15061237
Chicago/Turabian StyleRen, Jianle, Shanshan Tan, Xinxin Chen, Jiying Yao, Zhihong Niu, Ying Wang, Lei Ma, Xiaolong Gao, Sheng Niu, Libin Liang, and et al. 2023. "Genomic Characterization and gE/gI-Deleted Strain Construction of Novel PRV Variants Isolated in Central China" Viruses 15, no. 6: 1237. https://doi.org/10.3390/v15061237