
Polypyrimidine-Tract-Binding Protein Isoforms Differentially Regulate the Hepatitis C Virus Internal Ribosome Entry Site
 ,
,  ,
,  , , and
, , and 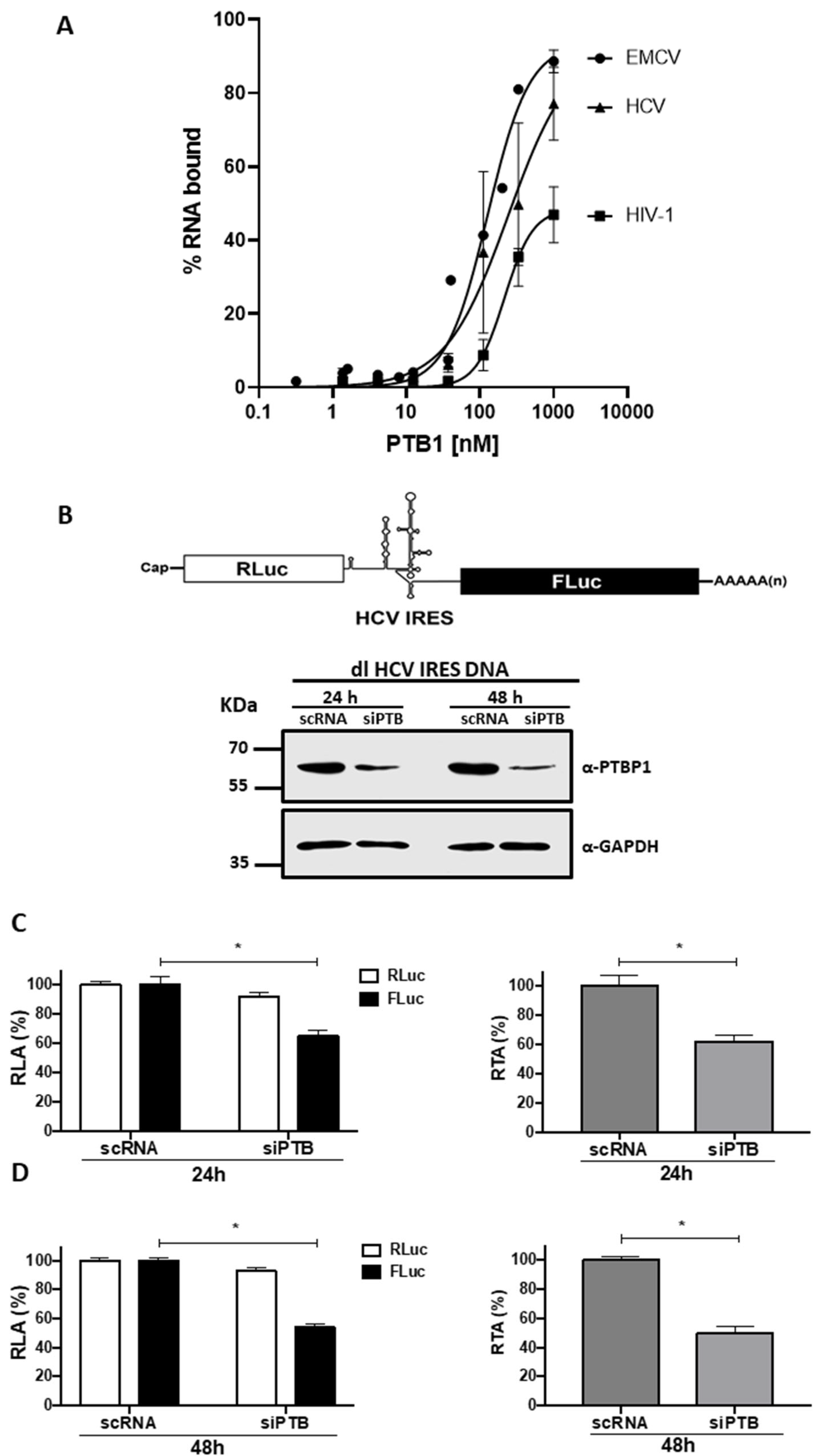{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Material and Methods
3. Results
3.1. Knockdown of Endogenous PTB Negatively Impacts HCV IRES Activity in HuH-7 Cells
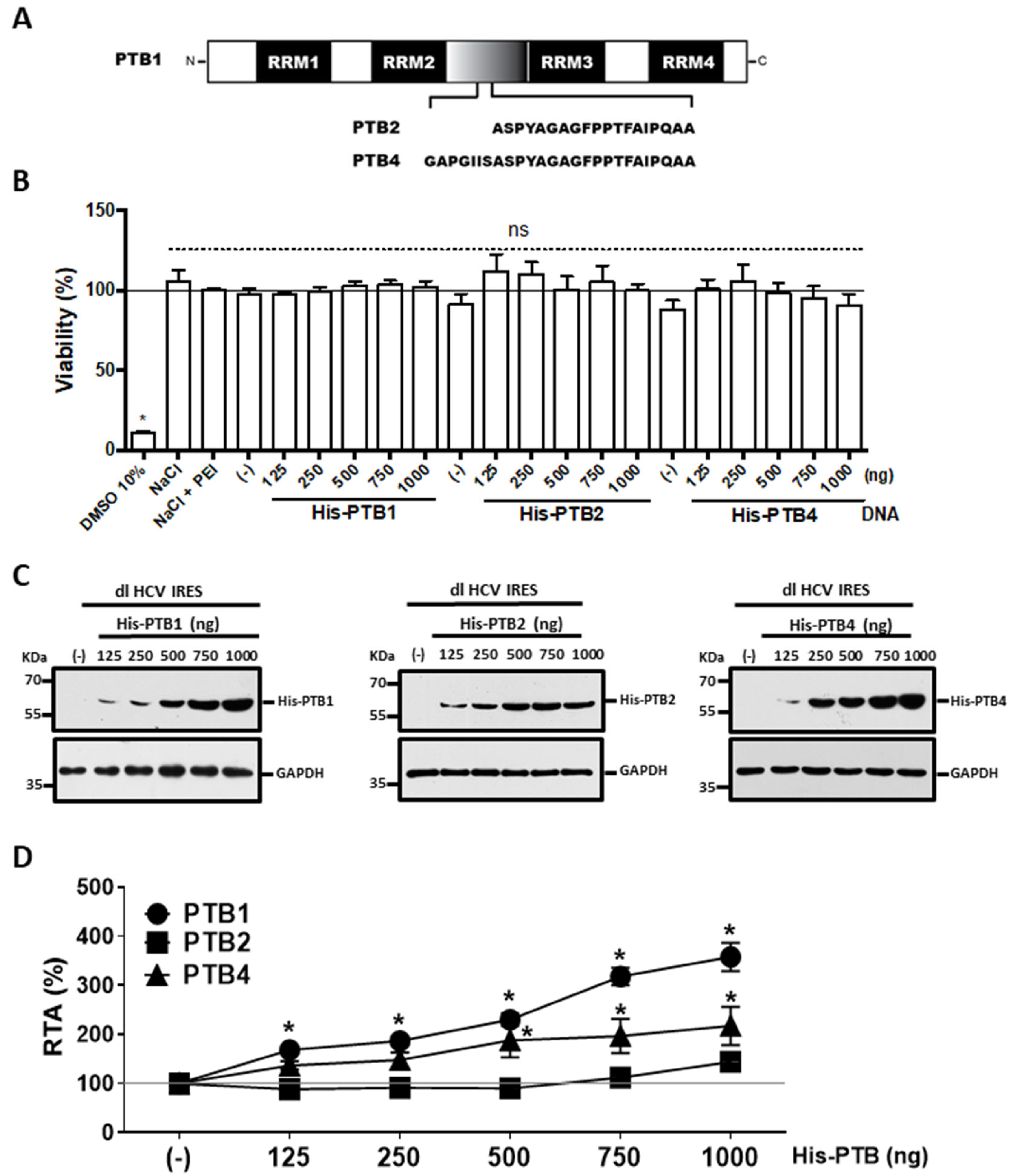
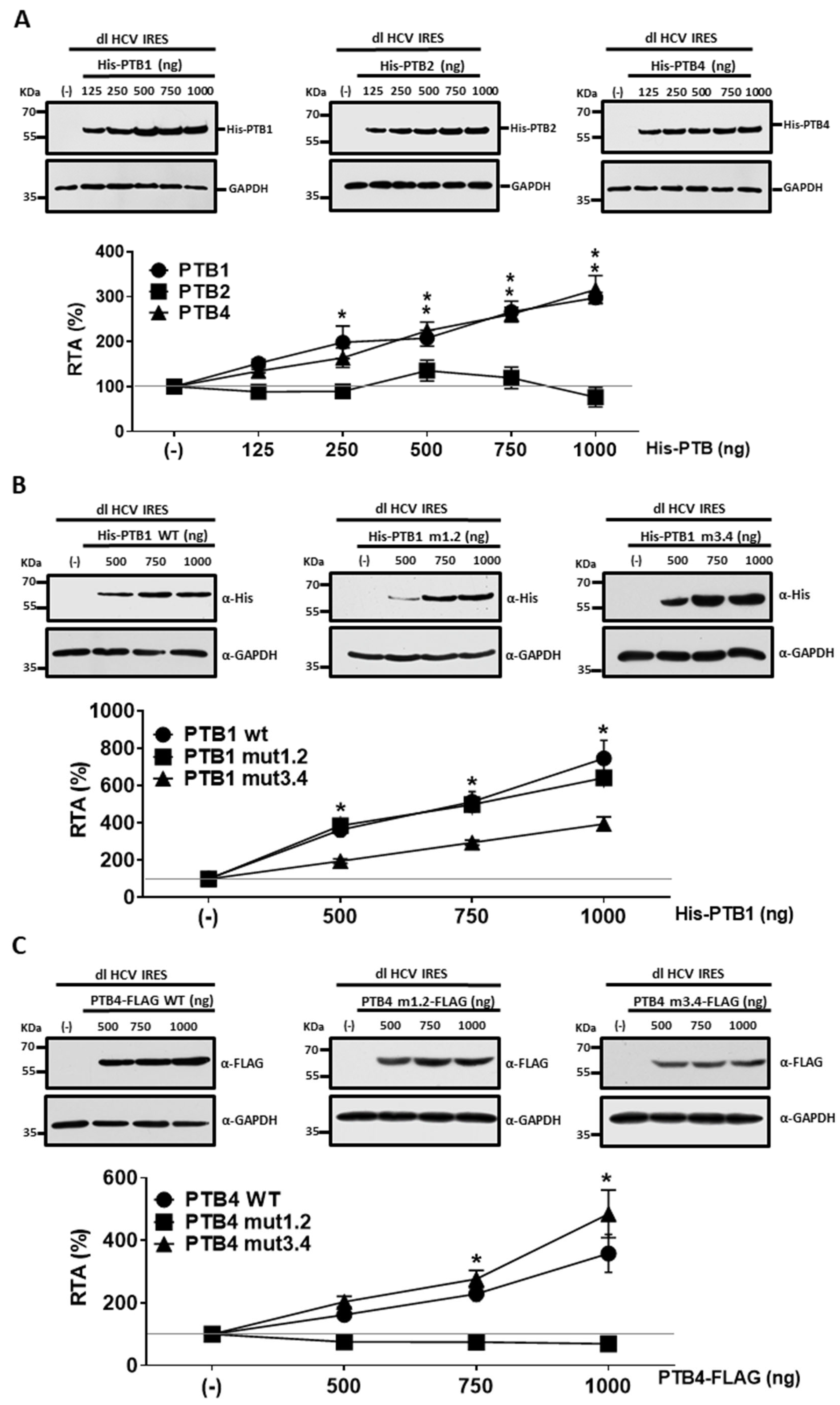
3.2. Overexpression of PTB1 and PTB4, but Not PTB2, Promotes HCV IRES Activity
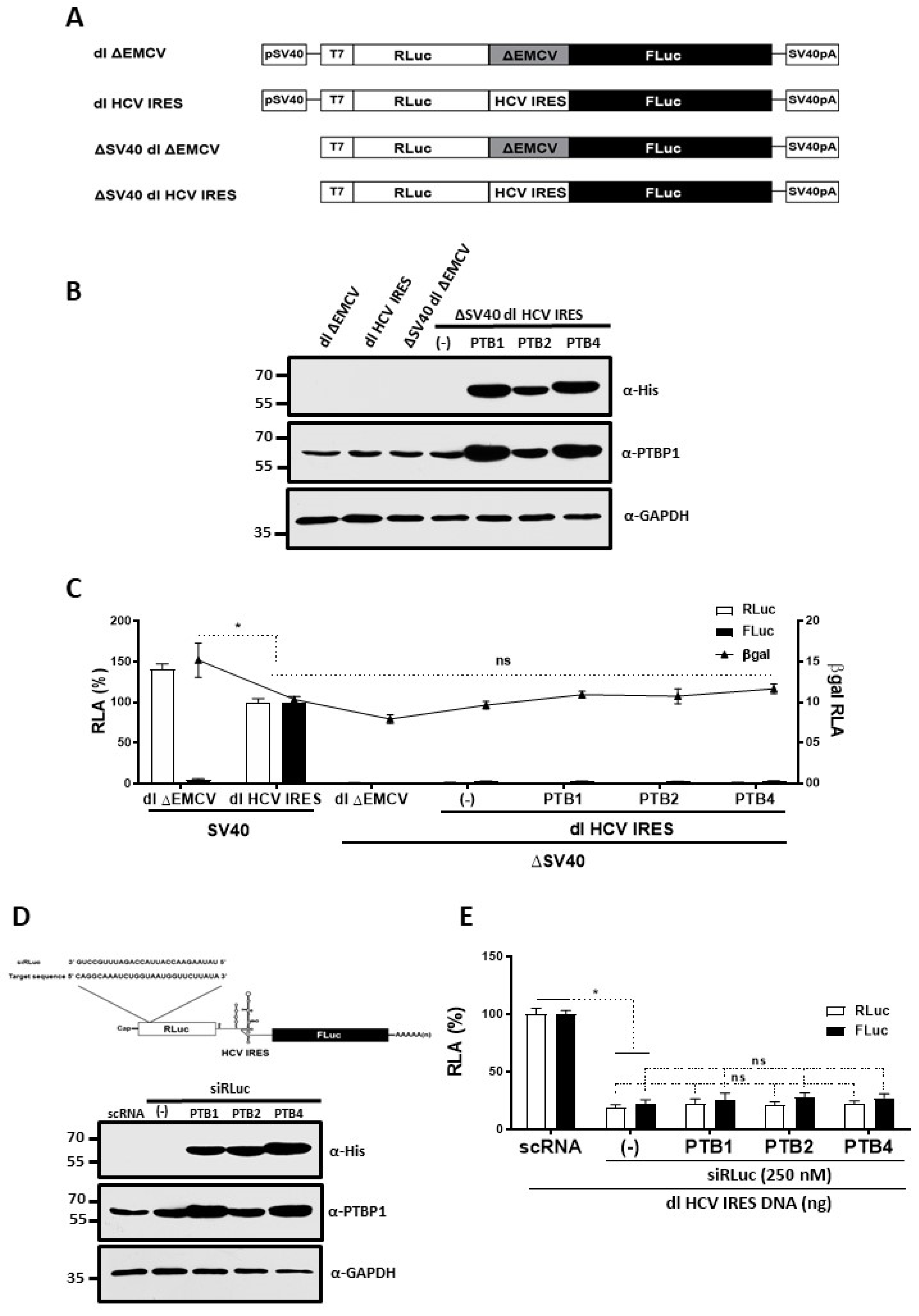
3.3. Overexpression of PTB Isoforms Does Not Induce Alternative Splicing nor Cryptic Promoter Activity from the dl HCV IRES Reporter
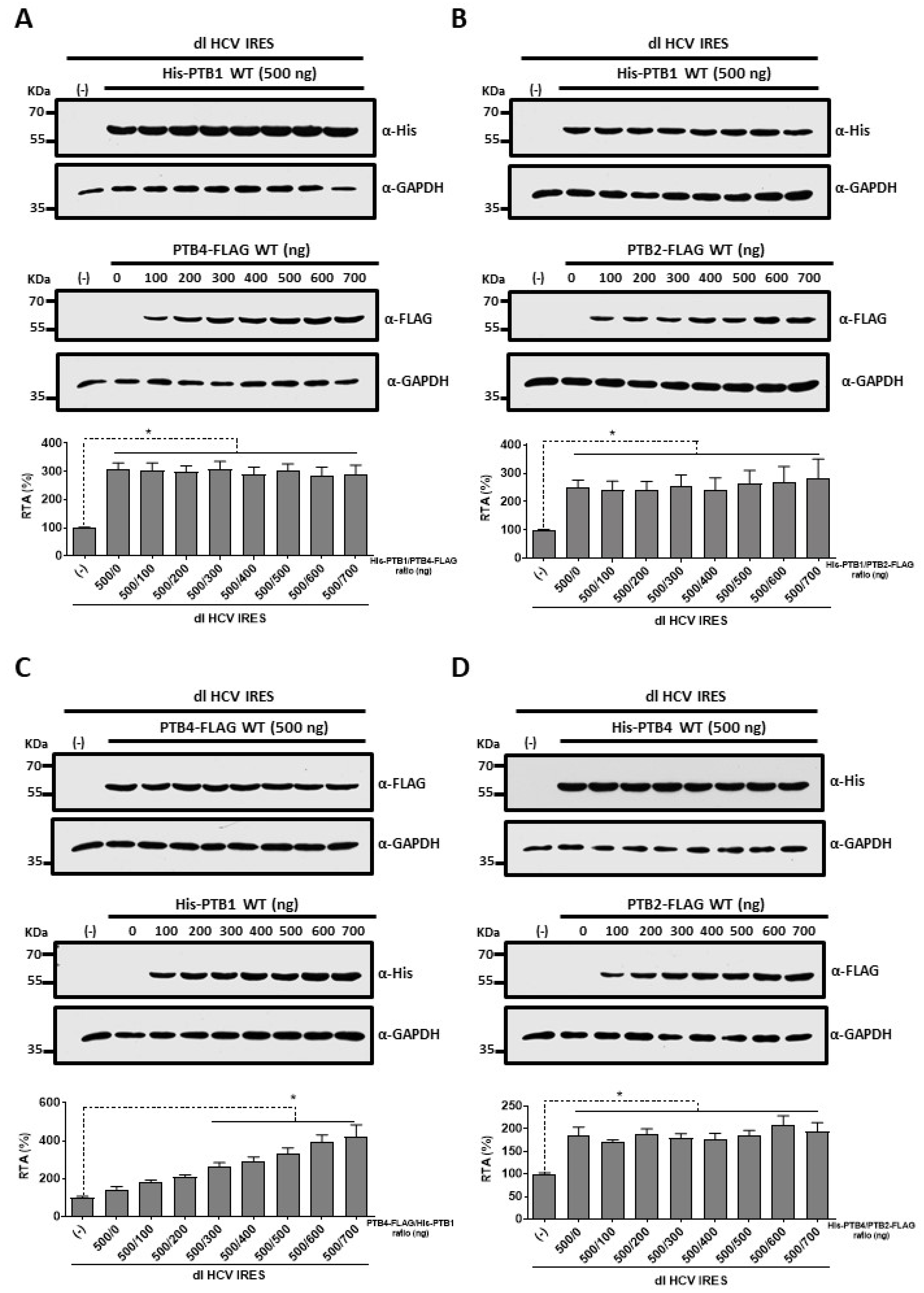
3.4. PTB1 and PTB4 Hierarchy in Promoting HCV IRES-Mediated Initiation in HuH-7 Cells
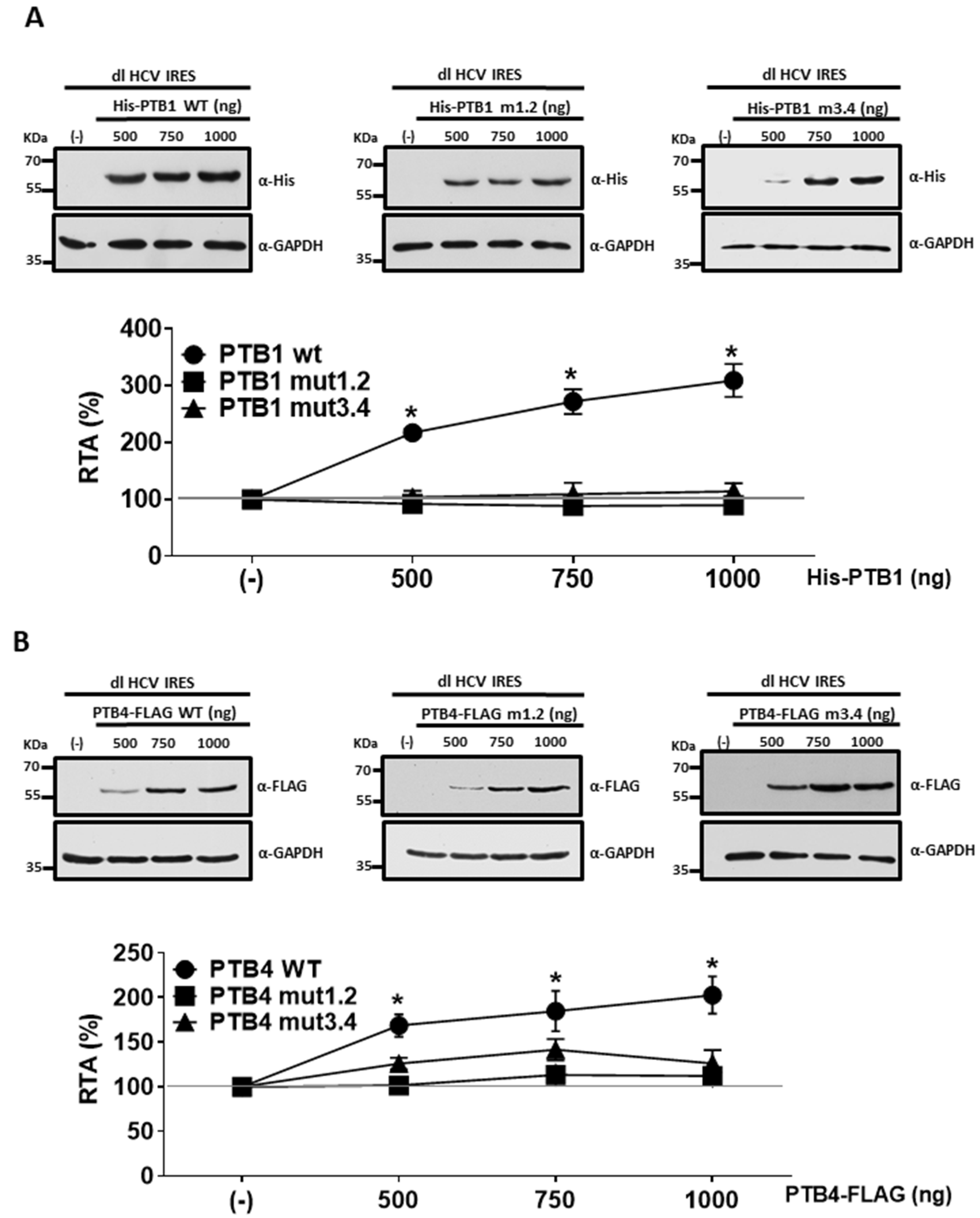
3.5. PTBs RRM1/RRM2 and RRM3/RRM4 Are Required for HCV IRES Stimulation in HuH-7 Cells
3.6. Impacts of PTB1 and PTB4 RRM1/RRM2 and RRM3/RRM4 Mutations on HCV IRES Activity in HEK293T Cells
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Reynolds, J.E.; Kaminski, A.; Kettinen, H.J.; Grace, K.; Clarke, B.E.; Carroll, A.R.; Rowlands, D.J.; Jackson, R.J. Unique features of internal initiation of hepatitis C virus RNA translation. EMBO J. 1995, 14, 6010–6020. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Sarnow, P.; Siddiqui, A. Translation of human hepatitis C virus RNA in cultured cells is mediated by an internal ribosome-binding mechanism. J. Virol. 1993, 67, 3338–3344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsukiyama-Kohara, K.; Iizuka, N.; Kohara, M.; Nomoto, A. Internal ribosome entry site within hepatitis C virus RNA. J. Virol. 1992, 66, 1476–1483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pestova, T.V.; Shatsky, I.N.; Fletcher, S.P.; Jackson, R.J.; Hellen, C.U. A prokaryotic-like mode of cytoplasmic eukaryotic ribosome binding to the initiation codon during internal translation initiation of hepatitis C and classical swine fever virus RNAs. Genes Dev. 1998, 12, 67–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reynolds, J.E.; Kaminski, A.; Carroll, A.R.; Clarke, B.E.; Rowlands, D.J.; Jackson, R.J. Internal initiation of translation of hepatitis C virus RNA: The ribosome entry site is at the authentic initiation codon. RNA 1996, 2, 867–878. [Google Scholar]
- Ali, N.; Siddiqui, A. The La antigen binds 5′ noncoding region of the hepatitis C virus RNA in the context of the initiator AUG codon and stimulates internal ribosome entry site-mediated translation. Proc. Natl. Acad. Sci. USA 1997, 94, 2249–2254. [Google Scholar] [CrossRef] [Green Version]
- Ali, N.; Pruijn, G.J.; Kenan, D.J.; Keene, J.D.; Siddiqui, A. Human La antigen is required for the hepatitis C virus internal ribosome entry site-mediated translation. J. Biol. Chem. 2000, 275, 27531–27540. [Google Scholar] [CrossRef] [Green Version]
- Hwang, B.; Lim, J.H.; Hahm, B.; Jang, S.K.; Lee, S.W. hnRNP L is required for the translation mediated by HCV IRES. Biochem. Biophys. Res. Commun. 2009, 378, 584–588. [Google Scholar] [CrossRef]
- Paek, K.Y.; Kim, C.S.; Park, S.M.; Kim, J.H.; Jang, S.K. RNA-binding protein hnRNP D modulates internal ribosome entry site-dependent translation of hepatitis C virus RNA. J. Virol. 2008, 82, 12082–12093. [Google Scholar] [CrossRef] [Green Version]
- Korf, M.; Jarczak, D.; Beger, C.; Manns, M.P.; Kruger, M. Inhibition of hepatitis C virus translation and subgenomic replication by siRNAs directed against highly conserved HCV sequence and cellular HCV cofactors. J. Hepatol. 2005, 43, 225–234. [Google Scholar] [CrossRef]
- Kruger, M.; Beger, C.; Welch, P.J.; Barber, J.R.; Manns, M.P.; Wong-Staal, F. Involvement of proteasome alpha-subunit PSMA7 in hepatitis C virus internal ribosome entry site-mediated translation. Mol. Cell. Biol. 2001, 21, 8357–8364. [Google Scholar] [CrossRef]
- Kim, J.H.; Paek, K.Y.; Ha, S.H.; Cho, S.; Choi, K.; Kim, C.S.; Ryu, S.H.; Jang, S.K. A cellular RNA-binding protein enhances internal ribosomal entry site-dependent translation through an interaction downstream of the hepatitis C virus polyprotein initiation codon. Mol. Cell. Biol. 2004, 24, 7878–7890. [Google Scholar] [CrossRef] [Green Version]
- Park, S.M.; Paek, K.Y.; Hong, K.Y.; Jang, C.J.; Cho, S.; Park, J.H.; Kim, J.H.; Jan, E.; Jang, S.K. Translation-competent 48S complex formation on HCV IRES requires the RNA-binding protein NSAP1. Nucleic Acids Res. 2011, 39, 7791–7802. [Google Scholar] [CrossRef] [Green Version]
- Scheller, N.; Mina, L.B.; Galao, R.P.; Chari, A.; Gimenez-Barcons, M.; Noueiry, A.; Fischer, U.; Meyerhans, A.; Diez, J. Translation and replication of hepatitis C virus genomic RNA depends on ancient cellular proteins that control mRNA fates. Proc. Natl. Acad. Sci. USA 2009, 106, 13517–13522. [Google Scholar] [CrossRef] [Green Version]
- Izumi, R.E.; Valdez, B.; Banerjee, R.; Srivastava, M.; Dasgupta, A. Nucleolin stimulates viral internal ribosome entry site-mediated translation. Virus Res. 2001, 76, 17–29. [Google Scholar] [CrossRef]
- Pacheco, A.; Lopez de Quinto, S.; Ramajo, J.; Fernandez, N.; Martinez-Salas, E. A novel role for Gemin5 in mRNA translation. Nucleic Acids Res. 2009, 37, 582–590. [Google Scholar] [CrossRef]
- Cao, H.; Zhao, K.; Yao, Y.; Guo, J.; Gao, X.; Yang, Q.; Guo, M.; Zhu, W.; Wang, Y.; Wu, C.; et al. RNA binding protein 24 regulates the translation and replication of hepatitis C virus. Protein Cell 2018, 9, 930–944. [Google Scholar] [CrossRef] [Green Version]
- Ali, N.; Siddiqui, A. Interaction of polypyrimidine tract-binding protein with the 5′ noncoding region of the hepatitis C virus RNA genome and its functional requirement in internal initiation of translation. J. Virol. 1995, 69, 6367–6375. [Google Scholar] [CrossRef] [Green Version]
- Anwar, A.; Ali, N.; Tanveer, R.; Siddiqui, A. Demonstration of functional requirement of polypyrimidine tract-binding protein by SELEX RNA during hepatitis C virus internal ribosome entry site-mediated translation initiation. J. Biol. Chem. 2000, 275, 34231–34235. [Google Scholar] [CrossRef] [Green Version]
- Gosert, R.; Chang, K.H.; Rijnbrand, R.; Yi, M.; Sangar, D.V.; Lemon, S.M. Transient expression of cellular polypyrimidine-tract binding protein stimulates cap-independent translation directed by both picornaviral and flaviviral internal ribosome entry sites In vivo. Mol. Cell. Biol. 2000, 20, 1583–1595. [Google Scholar] [CrossRef] [Green Version]
- Nishimura, T.; Saito, M.; Takano, T.; Nomoto, A.; Kohara, M.; Tsukiyama-Kohara, K. Comparative aspects on the role of polypyrimidine tract-binding protein in internal initiation of hepatitis C virus and picornavirus RNAs. Comp. Immunol. Microbiol. Infect. Dis. 2008, 31, 435–448. [Google Scholar] [CrossRef] [PubMed]
- Shirasaki, T.; Honda, M.; Mizuno, H.; Shimakami, T.; Okada, H.; Sakai, Y.; Murakami, S.; Wakita, T.; Kaneko, S. La protein required for internal ribosome entry site-directed translation is a potential therapeutic target for hepatitis C virus replication. J. Infect. Dis. 2010, 202, 75–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tischendorf, J.J.; Beger, C.; Korf, M.; Manns, M.P.; Kruger, M. Polypyrimidine tract-binding protein (PTB) inhibits Hepatitis C virus internal ribosome entry site (HCV IRES)-mediated translation, but does not affect HCV replication. Arch. Virol. 2004, 149, 1955–1970. [Google Scholar] [CrossRef] [PubMed]
- Kaminski, A.; Hunt, S.L.; Patton, J.G.; Jackson, R.J. Direct evidence that polypyrimidine tract binding protein (PTB) is essential for internal initiation of translation of encephalomyocarditis virus RNA. RNA 1995, 1, 924–938. [Google Scholar] [PubMed]
- Brocard, M.; Paulous, S.; Komarova, A.V.; Deveaux, V.; Kean, K.M. Evidence that PTB does not stimulate HCV IRES-driven translation. Virus Genes 2007, 35, 5–15. [Google Scholar] [CrossRef]
- Rosenfeld, A.B.; Racaniello, V.R. Hepatitis C virus internal ribosome entry site-dependent translation in Saccharomyces cerevisiae is independent of polypyrimidine tract-binding protein, poly(rC)-binding protein 2, and La protein. J. Virol. 2005, 79, 10126–10137. [Google Scholar] [CrossRef] [Green Version]
- Kamath, R.V.; Leary, D.J.; Huang, S. Nucleocytoplasmic shuttling of polypyrimidine tract-binding protein is uncoupled from RNA export. Mol. Biol. Cell 2001, 12, 3808–3820. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Yen, T.S. Characterization of the nuclear export signal of polypyrimidine tract-binding protein. J. Biol. Chem. 2002, 277, 10306–10314. [Google Scholar] [CrossRef] [Green Version]
- Sawicka, K.; Bushell, M.; Spriggs, K.A.; Willis, A.E. Polypyrimidine-tract-binding protein: A multifunctional RNA-binding protein. Biochem. Soc. Trans. 2008, 36, 641–647. [Google Scholar] [CrossRef] [Green Version]
- Gil, A.; Sharp, P.A.; Jamison, S.F.; Garcia-Blanco, M.A. Characterization of cDNAs encoding the polypyrimidine tract-binding protein. Genes Dev. 1991, 5, 1224–1236. [Google Scholar] [CrossRef] [Green Version]
- Romanelli, M.G.; Lorenzi, P.; Morandi, C. Organization of the human gene encoding heterogeneous nuclear ribonucleoprotein type I (hnRNP I) and characterization of hnRNP I related pseudogene. Gene 2000, 255, 267–272. [Google Scholar] [CrossRef]
- Wollerton, M.C.; Gooding, C.; Robinson, F.; Brown, E.C.; Jackson, R.J.; Smith, C.W. Differential alternative splicing activity of isoforms of polypyrimidine tract binding protein (PTB). RNA 2001, 7, 819–832. [Google Scholar] [CrossRef] [Green Version]
- Patton, J.G.; Mayer, S.A.; Tempst, P.; Nadal-Ginard, B. Characterization and molecular cloning of polypyrimidine tract-binding protein: A component of a complex necessary for pre-mRNA splicing. Genes Dev. 1991, 5, 1237–1251. [Google Scholar] [CrossRef] [Green Version]
- Wagner, E.J.; Carstens, R.P.; Garcia-Blanco, M.A. A novel isoform ratio switch of the polypyrimidine tract binding protein. Electrophoresis 1999, 20, 1082–1086. [Google Scholar] [CrossRef]
- Wang, C.; Norton, J.T.; Ghosh, S.; Kim, J.; Fushimi, K.; Wu, J.Y.; Stack, M.S.; Huang, S. Polypyrimidine tract-binding protein (PTB) differentially affects malignancy in a cell line-dependent manner. J. Biol. Chem. 2008, 283, 20277–20287. [Google Scholar] [CrossRef] [Green Version]
- Ghetti, A.; Pinol-Roma, S.; Michael, W.M.; Morandi, C.; Dreyfuss, G. hnRNP I, the polypyrimidine tract-binding protein: Distinct nuclear localization and association with hnRNAs. Nucleic Acids Res. 1992, 20, 3671–3678. [Google Scholar] [CrossRef]
- Perez, I.; McAfee, J.G.; Patton, J.G. Multiple RRMs contribute to RNA binding specificity and affinity for polypyrimidine tract binding protein. Biochemistry 1997, 36, 11881–11890. [Google Scholar] [CrossRef]
- Conte, M.R.; Grune, T.; Ghuman, J.; Kelly, G.; Ladas, A.; Matthews, S.; Curry, S. Structure of tandem RNA recognition motifs from polypyrimidine tract binding protein reveals novel features of the RRM fold. EMBO J. 2000, 19, 3132–3141. [Google Scholar] [CrossRef]
- Oberstrass, F.C.; Auweter, S.D.; Erat, M.; Hargous, Y.; Henning, A.; Wenter, P.; Reymond, L.; Amir-Ahmady, B.; Pitsch, S.; Black, D.L.; et al. Structure of PTB bound to RNA: Specific binding and implications for splicing regulation. Science 2005, 309, 2054–2057. [Google Scholar] [CrossRef]
- Vitali, F.; Henning, A.; Oberstrass, F.C.; Hargous, Y.; Auweter, S.D.; Erat, M.; Allain, F.H. Structure of the two most C-terminal RNA recognition motifs of PTB using segmental isotope labeling. EMBO J. 2006, 25, 150–162. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Blanco, M.A.; Jamison, S.F.; Sharp, P.A. Identification and purification of a 62,000-dalton protein that binds specifically to the polypyrimidine tract of introns. Genes Dev. 1989, 3, 1874–1886. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Valcarcel, J.; Green, M.R. Distinct binding specificities and functions of higher eukaryotic polypyrimidine tract-binding proteins. Science 1995, 268, 1173–1176. [Google Scholar] [CrossRef] [PubMed]
- Clerte, C.; Hall, K.B. The domains of polypyrimidine tract binding protein have distinct RNA structural preferences. Biochemistry 2009, 48, 2063–2074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitchell, S.A.; Spriggs, K.A.; Coldwell, M.J.; Jackson, R.J.; Willis, A.E. The Apaf-1 internal ribosome entry segment attains the correct structural conformation for function via interactions with PTB and unr. Mol. Cell 2003, 11, 757–771. [Google Scholar] [CrossRef] [PubMed]
- Xue, Y.; Zhou, Y.; Wu, T.; Zhu, T.; Ji, X.; Kwon, Y.S.; Zhang, C.; Yeo, G.; Black, D.L.; Sun, H.; et al. Genome-wide analysis of PTB-RNA interactions reveals a strategy used by the general splicing repressor to modulate exon inclusion or skipping. Mol. Cell 2009, 36, 996–1006. [Google Scholar] [CrossRef] [Green Version]
- Caceres, C.J.; Contreras, N.; Angulo, J.; Vera-Otarola, J.; Pino-Ajenjo, C.; Llorian, M.; Ameur, M.; Lisboa, F.; Pino, K.; Lowy, F.; et al. Polypyrimidine tract-binding protein binds to the 5′ untranslated region of the mouse mammary tumor virus mRNA and stimulates cap-independent translation initiation. FEBS J. 2016, 283, 1880–1901. [Google Scholar] [CrossRef] [Green Version]
- Barria, M.I.; Gonzalez, A.; Vera-Otarola, J.; Leon, U.; Vollrath, V.; Marsac, D.; Monasterio, O.; Perez-Acle, T.; Soza, A.; Lopez-Lastra, M. Analysis of natural variants of the hepatitis C virus internal ribosome entry site reveals that primary sequence plays a key role in cap-independent translation. Nucleic Acids Res. 2009, 37, 957–971. [Google Scholar] [CrossRef]
- Wilson, J.E.; Powell, M.J.; Hoover, S.E.; Sarnow, P. Naturally occurring dicistronic cricket paralysis virus RNA is regulated by two internal ribosome entry sites. Mol. Cell Biol. 2000, 20, 4990–4999. [Google Scholar] [CrossRef] [Green Version]
- Gooding, C.; Edge, C.; Lorenz, M.; Coelho, M.B.; Winters, M.; Kaminski, C.F.; Cherny, D.; Eperon, I.C.; Smith, C.W. MBNL1 and PTB cooperate to repress splicing of Tpm1 exon 3. Nucleic Acids Res. 2013, 41, 4765–4782. [Google Scholar] [CrossRef] [Green Version]
- Caceres, C.J.; Angulo, J.; Lowy, F.; Contreras, N.; Walters, B.; Olivares, E.; Allouche, D.; Merviel, A.; Pino, K.; Sargueil, B.; et al. Non-canonical translation initiation of the spliced mRNA encoding the human T-cell leukemia virus type 1 basic leucine zipper protein. Nucleic Acids Res. 2018, 46, 11030–11047. [Google Scholar] [CrossRef]
- Angulo, J.; Ulryck, N.; Deforges, J.; Chamond, N.; Lopez-Lastra, M.; Masquida, B.; Sargueil, B. LOOP IIId of the HCV IRES is essential for the structural rearrangement of the 40S-HCV IRES complex. Nucleic Acids Res. 2016, 44, 1309–1325. [Google Scholar] [CrossRef]
- Sargueil, B.; McKenna, J.; Burke, J.M. Analysis of the functional role of a G.A sheared base pair by in vitro genetics. J. Biol. Chem. 2000, 275, 32157–32166. [Google Scholar] [CrossRef] [Green Version]
- Weill, L.; Louis, D.; Sargueil, B. Selection and evolution of NTP-specific aptamers. Nucleic Acids Res. 2004, 32, 5045–5058. [Google Scholar] [CrossRef] [Green Version]
- De Bisschop, G.; Ameur, M.; Ulryck, N.; Benattia, F.; Ponchon, L.; Sargueil, B.; Chamond, N. HIV-1 gRNA, a biological substrate, uncovers the potency of DDX3X biochemical activity. Biochimie 2019, 164, 83–94. [Google Scholar] [CrossRef]
- Olivares, E.; Landry, D.M.; Caceres, C.J.; Pino, K.; Rossi, F.; Navarrete, C.; Huidobro-Toro, J.P.; Thompson, S.R.; Lopez-Lastra, M. The 5′ untranslated region of the human T-cell lymphotropic virus type 1 mRNA enables cap-independent translation initiation. J. Virol. 2014, 88, 5936–5955. [Google Scholar] [CrossRef] [Green Version]
- Locker, N.; Chamond, N.; Sargueil, B. A conserved structure within the HIV gag open reading frame that controls translation initiation directly recruits the 40S subunit and eIF3. Nucleic Acids Res. 2011, 39, 2367–2377. [Google Scholar] [CrossRef]
- Kafasla, P.; Morgner, N.; Poyry, T.A.; Curry, S.; Robinson, C.V.; Jackson, R.J. Polypyrimidine tract binding protein stabilizes the encephalomyocarditis virus IRES structure via binding multiple sites in a unique orientation. Mol. Cell 2009, 34, 556–568. [Google Scholar] [CrossRef] [Green Version]
- Waysbort, A.; Bonnal, S.; Audigier, S.; Esteve, J.P.; Prats, A.C. Pyrimidine tract binding protein and La autoantigen interact differently with the 5′ untranslated regions of lentiviruses and oncoretrovirus mRNAs. FEBS Lett. 2001, 490, 54–58. [Google Scholar] [CrossRef]
- Han, Y.; Wang, S.; Zhang, Z.; Ma, X.; Li, W.; Zhang, X.; Deng, J.; Wei, H.; Li, Z.; Zhang, X.E.; et al. In vivo imaging of protein-protein and RNA-protein interactions using novel far-red fluorescence complementation systems. Nucleic Acids Res. 2014, 42, e103. [Google Scholar] [CrossRef] [Green Version]
- Luo, G. Cellular proteins bind to the poly(U) tract of the 3′ untranslated region of hepatitis C virus RNA genome. Virology 1999, 256, 105–118. [Google Scholar] [CrossRef] [Green Version]
- Ito, T.; Lai, M.M. An internal polypyrimidine-tract-binding protein-binding site in the hepatitis C virus RNA attenuates translation, which is relieved by the 3′-untranslated sequence. Virology 1999, 254, 288–296. [Google Scholar] [CrossRef] [PubMed]
- Chung, R.T.; Kaplan, L.M. Heterogeneous nuclear ribonucleoprotein I (hnRNP-I/PTB) selectively binds the conserved 3′ terminus of hepatitis C viral RNA. Biochem. Biophys. Res. Commun. 1999, 254, 351–362. [Google Scholar] [CrossRef] [PubMed]
- Chang, K.S.; Luo, G. The polypyrimidine tract-binding protein (PTB) is required for efficient replication of hepatitis C virus (HCV) RNA. Virus Res. 2006, 115, 1–8. [Google Scholar] [CrossRef]
- Domitrovich, A.M.; Diebel, K.W.; Ali, N.; Sarker, S.; Siddiqui, A. Role of La autoantigen and polypyrimidine tract-binding protein in HCV replication. Virology 2005, 335, 72–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pilipenko, E.V.; Viktorova, E.G.; Guest, S.T.; Agol, V.I.; Roos, R.P. Cell-specific proteins regulate viral RNA translation and virus-induced disease. EMBO J. 2001, 20, 6899–6908. [Google Scholar] [CrossRef]
- Dumas, E.; Staedel, C.; Colombat, M.; Reigadas, S.; Chabas, S.; Astier-Gin, T.; Cahour, A.; Litvak, S.; Ventura, M. A promoter activity is present in the DNA sequence corresponding to the hepatitis C virus 5′ UTR. Nucleic Acids Res. 2003, 31, 1275–1281. [Google Scholar] [CrossRef]
- Van Eden, M.E.; Byrd, M.P.; Sherrill, K.W.; Lloyd, R.E. Demonstrating internal ribosome entry sites in eukaryotic mRNAs using stringent RNA test procedures. RNA 2004, 10, 720–730. [Google Scholar] [CrossRef] [Green Version]
- Wagner, E.J.; Garcia-Blanco, M.A. Polypyrimidine tract binding protein antagonizes exon definition. Mol. Cell Biol. 2001, 21, 3281–3288. [Google Scholar] [CrossRef] [Green Version]
- Kafasla, P.; Lin, H.; Curry, S.; Jackson, R.J. Activation of picornaviral IRESs by PTB shows differential dependence on each PTB RNA-binding domain. RNA 2011, 17, 1120–1131. [Google Scholar] [CrossRef] [Green Version]
- Zignego, A.L.; Macchia, D.; Monti, M.; Thiers, V.; Mazzetti, M.; Foschi, M.; Maggi, E.; Romagnani, S.; Gentilini, P.; Brechot, C. Infection of peripheral mononuclear blood cells by hepatitis C virus. J. Hepatol. 1992, 15, 382–386. [Google Scholar] [CrossRef]
- Bartosch, B.; Dubuisson, J.; Cosset, F.L. Infectious hepatitis C virus pseudo-particles containing functional E1-E2 envelope protein complexes. J. Exp. Med. 2003, 197, 633–642. [Google Scholar] [CrossRef]
- Bung, C.; Bochkaeva, Z.; Terenin, I.; Zinovkin, R.; Shatsky, I.N.; Niepmann, M. Influence of the hepatitis C virus 3′-untranslated region on IRES-dependent and cap-dependent translation initiation. FEBS Lett. 2010, 584, 837–842. [Google Scholar] [CrossRef]
- Shi, G.; Yagyu, F.; Shimizu, Y.; Shimizu, K.; Oshima, M.; Iwamoto, A.; Gao, B.; Liu, W.; Gao, G.F.; Kitamura, Y. Flow cytometric assay using two fluorescent proteins for the function of the internal ribosome entry site of hepatitis C virus. Cytom. A 2011, 79, 653–660. [Google Scholar] [CrossRef]
- Romanelli, M.G.; Diani, E.; Lievens, P.M. New insights into functional roles of the polypyrimidine tract-binding protein. Int. J. Mol. Sci. 2013, 14, 22906–22932. [Google Scholar] [CrossRef] [Green Version]
- Hunt, S.L.; Jackson, R.J. Polypyrimidine-tract binding protein (PTB) is necessary, but not sufficient, for efficient internal initiation of translation of human rhinovirus-2 RNA. RNA 1999, 5, 344–359. [Google Scholar] [CrossRef]
- Kaminski, A.; Jackson, R.J. The polypyrimidine tract binding protein (PTB) requirement for internal initiation of translation of cardiovirus RNAs is conditional rather than absolute. RNA 1998, 4, 626–638. [Google Scholar] [CrossRef] [Green Version]
- Vopalensky, V.; Masek, T.; Horvath, O.; Vicenova, B.; Mokrejs, M.; Pospisek, M. Firefly luciferase gene contains a cryptic promoter. RNA 2008, 14, 1720–1729. [Google Scholar] [CrossRef] [Green Version]
- Honda, M.; Kaneko, S.; Matsushita, E.; Kobayashi, K.; Abell, G.A.; Lemon, S.M. Cell cycle regulation of hepatitis C virus internal ribosomal entry site-directed translation. Gastroenterology 2000, 118, 152–162. [Google Scholar] [CrossRef]
- Venkatesan, A.; Sharma, R.; Dasgupta, A. Cell cycle regulation of hepatitis C and encephalomyocarditis virus internal ribosome entry site-mediated translation in human embryonic kidney 293 cells. Virus Res. 2003, 94, 85–95. [Google Scholar] [CrossRef]
- Lamichhane, R.; Daubner, G.M.; Thomas-Crusells, J.; Auweter, S.D.; Manatschal, C.; Austin, K.S.; Valniuk, O.; Allain, F.H.; Rueda, D. RNA looping by PTB: Evidence using FRET and NMR spectroscopy for a role in splicing repression. Proc. Natl. Acad. Sci. USA 2010, 107, 4105–4110. [Google Scholar] [CrossRef] [Green Version]
- Chahal, J.; Gebert, L.F.R.; Gan, H.H.; Camacho, E.; Gunsalus, K.C.; MacRae, I.J.; Sagan, S.M. miR-122 and Ago interactions with the HCV genome alter the structure of the viral 5′ terminus. Nucleic Acids Res. 2019, 47, 5307–5324. [Google Scholar] [CrossRef] [PubMed]
- Jangra, R.K.; Yi, M.; Lemon, S.M. Regulation of hepatitis C virus translation and infectious virus production by the microRNA miR-122. J. Virol. 2010, 84, 6615–6625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roberts, A.P.; Lewis, A.P.; Jopling, C.L. miR-122 activates hepatitis C virus translation by a specialized mechanism requiring particular RNA components. Nucleic Acids Res. 2011, 39, 7716–7729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hunt, S.L.; Hsuan, J.J.; Totty, N.; Jackson, R.J. unr, a cellular cytoplasmic RNA-binding protein with five cold-shock domains, is required for internal initiation of translation of human rhinovirus RNA. Genes Dev. 1999, 13, 437–448. [Google Scholar] [CrossRef] [Green Version]
- Pilipenko, E.V.; Pestova, T.V.; Kolupaeva, V.G.; Khitrina, E.V.; Poperechnaya, A.N.; Agol, V.I.; Hellen, C.U. A cell cycle-dependent protein serves as a template-specific translation initiation factor. Genes Dev. 2000, 14, 2028–2045. [Google Scholar] [CrossRef]
- King, H.A.; Cobbold, L.C.; Pichon, X.; Poyry, T.; Wilson, L.A.; Booden, H.; Jukes-Jones, R.; Cain, K.; Lilley, K.S.; Bushell, M.; et al. Remodelling of a polypyrimidine tract-binding protein complex during apoptosis activates cellular IRESs. Cell Death Differ. 2014, 21, 161–171. [Google Scholar] [CrossRef] [Green Version]
- Jahan, N.; Wimmer, E.; Mueller, S. Polypyrimidine tract binding protein-1 (PTB1) is a determinant of the tissue and host tropism of a human rhinovirus/poliovirus chimera PV1(RIPO). PLoS ONE 2013, 8, e60791. [Google Scholar] [CrossRef] [Green Version]
- Guest, S.; Pilipenko, E.; Sharma, K.; Chumakov, K.; Roos, R.P. Molecular mechanisms of attenuation of the Sabin strain of poliovirus type 3. J. Virol. 2004, 78, 11097–11107. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Angulo, J.; Cáceres, C.J.; Contreras, N.; Fernández-García, L.; Chamond, N.; Ameur, M.; Sargueil, B.; López-Lastra, M. Polypyrimidine-Tract-Binding Protein Isoforms Differentially Regulate the Hepatitis C Virus Internal Ribosome Entry Site. Viruses 2023, 15, 8. https://doi.org/10.3390/v15010008
Angulo J, Cáceres CJ, Contreras N, Fernández-García L, Chamond N, Ameur M, Sargueil B, López-Lastra M. Polypyrimidine-Tract-Binding Protein Isoforms Differentially Regulate the Hepatitis C Virus Internal Ribosome Entry Site. Viruses. 2023; 15(1):8. https://doi.org/10.3390/v15010008
Chicago/Turabian StyleAngulo, Jenniffer, C. Joaquín Cáceres, Nataly Contreras, Leandro Fernández-García, Nathalie Chamond, Melissa Ameur, Bruno Sargueil, and Marcelo López-Lastra. 2023. "Polypyrimidine-Tract-Binding Protein Isoforms Differentially Regulate the Hepatitis C Virus Internal Ribosome Entry Site" Viruses 15, no. 1: 8. https://doi.org/10.3390/v15010008