
Use of Antiandrogens as Therapeutic Agents in COVID-19 Patients

1
Department of Infectious Diseases, Imperial College London, London W2 1PG, UK
2
School of Life Sciences, University of Essex, Colchester CO4 3SQ, UK
*
Author to whom correspondence should be addressed.
Viruses 2022, 14(12), 2728; https://doi.org/10.3390/v14122728
Submission received: 31 October 2022
/
Revised: 23 November 2022
/
Accepted: 1 December 2022
/
Published: 7 December 2022
(This article belongs to the Special Issue SARS-CoV-2 inside and outside the Respiratory Tract: From Diagnostics to Therapies)
{kind=link}
{kind=link}
Abstract
:COVID-19, caused by the severe acute respiratory syndrome coronavirus 2 (SARS CoV-2), is estimated to have caused over 6.5 million deaths worldwide. The emergence of fast-evolving SARS-CoV-2 variants of concern alongside increased transmissibility and/or virulence, as well as immune and vaccine escape capabilities, highlight the urgent need for more effective antivirals to combat the disease in the long run along with regularly updated vaccine boosters. One of the early risk factors identified during the COVID-19 pandemic was that men are more likely to become infected by the virus, more likely to develop severe disease and exhibit a higher likelihood of hospitalisation and mortality rates compared to women. An association exists between SARS-CoV-2 infectiveness and disease severity with sex steroid hormones and, in particular, androgens. Several studies underlined the importance of the androgen-mediated regulation of the host protease TMPRSS2 and the cell entry protein ACE2, as well as the key role of these factors in the entry of the virus into target cells. In this context, modulating androgen signalling is a promising strategy to block viral infection, and antiandrogens could be used as a preventative measure at the pre- or early hospitalisation stage of COVID-19 disease. Different antiandrogens, including commercial drugs used to treat metastatic castration-sensitive prostate cancer and other conditions, have been tested as antivirals with varying success. In this review, we summarise the most recent updates concerning the use of antiandrogens as prophylactic and therapeutic options for COVID-19.
1. Emergence of COVID-19 and Variants of Concern
Coronaviruses (CoV) are a large family of positive-sense, enveloped, single-stranded RNA viruses that are common in people and many animal species. Human coronaviruses (hCoV) include the common cold CoV 229E, OC43, NL63 and HKU1, which predominantly cause mild respiratory illness, and three highly pathogenic coronaviruses, which cause more severe and fatal diseases in people, e.g., severe acute respiratory syndrome coronavirus (SARS-CoV, 2003), Middle East respiratory syndrome coronavirus (MERS-CoV, 2012) and more recently, SARS-CoV-2 (2019), the causative agent of COVID-19 [1]. At the end of September 2022, over 600 million cases of COVID-19 were confirmed globally, with a 4–5% crude mortality risk (deaths per 100 patients hospitalized primarily for COVID-19) [2]. COVID-19 may present with mild, moderate or severe symptoms in patients. Mild symptoms include fever, dry cough, dyspnoea and fatigue, while COVID-19-associated mortality is attributed to severe respiratory failure resembling acute respiratory distress syndrome (ARDS) [3,4,5]. Severe pneumonia COVID-19 patients exhibit exacerbated inflammatory response resulting in excessive release of pro-inflammatory cytokines, known as the “cytokine storm”, which leads to alveolar damage, fibrosis and progressive respiratory failure [5,6,7,8]. It is becoming increasingly clear that COVID-19 is a multisystem disease and can also lead to heart failure, acute kidney failure and neurovascular injuries [5,9,10]. In addition, growing patient testimony and scientific evidence demonstrate that a large number of patients (approximately 2% of the UK population alone) also suffer from post-COVID syndrome (long COVID or post-acute sequelae of COVID-19 (PASC)), which causes debilitating physical or mental symptoms for long periods after infection [5,11,12,13,14].
Vaccination programmes have been key to reducing COVID-19 hospital admissions and mortality rates, but for some countries and groups of people, the uptake is still low due to various socioeconomic factors and accessibility inequalities. In addition, vaccination-acquired immunity wanes substantially with time and boosters are required to restore the efficiency of vaccines against severe diseases [15,16]. Despite progress in countries with high vaccination rates, the pandemic is still actively unfolding with emerging virus variants that can evade vaccine or natural immunity triggering new public health challenges. Virus variants are characterised by the WHO as variants of concern (VOC) if they demonstrate increased transmissibility, virulence or cause reduced effectiveness of vaccine-induced protection, diagnostic tests and management measures [17,18]. Important variants include B.1.1.7 (Alpha), B.1.351 (Beta), P.1 (Gamma) and B.1.617.2 (Delta) [18,19,20]. Two new sub-variants (B.1.1.529; Omicron) designated as BA.4 and BA.5 emerged in December 2021 and January 2022, respectively, which replaced the other circulating Omicron lineages and have driven the current wave of infections in Africa, the United States and Europe [21]. These two subvariants appear more contagious and circumvent immunity from previous infections or vaccination [22,23]. As a result of the emergence of Omicron variants, several updated COVID-19 vaccines have been developed and approved for use in the United States, United Kingdom and elsewhere. However, a recent study suggested that the direct protective benefit of the Omicron-updated bivalent vaccines is only marginal compared to ancestral-based vaccines [24].
COVID-19 hospitalised patient management in high-income countries is largely focused on highly efficient albeit high-priced monoclonal antibodies [25], anti-inflammatory drugs such as dexamethasone, which reduce cytokine levels and immune-mediated pathogenesis in the hyper-inflammatory phase of the disease [26], and/or drugs which directly interfere with viral replication. The latter act either by inhibition of the viral 3CLpro protease (paxlovid), by inhibition of the RNA-dependent RNA polymerase (remdesivir) [27] or by introducing copying errors in the viral genome (molnupiravir) [28]. Recent clinical studies reported that remdesivir has no significant effect on patients who are already being ventilated [29] and that some paxlovid- or molnupiravir-treated patients experience rebound infections after completing a course of treatment [30,31]. Taken together, more efficient and less expensive antiviral drugs for pre-emptive protection against emerging variants, reducing reliance on boosters, are required, especially in low- and middle-income countries.
2. Risk Factors and Gender Disparity in COVID-19 Patients
Most COVID-19 patients are expected to have a favourable prognosis. However, some groups are at higher risk of developing severe or fatal illness due to underlying health conditions. These include demographic factors (i.e., older age), gender, occupational exposure, ethnicity and, more importantly, comorbidities such as diabetes, hypertension, obesity, neurological disorders, cancer, heart, lung, liver and kidney disease [32,33,34,35,36]. Researchers are in the early stages of understanding the risk factors of viral persistence and long COVID. Accumulating evidence shows that people who experienced multisystem inflammatory syndrome (MIS) with underlying health conditions and who were not vaccinated were more likely to develop long-term post-COVID sequelae [37,38,39].
Since the start of the coronavirus pandemic, emerging gender-disaggregated data from multiple countries suggested that men were disproportionally affected by COVID-19. Initially, it was hypothesised that the higher occupational exposure and comorbidities of males compared to females were contributory to this discrepancy. Follow-up studies confirmed that men globally exhibit higher infection, hospitalisation and mortality rates compared to women [40,41,42]. Female COVID-19 patients were also found to clear the virus and recover substantially earlier compared to male patients [43,44]. Sex differences in prevalence, severity and outcome of viral infection were previously reported for other viruses such as influenza viruses, HIV, herpes simplex viruses, hepatitis B, measles and West Nile virus [45,46,47]. The reasons accounting for the higher vulnerability of males to viruses are not clear, but several immunological, as well as genetic, hormonal and socio-behavioural explanations have been put forward.
The innate immune system serves as the first line of host defence against invading viruses and involves sensing viruses through pattern recognition receptors and activating inflammatory pathways, which eventually result in viral clearance. Innate immune cells, i.e., monocytes, dendritic cells and macrophages, are functionally more active in females [48,49,50]. Females are typically able to launch more vigorous innate and adaptive immune responses, have higher resistance to most viral infections and exhibit higher antibody responses following infection and vaccination compared to males [48,51,52,53]. The bi-allelic expression of immune-related genes present on the X chromosome, such as the RNA receptors TLR7 and TLR8, may be responsible for sexually dimorphic immunophenotypes [52,53,54]. Hormonal regulation may also be responsible for the gender disparity in disease progression and outcomes following viral challenges. Steroid hormones, such as androgens, oestrogens, progesterone and glucocorticoids, control the activity of innate immune cells and the survival and differentiation of T and B lymphocytes in both men and women [48,54,55,56]. Many studies have been published on the effects of steroid hormones on immune responses. The consensus is that androgens (primarily male hormones) are predominantly immunosuppressive while oestrogens (the primary female hormones) are mostly immunoprotective [49,50,57,58,59,60], and they can thus potentially suppress or enhance antiviral responses, respectively. In addition to their effects on antiviral responses, androgens can also directly influence virus activity, i.e., HBV and HCV [61]. Conversely, viruses are also able to manipulate sex steroid receptor signalling mechanisms to serve their own survival and enhance their replication rate [62]. The extent to which male and female immune responses differ during SARS-CoV-2 infection and the possibility of a bi-directional interaction between the virus and androgens have been overlooked so far and deserve greater attention.
3. Cell Entry of SARS-CoV-2
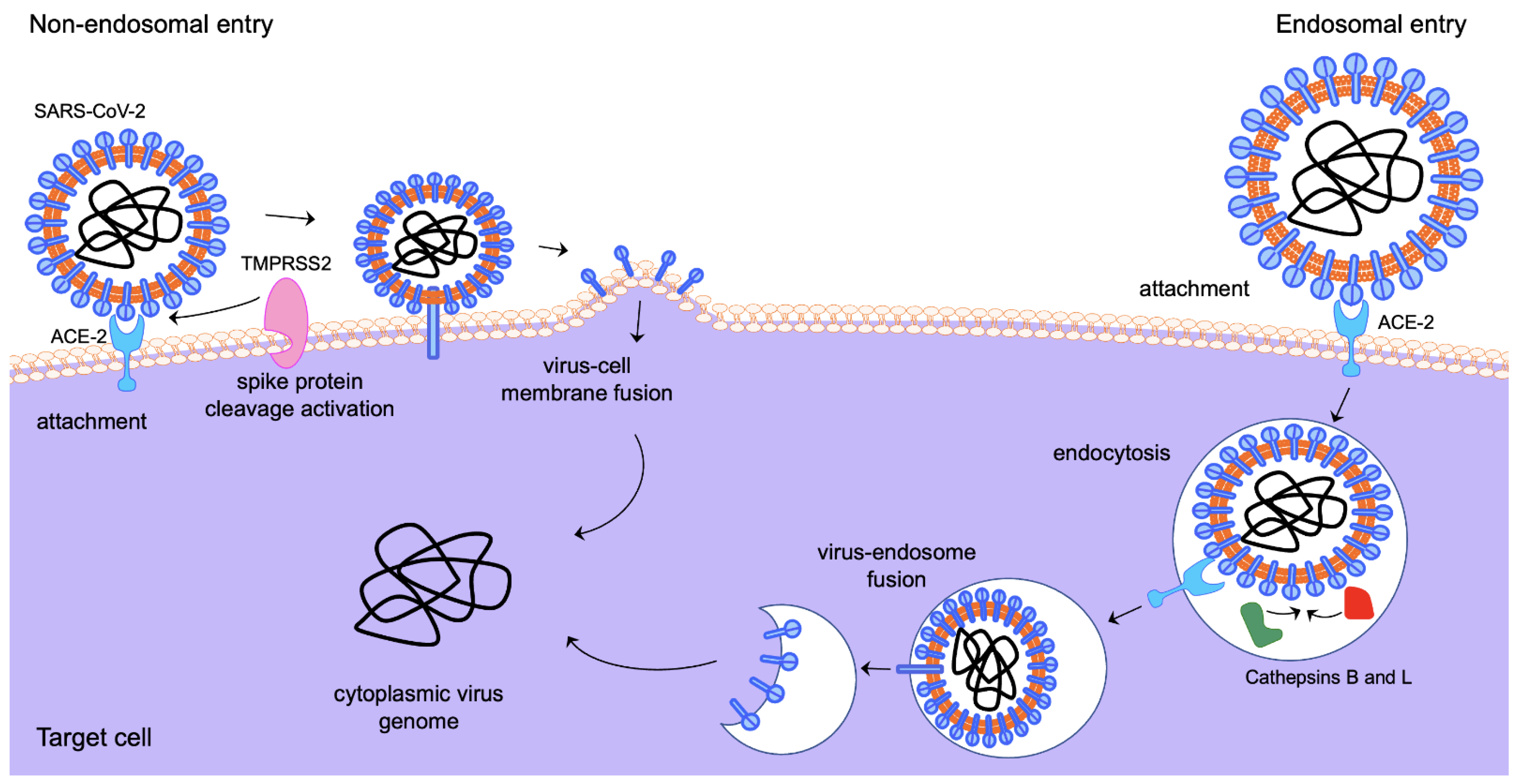
Viral attachment, fusion and entry of SARS-CoV-2 into target cells are mediated by the virus’ envelope spike (S) glycoprotein [63,64,65]. The S protein is the main viral antigenic site for inducing antibody responses and is, therefore, a key target for the development of antivirals and vaccines. S is synthesised as a precursor protein that is cleaved into two subunits: the N-terminal S1 subunit, which contains the receptor binding domain, and the C-terminal S2 subunit, which harbours the viral fusion machinery [63,64,66]. SARS-CoV-2 initiates cell entry by binding its S to the transmembrane glycoprotein Angiotensin-Converting Enzyme 2 (ACE2), a negative controller of the Renin–Angiotensin system [67,68]. Following binding, the virus gains access to the cell by two alternative routes (Figure 1), either via direct fusion with the cell membrane or via clathrin-mediated endocytosis [64,65,66]. Both entry mechanisms require priming of the S protein by proteolytic cleavage at two sites in a manner reminiscent of that of the proteolytic cleavage and maturation of influenza A viruses’ entry protein haemagglutinin [69,70]. The first cleavage site is a polybasic insertion (PRRAR) at the S1–S2 boundary and the second cleavage occurs at the S2′ site in the S2 subunit, a crucial step for triggering the fusion of viral and host cell membranes [64,65,71,72]. The S1–S2 site is cleaved by the serine protease furin, which is ubiquitously expressed in the respiratory tract, while the S2′ site can be cleaved by either the endosomal cathepsins (B and L) in the endolysosome or by the cell-surface transmembrane protease serine 2 (TMPRSS2) at the plasma membrane [63,73,74]. TMPRSS2 is part of the type 2 transmembrane serine protease (TTSP) family and has been extensively studied in the context of prostate cancer as its expression is regulated in response to androgens through direct transcriptional regulation by the androgen receptor (AR) [75,76]. Other TTSPs, such as TMPRSS4, may also act as SARS-CoV-2 cell entry mediators [77]. The aftermath of SARS-CoV-2 cell entry is governed by a complicated network of host–pathogen interactions, where a range of viral and host factors can demarcate the disease outcome.
4. ACE2 and TMPRSS2 as Candidate Targets for Antiviral Therapy
A large number of viral and host factors that play key roles in virus pathogenesis have been discovered. Among them, ACE2 and TMPRSS2 have been proposed as the most attractive targets for the development of antivirals that could inhibit or delay virus infection, thus possibly reducing viral transmission and symptom severity. Both factors are expressed in the nasal and bronchial secretory cells, including alveolar epithelial type II cells (AT2), which is the main target cell subpopulation of SARS-CoV-2, as well as gastrointestinal and other virus-targeted tissues [78,79,80,81,82]. ACE2 regulates key processes in the human body, such as blood pressure, cardiovascular function, wound healing and inflammation and acts as the main cellular receptor for SARS-CoV and hCoV NL63 [83,84]. TMPRSS2 is also essential for the cell entry of other viruses, i.e., influenza A and B, HCV, as well as other coronaviruses [85,86,87,88,89].
At the onset of the COVID-19 pandemic, there were studies suggesting that higher ACE2 and TMPRSS2 levels may be associated with higher susceptibility to SARS-CoV-2 infection, and this spurred extensive research that unveiled novel insights into the function and biology of ACE2/TMPRSS2. Several reports revealed an association between the variable expression of ACE2 and TMPRSS2 in different tissues across individuals with COVID-19 severity/fatality variations [90,91,92,93,94]. The lung airway expression of both ACE2 and TMPRSS2 is lower in children who are less susceptible to infection compared with adults, and significantly higher in smokers compared to non-smokers and in patients with chronic obstructive pulmonary disease (COPD) compared to healthy individuals [95,96].
Targeting ACE2 was considered the obvious first-choice target for prophylactic and therapeutic interventions aiming to block virus accessibility to respiratory cells early on in infection [97,98]. Several strategies have been evaluated in vitro and in vivo with varied success, including decoy or soluble ACE2 molecules, pseudoligands with a high ACE2 affinity and blocking antibodies [67,99,100]. However, a disadvantage of these approaches is the potential dysregulation of ACE2-mediated vasodilation, amino acid transport and pancreatic insulin secretion [100,101,102,103]. TMPRSS2-inhibitors are arguably better candidates for COVID-19 antivirals than those for ACE2 as knockout of TMPRSS2 protein causes no overt detrimental phenotype [104,105]. Numerous TMPRSS2 inhibitors have been tested in in vitro studies that showed they inhibit entry of SARS-CoV-2 in lung cells, including notably camostat, nafamostat and several peptidomimetic inhibitors [67,106,107,108,109]. These inhibitors were either ‘repurposed’ commercially available drugs, designed de novo or identified by virtual screenings and have been reviewed extensively elsewhere [110,111,112,113,114,115]. Several of these compounds have already been or are currently being investigated in randomised, controlled trials to assess their use as monotherapies against COVID-19. Clinical trials testing the efficacy of camostat and nafamostat have shown contradictory results. Some studies demonstrated a clear, beneficial effect for COVID-19 patients [116,117,118], while others showed only modest clinical improvements [107]. A possible caveat for these inhibitors is that the currently prevailing Omicron subvariants favour cell entry via the endosomal pathway [119,120,121], and it is possible that inhibitors targeting the TMPRSS2 (non-endosomal) entry route might not have a significant effect. Separate studies have targeted the expression of TMPRSS2 using either androgen receptor (AR) antagonists or drugs that lower circulating androgen levels. The relationship between androgens and COVID-19 is not entirely clear and is even considered controversial by some researchers. Conflicting studies have been published reporting that both subdued or excessive testosterone levels can lead to severe COVID-19 disease, e.g., in hypogonadism in older men and testosterone-treated postmenopausal women, respectively, which suggests that the androgen-mediated effects of COVID-19 patients are likely multidimensional and interdependent on a range of confounding factors [122].
5. Androgen Receptor Structure and Signalling
The androgen receptor (AR), a member of the steroid receptor family, is a ligand-dependent transcription factor with a modular structure [123]. The N-terminal activation function (AF-1) is a region that mediates protein–protein interactions and is important for transcriptional activity. Adjacent to this region is the DNA binding domain (DBD), consisting of two zinc-finger-like modules, which modulate the interaction of the receptors with specific DNA response elements present in the regulatory regions of target genes. The C-terminus houses the ligand binding domain (LBD)/activation function 2 (AF2). The AR LBD is formed of 12 α-helices which fold to form a pocket into which androgen fits. Ligand binding promotes a conformational change, resulting in receptor activation [124,125].
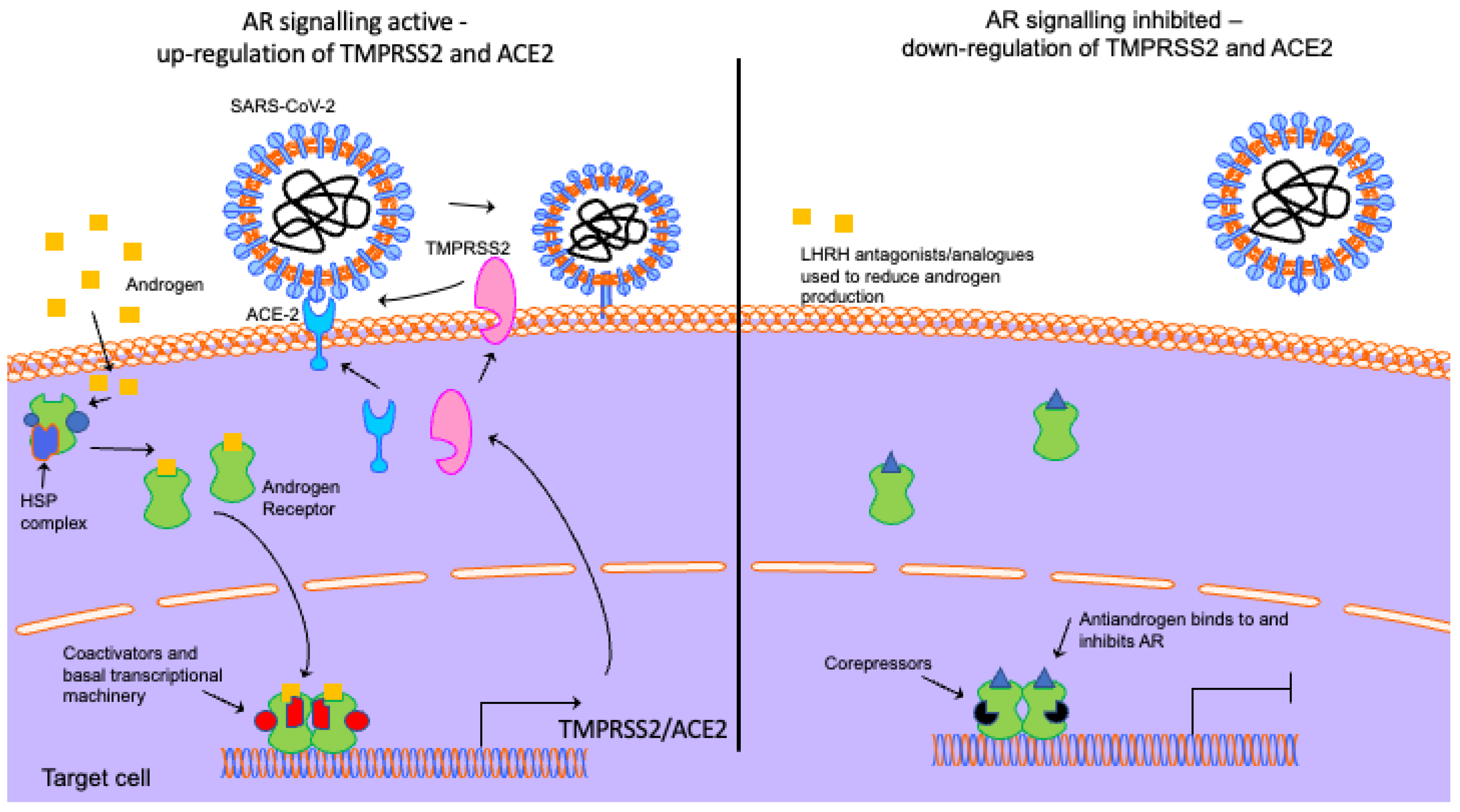
In the absence of a ligand, the AR is located in the cytoplasm and is held in a ligand-binding competent state by a heat shock protein complex (Figure 2). Testosterone can diffuse into the cell and is converted to the more potent androgen dihydrotestosterone (DHT) by 5α-reductase. Upon ligand binding, the AR undergoes a conformational change which promotes dissociation of the heat shock protein complex, dimerisation and nuclear localisation. Next, the AR binds to androgen response elements (AREs) in the regulatory regions of target genes and, via the recruitment of coactivators (proteins that enhance the receptor’s transcriptional activity) and the basal transcriptional machinery, enhances or represses gene expression [123,124,125,126].
6. Androgen Receptor Activity in the Lung and Regulation of TMPRSS2 and ACE2
Our understanding of AR action has predominantly come from studies that have investigated the receptor’s role in the prostate, a small secretory gland at the base of the bladder, and prostate cancer [126]. In the prostate, the AR regulates various genes important in the development and function of the gland. In prostate cancer, the AR has been shown to regulate a transcriptional profile that promotes tumour growth. However, it is well documented that the AR is expressed in, and plays an important role in, other organs [125]. For example, analysis of an AR reporter mouse demonstrated that the AR is expressed and transcriptionally active in multiple organs in male and female mice, including the testes, prostate, ovaries, uterus, salivary glands and spleen [127]. Importantly, the study also demonstrated that AR is transcriptionally active in the lung, albeit weakly. Further, microarray analysis of the AR transcriptome in the lung adenocarcinoma cell line A549 demonstrated that the AR regulates pathways involved in, e.g., oxygen transport, DNA repair and DNA recombination [128].
Several studies have investigated AR expression in the lungs, with single-cell analysis demonstrating that the receptor is expressed in multiple cell types, e.g., club cells [129,130,131]. Importantly, the AR was shown to be co-expressed with TMPRSS2 and ACE2 in AT2 cells [130], the cell type predominantly targeted by SARS-CoV-2 [132]. The AR has been shown to regulate the expression of TMPRSS2 in multiple cell types, including prostate, breast and lung cells [75,128,130,133]. Analysis of the regions upstream of TMPRSS2 identified multiple potential AREs, with a binding site approximately 13 kb upstream from the start site being crucial for optimal androgen regulation of the gene [133]. Interestingly, chromatin immunoprecipitation analysis of AR binding upstream regions of TMPRSS2 demonstrated that this regulation might be tissue-specific [130,134]. In the prostate cancer cell line LNCaP, the AR was found to bind to response elements present in approximately the first 80 kb upstream of TMPRSS2. However, in the immortalised lung cells A549 and H1944, the AR is bound to more distal regions, e.g., binding sites present approximately 100–200 kb upstream of the transcriptional start site [130]. ACE2 expression has also been shown to be regulated by the AR. For example, Baratchian et al. demonstrated that ACE2 is regulated at the RNA and protein levels by the AR in LNCaP cells [129]. Similar to TMPRSS2, AR binding sites were also identified upstream of the ACE2 gene, suggesting direct gene regulation by androgens.
7. Androgen Receptor Modulators as Treatments for COVID
AR regulation of TMPRSS2 and ACE2 expression has led to speculation that androgens could contribute to SARS-CoV-2 infection and disease severity (e.g., [135]). Subsequently, it was hypothesised that inhibition of AR signalling could be a method to reduce SARS-CoV-2 infection/treat the disease [136]. Multiple inhibitors of the AR signalling pathway have been developed, and these either block androgen synthesis, also known as androgen deprivation therapies (ADT, e.g., LHRH antagonists/agonists), or bind directly to the receptor and inhibit its activity (antiandrogens). These androgen signalling inhibitors are widely used in men for the treatment of prostate cancer as well as for other diseases such as benign prostatic hyperplasia and male pattern baldness/androgenetic alopecia [137,138]. They have also been trialled/used in women to treat diseases such as breast cancer, ovarian cancer and polycystic ovarian syndrome [139,140,141,142].
Antiandrogens (e.g., enzalutamide) have been shown to significantly reduce ACE2 and TMRPSS2 expression in prostate and lung cell lines [130,134]. Castration of mice (removal of testicular androgen production) and treatment of mice with enzalutamide has also been shown to reduce ACE2 and/or TMPRSS2 expression in the mouse lung [130,143]. Importantly, inhibition of AR signalling has been shown to block cellular entry of pseudotyped virus expressing the SARS-CoV-2 spike protein and also the authentic virus in lung and prostate cancer cell lines [130]. However, Li et al. found that enzalutamide was unable to block SARS-CoV-2 infection in human lung organoids [134].
8. Retrospective/Observational Studies to Assess the Potential Efficacy of ADT in Relation to COVID-19
To assess the potential benefits of ADT in relation to COVID-19, several observational studies have been undertaken. The first clinical data to support the hypothesis that ADT could be useful for the management of COVID-19 came from Montopoli et al., who analysed infection rates in prostate cancer patients in the Veneto region of Italy [144]. A comparison of 5273 patients receiving ADT with 37,161 patients not receiving ADT found that although cancer patients were at higher risk of COVID-19 infection, ADT partially protected men from the disease. Soon after, a second but smaller study (22 prostate cancer patients receiving ADT and 36 that were not) performed at Mount Sinai Health System in New York City also found that ADT had potential as a treatment option for COVID-19 [145]. The study found that those receiving ADT were significantly less likely to require hospitalisation or oxygen support. However, ADT was not found to significantly reduce morbidity or intubation [145]. A more recent observational study of veterans in the US also demonstrated that men receiving ADT had reduced incidence of COVID-19 and were less likely to suffer from severe symptoms [146].
In addition to the analysis of prostate cancer patients receiving ADT, studies have also looked at the therapeutic potential of AR signalling inhibitors as a treatment option for COVID in other cohorts of men. For example, McCoy et al. investigated the potential benefit of 5-alpha-reductase inhibitors, molecules that block the conversion of testosterone into the more potent DHT, in men being treated for androgenetic alopecia such as spironolactone [147]. In agreement with the ADT studies, the authors found that there was a significant reduction in the frequency of COVID-19-related symptoms in the men receiving the 5-alpha-reductase inhibitor (n = 48) compared to those not receiving the therapy (n = 65). However, not all epidemiological studies have supported the use of ADT as a therapy for COVID-19. For example, Welén et al. investigated COVID-19 severity in prostate cancer patients in the Swedish national registers who had received different types of ADT [148]. After adjustment for age and comorbidities, the authors found no evidence that ADT protected the patients from infection, nor did it reduce the severity of the disease.
9. Clinical Trials to Assess the Efficacy of ADT as a Treatment Option for COVID-19
Due to the experimental and observational clinical data investigating ADT as a therapeutic approach for COVID-19, multiple clinical trials were initiated. Using novel and experimental antiandrogen proxalutamide, one study showed significant promise in clinical trials. The trial demonstrated a significant improvement in survival and a reduction in hospitalisations in the group receiving the antiandrogen. However, concerns were raised regarding the study design, and the publication has since been retracted [138]. A second larger follow-up multicentre clinical trial was performed for proxalutamide. The trial included a total of 778 subjects who had been hospitalised with COVID-19, 423 of whom received Proxalutamide in addition to standard care. Patients who received proxalutamide were found to have a significantly better recovery rate, reduced mortality rate and, on average, spent less time in the hospital [139].
In contrast to the proxalutamide results, several studies investigating alternative androgen signalling inhibitors have shown limited/no benefit to patients. For example, Nickols et al. undertook a clinical trial to assess the therapeutic value of degarelix, which blocks androgen production through inhibition of the hypothalamus–pituitary signalling axis. The study was terminated after an interim analysis demonstrated that there was no significant difference in outcomes (e.g., mortality, ongoing need for hospitalisation, or requirement for mechanical ventilation) between the degarelix (plus standard care) and placebo groups [149]. Similarly, the COVIDENZA trial also found that inhibition of AR signalling had no therapeutic value for COVID-19. COVIDENZA was a randomised phase 2 clinical trial that investigated the efficacy of the antiandrogen enzalutamide. The trial enrolled 42 hospitalised COVID-19 patients, and following a safety evaluation, the trial was halted as it was found that enzalutamide-treated patients required longer stays in the hospital. A number of other clinical trials that aimed to investigate antiandrogens as a therapeutic approach for COVID-19 were subsequently withdrawn. However, at the time of writing, two proxalutamide clinical trials are ongoing (NCT05009732 and NCT04869228); hopefully, these will provide some clarity on the efficacy of targeting the AR as a treatment option for COVID-19.
10. Conclusions and Further Perspectives
Considering the prohibiting cost of current COVID-19 drug regimens for low- and middle-income countries, the emerging SARS-CoV-2 variants and the COVID-19 vaccine rollout and efficacy challenges, the need for cost-effective, orally available and broad-spectrum antivirals that can act against a wide range of SARS-CoV-2 variants remains urgent [150,151]. Despite the promising antiviral effect that a range of antiandrogens display in vitro against SARS-CoV-2, the results of finalised clinical trials on the efficacy of ADT or antiandrogens in COVID-19 patients have not been conclusive enough to inform clinical practice. Various next-generation antiandrogens have been formulated, and the development of a lot more is underway, including apalutamide, darolutamide, orteronel and galeterone. These new drugs should be explored for their antiviral effects and clinical outcomes as they might be more effective against SARS-CoV-2 and perhaps more amenable for widespread use in COVID-19 patients.
A disadvantage of antiandrogens as standalone therapeutic agents is that alternative TMPRSS2-independent virus entry pathways can counteract the inhibitory effects of antiandrogens on TMPRSS2-dependent viral entry. This might explain the mixed or inconclusive results of clinical trials to date which evaluated the monotherapy of ADT/antiandrogens in COVID-19 patients. The use of ADT/antiandrogens in combination therapy has not been evaluated so far. Combinations of antiviral drugs are more likely to function synergistically if they have distinct mechanisms of action and target different stages of the virus lifecycle. Therefore, while we wait for the outcomes of the remaining ADT clinical trials and notably proxalutamide, the next wave of investigations should be focused on the combination of antiandrogen therapy with other treatments, such as viral replication inhibitors.
In addition, it is surprising that experimental approaches to target TMPRSS2-independent virus entry pathways have been comparably limited, and the crosstalk between androgen signalling and virus pathogenicity has not been investigated thoroughly. The concept that androgens may serve dual roles in SARS-CoV-2 infection is intriguing and remains understudied. Future efforts should be focused on simultaneously targeting alternative viral entry mechanisms and defining better the mechanistic roles of androgens in the respiratory tract. In conclusion, antiandrogens have more exploratory potential as antivirals. Building on results from former clinical trials, future trials should focus on antiandrogens or ADT in combinatorial therapeutic modalities against COVID-19.
Author Contributions
Conceptualisation, data curation, writing—original draft preparation: E.S.G. and G.N.B.; data curation: E.C. All authors have read and agreed to the published version of the manuscript.
Funding
Funding was provided by the University of Essex COVID-19 Rapid and Agile Fund awarded to E.S.G and G.N.B.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Cui, J.; Li, F.; Shi, Z.L. Origin and evolution of pathogenic coronaviruses. Nat. Rev. Microbiol. 2019, 17, 181–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adjei, S.; Hong, K.; Molinari, N.M.; Bull-Otterson, L.; Ajani, U.A.; Gundlapalli, A.V.; Harris, A.M.; Hsu, J.; Kadri, S.S.; Starnes, J.; et al. Mortality risk among patients hospitalized primarily for COVID-19 during the Omicron and Delta variant pandemic periods-United States, April 2020–June 2022. MMWR Morb. Mortal. Wkly. Rep. 2022, 71, 1182–1189. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Ji, P.; Pang, J.; Zhong, Z.; Li, H.; He, C.; Zhang, J.; Zhao, C. Clinical characteristics of 3,062 COVID-19 patients: A meta-analysis. J. Med. Virol. 2020, 92, 1902–1914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whitaker, M.; Elliott, J.; Chadeau-Hyam, M.; Riley, S.; Darzi, A.; Cooke, G.; Ward, H.; Elliott, P. Persistent COVID-19 symptoms in a community study of 606,434 people in England. Nat. Commun. 2022, 13, 1957. [Google Scholar] [CrossRef] [PubMed]
- Evans, R.A.; McAuley, H.; Harrison, E.M.; Shikotra, A.; Singapuri, A.; Sereno, M.; Elneima, O.; Docherty, A.B.; Lone, N.I.; Leavy, O.C.; et al. Physical, cognitive, and mental health impacts of COVID-19 after hospitalisation (PHOSP-COVID): A UK multicentre, prospective cohort study. Lancet Respir. Med. 2021, 9, 1275–1287. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Penn, R.; Barclay, W.S.; Giotis, E.S. Naive human macrophages are refractory to SARS-CoV-2 infection and exhibit a modest inflammatory response early in infection. Viruses 2022, 14, 441. [Google Scholar] [CrossRef] [PubMed]
- Montazersaheb, S.; Hosseiniyan Khatibi, S.M.; Hejazi, M.S.; Tarhriz, V.; Farjami, A.; Ghasemian Sorbeni, F.; Farahzadi, R.; Ghasemnejad, T. COVID-19 infection: An overview on cytokine storm and related interventions. Virol. J. 2022, 19, 92. [Google Scholar] [CrossRef]
- Henderson, L.A.; Canna, S.W.; Schulert, G.S.; Volpi, S.; Lee, P.Y.; Kernan, K.F.; Caricchio, R.; Mahmud, S.; Hazen, M.M.; Halyabar, O.; et al. On the alert for cytokine storm: Immunopathology in COVID-19. Arthritis Rheumatol. 2020, 72, 1059–1063. [Google Scholar] [CrossRef] [Green Version]
- Elrobaa, I.H.; New, K.J. COVID-19: Pulmonary and extra pulmonary manifestations. Front. Public Health 2021, 9, 711616. [Google Scholar] [CrossRef]
- Lee, M.H.; Perl, D.P.; Steiner, J.; Pasternack, N.; Li, W.; Maric, D.; Safavi, F.; Horkayne-Szakaly, I.; Jones, R.; Stram, M.N.; et al. neurovascular injury with complement activation and inflammation in COVID-19. Brain 2022, 145, 2555–2568. [Google Scholar] [CrossRef]
- Greenhalgh, T.; Sivan, M.; Delaney, B.; Evans, R.; Milne, R. Long COVID-an update for primary care. BMJ 2022, 378, e072117. [Google Scholar] [CrossRef] [PubMed]
- Brightling, C.E.; Evans, R.A. Long COVID: Which symptoms can be attributed to SARS-CoV-2 infection? Lancet 2022, 400, 411–413. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Li, X.; Gu, X.; Zhang, H.; Ren, L.; Guo, L.; Liu, M.; Wang, Y.; Cui, D.; Wang, Y.; et al. Health outcomes in people 2 years after surviving hospitalisation with COVID-19: A longitudinal cohort study. Lancet Respir. Med. 2022, 10, 863–876. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Yang, L. Post-acute sequelae of SARS-CoV-2 infection: A neglected public health issue. Front. Public Health 2022, 10, 908757. [Google Scholar] [CrossRef] [PubMed]
- Feikin, D.R.; Abu-Raddad, L.J.; Andrews, N.; Davies, M.A.; Higdon, M.M.; Orenstein, W.A.; Patel, M.K. Assessing vaccine effectiveness against severe COVID-19 disease caused by omicron variant. Report from a meeting of the World Health Organization. Vaccine 2022, 40, 3516–3527. [Google Scholar] [CrossRef]
- Andrews, N.; Tessier, E.; Stowe, J.; Gower, C.; Kirsebom, F.; Simmons, R.; Gallagher, E.; Thelwall, S.; Groves, N.; Dabrera, G.; et al. Duration of protection against mild and severe disease by COVID-19 Vaccines. N. Engl. J. Med. 2022, 386, 340–350. [Google Scholar] [CrossRef]
- Shrestha, L.B.; Foster, C.; Rawlinson, W.; Tedla, N.; Bull, R.A. Evolution of the SARS-CoV-2 omicron variants BA.1 to BA.5: Implications for immune escape and transmission. Rev. Med. Virol. 2022, 32, e2381. [Google Scholar] [CrossRef]
- Joshi, N.; Tyagi, A.; Nigam, S. Molecular Level dissection of critical spike mutations in SARS-CoV-2 variants of concern (VOCs): A simplified review. ChemistrySelect 2021, 6, 7981–7998. [Google Scholar] [CrossRef]
- Tao, K.; Tzou, P.L.; Nouhin, J.; Gupta, R.K.; de Oliveira, T.; Kosakovsky Pond, S.L.; Fera, D.; Shafer, R.W. The biological and clinical significance of emerging SARS-CoV-2 variants. Nat. Rev. Genet. 2021, 22, 757–773. [Google Scholar] [CrossRef]
- Plante, J.A.; Mitchell, B.M.; Plante, K.S.; Debbink, K.; Weaver, S.C.; Menachery, V.D. The variant gambit: COVID-19’s next move. Cell Host Microbe 2021, 29, 508–515. [Google Scholar] [CrossRef]
- Tegally, H.; Moir, M.; Everatt, J.; Giovanetti, M.; Scheepers, C.; Wilkinson, E.; Subramoney, K.; Makatini, Z.; Moyo, S.; Amoako, D.G.; et al. Emergence of SARS-CoV-2 omicron lineages BA.4 and BA.5 in South Africa. Nat. Med. 2022, 28, 1785–1790. [Google Scholar] [CrossRef] [PubMed]
- Tallei, T.E.; Alhumaid, S.; AlMusa, Z.; Fatimawali; Kusumawaty, D.; Alynbiawi, A.; Alshukairi, A.N.; Rabaan, A.A. Update on the omicron sub-variants BA.4 and BA.5. Rev. Med. Virol. 2022, 26, e2391. [Google Scholar] [CrossRef] [PubMed]
- Thompson, M.G.; Natarajan, K.; Irving, S.A.; Rowley, E.A.; Griggs, E.P.; Gaglani, M.; Klein, N.P.; Grannis, S.J.; DeSilva, M.B.; Stenehjem, E.; et al. Effectiveness of a third dose of mRNA vaccines against COVID-19-associated emergency department and urgent care encounters and hospitalizations among adults during periods of Delta and Omicron variant predominance-VISION Network, 10 States, August 2021–January 2022. MMWR Morb. Mortal. Wkly. Rep. 2022, 71, 139–145. [Google Scholar] [PubMed]
- Khoury, D.S.; Docken, S.S.; Subbarao, K.; Kent, S.J.; Davenport, M.P.; Cromer, D. Predicting the efficacy of variant-modified COVID-19 vaccine boosters. bioRxiv 2022. [Google Scholar] [CrossRef]
- Weinreich, D.M.; Sivapalasingam, S.; Norton, T.; Ali, S.; Gao, H.; Bhore, R.; Musser, B.J.; Soo, Y.; Rofail, D.; Im, J.; et al. REGN-COV2, a neutralizing antibody cocktail, in outpatients with Covid-19. N. Engl. J. Med. 2021, 384, 238–251. [Google Scholar] [CrossRef]
- Group, R.C.; Horby, P.; Lim, W.S.; Emberson, J.R.; Mafham, M.; Bell, J.L.; Linsell, L.; Staplin, N.; Brightling, C.; Ustianowski, A.; et al. Dexamethasone in hospitalized patients with COVID-19. N. Engl. J. Med. 2021, 384, 693–704. [Google Scholar]
- Beigel, J.H.; Tomashek, K.M.; Dodd, L.E.; Mehta, A.K.; Zingman, B.S.; Kalil, A.C.; Hohmann, E.; Chu, H.Y.; Luetkemeyer, A.; Kline, S.; et al. Remdesivir for the treatment of COVID-19-Final report. N. Engl. J. Med. 2020, 383, 1813–1826. [Google Scholar] [CrossRef]
- Vangeel, L.; Chiu, W.; De Jonghe, S.; Maes, P.; Slechten, B.; Raymenants, J.; Andre, E.; Leyssen, P.; Neyts, J.; Jochmans, D. Remdesivir, molnupiravir and nirmatrelvir remain active against SARS-CoV-2 omicron and other variants of concern. Antiviral Res. 2022, 198, 105252. [Google Scholar] [CrossRef]
- WHO Solidarity Trial Consortium. Remdesivir and three other drugs for hospitalised patients with COVID-19: Final results of the WHO Solidarity randomised trial and updated meta-analyses. Lancet 2022, 399, 1941–1953. [Google Scholar] [CrossRef]
- Wang, L.; Berger, N.A.; Davis, P.B.; Kaelber, D.C.; Volkow, N.D.; Xu, R. COVID-19 rebound after paxlovid and molnupiravir during January–June 2022. medRxiv 2022. [Google Scholar] [CrossRef]
- Rubin, R. From positive to negative to positive again-The mystery of why COVID-19 rebounds in some Patients who take paxlovid. JAMA 2022, 327, 2380–2382. [Google Scholar] [CrossRef] [PubMed]
- Li, L.Q.; Huang, T.; Wang, Y.Q.; Wang, Z.P.; Liang, Y.; Huang, T.B.; Zhang, H.Y.; Sun, W.; Wang, Y. COVID-19 patients’ clinical characteristics, discharge rate, and fatality rate of meta-analysis. J. Med. Virol. 2020, 92, 577–583. [Google Scholar] [CrossRef] [PubMed]
- Ioannidis, J.P.A.; Axfors, C.; Contopoulos-Ioannidis, D.G. Population-level COVID-19 mortality risk for non-elderly individuals overall and for non-elderly individuals without underlying diseases in pandemic epicenters. Environ. Res. 2020, 188, 109890. [Google Scholar] [CrossRef] [PubMed]
- Williamson, E.J.; Walker, A.J.; Bhaskaran, K.; Bacon, S.; Bates, C.; Morton, C.E.; Curtis, H.J.; Mehrkar, A.; Evans, D.; Inglesby, P.; et al. Factors associated with COVID-19-related death using OpenSAFELY. Nature 2020, 584, 430–436. [Google Scholar] [CrossRef] [PubMed]
- Fauci, A.S.; Lane, H.C.; Redfield, R.R. Covid-19-Navigating the uncharted. N. Engl. J. Med. 2020, 382, 1268–1269. [Google Scholar] [CrossRef]
- Zhang, J.J.; Dong, X.; Liu, G.H.; Gao, Y.D. Risk and protective factors for COVID-19 morbidity, severity, and mortality. Clin. Rev. Allergy Immunol. 2022, 19, 1–18. [Google Scholar] [CrossRef]
- Antonelli, M.; Pujol, J.C.; Spector, T.D.; Ourselin, S.; Steves, C.J. Risk of long COVID associated with delta versus omicron variants of SARS-CoV-2. Lancet 2022, 399, 2263–2264. [Google Scholar] [CrossRef]
- Antonelli, M.; Penfold, R.S.; Merino, J.; Sudre, C.H.; Molteni, E.; Berry, S.; Canas, L.S.; Graham, M.S.; Klaser, K.; Modat, M.; et al. Risk factors and disease profile of post-vaccination SARS-CoV-2 infection in UK users of the COVID Symptom Study app: A prospective, community-based, nested, case-control study. Lancet Infect Dis. 2022, 22, 43–55. [Google Scholar] [CrossRef]
- Fernandez-de-Las-Penas, C.; de-la-Llave-Rincon, A.I.; Ortega-Santiago, R.; Ambite-Quesada, S.; Gomez-Mayordomo, V.; Cuadrado, M.L.; Arias-Navalon, J.A.; Hernandez-Barrera, V.; Martin-Guerrero, J.D.; Pellicer-Valero, O.J.; et al. Prevalence and risk factors of musculoskeletal pain symptoms as long-term post-COVID sequelae in hospitalized COVID-19 survivors: A multicenter study. Pain 2022, 163, e989–e996. [Google Scholar] [CrossRef]
- Wenham, C.; Smith, J.; Morgan, R.; on behalf of the Gender and COVID-19 Working Group. COVID-19: The gendered impacts of the outbreak. Lancet 2020, 395, 846–848. [Google Scholar] [CrossRef] [Green Version]
- Stasi, V.D.; Rastrelli, G. The role of sex hormones in the disparity of COVID-19 outcomes based on gender. J. Sex Med. 2021, 18, 1950–1954. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.M.; Bai, P.; He, W.; Wu, F.; Liu, X.F.; Han, D.M.; Liu, S.; Yang, J.K. Gender differences in patients with COVID-19: Focus on severity and mortality. Front. Public Health 2020, 8, 152. [Google Scholar] [CrossRef] [PubMed]
- Xu, K.; Chen, Y.; Yuan, J.; Yi, P.; Ding, C.; Wu, W.; Li, Y.; Ni, Q.; Zou, R.; Li, X.; et al. Factors associated with prolonged viral RNA shedding in patients with coronavirus disease 2019 (COVID-19). Clin. Infect Dis. 2020, 71, 799–806. [Google Scholar] [CrossRef] [PubMed]
- Chakravarty, D.; Nair, S.S.; Hammouda, N.; Ratnani, P.; Gharib, Y.; Wagaskar, V.; Mohamed, N.; Lundon, D.; Dovey, Z.; Kyprianou, N.; et al. Sex differences in SARS-CoV-2 infection rates and the potential link to prostate cancer. Commun. Biol. 2020, 3, 374. [Google Scholar] [CrossRef] [PubMed]
- Klein, S.L.; Huber, S. Sex differences in susceptibility to viral infection. In Sex Hormones and Immunity to Infection; Klein, S.L., Roberts, C., Eds.; Springer: Berlin, Germany, 2010; pp. 93–122. [Google Scholar]
- Klein, S.L.; Hodgson, A.; Robinson, D.P. Mechanisms of sex disparities in influenza pathogenesis. J. Leukoc. Biol. 2012, 92, 67–73. [Google Scholar] [CrossRef] [Green Version]
- Klein, S.L. Sex differences in prophylaxis and therapeutic treatments for viral diseases. Handb. Exp. Pharmacol. 2012, 499–522. [Google Scholar]
- Klein, S.L.; Flanagan, K.L. Sex differences in immune responses. Nat. Rev. Immunol. 2016, 16, 626–638. [Google Scholar] [CrossRef]
- Liao, Z.H.; Huang, T.; Xiao, J.W.; Gu, R.C.; Ouyang, J.; Wu, G.; Liao, H. Estrogen signaling effects on muscle-specific immune responses through controlling the recruitment and function of macrophages and T cells. Skelet Muscle 2019, 9, 20. [Google Scholar] [CrossRef]
- Al-Attar, A.; Presnell, S.R.; Peterson, C.A.; Thomas, D.T.; Lutz, C.T. The effect of sex on immune cells in healthy aging: Elderly women have more robust natural killer lymphocytes than do elderly men. Mech. Ageing Dev. 2016, 156, 25–33. [Google Scholar] [CrossRef]
- Fish, E.N. The X-files in immunity: Sex-based differences predispose immune responses. Nat. Rev. Immunol. 2008, 8, 737–744. [Google Scholar] [CrossRef]
- Libert, C.; Dejager, L.; Pinheiro, I. The X chromosome in immune functions: When a chromosome makes the difference. Nat. Rev. Immunol. 2010, 10, 594–604. [Google Scholar] [CrossRef] [PubMed]
- vom Steeg, L.G.; Klein, S.L. Sex matters in infectious disease pathogenesis. PLoS Pathog. 2016, 12, e1005374. [Google Scholar] [CrossRef] [PubMed]
- Oertelt-Prigione, S. The influence of sex and gender on the immune response. Autoimmun. Rev. 2012, 11, A479–A485. [Google Scholar] [CrossRef]
- Straub, R.H. The complex role of estrogens in inflammation. Endocr. Rev. 2007, 28, 521–574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilliver, S.C. Sex steroids as inflammatory regulators. J. Steroid. Biochem. Mol. Biol. 2010, 120, 105–115. [Google Scholar] [CrossRef]
- Khan, D.; Ansar Ahmed, S. The immune System is a natural target for estrogen action: Opposing effects of estrogen in two prototypical autoimmune diseases. Front. Immunol. 2015, 6, 635. [Google Scholar] [CrossRef] [Green Version]
- Foo, Y.Z.; Nakagawa, S.; Rhodes, G.; Simmons, L.W. The effects of sex hormones on immune function: A meta-analysis. Biol. Rev. Camb. Philos. Soc. 2017, 92, 551–571. [Google Scholar] [CrossRef] [Green Version]
- Taneja, V. Sex hormones determine immune response. Front. Immunol. 2018, 9, 1931. [Google Scholar] [CrossRef]
- Bouman, A.; Heineman, M.J.; Faas, M.M. Sex hormones and the immune response in humans. Hum. Reprod. Update 2005, 11, 411–423. [Google Scholar] [CrossRef] [Green Version]
- Ruggieri, A.; Gagliardi, M.C.; Anticoli, S. Sex-dependent outcome of Hepatitis B and C viruses infections: Synergy of sex hormones and immune responses? Front. Immunol. 2018, 9, 2302. [Google Scholar] [CrossRef] [Green Version]
- Tian, Y.; Kuo, C.F.; Chen, W.L.; Ou, J.H. Enhancement of hepatitis B virus replication by androgen and its receptor in mice. J. Virol. 2012, 86, 1904–1910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jackson, C.B.; Farzan, M.; Chen, B.; Choe, H. Mechanisms of SARS-CoV-2 entry into cells. Nat. Rev. Mol. Cell Biol. 2022, 23, 3–20. [Google Scholar] [CrossRef] [PubMed]
- Yadav, R.; Chaudhary, J.K.; Jain, N.; Chaudhary, P.K.; Khanra, S.; Dhamija, P.; Sharma, A.; Kumar, A.; Handu, S. Role of structural and non-structural proteins and therapeutic targets of SARS-CoV-2 for COVID-19. Cells 2021, 10, 821. [Google Scholar] [CrossRef] [PubMed]
- Bayati, A.; Kumar, R.; Francis, V.; McPherson, P.S. SARS-CoV-2 infects cells after viral entry via clathrin-mediated endocytosis. J. Biol. Chem. 2021, 296, 100306. [Google Scholar] [CrossRef]
- Shang, J.; Wan, Y.; Luo, C.; Ye, G.; Geng, Q.; Auerbach, A.; Li, F. Cell entry mechanisms of SARS-CoV-2. Proc. Natl. Acad. Sci. USA 2020, 117, 11727–11734. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, M.; Kleine-Weber, H.; Schroeder, S.; Kruger, N.; Herrler, T.; Erichsen, S.; Schiergens, T.S.; Herrler, G.; Wu, N.H.; Nitsche, A.; et al. SARS-CoV-2 cell entry depends on ACE2 and TMPRSS2 and is blocked by a clinically proven protease inhibitor. Cell 2020, 181, 271–280. [Google Scholar] [CrossRef]
- Jia, H. Pulmonary angiotensin-converting enzyme 2 (ACE2) and inflammatory lung disease. Shock 2016, 46, 239–248. [Google Scholar] [CrossRef]
- Giotis, E.S.; Carnell, G.; Young, E.F.; Ghanny, S.; Soteropoulos, P.; Wang, L.F.; Barclay, W.S.; Skinner, M.A.; Temperton, N. Entry of the bat influenza H17N10 virus into mammalian cells is enabled by the MHC class II HLA-DR receptor. Nat. Microbiol. 2019, 4, 2035–2038. [Google Scholar] [CrossRef] [Green Version]
- Long, J.S.; Mistry, B.; Haslam, S.M.; Barclay, W.S. Host and viral determinants of influenza A virus species specificity. Nat. Rev. Microbiol. 2019, 17, 67–81. [Google Scholar] [CrossRef]
- Matsuyama, S.; Nagata, N.; Shirato, K.; Kawase, M.; Takeda, M.; Taguchi, F. Efficient activation of the severe acute respiratory syndrome coronavirus spike protein by the transmembrane protease TMPRSS2. J. Virol. 2010, 84, 12658–12664. [Google Scholar] [CrossRef] [Green Version]
- Shulla, A.; Heald-Sargent, T.; Subramanya, G.; Zhao, J.; Perlman, S.; Gallagher, T. A transmembrane serine protease is linked to the severe acute respiratory syndrome coronavirus receptor and activates virus entry. J. Virol. 2011, 85, 873–882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belouzard, S.; Millet, J.K.; Licitra, B.N.; Whittaker, G.R. Mechanisms of coronavirus cell entry mediated by the viral spike protein. Viruses 2012, 4, 1011–1033. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, I.M.; Abdelmalek, D.H.; Elshahat, M.E.; Elfiky, A.A. COVID-19 spike-host cell receptor GRP78 binding site prediction. J. Infect. 2020, 80, 554–562. [Google Scholar] [CrossRef] [PubMed]
- Lin, B.; Ferguson, C.; White, J.T.; Wang, S.; Vessella, R.; True, L.D.; Hood, L.; Nelson, P.S. Prostate-localized and androgen-regulated expression of the membrane-bound serine protease TMPRSS2. Cancer Res. 1999, 59, 4180–4184. [Google Scholar] [PubMed]
- Lucas, J.M.; Heinlein, C.; Kim, T.; Hernandez, S.A.; Malik, M.S.; True, L.D.; Morrissey, C.; Corey, E.; Montgomery, B.; Mostaghel, E.; et al. The androgen-regulated protease TMPRSS2 activates a proteolytic cascade involving components of the tumor microenvironment and promotes prostate cancer metastasis. Cancer Discov. 2014, 4, 1310–1325. [Google Scholar] [CrossRef] [Green Version]
- Katopodis, P.; Kerslake, R.; Davies, J.; Randeva, H.S.; Chatha, K.; Hall, M.; Spandidos, D.A.; Anikin, V.; Polychronis, A.; Robertus, J.L.; et al. COVID-19 and SARS-CoV-2 host cell entry mediators: Expression profiling of TMRSS4 in health and disease. Int. J. Mol. Med. 2021, 47, 64. [Google Scholar] [CrossRef] [PubMed]
- Lukassen, S.; Chua, R.L.; Trefzer, T.; Kahn, N.C.; Schneider, M.A.; Muley, T.; Winter, H.; Meister, M.; Veith, C.; Boots, A.W.; et al. SARS-CoV-2 receptor ACE2 and TMPRSS2 are primarily expressed in bronchial transient secretory cells. EMBO J. 2020, 39, e105114. [Google Scholar] [CrossRef]
- Kumar, A.; Faiq, M.A.; Pareek, V.; Raza, K.; Narayan, R.K.; Prasoon, P.; Kumar, P.; Kulandhasamy, M.; Kumari, C.; Kant, K.; et al. Relevance of SARS-CoV-2 related factors ACE2 and TMPRSS2 expressions in gastrointestinal tissue with pathogenesis of digestive symptoms, diabetes-associated mortality, and disease recurrence in COVID-19 patients. Med. Hypotheses 2020, 144, 110271. [Google Scholar] [CrossRef]
- Collin, J.; Queen, R.; Zerti, D.; Dorgau, B.; Georgiou, M.; Djidrovski, I.; Hussain, R.; Coxhead, J.M.; Joseph, A.; Rooney, P.; et al. Co-expression of SARS-CoV-2 entry genes in the superficial adult human conjunctival, limbal and corneal epithelium suggests an additional route of entry via the ocular surface. Ocul. Surf. 2021, 19, 190–200. [Google Scholar] [CrossRef]
- Zou, X.; Chen, K.; Zou, J.; Han, P.; Hao, J.; Han, Z. Single-cell RNA-seq data analysis on the receptor ACE2 expression reveals the potential risk of different human organs vulnerable to 2019-nCoV infection. Front. Med. 2020, 14, 185–192. [Google Scholar] [CrossRef] [Green Version]
- Vaarala, M.H.; Porvari, K.S.; Kellokumpu, S.; Kyllonen, A.P.; Vihko, P.T. Expression of transmembrane serine protease TMPRSS2 in mouse and human tissues. J. Pathol. 2001, 193, 134–140. [Google Scholar] [CrossRef] [PubMed]
- de Kloet, A.D.; Krause, E.G.; Woods, S.C. The renin angiotensin system and the metabolic syndrome. Physiol. Behav. 2010, 100, 525–534. [Google Scholar] [CrossRef] [PubMed]
- Samavati, L.; Uhal, B.D. ACE2, much more than just a receptor for SARS-COV-2. Front. Cell Infect. Microbiol. 2020, 10, 317. [Google Scholar] [CrossRef] [PubMed]
- Shen, L.W.; Qian, M.Q.; Yu, K.; Narva, S.; Yu, F.; Wu, Y.L.; Zhang, W. Inhibition of Influenza A virus propagation by benzoselenoxanthenes stabilizing TMPRSS2 Gene G-quadruplex and hence down-regulating TMPRSS2 expression. Sci. Rep. 2020, 10, 7635. [Google Scholar] [CrossRef] [PubMed]
- Esumi, M.; Ishibashi, M.; Yamaguchi, H.; Nakajima, S.; Tai, Y.; Kikuta, S.; Sugitani, M.; Takayama, T.; Tahara, M.; Takeda, M.; et al. Transmembrane serine protease TMPRSS2 activates hepatitis C virus infection. Hepatology 2015, 61, 437–446. [Google Scholar] [CrossRef]
- Shirato, K.; Kawase, M.; Matsuyama, S. Middle East respiratory syndrome coronavirus infection mediated by the transmembrane serine protease TMPRSS2. J. Virol. 2013, 87, 12552–12561. [Google Scholar] [CrossRef] [Green Version]
- Gierer, S.; Bertram, S.; Kaup, F.; Wrensch, F.; Heurich, A.; Kramer-Kuhl, A.; Welsch, K.; Winkler, M.; Meyer, B.; Drosten, C.; et al. The spike protein of the emerging betacoronavirus EMC uses a novel coronavirus receptor for entry, can be activated by TMPRSS2, and is targeted by neutralizing antibodies. J. Virol. 2013, 87, 5502–5511. [Google Scholar] [CrossRef] [Green Version]
- Limburg, H.; Harbig, A.; Bestle, D.; Stein, D.A.; Moulton, H.M.; Jaeger, J.; Janga, H.; Hardes, K.; Koepke, J.; Schulte, L.; et al. TMPRSS2 is the major activating protease of influenza A virus in primary human airway cells and influenza B virus in human type II pneumocytes. J. Virol. 2019, 93, e00649-19. [Google Scholar] [CrossRef] [Green Version]
- Iwata-Yoshikawa, N.; Okamura, T.; Shimizu, Y.; Hasegawa, H.; Takeda, M.; Nagata, N. TMPRSS2 contributes to virus spread and immunopathology in the airways of murine models after coronavirus infection. J. Virol. 2019, 93, e01815-18. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Jiang, Q.; Xia, X.; Liu, K.; Yu, Z.; Tao, W.; Gong, W.; Han, J.J. Individual variation of the SARS-CoV-2 receptor ACE2 gene expression and regulation. Aging Cell 2020, 19, e13168. [Google Scholar] [CrossRef]
- Singh, H.; Choudhari, R.; Nema, V.; Khan, A.A. ACE2 and TMPRSS2 polymorphisms in various diseases with special reference to its impact on COVID-19 disease. Microb. Pathog. 2021, 150, 104621. [Google Scholar] [CrossRef] [PubMed]
- Calcagnile, M.; Forgez, P.; Iannelli, A.; Bucci, C.; Alifano, M.; Alifano, P. Molecular docking simulation reveals ACE2 polymorphisms that may increase the affinity of ACE2 with the SARS-CoV-2 spike protein. Biochimie 2021, 180, 143–148. [Google Scholar] [CrossRef] [PubMed]
- Irham, L.M.; Chou, W.H.; Calkins, M.J.; Adikusuma, W.; Hsieh, S.L.; Chang, W.C. Genetic variants that influence SARS-CoV-2 receptor TMPRSS2 expression among population cohorts from multiple continents. Biochem. Biophys. Res. Commun. 2020, 529, 263–269. [Google Scholar] [CrossRef] [PubMed]
- Saheb Sharif-Askari, N.; Saheb Sharif-Askari, F.; Alabed, M.; Temsah, M.H.; Al Heialy, S.; Hamid, Q.; Halwani, R. Airways expression of SARS-CoV-2 receptor, ACE2, and TMPRSS2 is lower in children than adults and increases with smoking and COPD. Mol. Ther. Methods Clin. Dev. 2020, 18, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Wang, A.; Chiou, J.; Poirion, O.B.; Buchanan, J.; Valdez, M.J.; Verheyden, J.M.; Hou, X.; Kudtarkar, P.; Narendra, S.; Newsome, J.M.; et al. Single-cell multiomic profiling of human lungs reveals cell-type-specific and age-dynamic control of SARS-CoV-2 host genes. Elife 2020, 9, e62522. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Yang, L.; Zhao, X.E. Co-crystallization and structure determination: An effective direction for anti-SARS-CoV-2 drug discovery. Comput. Struct. Biotechnol. J. 2021, 19, 4684–4701. [Google Scholar] [CrossRef]
- Mackin, R.T.; Edwards, J.V.; Atuk, E.B.; Beltrami, N.; Condon, B.D.; Jayawickramarajah, J.; French, A.D. Structure/function analysis of truncated amino-terminal ACE2 peptide analogs that bind to SARS-CoV-2 spike glycoprotein. Molecules 2022, 27, 2070. [Google Scholar] [CrossRef]
- Monteil, V.; Dyczynski, M.; Lauschke, V.M.; Kwon, H.; Wirnsberger, G.; Youhanna, S.; Zhang, H.; Slutsky, A.S.; Hurtado Del Pozo, C.; Horn, M.; et al. Human soluble ACE2 improves the effect of remdesivir in SARS-CoV-2 infection. EMBO Mol. Med. 2021, 13, e13426. [Google Scholar] [CrossRef]
- Jia, H.; Neptune, E.; Cui, H. Targeting ACE2 for COVID-19 Therapy: Opportunities and challenges. Am. J. Respir. Cell Mol. Biol. 2021, 64, 416–425. [Google Scholar] [CrossRef]
- Lelis, D.F.; Freitas, D.F.; Machado, A.S.; Crespo, T.S.; Santos, S.H.S. Angiotensin-(1-7), adipokines and inflammation. Metabolism 2019, 95, 36–45. [Google Scholar] [CrossRef]
- Zhang, H.; Penninger, J.M.; Li, Y.; Zhong, N.; Slutsky, A.S. Angiotensin-converting enzyme 2 (ACE2) as a SARS-CoV-2 receptor: Molecular mechanisms and potential therapeutic target. Intensive Care Med. 2020, 46, 586–590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Xie, B.; Hashimoto, K. Current status of potential therapeutic candidates for the COVID-19 crisis. Brain Behav. Immun. 2020, 87, 59–73. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.S.; Heinlein, C.; Hackman, R.C.; Nelson, P.S. Phenotypic analysis of mice lacking the TMPRSS2-encoded protease. Mol. Cell Biol. 2006, 26, 965–975. [Google Scholar] [CrossRef] [Green Version]
- Stopsack, K.H.; Mucci, L.A.; Antonarakis, E.S.; Nelson, P.S.; Kantoff, P.W. TMPRSS2 and COVID-19: Serendipity or opportunity for intervention? Cancer Discov. 2020, 10, 779–782. [Google Scholar] [CrossRef] [Green Version]
- Bestle, D.; Heindl, M.R.; Limburg, H.; Van Lam van, T.; Pilgram, O.; Moulton, H.; Stein, D.A.; Hardes, K.; Eickmann, M.; Dolnik, O.; et al. TMPRSS2 and furin are both essential for proteolytic activation of SARS-CoV-2 in human airway cells. Life Sci. Alliance 2020, 3, e202000786. [Google Scholar] [CrossRef] [PubMed]
- Gunst, J.D.; Staerke, N.B.; Pahus, M.H.; Kristensen, L.H.; Bodilsen, J.; Lohse, N.; Dalgaard, L.S.; Bronnum, D.; Frobert, O.; Honge, B.; et al. Efficacy of the TMPRSS2 inhibitor camostat mesilate in patients hospitalized with COVID-19- A double-blind randomized controlled trial. EClinicalMedicine 2021, 35, 100849. [Google Scholar] [CrossRef]
- Shapira, T.; Monreal, I.A.; Dion, S.P.; Buchholz, D.W.; Imbiakha, B.; Olmstead, A.D.; Jager, M.; Desilets, A.; Gao, G.; Martins, M.; et al. A TMPRSS2 inhibitor acts as a pan-SARS-CoV-2 prophylactic and therapeutic. Nature 2022, 605, 340–348. [Google Scholar] [CrossRef]
- Mahoney, M.; Damalanka, V.C.; Tartell, M.A.; Chung, D.H.; Lourenco, A.L.; Pwee, D.; Mayer Bridwell, A.E.; Hoffmann, M.; Voss, J.; Karmakar, P.; et al. A novel class of TMPRSS2 inhibitors potently block SARS-CoV-2 and MERS-CoV viral entry and protect human epithelial lung cells. Proc. Natl. Acad. Sci. USA 2021, 118, e2108728118. [Google Scholar] [CrossRef]
- Rahman, N.; Basharat, Z.; Yousuf, M.; Castaldo, G.; Rastrelli, L.; Khan, H. Virtual screening of natural products against Type II transmembrane serine protease (TMPRSS2), the priming agent of coronavirus 2 (SARS-CoV-2). Molecules 2020, 25, 2271. [Google Scholar] [CrossRef]
- Rahbar Saadat, Y.; Hosseiniyan Khatibi, S.M.; Zununi Vahed, S.; Ardalan, M. Host serine proteases: A potential targeted therapy for COVID-19 and influenza. Front. Mol. Biosci. 2021, 8, 725528. [Google Scholar] [CrossRef]
- Hu, X.; Shrimp, J.H.; Guo, H.; Xu, M.; Chen, C.Z.; Zhu, W.; Zakharov, A.V.; Jain, S.; Shinn, P.; Simeonov, A.; et al. Discovery of TMPRSS2 inhibitors from virtual screening as a potential treatment of COVID-19. ACS Pharmacol. Transl. Sci. 2021, 4, 1124–1135. [Google Scholar] [CrossRef] [PubMed]
- Alzain, A.A.; Elbadwi, F.A.; Alsamani, F.O. Discovery of novel TMPRSS2 inhibitors for COVID-19 using in silico fragment-based drug design, molecular docking, molecular dynamics, and quantum mechanics studies. Inform. Med. Unlocked 2022, 29, 100870. [Google Scholar] [CrossRef] [PubMed]
- Mahgoub, M.A.; Alnaem, A.; Fadlelmola, M.; Abo-Idris, M.; Makki, A.A.; Abdelgadir, A.A.; Alzain, A.A. Discovery of novel potential inhibitors of TMPRSS2 and Mpro of SARS-CoV-2 using E-pharmacophore and docking-based virtual screening combined with molecular dynamic and quantum mechanics. J. Biomol. Struct. Dyn. 2022, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Manandhar, S.; Pai, K.S.R.; Krishnamurthy, P.T.; Kiran, A.; Kumari, G.K. Identification, virtual screening and molecular dynamic analysis of novel TMPRSS2 inhibitors from natural compound database as potential entry-blocking agents in SARS-CoV-2 therapy. Struct. Chem. 2022, 33, 1609–1617. [Google Scholar] [CrossRef] [PubMed]
- Hofmann-Winkler, H.; Moerer, O.; Alt-Epping, S.; Brauer, A.; Buttner, B.; Muller, M.; Fricke, T.; Grundmann, J.; Harnisch, L.O.; Heise, D.; et al. Camostat mesylate may reduce severity of coronavirus disease 2019 sepsis: A first observation. Crit. Care Explor. 2020, 2, e0284. [Google Scholar] [CrossRef]
- Doi, K.; Ikeda, M.; Hayase, N.; Moriya, K.; Morimura, N.; the COVID-UTH Study Group. Nafamostat mesylate treatment in combination with favipiravir for patients critically ill with COVID-19: A case series. Crit. Care 2020, 24, 392. [Google Scholar] [CrossRef]
- Jang, S.; Rhee, J.Y. Three cases of treatment with nafamostat in elderly patients with COVID-19 pneumonia who need oxygen therapy. Int. J. Infect. Dis. 2020, 96, 500–502. [Google Scholar] [CrossRef]
- Willett, B.J.; Grove, J.; MacLean, O.A.; Wilkie, C.; De Lorenzo, G.; Furnon, W.; Cantoni, D.; Scott, S.; Logan, N.; Ashraf, S.; et al. SARS-CoV-2 Omicron is an immune escape variant with an altered cell entry pathway. Nat. Microbiol. 2022, 7, 1161–1179. [Google Scholar] [CrossRef]
- Pia, L.; Rowland-Jones, S. Omicron entry route. Nat. Rev. Immunol. 2022, 22, 144. [Google Scholar] [CrossRef]
- Peacock, T.P.; Brown, J.C.; Zhou, J.; Thakur, N.; Sukhova, K.; Newman, J.; Kugathasan, R.; Yan, A.W.C.; Furnon, W.; Lorenzo, G.D.; et al. The altered entry pathway and antigenic distance of the SARS-CoV-2 Omicron variant map to separate domains of spike protein. bioRxiv 2021. [Google Scholar] [CrossRef]
- Punjani, N.; Flannigan, R. Androgens and COVID-19: Exploring the role of testosterone replacement therapy. Int. J. Impot. Res. 2022, 34, 649–651. [Google Scholar] [CrossRef] [PubMed]
- Brooke, G.N.; Waxman, J.; Bevan, C.L. The role of androgen receptor mutations in progression of prostate cancer. In Advances in Genome Science, Changing Views on Living Organisms; Neri, C., Ed.; Bentham Science Publishers: Sharjah, United Arab Emirates, 2013; Volume 1, pp. 45–65. [Google Scholar]
- Brooke, G.N.; Bevan, C.L. The role of androgen receptor mutations in prostate cancer progression. Current Genom. 2009, 10, 18–25. [Google Scholar] [CrossRef] [PubMed]
- Bleach, R.; McIlroy, M. The divergent function of androgen receptor in breast cancer; analysis of steroid mediators and tumor intracrinology. Front. Endocrinol. 2018, 9, 594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, Q.; He, B. Androgen receptor signaling in the development of castration-resistant prostate cancer. Front. Oncol. 2019, 9, 858. [Google Scholar] [CrossRef] [Green Version]
- Dart, D.A.; Waxman, J.; Aboagye, E.O.; Bevan, C.L. Visualising androgen receptor activity in male and female mice. PLoS ONE 2013, 8, e71694. [Google Scholar] [CrossRef] [Green Version]
- Mikkonen, L.; Pihlajamaa, P.; Sahu, B.; Zhang, F.P.; Janne, O.A. Androgen receptor and androgen-dependent gene expression in lung. Mol. Cell Endocrinol. 2010, 317, 14–24. [Google Scholar] [CrossRef] [Green Version]
- Baratchian, M.; McManus, J.M.; Berk, M.P.; Nakamura, F.; Mukhopadhyay, S.; Xu, W.; Erzurum, S.; Drazba, J.; Peterson, J.; Klein, E.A.; et al. Androgen regulation of pulmonary AR, TMPRSS2 and ACE2 with implications for sex-discordant COVID-19 outcomes. Sci. Rep. 2021, 11, 11130. [Google Scholar] [CrossRef]
- Leach, D.A.; Mohr, A.; Giotis, E.S.; Cil, E.; Isac, A.M.; Yates, L.L.; Barclay, W.S.; Zwacka, R.M.; Bevan, C.L.; Brooke, G.N. The antiandrogen enzalutamide downregulates TMPRSS2 and reduces cellular entry of SARS-CoV-2 in human lung cells. Nat. Commun. 2021, 12, 4068. [Google Scholar] [CrossRef]
- McManus, J.M.; Gaston, B.; Zein, J.; Sharifi, N. Association Between Asthma and Reduced Androgen Receptor Expression in Airways. J. Endocr. Soc. 2022, 6, bvac047. [Google Scholar] [CrossRef]
- Rockx, B.; Kuiken, T.; Herfst, S.; Bestebroer, T.; Lamers, M.M.; Oude Munnink, B.B.; de Meulder, D.; van Amerongen, G.; van den Brand, J.; Okba, N.M.A.; et al. Comparative pathogenesis of COVID-19, MERS, and SARS in a nonhuman primate model. Science 2020, 368, 1012–1015. [Google Scholar] [CrossRef] [Green Version]
- Clinckemalie, L.; Spans, L.; Dubois, V.; Laurent, M.; Helsen, C.; Joniau, S.; Claessens, F. Androgen regulation of the TMPRSS2 gene and the effect of a SNP in an androgen response element. Mol. Endocrinol. 2013, 27, 2028–2040. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Han, M.; Dai, P.; Xu, W.; He, J.; Tao, X.; Wu, Y.; Tong, X.; Xia, X.; Guo, W.; et al. Distinct mechanisms for TMPRSS2 expression explain organ-specific inhibition of SARS-CoV-2 infection by enzalutamide. Nat. Commun. 2021, 12, 866. [Google Scholar] [CrossRef] [PubMed]
- Science. Wadman, M. Why Coronavirus Hits Men Harder: Sex Hormones Offer Clues. Available online: https://www.science.org/content/article/why-coronavirus-hits-men-harder-sex-hormones-offer-clues (accessed on 1 November 2022).
- Mauvais-Jarvis, F. Do anti-androgens have potential as therapeutics for COVID-19? Endocrinology 2021, 162, bqab114. [Google Scholar] [CrossRef] [PubMed]
- Yip, L.; Sinclair, R.D. Antiandrogen therapy for androgenetic alopecia. Expert Rev. Dermatol. 2014, 1, 261–269. [Google Scholar] [CrossRef]
- Lanz, C.; Bennamoun, M.; Macek, P.; Cathelineau, X.; Sanchez-Salas, R. The importance of antiandrogen in prostate cancer treatment. Ann. Transl. Med. 2019, 7 (Suppl. S8), S362. [Google Scholar] [CrossRef]
- Papadatos-Pastos, D.; Dedes, K.J.; de Bono, J.S.; Kaye, S.B. Revisiting the role of antiandrogen strategies in ovarian cancer. Oncologist 2011, 16, 1413–1421. [Google Scholar] [CrossRef] [Green Version]
- Fioretti, F.M.; Sita-Lumsden, A.; Bevan, C.L.; Brooke, G.N. Revising the role of the androgen receptor in breast cancer. J. Mol. Endocrinol. 2014, 52, R257–R265. [Google Scholar] [CrossRef]
- Anestis, A.; Zoi, I.; Papavassiliou, A.G.; Karamouzis, M.V. Androgen receptor in breast cancer-clinical and preclinical research insights. Molecules 2020, 25, 358. [Google Scholar] [CrossRef] [Green Version]
- Alpanes, M.; Fernandez-Duran, E.; Escobar-Morreale, H.F. Androgens and polycystic ovary syndrome. Expert Rev. Endocrinol. Metab. 2012, 7, 91–102. [Google Scholar] [CrossRef]
- Deng, Q.; Rasool, R.U.; Russell, R.M.; Natesan, R.; Asangani, I.A. Targeting androgen regulation of TMPRSS2 and ACE2 as a therapeutic strategy to combat COVID-19. iScience 2021, 24, 102254. [Google Scholar] [CrossRef]
- Montopoli, M.; Zumerle, S.; Vettor, R.; Rugge, M.; Zorzi, M.; Catapano, C.V.; Carbone, G.M.; Cavalli, A.; Pagano, F.; Ragazzi, E.; et al. Androgen-deprivation therapies for prostate cancer and risk of infection by SARS-CoV-2: A population-based study (N = 4532). Ann. Oncol. 2020, 31, 1040–1045. [Google Scholar] [CrossRef] [PubMed]
- Patel, V.G.; Zhong, X.; Liaw, B.; Tremblay, D.; Tsao, C.K.; Galsky, M.D.; Oh, W.K. Does androgen deprivation therapy protect against severe complications from COVID-19? Ann. Oncol. 2020, 31, 1419–1420. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.M.; Heberer, K.; Gao, A.; Becker, D.J.; Loeb, S.; Makarov, D.V.; Gulanski, B.; DuVall, S.L.; Aslan, M.; Lee, J.; et al. A population-level analysis of the protective effects of androgen deprivation therapy against COVID-19 disease incidence and severity. Front. Med. 2022, 9, 774773. [Google Scholar] [CrossRef] [PubMed]
- McCoy, J.; Cadegiani, F.A.; Wambier, C.G.; Herrera, S.; Vano-Galvan, S.; Mesinkovska, N.A.; Ramos, P.M.; Shapiro, J.; Sinclair, R.; Tosti, A.; et al. 5-alpha-reductase inhibitors are associated with reduced frequency of COVID-19 symptoms in males with androgenetic alopecia. J. Eur. Acad. Dermatol. Venereol. 2021, 35, e243–e246. [Google Scholar] [CrossRef] [PubMed]
- Welen, K.; Rosendal, E.; Gisslen, M.; Lenman, A.; Freyhult, E.; Fonseca-Rodriguez, O.; Bremell, D.; Stranne, J.; Balkhed, A.O.; Niward, K.; et al. A Phase 2 Trial of the effect of antiandrogen therapy on COVID-19 outcome: No evidence of benefit, supported by epidemiology and in vitro data. Eur. Urol. 2022, 81, 285–293. [Google Scholar] [CrossRef]
- Nickols, N.G.; Mi, Z.; DeMatt, E.; Biswas, K.; Clise, C.E.; Huggins, J.T.; Maraka, S.; Ambrogini, E.; Mirsaeidi, M.S.; Levin, E.R.; et al. Effect of androgen suppression on clinical outcomes in hospitalized men with COVID-19: The HITCH randomized clinical trial. JAMA Netw. Open 2022, 5, e227852. [Google Scholar] [CrossRef]
- Luttens, A.; Gullberg, H.; Abdurakhmanov, E.; Vo, D.D.; Akaberi, D.; Talibov, V.O.; Nekhotiaeva, N.; Vangeel, L.; De Jonghe, S.; Jochmans, D.; et al. Ultralarge virtual screening identifies SARS-CoV-2 main protease inhibitors with broad-spectrum activity against coronaviruses. J. Am. Chem. Soc. 2022, 144, 2905–2920. [Google Scholar] [CrossRef]
- Wang, Z.; Yang, L. Broad-spectrum prodrugs with anti-SARS-CoV-2 activities: Strategies, benefits, and challenges. J. Med. Virol. 2022, 94, 1373–1390. [Google Scholar] [CrossRef]
Figure 1.
Alternative entry routes of SARS-CoV-2 into target cells. Elements of the schematic are from Motifolio.
Figure 1.
Alternative entry routes of SARS-CoV-2 into target cells. Elements of the schematic are from Motifolio.

Figure 2.
Androgen receptor signalling and how androgen deprivation therapies might block viral uptake. In the absence of androgen, the AR is located in the cytoplasm bound to a heat shock protein (HSP) complex. Ligand binding promotes dissociation of this complex, nuclear localisation and dimerisation. The AR binds to androgen response elements in the regulatory regions of target genes (e.g., TMPRSS2 and ACE2) and, through the recruitment of coactivators and the basal transcriptional machinery, regulates transcription. LHRH analogues/antagonists reduce androgen production to inhibit AR signalling. Antiandrogens bind to and inhibit the AR. The downregulation of AR signalling reduces TMPRSS2 and ACE2 expression, reducing COVID-19 entry. Elements of the schematic are from Motifolio.
Figure 2.
Androgen receptor signalling and how androgen deprivation therapies might block viral uptake. In the absence of androgen, the AR is located in the cytoplasm bound to a heat shock protein (HSP) complex. Ligand binding promotes dissociation of this complex, nuclear localisation and dimerisation. The AR binds to androgen response elements in the regulatory regions of target genes (e.g., TMPRSS2 and ACE2) and, through the recruitment of coactivators and the basal transcriptional machinery, regulates transcription. LHRH analogues/antagonists reduce androgen production to inhibit AR signalling. Antiandrogens bind to and inhibit the AR. The downregulation of AR signalling reduces TMPRSS2 and ACE2 expression, reducing COVID-19 entry. Elements of the schematic are from Motifolio.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Giotis, E.S.; Cil, E.; Brooke, G.N. Use of Antiandrogens as Therapeutic Agents in COVID-19 Patients. Viruses 2022, 14, 2728. https://doi.org/10.3390/v14122728
AMA Style
Giotis ES, Cil E, Brooke GN. Use of Antiandrogens as Therapeutic Agents in COVID-19 Patients. Viruses. 2022; 14(12):2728. https://doi.org/10.3390/v14122728
Chicago/Turabian StyleGiotis, Efstathios S., Emine Cil, and Greg N. Brooke. 2022. "Use of Antiandrogens as Therapeutic Agents in COVID-19 Patients" Viruses 14, no. 12: 2728. https://doi.org/10.3390/v14122728
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.