

Re-Introduction of West Nile Virus Lineage 1 in Senegal from Europe and Subsequent Circulation in Human and Mosquito Populations between 2012 and 2021
 , ,
, ,  , , , , , ,
, , , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Human Surveillance
2.1.1. Presentation of the Syndromic Sentinel Surveillance Network in Senegal (4S Network)
2.1.2. Human Sample Collection
2.2. Mosquito Collection
2.3. Serological Diagnostic
2.4. Molecular Diagnostic
2.5. Virus Isolation and Identification
2.6. Viral Genome Sequencing
2.7. Phylogenetic Analyses
3. Results
3.1. Human Samples
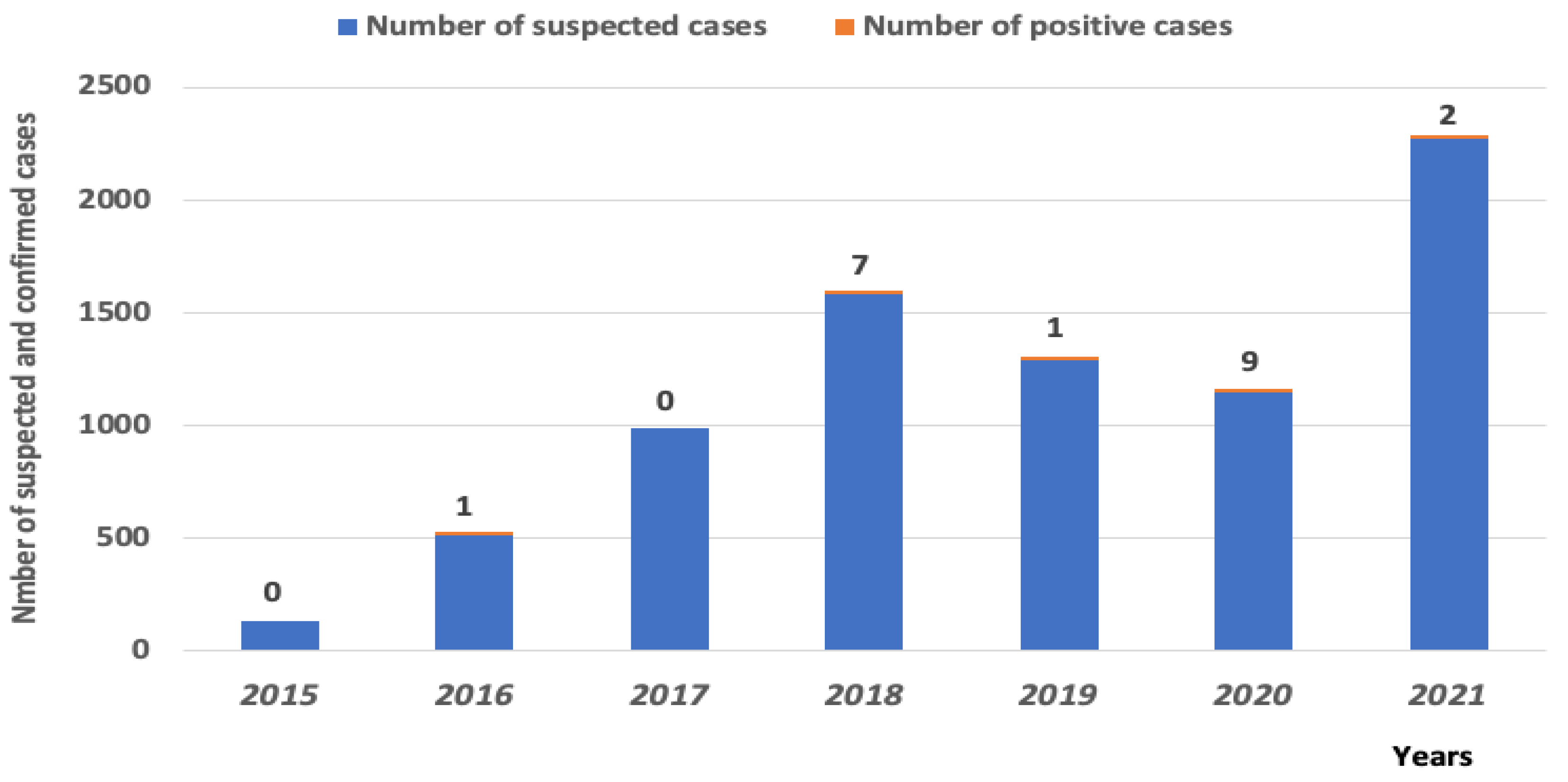
3.1.1. WNV Detection
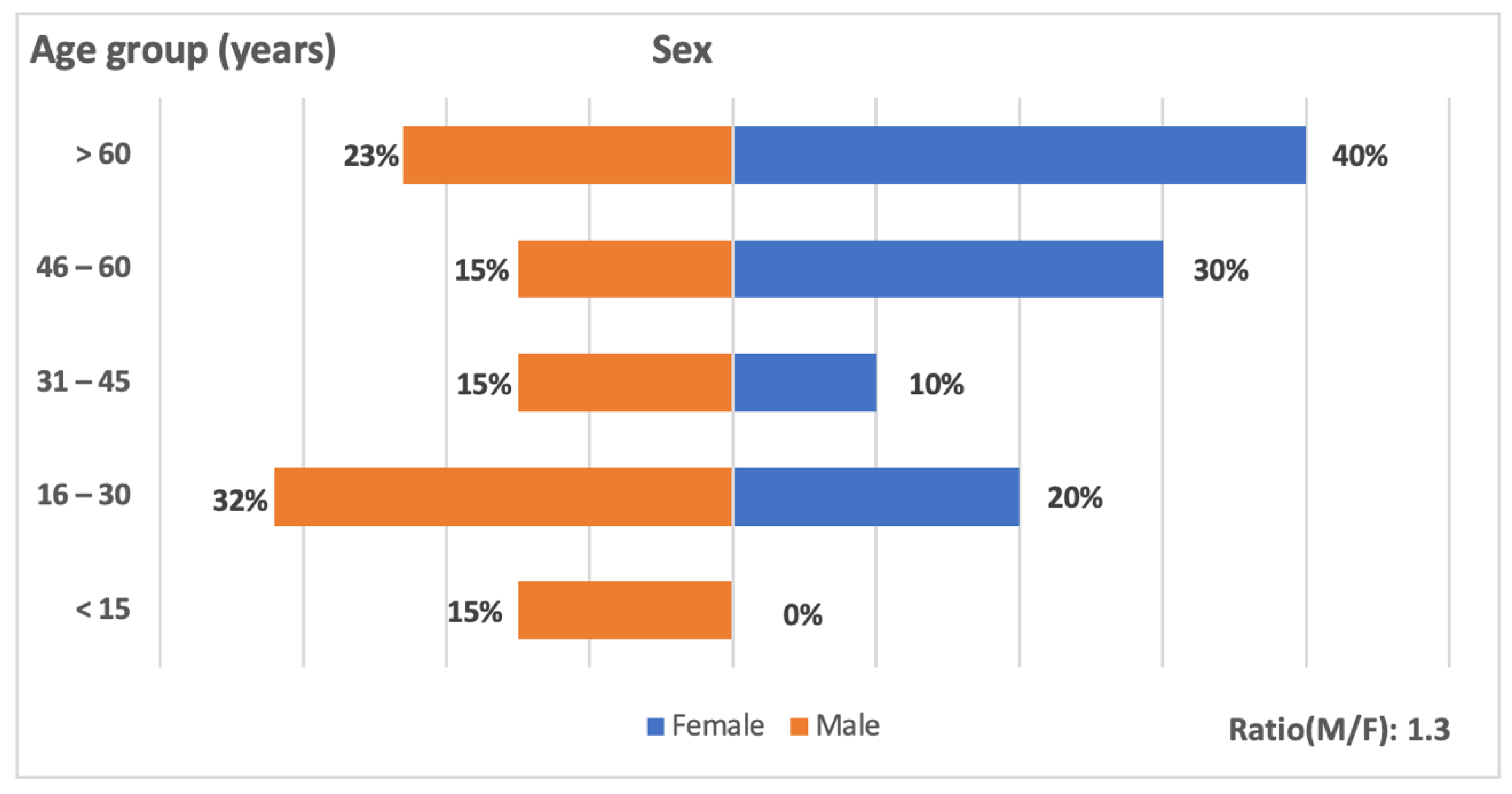
3.1.2. Distribution of WNV Positive Cases by Age and Sex
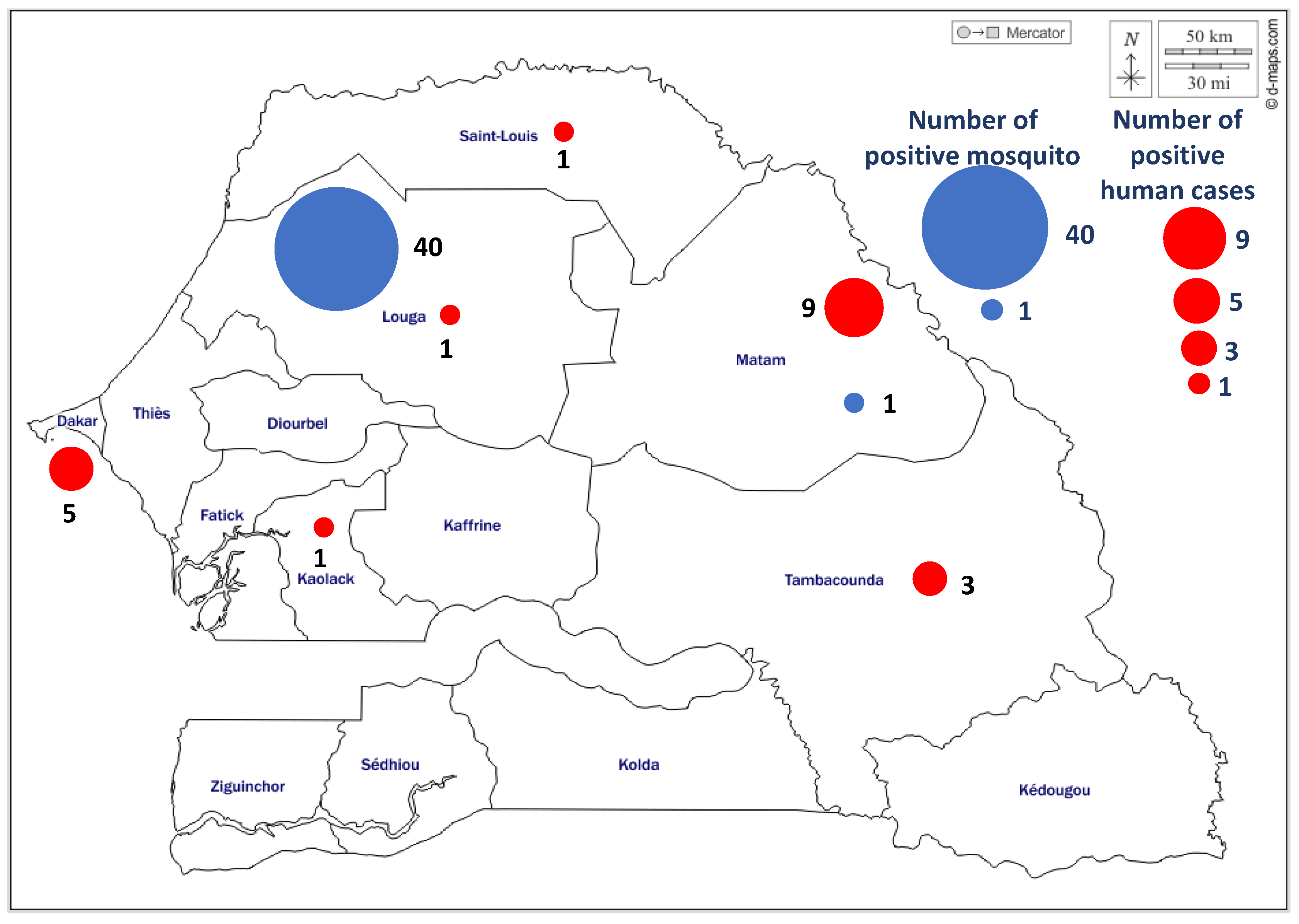
3.1.3. Spatio-Temporal Distibution of WNV Positive Cases
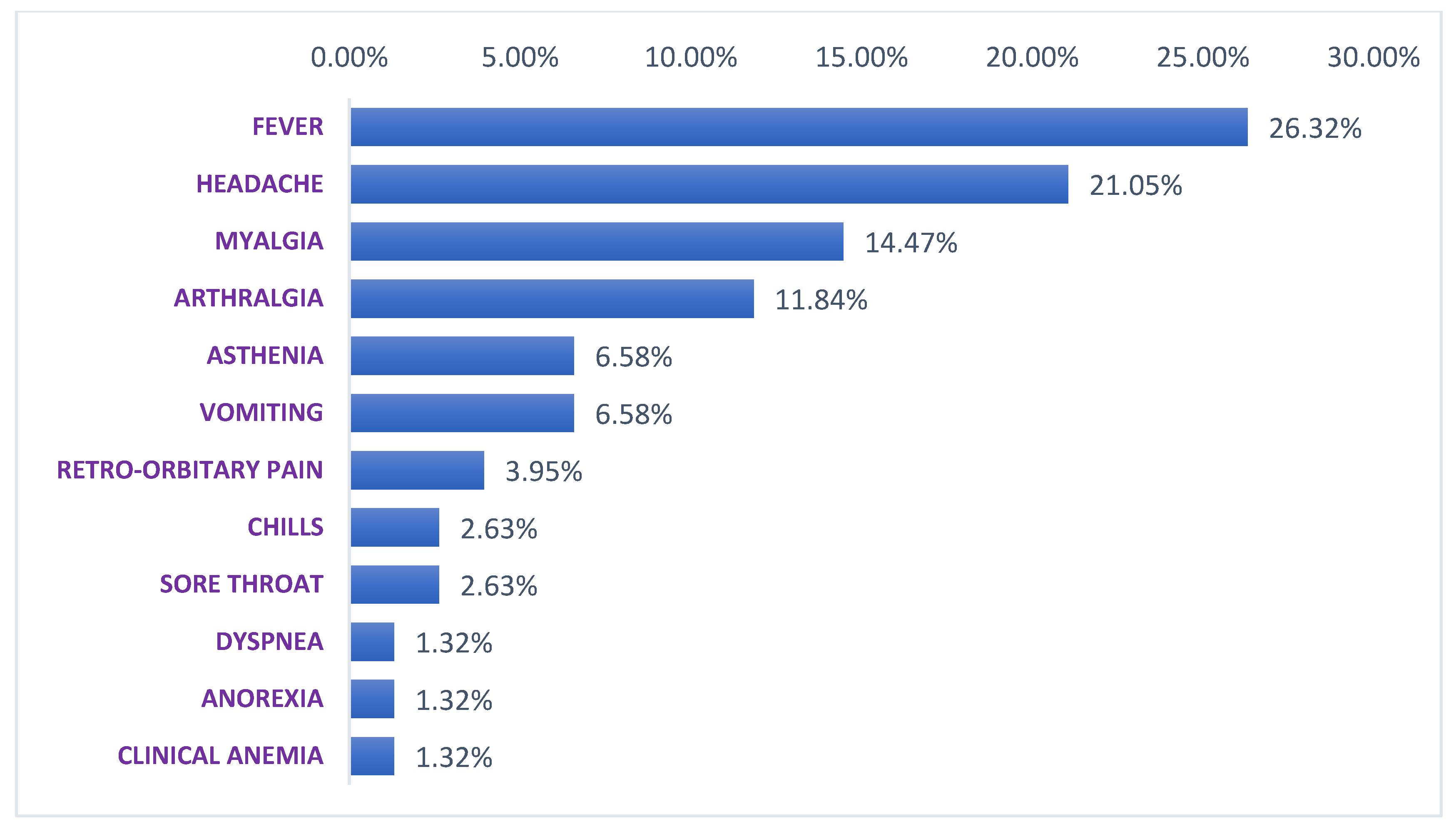
3.1.4. Symptoms Associated to WNV Disease
3.2. Mosquito Samples
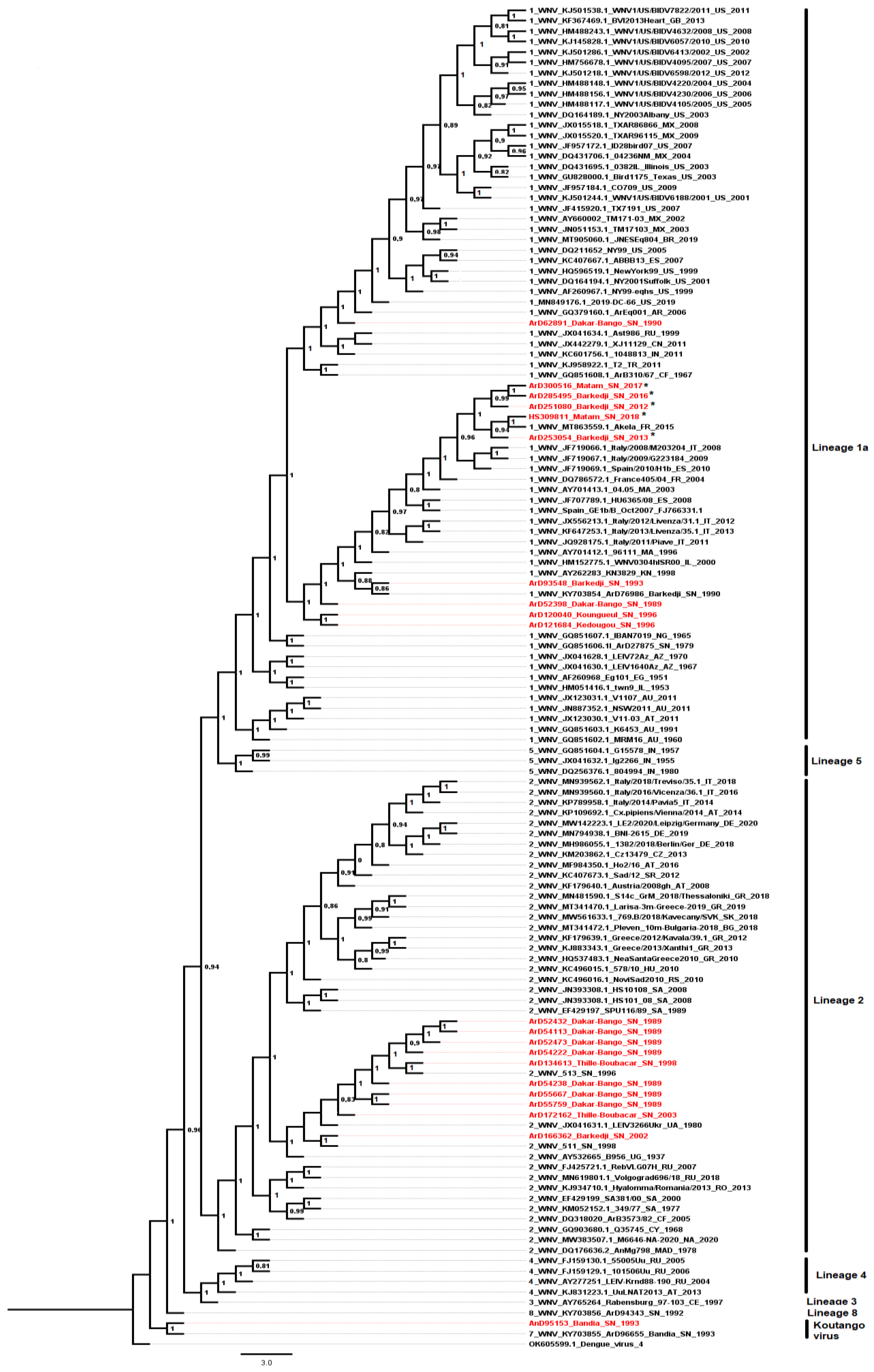
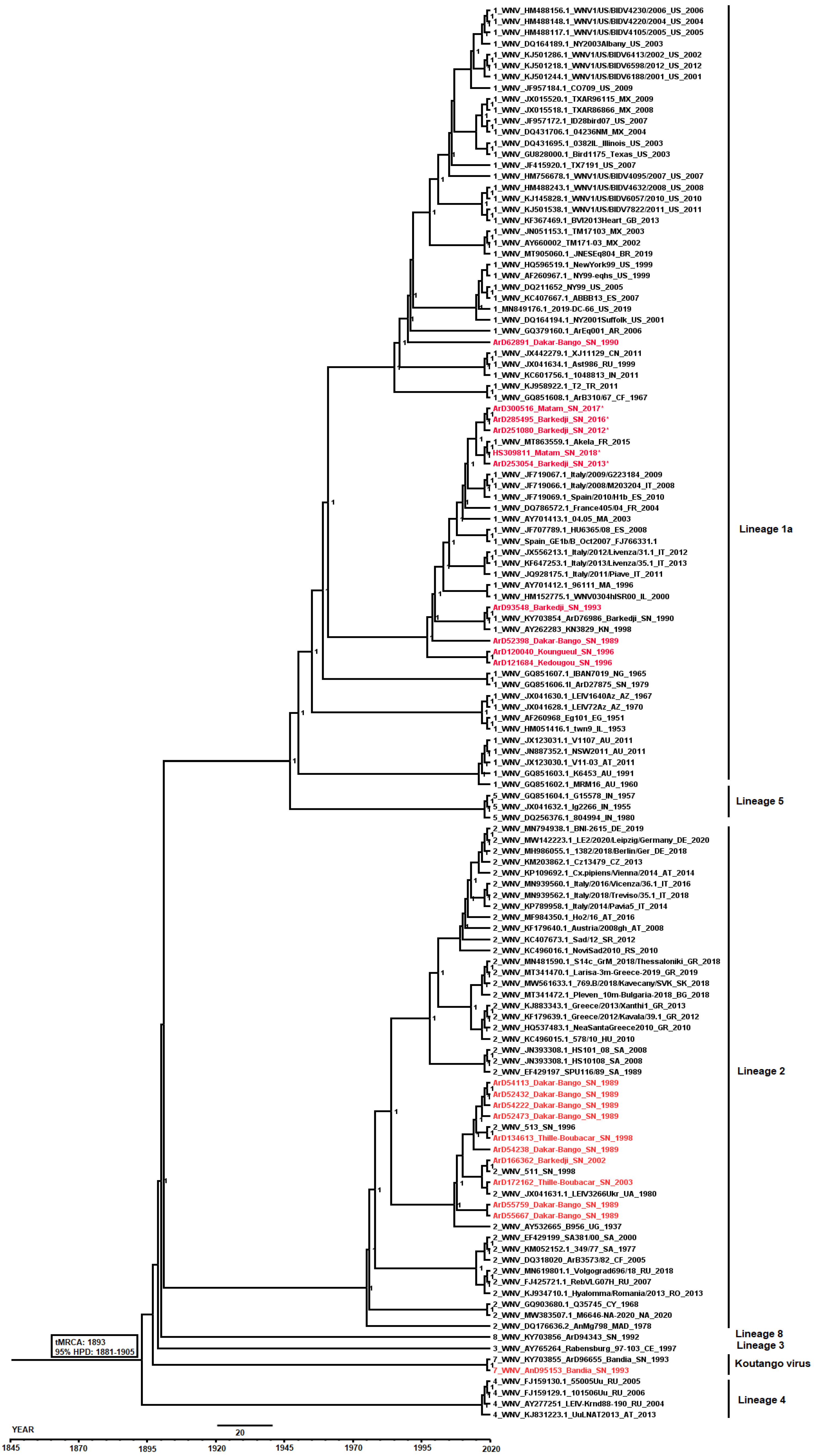
3.3. Phylogenetic Analyses of WNV Strains Isolated in Senegal
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kramer, L.D.; Styer, L.M.; Ebel, G.D. A global perspective on the epidemiology of West Nile virus. Annu. Rev. Entomol. 2008, 53, 61–81. [Google Scholar] [CrossRef] [Green Version]
- Smithburn, K.C.; Hughs, T.P.; Burke, A.W.; Paul, J.H. A neurotropic virus isolated from the blood of a native of Uganda. Am. J. Trop. Med. Hyg. 1940, 20, 471–492. [Google Scholar] [CrossRef]
- Fall, G.; Diallo, M.; Loucoubar, C.; Faye, O.; Sall, A.A. Vector competence of Culex neavei and Culex quinquefasciatus (Diptera: Culicidae) from Senegal for lineages 1, 2, Koutango and a putative new lineage of West Nile virus. Am. J. Trop. Med. Hyg. 2014, 90, 747–754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Available online: https://talk.ictvonline.org/ictv-reports/ictv_online_report/positive-sense-rna-viruses/w/flaviviridae/360/genus-flavivirus (accessed on 23 April 2022).
- Lanciotti, R.S.; Roehrig, J.T.; Deubel, V.; Smith, J.; Parker, M.; Steele, K.; Crise, B.; Volpe, K.E.; Crabtree, M.B.; Scherret, J.H.; et al. Origin of the West Nile virus responsible for an outbreak of encephalitis in the northeastern United States. Science 1999, 286, 2333–2337. [Google Scholar] [CrossRef] [Green Version]
- Hall, R.A.; Scherret, J.H.; Mackenzie, J.S. Kunjin virus: An Australian variant of West Nile? Ann. NY Acad. Sci. 2001, 951, 153–160. [Google Scholar] [CrossRef]
- Murray, K.O.; Mertens, E.; Despres, P. West Nile virus and its emergence in the United States of America. Vet. Res. 2010, 41, 67. [Google Scholar] [CrossRef] [Green Version]
- Bakonyi, T.; Ivanics, E.; Erdelyi, K.; Ursu, K.; Ferenczi, E.; Weissenbock, H.; Nowotny, N. Lineage 1 and 2 strains of encephalitic West Nile virus, central Europe. Emerg. Infect. Dis. 2006, 12, 618–623. [Google Scholar] [CrossRef] [PubMed]
- Bagnarelli, P.; Marinelli, K.; Trotta, D.; Monachetti, A.; Tavio, M.; Del Gobbo, R.; Capobianchi, M.R.; Menzo, S.; Nicoletti, L.; Magurano, F.; et al. Human case of autochthonous West Nile virus lineage 2 infection in Italy, September 2011. Eurosurveillance 2011, 16, 20002. [Google Scholar] [CrossRef] [PubMed]
- Mencattelli, G.; Ndione, M.H.D.; Rosà, R.; Marini, G.; Diagne, C.T.; Diagne, M.M.; Fall, G.; Faye, O.; Diallo, M.; Faye, O.; et al. Epidemiology of West Nile virus in Africa: An underestimated threat. PLoS Negl. Trop. Dis. 2022, 16, e0010075. [Google Scholar] [CrossRef]
- Van der Meulen, K.M.; Pensaert, M.B.; Nauwynck, H.J. West Nile virus in the vertebrate world. Arch. Virol. 2005, 150, 637–657. [Google Scholar] [CrossRef]
- Hayes, E.B.; Sejvar, J.J.; Zaki, S.R.; Lanciotti, R.S.; Bode, A.V.; Campbell, G.L. Virology, pathology, and clinical manifestations of West Nile virus disease. Emerg. Infect. Dis. 2005, 11, 1174. [Google Scholar] [CrossRef]
- Bergsman, L.D.; Hyman, J.M.; Manore, C.A. A mathematical model for the spread of West Nile virus in migratory and resident birds. Math. Biosci. Eng. 2016, 13, 401–424. [Google Scholar] [CrossRef] [PubMed]
- Martin, M.F.; Nisole, S. West Nile Virus Restriction in Mosquito and Human Cells: A Virus under Confinement. Vaccines 2020, 8, 256. [Google Scholar] [CrossRef] [PubMed]
- Monastiri, A.; Mechri, B.; Vázquez-González, A.; Ar Gouilh, M.; Chakroun, M.; Loussaief, C.; Mastouri, M.; Dimassi, N.; Boughzala, L.; Aouni, M.; et al. A four-year survey (2011–2014) of West Nile virus infection in humans, mosquitoes and birds, including the 2012 meningoencephalitis outbreak in Tunisia. Emerg. Microbes Infect. 2018, 7, 28. [Google Scholar] [CrossRef] [Green Version]
- Kolawole, O.M.; Seriki, A.A.; Irekeola, A.A.; Ogah, J.I. The Neglect and Fast Spread of Some Arboviruses: A Note for Healthcare Providers in Nigeria. Diseases 2018, 6, 99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parkash, V.; Woods, K.; Kafetzopoulou, L.; Osborne, J.; Aarons, E.; Cartwright, K. West Nile Virus Infection in Travelers Returning to United Kingdom from South Africa. Emerg. Infect. Dis. 2019, 25, 367–369. [Google Scholar] [CrossRef] [PubMed]
- Bai, F.; Thompson, E.A.; Vig, P.J.S.; Leis, A.A. Current Understanding of West Nile Virus Clinical Manifestations, Immune Responses, Neuroinvasion, and Immunotherapeutic Implications. Pathogens 2019, 8, 193. [Google Scholar] [CrossRef] [Green Version]
- Fall, G.; Di Paola, N.; Faye, M.; Dia, M.; Freire, C.C.d.M.; Loucoubar, C.; Zanotto, P.M.D.A.; Faye, O.; Sall, A.A. Biological and phylogenetic characteristics of West African lineages of West Nile virus. PLoS Negl. Trop. Dis. 2017, 11, e0006078. [Google Scholar] [CrossRef] [Green Version]
- Tsai, T.F.; Popovici, F.; Cernescu, C.; Campbell, G.L.; Nedelcu, N.I. West Nile encephalitis epidemic in southeastern Romania. Lancet 1998, 352, 767–771. [Google Scholar] [CrossRef]
- Kilpatrick, A.M.; Randolph, S.E. Drivers, dynamics, and control of emerging vector-borne zoonotic diseases. Lancet 2012, 380, 1946–1955. [Google Scholar] [CrossRef]
- Weinberger, M.; Pitlik, S.D.; Gandacu, D.; Lang, R.; Nassar, F.; Ben David, D.; Rubinstein, E.; Izthaki, A.; Mishal, J.; Kitzes, R.; et al. West Nile fever outbreak, Israel, 2000: Epidemiologic aspects. Emerg. Infect. Dis. 2001, 7, 686–691. [Google Scholar] [CrossRef] [PubMed]
- Zeller, H.G.; Schuffenecker, I. West Nile virus: An overview of its spread in Europe and the Mediterranean basin in contrast to its spread in the Americas. Eur. J. Clin. Microbiol. Infect. Dis. 2004, 23, 147–156. [Google Scholar] [CrossRef] [PubMed]
- Rossini, G.; Carletti, F.; Bordi, L.; Cavrini, F.; Gaibani, P.; Landini, M.P.; Pierro, A.; Capobianchi, M.R.; Di Caro, A.; Sambri, V. Phylogenetic analysis of west nile virus isolates, Italy, 2008–2009. Emerg. Infect. Dis. 2011, 17, 903–906. [Google Scholar] [CrossRef] [PubMed]
- Barzon, L.; Papa, A.; Lavezzo, E.; Franchin, E.; Pacenti, M.; Sinigaglia, A.; Masi, G.; Trevisan, M.; Squarzon, L.; Toppo, S. Phylogenetic characterization of Central/Southern European lineage 2 West Nile virus: Analysis of human outbreaks in Italy and Greece, 2013–2014. Clin. Microbiol. Infect. 2015, 21, 1122.e1–1122.e10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bakonyi, T.; Ferenczi, E.; Erdélyi, K.; Kutasi, O.; Csörgő, T.; Seidel, B.; Weissenböck, H.; Brugger, K.; Bán, E.; Nowotny, N. Explosive spread of a neuroinvasive lineage 2 West Nile virus in Central Europe, 2008/2009. Vet. Microbiol. 2013, 165, 61–70. [Google Scholar] [CrossRef]
- Popović, N.; Milošević, B.; Urošević, A.; Poluga, J.; Lavadinović, L.; Nedelijković, J.; Jevtović, D.; Dulović, O. Outbreak of West Nile virus infection among humans in Serbia, August to October 2012. Eurosurveillance 2013, 18, 20613. [Google Scholar] [CrossRef] [Green Version]
- CDC. Available online: https://www.cdc.gov/westnile/statsmaps/cumMapsData.html (accessed on 23 April 2022).
- Depoortere, E.; Kavle, J.; Keus, K.; Zeller, H.; Murri, S.; Legros, D. Outbreak of West Nile virus causing severe neurological involvement in children, Nuba Mountains, Sudan, 2002. Trop. Med. Int. Health 2004, 9, 730–736. [Google Scholar] [CrossRef] [Green Version]
- Abroug, F.; Ouanes-Besbes, L.; Letaief, M.; Ben, R.F.; Khairallah, M.; Triki, H.; Bouzouiaia, N. A cluster study of predictors of severe West Nile virus infection. Mayo Clin. Proc. 2006, 81, 12–16. [Google Scholar] [CrossRef]
- Hammami, S.; Hassine, T.B.; Conte, A.; Amdouni, J.; De Massis, F.; Sghaier, S.; Hassen, S.B. West Nile disease in Tunisia: An overview of 60 years. Vet. Ital. 2017, 53, 225–234. [Google Scholar]
- Zaayman, D.; Venter, M. West Nile virus neurologic disease in humans, South Africa, September 2008–May 2009. Emerg. Infect. Dis. 2012, 18, 2051–2054. [Google Scholar] [CrossRef] [PubMed]
- Chevalier, V.; Lancelot, R.; Diaité, A.; Mondet, B.; Sall, B.; De Lamballerie, X. Serological Assessment of West Nile Fever Virus Activity in the Pastoral System of Ferlo, Senegal. Ann. N. Y. Acad. Sci. 2006, 1081, 216–225. [Google Scholar] [CrossRef] [PubMed]
- Chevalier, V.; Reynaud, P.; Lefrancois, T.; Durand, B.; Baillon, F.; Balanca, G.; Gaidet, N.; Mondet, B.; Lancelot, R. Predicting West Nile virus seroprevalence in wild birds in Senegal. Vector Borne Zoonotic Dis. 2009, 10, 589–596. [Google Scholar] [CrossRef]
- Cabre, O.; Durand, J.P.; Prange, A.; Gomez, J.; Maurizi, L.; Tolou, H.; Davoust, B. West Nile virus infection: Serological investigation among horses in France and in Africa. Med. Trop. 2005, 65, 439–443. [Google Scholar]
- Chevalier, V.; Dupressoir, A.; Tran, A.; Diop, O.M.; Gottland, C.; Iallo, M.D.; Etter, E.; Ndiaye, M.; Grosbois, V.; Dia, M.; et al. Environmental risk factors of West Nile virus infection of horses in the Senegal River basin. Epidemiol. Infect. 2010, 138, 1601–1609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gordon, S.W.; Tammariello, R.F.; Linthicum, K.J.; Dohm, O.J.; Digoutte, J.P.; Calvo-Wilson, M.A. Arbovirus isolations from mosquitoes collected during 1988 in the Senegal River basin. Am. J. Trop. Med. Hyg. 1992, 47, 742–748. [Google Scholar] [CrossRef] [Green Version]
- Traore-Lamizana, M.; Zeller, H.G.; Mondo, M.; Hervy, J.P.; Adam, F.; And Digoutte, J.P. Isolations of West Nile and Bagaza Viruses from Mosquitoes (Diptera: Culicidae) in Central Senegal (Ferlo). J. Med. Entomol. 1994, 31, 934–938. [Google Scholar] [CrossRef]
- Renaudet, J.; Jan, C.; Ridet, J.; Adam, C.; Robin, Y. A serological survey of arboviruses in the human population of Senegal. Bull. Soc. Pathol. Exot. Fil. 1977, 71, 131–140. [Google Scholar]
- Monlun, E.; Zeller, H.; Le Guenno, B.; Traoré-Lamizana, M.; Hervy, J.P.; Adam, F.; Ferrara, L.; Fontenille, D.; Sylla, R.; Mondo, M. Surveillance of the circulation of arbovirus of medical interest in the region of eastern Senegal. Bull. Soc. Pathol. Exot. 1990 1993, 86, 21–28. [Google Scholar]
- Murgue, B.; Zeller, H.; Deubel, V. The Ecology and Epidemiology of West Nile Virus in Africa, Europe and Asia. In Japanese Encephalitis and West Nile Viruses; Springer: Berlin/Heidelberg, Germany, 2002; pp. 195–221. [Google Scholar] [CrossRef]
- Barry, M.A.; Arinal, F.; Talla, C.; Hedible, B.G.; Sarr, F.D.; Ba, I.O.; Diop, B.; Dia, N.; Vray, M. Performance of case definitions and clinical predictors for influenza surveillance among patients followed in a rural cohort in Senegal. BMC Infect. Dis. 2021, 21, 31. [Google Scholar] [CrossRef]
- Bob, N.S.; Barry, M.A.; Diagne, M.M.; Faye, M.; Ndione, M.H.D.; Diallo, A.; Diop, M.; Diop, B.; Faye, O.; Loucoubar, C.; et al. Detection of Rift Valley Fever Virus Lineage H From South Africa Through the Syndromic Sentinel Surveillance Network in Senegal. Open Forum Infect. Dis. 2021, 9, ofab655. [Google Scholar] [CrossRef]
- Ndiaye, E.H.; Diallo, D.; Fall, G.; Ba, Y.; Faye, O.; Dia, I.; Diallo, M. Arboviruses isolated from the Barkedji mosquito based surveillance system, 2012–2013. BMC Infect. Dis. 2018, 18, 642. [Google Scholar] [CrossRef]
- Traore-Lamizana, M.; Fontenille, D.; Diallo, M.; Ba, Y.; Zeller, H.G.; Mondo, M.; Adam, F.; Thonnon, J.; Maiga, A. Arbovirus surveillance from 1990 to 1995 in the Barkedji area (Ferlo) of Senegal, a possible natural focus of Rift Valley fever virus. J. Med. Entomol. 2001, 38, 480–492. [Google Scholar] [CrossRef]
- Clark, G.G.; Seda, H.; Gubler, D.J. Use of the “CDC backpack aspirator” for surveillance of aedes aegyptiin San Juan, Puerto rico. J. Am. Mosq. Control. Assoc. 1994, 10, 119–124. [Google Scholar] [PubMed]
- Sudia, W.D.; Chamberlain, R.W. Battery-operated light trap, an improved model. By W. D. Sudia and R. W. Chamberlain, 1962. J. Am. Mosq. Control Assoc. 1988, 4, 536–538. [Google Scholar] [PubMed]
- Service, M.W. Mosquito Ecology Field Sampling Methods; Springer: Heidelberg, The Netherlands, 1993. [Google Scholar]
- Edwards, F. Mosquitoes of the Ethiopian Region: III Culicine Adults and Pupae; British Museum (Natural History): London, UK, 1941. [Google Scholar]
- Diagne, N.; Fontenille, D.; Konate, L.; Faye, O.; Lamizana, M.T.; Legros, F.; Molez, J.F.; Trape, J.F. Anopheles of Senegal. An annotated and illustrated list. Bull. Soc. Pathol. Exot. 1994, 87, 267–277. [Google Scholar] [PubMed]
- Faye, O.; de Lourdes Monteiro, M.; Vrancken, B.; Prot, M.; Lequime, S.; Diarra, M.; Ndiaye, O.; Valdez, T.; Tavarez, S.; Ramos, J.; et al. Genomic Epidemiology of 2015–2016 Zika Virus Outbreak in Cape Verde. Emerg. Infect. Dis. 2020, 26, 1084–1090. [Google Scholar] [CrossRef]
- Fall, G.; Faye, M.; Weidmann, M.; Kaiser, M.; Dupressoir, A.; Ndiaye, E.H.; Ba, Y.; Diallo, M.; Faye, O.; Sall, A.A. Real-Time RT-PCR Assays for Detection and Genotyping of West Nile Virus Lineages Circulating in Africa. Vector Borne Zoonotic Dis. 2016, 16, 781–789. [Google Scholar] [CrossRef]
- Digoutte, J.P.; Calvo-Wilson, M.A.; Mondo, M.; Traore-Lamizana, M.; Adam, F. Continuous cell lines and immune ascitic fluid pools in arbovirus detection. Res. Virol. 1992, 143, 417–422. [Google Scholar] [CrossRef]
- Allander, T.; Emerson, S.U.; Engle, R.E.; Purcell, R.H.; Bukh, J. A virus discovery method incorporating DNase treatment and its application to the identification of two bovine parvovirus species. Proc. Natl. Acad. Sci. USA 2001, 98, 11609–11614. [Google Scholar] [CrossRef] [Green Version]
- Li, P.E.; Lo, C.C.; Anderson, J.J.; Davenport, K.W.; Bishop-Lilly, K.A.; Xu, Y.; Ahmed, S.; Feng, S.; Mokashi, V.P.; Chain, P.S. Enabling the democratization of the genomics revolution with a fully integrated web-based bioinformatics platform. Nucleic Acids Res. 2017, 45, 67–80. [Google Scholar] [CrossRef] [Green Version]
- Okonechnikov, K.; Golosova, O.; Fursov, M. The UGENE team. Unipro UGENE: A unified bioinformatics toolkit. Bioinformatics 2012, 28, 1166–1167. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [Green Version]
- Kalyaanamoorthy, S.; Minh, B.; Wong, T.; Von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef]
- Nguyen, L.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A Fast and Effective Stochastic Algorithm for Estimating Maximum-Likelihood Phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Price, M.N.; Paramvir, S.D.; Arkin, A.P. FastTree 2: Approximately Maximum-Likelihood Trees for Large Alignments. PLoS ONE 2010, 5, e9490. [Google Scholar] [CrossRef]
- Figtree. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 5 May 2022).
- Drummond, A.J.; Suchard, M.A.; Xie, D.; Rambaut, A. Bayesian phylogenetics with BEAUti and the BEAST 1.7. Mol. Biol. Evol. 2012, 29, 1969–1973. [Google Scholar] [CrossRef] [Green Version]
- Tracer. Available online: http://beast.bio.ed.ac.uk/Tracer (accessed on 5 May 2022).
- Treeannotator. Available online: http://beast.community/treeannotator (accessed on 5 May 2022).
- Dieng, I.; Ndione, M.H.D.; Fall, C.; Diagne, M.M.; Diop, M.; Gaye, A.; Barry, M.A.; Diop, B.; Ndiaye, M.; Bousso, A.; et al. Multifoci and multiserotypes circulation of dengue virus in Senegal between 2017 and 2018. BMC Infect. Dis. 2021, 21, 867. [Google Scholar] [CrossRef]
- Dieng, I.; Barry, M.A.; Diagne, M.M.; Diop, B.; Ndiaye, M.; Faye, M.; Ndione, M.H.D.; Dieng, M.M.; Bousso, A.; Fall, G.; et al. Detection of Crimean Congo haemorrhagic fever virus in North-eastern Senegal, Bokidiawé 2019. Emerg. Microbes Infect. 2020, 9, 2485–2487. [Google Scholar] [CrossRef]
- Petersen, L.R.; Brault, A.C.; Nasci, R.S. West Nile virus: Review of the literature. JAMA 2013, 310, 308–315. [Google Scholar] [CrossRef]
- Popescu, C.P.; Florescu, S.A.; Cotar, A.I.; Badescu, D.; Ceianu, C.S.; Zaharia, M.; Tardei, G.; Codreanu, D.; Ceausu, E.; Ruta, S.M. Re-emergence of severe West Nile virus neuroinvasive disease in humans in Romania, 2012 to 2017-implications for travel medicine. Travel Med. Infect. Dis. 2018, 22, 30–35. [Google Scholar] [CrossRef]
- Habarugira, G.; Suen, W.W.; Hobson-Peters, J.; Hall, R.A.; Bielefeldt-Ohmann, H. West Nile Virus: An Update on Pathobiology, Epidemiology, Diagnostics, Control and “One Health” Implications. Pathogens 2020, 9, 589. [Google Scholar] [CrossRef] [PubMed]
- O’Leary, D.R.; Marfin, A.A.; Montgomery, S.P.; Kipp, A.M.; Lehman, J.A.; Biggerstaff, B.J.; Elko, V.L.; Collins, P.D.; Jones, J.E.; Campbell, G.L. The epidemic of West Nile virus in the United States, 2002. Vector Borne Zoonotic Dis. 2004, 4, 61–70. [Google Scholar] [CrossRef] [PubMed]
- Barzon, L.; Pacenti, M.; Franchin, E.; Pagni, S.; Martello, T.; Cattai, M.; Cusinato, R.; Palù, G. Excretion of West Nile virus in urine during acute infection. J. Infect. Dis. 2013, 208, 1086–1092. [Google Scholar] [CrossRef] [PubMed]
- Nagy, A.; Bán, E.; Nagy, O.; Ferenczi, E.; Farkas, Á.; Bányai, K.; Farkas, S.; Takács, M. Detection and sequencing of West Nile virus RNA from human urine and serum samples during the 2014 seasonal period. Arch. Virol. 2016, 161, 1797–1806. [Google Scholar] [CrossRef]
- Chevalier, V.; Lancelot, R.; Thiongane, Y.; Sall, B.; Diaité, A.; Mondet, B. Rift Valley fever in small ruminants, Senegal, 2003. Emerg. Infect. Dis. 2005, 11, 1693–1700. [Google Scholar] [CrossRef]
- Biteye, B.; Fall, A.G.; Ciss, M.; Seck, M.T.; Apolloni, A.; Fall, M.; Tran, A.; Gimonneau, G. Ecological distribution and population dynamics of Rift Valley fever virus mosquito vectors (Diptera, Culicidae) in Senegal. Parasit. Vectors 2018, 11, 27. [Google Scholar] [CrossRef] [PubMed]
- Bakonyi, T.; Haussig, J.M. West Nile virus keeps on moving up in Europe. Eurosurveillance 2020, 25, 2001938. [Google Scholar] [CrossRef]
- Gu, W.; Unnasch, T.R.; Katholi, C.R.; Lampman, R.; Novak, R.J. Fundamental issues in mosquito surveillance for arboviral transmission. Trans. R Soc. Trop. Med. Hyg. 2008, 102, 817–822. [Google Scholar] [CrossRef] [Green Version]
- Gowda, N.N.; Vijayan, V.A. Indoor resting density, survival rate and host preference of Culex quinquefasciatus say (Diptera: Culicidae) in Mysore City. J. Commun. Dis. 1992, 24, 20–28. [Google Scholar]
- Rappole, J.H.; Derrickson, S.R.; HubaÂlek, Z.; HubaÂlek, Z. Migratory birds and spread of West Nile virus in the Western Hemisphere. Emerg. Infect. Dis. 2000, 6, 319. [Google Scholar] [CrossRef]
- Camp, J.V.; Nowotny, N. The knowns and unknowns of West Nile virus in Europe: What did we learn from the 2018 outbreak? Expert Rev. Anti-Infect. Ther. 2020, 18, 145–154. [Google Scholar] [CrossRef] [PubMed]
- Pietsch, C.; Michalski, D.; Münch, J.; Petros, S.; Bergs, S.; Trawinski, H.; Lübbert, C.; Liebert, U.G. Autochthonous West Nile virus infection outbreak in humans, Leipzig, Germany, August to September 2020. Eurosurveillance 2020, 25, 2001786. [Google Scholar] [CrossRef] [PubMed]
- Young, J.J.; Haussig, J.M.; Aberle, S.W.; Pervanidou, D.; Riccardo, F.; Sekulić, N.; Bakonyi, T.; Gossner, C.M. Epidemiology of human West Nile virus infections in the European Union and European Union enlargement countries, 2010 to 2018. Eurosurveillance 2021, 26, 2001095. [Google Scholar] [CrossRef] [PubMed]
- Platonov, A.E.; Karan, L.S.; Shopenskaia, T.A.; Fedorova, M.V.; Koliasnikova, N.M.; Rusakova, N.M.; Shishkina, L.V.; Arshba, T.E.; Zhuravlev, V.I.; Govorukhina, M.V.; et al. Genotyping of West Nile fever virus strains circulating in southern Russia as an epidemiological investigation method: Principles and results. Zh. Mikrobiol. Epidemiol. Immunobiol. 2011, 2, 29–37. [Google Scholar]
- Bakonyi, T.; Hubálek, Z.; Rudolf, I.; Nowotny, N. Novel flavivirus or new lineage of West Nile virus, central Europe. Emerg. Infect. Dis. 2005, 11, 225–231. [Google Scholar] [CrossRef] [PubMed]
- Zehender, G.; Veo, C.; Ebranati, E.; Carta, V.; Rovida, F.; Percivalle, E.; Moreno, A.; Lelli, D.; Calzolari, M.; Lavazza, A.; et al. Reconstructing the recent West Nile virus lineage 2 epidemic in Europe and Italy using discrete and continuous phylogeography. PLoS ONE 2017, 12, e0179679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chancey, C.; Grinev, A.; Volkova, E.; Rios, M. The global ecology and epidemiology of West Nile virus. Biomed. Res. Int. 2015, 2015, 376230. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Years | Number of Tested Samples |
|---|---|
| 2015 | 123 |
| 2016 | 515 |
| 2017 | 980 |
| 2018 | 1579 |
| 2019 | 1293 |
| 2020 | 1149 |
| 2021 | 2273 |
| IPD Number | Center | Reception Date | IgM | Titer /PRNT | Signs | Age | Sex | T° |
|---|---|---|---|---|---|---|---|---|
| 283397 | PS PONT | 18/10/2016 | POS | 10 | FEVER, CEPHALEAS, MYALGIAS, VOMITING | 37 | M | 38.9 |
| 309809 | PS BOKIDIAWE | 18/10/2018 | POS | 40 | FEVER, CEPHALEAS, ARTHRALGIAS, | 48 | M | 39 |
| 310012 | CS COKI | 23/10/2018 | POS | 80 | FEVER, CEPHALEAS, MYALGIAS, ARTHRALGIAS, ASTHENIA, DYSPNEA, CHILLS | 30 | M | 39.1 |
| 310192 | PS BOKIDIAWE | 26/10/2018 | POS | >320 | FEVER, CEPHALEAS, MYALGIAS | 21 | F | 38 |
| 310255 | PS SAINT LOUIS | 30/10/2018 | POS | 20 | FEVER | 3 MONTHS | M | 38.8 |
| 317510 | CS HANN MARISTES | 13/11/2018 | POS | 160 | FEVER, CEPHALEAS, MYALGIA, ASTHENIA, CHILLS, VOMITING | 28 | M | 39 |
| 317575 | DAKAR | 15/11/2018 | POS | 80 | FEVER | 33 | M | |
| 323078 | PS BOKIDIAWE | 25/11/2019 | POS | 10 | FEVER, CEPHALEAS, ARTHRALGIAS, SORE THROAT | 21 | F | 38.9 |
| 326040 | PS PONT | 24/09/2020 | POS | 80 | FEVER, CEPHALEAS, MYALGIAS, ARTHRALGIAS, ANOREXIA | 58 | M | 39.9 |
| 327049 | PS BOKIDIAWE | 15/10/2020 | POS | 80 | FEVER, CEPHALEAS, ARTHRALGIAS, SORE THROAT | NA | F | 38.2 |
| 328043 | PS BOKIDIAWE | 22/10/2020 | POS | >320 | FEVER, CEPHALEAS, ARTHRALGIAS, CLINICAL ANEMIA, VOMITING | 8 | M | 38 |
| 329097 | DAKAR | 02/11/2020 | POS | 80 | FEVER, CEPHALEAS, MYALGIAS | 77 | F | 38 |
| 330006 | DISTRICT OUEST | 03/11/2020 | POS | 40 | FEVER, CEPHALEAS, MYALGIAS, ARTHRALGIAS, ASTHENIA, VOMITING, RETRO-ORBITARY PAIN | 58 | F | >38 |
| 350013 | DISTRICT OUEST | 06/11/2020 | POS | >320 | FEVER, CEPHALEAS, MYALGIAS, ARTHRALGIAS, ASTHENIA, RETRO-ORBITARY PAIN | 21 | M | 39.5 |
| 363215 | PS BOKIDIAWE | 10/12/2020 | POS | 80 | FEVER, CEPHALEAS, MYALGIAS | 60 | F | 39.4 |
| 363216 | PS BOKIDIAWE | 10/12/2020 | POS | >320 | FEVER, CEPHALEAS, MYALGIAS, ASTHENIAS | 65 | M | 38.7 |
| 363264 | PS OREFONDE | 21/12/2020 | POS | 20 | FEVER | 19 | M | NA |
| 363431 | PS PONT | 14/01/2021 | POS | 10 | FEVER, CEPHALEAS, MYALGIAS, ARTHRALGIAS, VOMITING | 56 | F | 39.4 |
| 377948 | GOSSAS | 24/11/2021 | POS | 20 | FEVER | 68 | M | 36 |
| Year | Number of Mosquito Pools Collected | Number of WNV Lineage 1 Positive Pools | Area of Virus Isolation | Mosquito Species |
|---|---|---|---|---|
| 2012 | 2185 | 18 | Barkédji | Cx. quinquefasciatus (1), Cx. perfuscus (1), Cx. neavei (9), Cx. poicilipes (1), Cx. tritaeniorhynchus (1), Cx. antennatus (3), An. rufipes (1) and Ae. dalzieli (1) |
| 2013 | 2069 | 14 | Barkédji | Cx. neavei (13), Cx. tritaeniorhynchus (1) |
| 2014 | 517 | 0 | NA | |
| 2015 | 722 | 0 | NA | |
| 2016 | 1214 | 8 | Barkédji | Cx. neavei (3), Cx. poicilipes (4) and Cx. ethiopicus (1) |
| 2017 | 2761 | 1 | Matam | Cx. neavei (1) |
| 2018 | 727 | 0 | NA | |
| 2019 | 859 | 0 | NA | |
| 2020 | 1955 | 0 | NA |
| Strain | Origin | Year of Isolation | Locality | Genbank Accession Number |
|---|---|---|---|---|
| ArD52398 | Mosquito | 1989 | Dakar Bango | OP846971 |
| ArD52432 | Mosquito | 1989 | Dakar Bango | OP870453 |
| ArD54113 | Mosquito | 1989 | Dakar Bango | OP870454 |
| ArD52473 | Mosquito | 1989 | Dakar Bango | OP870455 |
| ArD54222 | Mosquito | 1989 | Dakar Bango | OP870456 |
| ArD54238 | Mosquito | 1989 | Dakar Bango | OP846976 |
| ArD55667 | Mosquito | 1989 | Dakar Bango | OP870457 |
| ArD55759 | Mosquito | 1989 | Dakar Bango | OP870458 |
| ArD62891 | Mosquito | 1990 | Dakar Bango | OP846977 |
| AnD95153 | Rodent | 1993 | Bandia | OP846972 |
| ArD93548 | Mosquito | 1993 | Barkédji | OP846973 |
| ArD120040 | Mosquito | 1996 | Koungueul | OP846974 |
| ArD121684 | Mosquito | 1996 | Kédougou | OP846975 |
| ArD134613 | Mosquito | 1998 | Thille Boubacar | OP870459 |
| ArD166362 | Mosquito | 2002 | Barkédji | OP870461 |
| ArD172162 | Mosquito | 2003 | Thille Boubacar | OP870460 |
| ArD251080 | Mosquito | 2012 | Barkédji | OP846978 |
| ArD253054 | Mosquito | 2013 | Barkédji | OP846979 |
| ArD285495 | Mosquito | 2016 | Barkédji | OP846982 |
| ArD300516 | Mosquito | 2017 | Matam | OP846980 |
| HS309811 | Human | 2018 | Matam | OP846981 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ndione, M.H.D.; Ndiaye, E.H.; Faye, M.; Diagne, M.M.; Diallo, D.; Diallo, A.; Sall, A.A.; Loucoubar, C.; Faye, O.; Diallo, M.; et al. Re-Introduction of West Nile Virus Lineage 1 in Senegal from Europe and Subsequent Circulation in Human and Mosquito Populations between 2012 and 2021. Viruses 2022, 14, 2720. https://doi.org/10.3390/v14122720
Ndione MHD, Ndiaye EH, Faye M, Diagne MM, Diallo D, Diallo A, Sall AA, Loucoubar C, Faye O, Diallo M, et al. Re-Introduction of West Nile Virus Lineage 1 in Senegal from Europe and Subsequent Circulation in Human and Mosquito Populations between 2012 and 2021. Viruses. 2022; 14(12):2720. https://doi.org/10.3390/v14122720
Chicago/Turabian StyleNdione, Marie Henriette Dior, El Hadji Ndiaye, Martin Faye, Moussa Moïse Diagne, Diawo Diallo, Amadou Diallo, Amadou Alpha Sall, Cheikh Loucoubar, Oumar Faye, Mawlouth Diallo, and et al. 2022. "Re-Introduction of West Nile Virus Lineage 1 in Senegal from Europe and Subsequent Circulation in Human and Mosquito Populations between 2012 and 2021" Viruses 14, no. 12: 2720. https://doi.org/10.3390/v14122720