
Subtherapeutic Doses of Vancomycin Synergize with Bacteriophages for Treatment of Experimental Methicillin-Resistant Staphylococcus aureus Infective Endocarditis


{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains, Bacterial Genomic DNA, and Bacteriophages
2.2. In Vitro Activities of Phages and Vancomycin against Planktonic Cultures and Biofilms
2.3. Murine Model of EE
2.4. Treatment Protocol
2.5. Outcomes
2.6. Statistical Analysis
3. Results
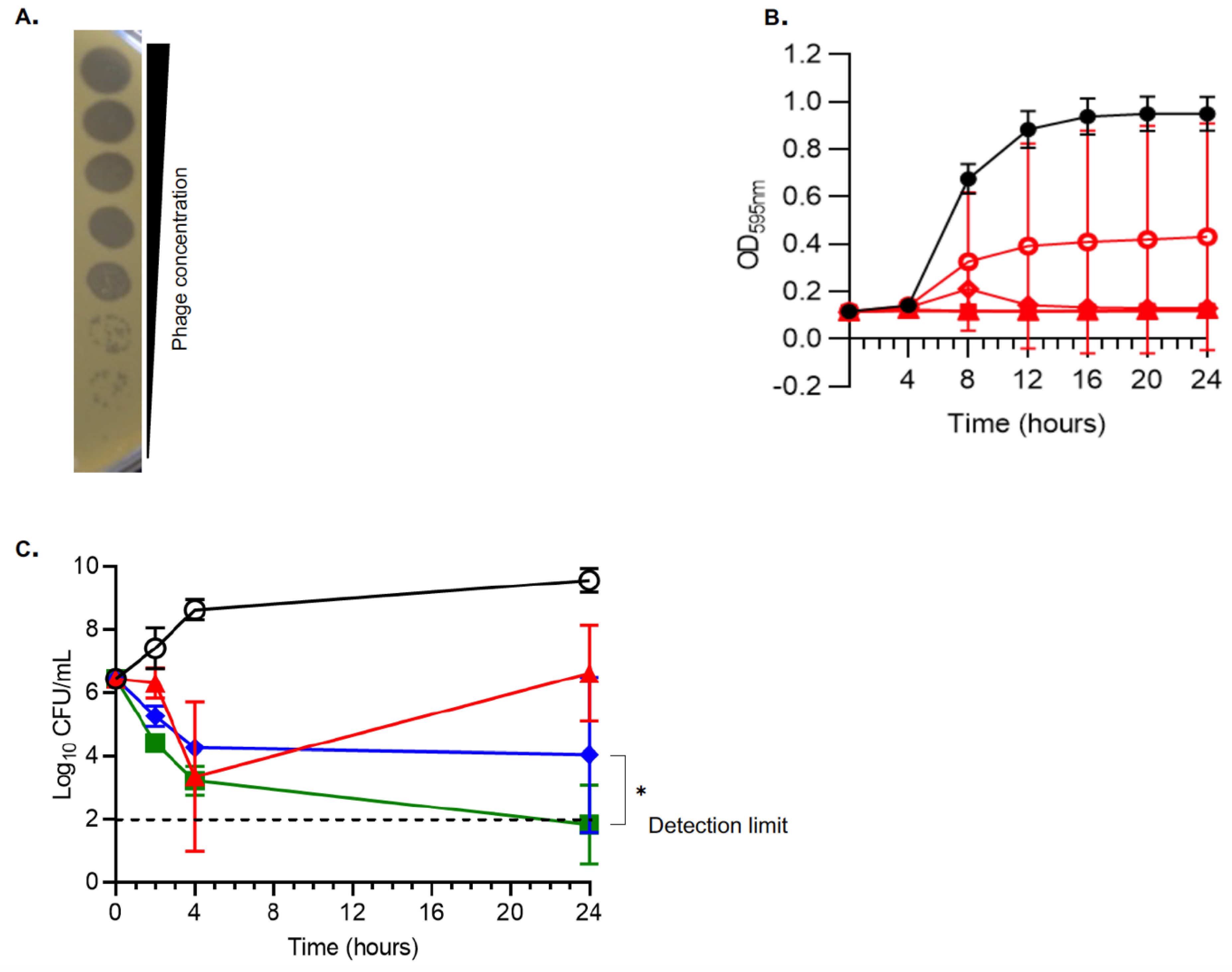
3.1. In Vitro Activity of vB_SauH_2002/Phage 66 Cocktail on MRSA AW7
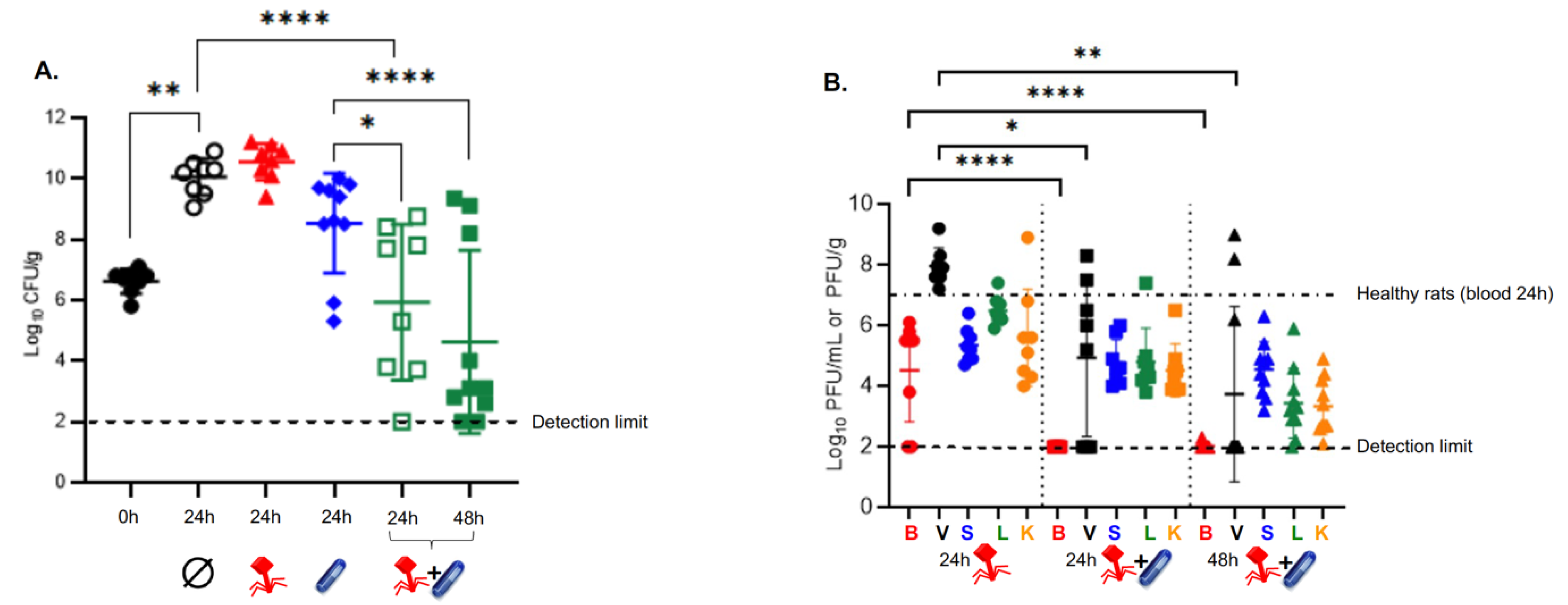
3.2. Subtherapeutic Doses of Vancomycin Synergize with the Phage Cocktail In Vivo
3.3. Subtherapeutic Doses of Vancomycin Impair the PK Profile of Phage In Vivo
3.4. The Phage 66-Resistance Phenotype Observed In Vivo Is Not Linked to Genetic Determinants
3.5. Despite Efficient Infection, Phage 66 Is Not Able to Control the In Vitro Growth of Planktonic MRSA AW7 Cells
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Que, Y.A.; Moreillon, P. Infective endocarditis. Nat. Rev. Cardiol. 2011, 8, 322–336. [Google Scholar] [CrossRef] [PubMed]
- de Kraker, M.E.; Wolkewitz, M.; Davey, P.G.; Grundmann, H. Clinical impact of antimicrobial resistance in European hospitals: Excess mortality and length of hospital stay related to methicillin-resistant Staphylococcus aureus bloodstream infections. Antimicrob. Agents Chemother. 2011, 55, 1598–1605. [Google Scholar] [CrossRef] [PubMed]
- Baddour, L.M.; Baddour, L.M.; Wilson, W.R.; Bayer, A.S.; Fowler, V.G., Jr.; Tleyjeh, I.M.; Rybak, M.J.; Barsic, B.; Lockhart, P.B.; Gewitz, M.H.; et al. Infective Endocarditis in Adults: Diagnosis, Antimicrobial Therapy, and Management of Complications: A Scientific Statement for Healthcare Professionals From the American Heart Association. Circulation 2015, 132, 1435–1486. [Google Scholar] [CrossRef] [PubMed]
- Cremieux, A.C.; Maziere, B.; Vallois, J.M.; Ottaviani, M.; Azancot, A.; Raffoul, H.; Bouvet, A.; Pocidalo, J.J.; Carbon, C. Evaluation of antibiotic diffusion into cardiac vegetations by quantitative autoradiography. J. Infect Dis. 1989, 159, 938–944. [Google Scholar] [CrossRef]
- Levine, D.P.; Fromm, B.S.; Reddy, B.R. Slow response to vancomycin or vancomycin plus rifampin in methicillin-resistant Staphylococcus aureus endocarditis. Ann. Intern. Med. 1991, 115, 674–680. [Google Scholar] [CrossRef]
- Neely, M.N.; Youn, G.; Jones, B.; Jelliffe, R.W.; Drusano, G.L.; Rodvold, K.A.; Lodise, T.P. Are vancomycin trough concentrations adequate for optimal dosing? Antimicrob. Agents Chemother. 2014, 58, 309–316. [Google Scholar] [CrossRef]
- Pfeltz, R.F.; Wilkinson, B.J. The escalating challenge of vancomycin resistance in Staphylococcus aureus. Curr. Drug Targets Infect. Disord. 2004, 4, 273–294. [Google Scholar] [CrossRef]
- Sulakvelidze, A.; Alavidze, Z.; Morris, J.G., Jr. Bacteriophage therapy. Antimicrob. Agents Chemother. 2001, 45, 649–659. [Google Scholar] [CrossRef]
- Petrovic Fabijan, A.; Lin, R.C.; Ho, J.; Maddocks, S.; Ben Zakour, N.L.; Iredell, J.R. Safety of bacteriophage therapy in severe Staphylococcus aureus infection. Nat. Microbiol. 2020, 5, 465–472. [Google Scholar] [CrossRef]
- Gilbey, T.; Ho, J.; Cooley, L.A.; Fabijan, A.P.; Iredell, J.R. Adjunctive bacteriophage therapy for prosthetic valve endocarditis due to Staphylococcus aureus. Med. J. Aust. 2019, 211, 142–143.e1. [Google Scholar] [CrossRef]
- Moelling, K.; Broecker, F.; Willy, C. A Wake-Up Call: We Need Phage Therapy Now. Viruses 2018, 10, 688. [Google Scholar] [CrossRef] [PubMed]
- Pirnay, J.P.; Ferry, T.; Resch, G. Recent progress toward the implementation of phage therapy in Western medicine. FEMS Microbiol. Rev. 2022, 46, fuab040. [Google Scholar] [CrossRef] [PubMed]
- Valente, L.; Prazak, J.; Que, Y.A.; Cameron, D.R. Progress and Pitfalls of Bacteriophage Therapy in Critical Care: A Concise Definitive Review. Crit. Care Explor. 2021, 3, e0351. [Google Scholar] [CrossRef] [PubMed]
- Save, J.; Que, Y.A.; Entenza, J.M.; Kolenda, C.; Laurent, F.; Resch, G. Bacteriophages Combined With Subtherapeutic Doses of Flucloxacillin Act Synergistically Against Staphylococcus aureus Experimental Infective Endocarditis. J. Am. Heart Assoc. 2022, 11, e023080. [Google Scholar] [CrossRef] [PubMed]
- Santoro, J.; Levison, M.E. Rat model of experimental endocarditis. Infect. Immun. 1978, 19, 915–918. [Google Scholar] [CrossRef] [PubMed]
- Heraief, E.; Glauser, M.P.; Freedman, L.R. Natural history of aortic valve endocarditis in rats. Infect. Immun. 1982, 37, 127–131. [Google Scholar] [CrossRef]
- Cameron, D.R.; Ramette, A.; Prazak, J.; Entenza, J.; Haenggi, M.; Que, Y.A.; Resch, G. Draft Genome Sequence of Methicillin-Resistant Staphylococcus aureus Strain AW7, Isolated from a Patient with Bacteremia. Microbiol. Resour. Announc. 2019, 8, e00806–19. [Google Scholar] [CrossRef]
- Vouillamoz, J.; Entenza, J.M.; Feger, C.; Glauser, M.P.; Moreillon, P. Quinupristin-dalfopristin combined with beta-lactams for treatment of experimental endocarditis due to Staphylococcus aureus constitutively resistant to macrolide-lincosamide-streptogramin B antibiotics. Antimicrob. Agents Chemother. 2000, 44, 1789–1795. [Google Scholar] [CrossRef]
- Prazak, J.; Valente, L.G.; Iten, M.; Federer, L.; Grandgirard, D.; Soto, S.; Resch, G.; Leib, S.L.; Jakob, S.M.; Haenggi, M.; et al. Benefits of aerosolized phages for the treatment of pneumonia due to methicillin-resistant Staphylococcus aureus (MRSA): An experimental study in rats. J. Infect. Dis. 2022, 225, 1452–1459. [Google Scholar] [CrossRef]
- Valente, L.G.; Federer, L.; Iten, M.; Grandgirard, D.; Leib, S.L.; Jakob, S.M.; Haenggi, M.; Cameron, D.R.; Que, Y.A.; Prazak, J. Searching for synergy: Combining systemic daptomycin treatment with localised phage therapy for the treatment of experimental pneumonia due to MRSA. BMC Res. Notes 2021, 14, 381. [Google Scholar] [CrossRef]
- Prazak, J.; Iten, M.; Cameron, D.R.; Save, J.; Grandgirard, D.; Resch, G.; Goepfert, C.; Leib, S.L.; Takala, J.; Jakob, S.M.; et al. Bacteriophages Improve Outcomes in Experimental Staphylococcus aureus Ventilator-associated Pneumonia. Am. J. Respir. Crit. Care Med. 2019, 200, 1126–1133. [Google Scholar] [CrossRef] [PubMed]
- Bae, T.; Glass, E.M.; Schneewind, O.; Missiakas, D. Generating a collection of insertion mutations in the Staphylococcus aureus genome using bursa aurealis. Methods Mol. Biol. 2008, 416, 103–116. [Google Scholar] [PubMed]
- Resch, G.; François, P.; Morisset, D.; Stojanov, M.; Bonetti, E.J.; Schrenzel, J.; Sakwinska, O.; Moreillon, P. Human-to-bovine jump of Staphylococcus aureus CC8 is associated with the loss of a beta-hemolysin converting prophage and the acquisition of a new staphylococcal cassette chromosome. PLoS ONE 2013, 8, e58187. [Google Scholar] [CrossRef] [PubMed]
- CLSI. Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria that Grow Aerobically, 11th ed.; CLSI Standards M07; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2018. [Google Scholar]
- Chin, J.N.; Jones, R.N.; Sader, H.S.; Savage, P.B.; Rybak, M.J. Potential synergy activity of the novel ceragenin, CSA-13, against clinical isolates of Pseudomonas aeruginosa, including multidrug-resistant P.aeruginosa. J. Antimicrob. Chemother. 2008, 61, 365–370. [Google Scholar] [CrossRef] [PubMed]
- Que, Y.A.; Entenza, J.M.; Francioli, P.; Moreillon, P. The impact of penicillinase on cefamandole treatment and prophylaxis of experimental endocarditis due to methicillin-resistant Staphylococcus aureus. J. Infect. Dis. 1998, 177, 146–154. [Google Scholar] [CrossRef]
- Moreillon, P.; Bizzini, A.; Giddey, M.; Vouillamoz, J.; Entenza, J.M. Vancomycin-intermediate Staphylococcus aureus selected during vancomycin therapy of experimental endocarditis are not detected by culture-based diagnostic procedures and persist after treatment arrest. J. Antimicrob. Chemother. 2012, 67, 652–660. [Google Scholar] [CrossRef]
- Dell, R.B.; Holleran, S.; Ramakrishnan, R. Sample size determination. ILAR J. 2002, 43, 207–213. [Google Scholar] [CrossRef]
- Entenza, J.M.; Veloso, T.R.; Vouillamoz, J.; Giddey, M.; Moreillon, P. Failure of Vancomycin Continuous Infusion against Experimental Endocarditis Due to Vancomycin-Intermediate Staphylococcus aureus. Antimicrob. Agents Chemother. 2011, 55, 385–387. [Google Scholar] [CrossRef]
- Xia, G.; Corrigan, R.M.; Winstel, V.; Goerke, C.; Gründling, A.; Peschel, A. Wall Teichoic Acid-Dependent Adsorption of Staphylococcal Siphovirus and Myovirus. J. Bacteriol. 2011, 193, 4006–4009. [Google Scholar] [CrossRef]
- Li, X.; Gerlach, D.; Du, X.; Larsen, J.; Stegger, M.; Kühner, P.; Peschel, A.; Xia, G.; Winstel, V. An accessory wall teichoic acid glycosyltransferase protects Staphylococcus aureus from the lytic activity of Podoviridae. Sci. Rep. 2015, 5, 17219. [Google Scholar] [CrossRef]
- Henry, M.; Lavigne, R.; Debarbieux, L. Predicting In Vivo Efficacy of Therapeutic Bacteriophages Used To Treat Pulmonary Infections. Antimicrob. Agents Chemother. 2013, 57, 5961–5968. [Google Scholar] [CrossRef] [PubMed]
- Abedon, S.T. Phage therapy dosing: The problem(s) with multiplicity of infection (MOI). Bacteriophage 2016, 6, e1220348. [Google Scholar] [CrossRef] [PubMed]
- Abedon, S.T. Phage-Antibiotic Combination Treatments: Antagonistic Impacts of Antibiotics on the Pharmacodynamics of Phage Therapy? Antibiotics 2019, 8, 182. [Google Scholar] [CrossRef] [PubMed]
- Torres-Barceló, C.; Arias-Sánchez, F.I.; Vasse, M.; Ramsayer, J.; Kaltz, O.; Hochberg, M.E. A Window of Opportunity to Control the Bacterial Pathogen Pseudomonas aeruginosa Combining Antibiotics and Phages. PLoS ONE 2014, 9, e106628. [Google Scholar] [CrossRef] [PubMed]
- Uchiyama, J.; Taniguchi, M.; Kurokawa, K.; Takemura-Uchiyama, I.; Ujihara, T.; Shimakura, H.; Sakaguchi, Y.; Murakami, H.; Sakaguchi, M.; Matsuzaki, S. Adsorption of Staphylococcus viruses S13′ and S24-1 on Staphylococcus aureus strains with different glycosidic linkage patterns of wall teichoic acids. J. Gen. Virol. 2017, 98, 2171–2180. [Google Scholar] [CrossRef] [PubMed]
- Ingmer, H.; Gerlach, D.; Wolz, C. Temperate Phages of Staphylococcus aureus. Microbiol. Spectr. 2019, 7, 7–5. [Google Scholar] [CrossRef]
- Mistretta, N.; Brossaud, M.; Telles, F.; Sanchez, V.; Talaga, P.; Rokbi, B. Glycosylation of Staphylococcus aureus cell wall teichoic acid is influenced by environmental conditions. Sci. Rep. 2019, 9, 3212. [Google Scholar] [CrossRef]
- Doub, J.B.; Urish, K.; Lee, M.; Fackler, J. Impact of Bacterial Phenotypic Variation with Bacteriophage therapy: A Pilot Study with Prosthetic Joint Infection Isolates. Int. J. Infect. Dis. 2022, 119, 44–46. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Save, J.; Que, Y.-A.; Entenza, J.; Resch, G. Subtherapeutic Doses of Vancomycin Synergize with Bacteriophages for Treatment of Experimental Methicillin-Resistant Staphylococcus aureus Infective Endocarditis. Viruses 2022, 14, 1792. https://doi.org/10.3390/v14081792
Save J, Que Y-A, Entenza J, Resch G. Subtherapeutic Doses of Vancomycin Synergize with Bacteriophages for Treatment of Experimental Methicillin-Resistant Staphylococcus aureus Infective Endocarditis. Viruses. 2022; 14(8):1792. https://doi.org/10.3390/v14081792
Chicago/Turabian StyleSave, Jonathan, Yok-Ai Que, José Entenza, and Grégory Resch. 2022. "Subtherapeutic Doses of Vancomycin Synergize with Bacteriophages for Treatment of Experimental Methicillin-Resistant Staphylococcus aureus Infective Endocarditis" Viruses 14, no. 8: 1792. https://doi.org/10.3390/v14081792