
Evidence of RedOX Imbalance during Zika Virus Infection Promoting the Formation of Disulfide-Bond-Dependent Oligomers of the Envelope Protein

 , , , , and
, , , , and 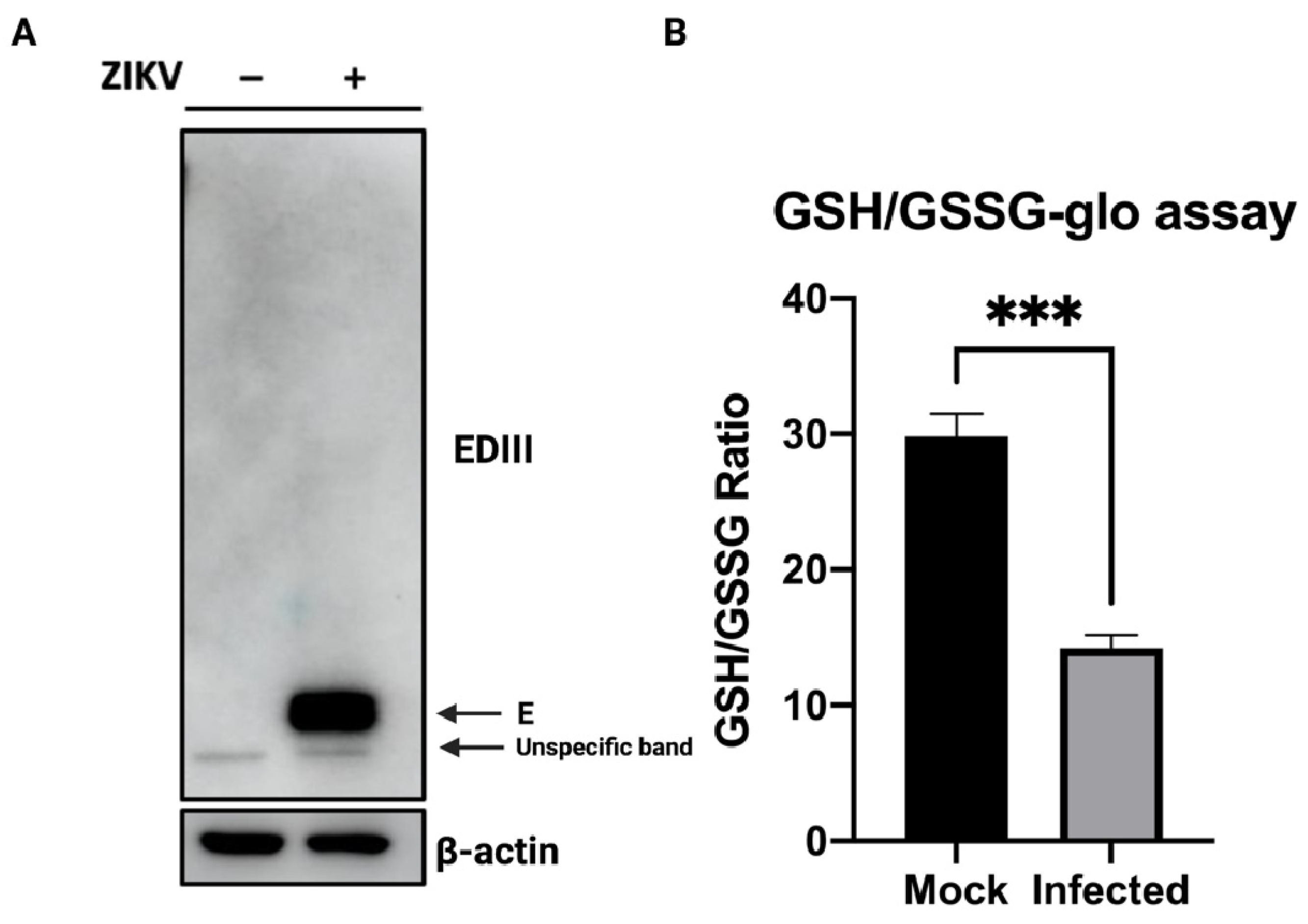{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells, Virus and Reagents
2.2. Cell Fractionation and Western Blot
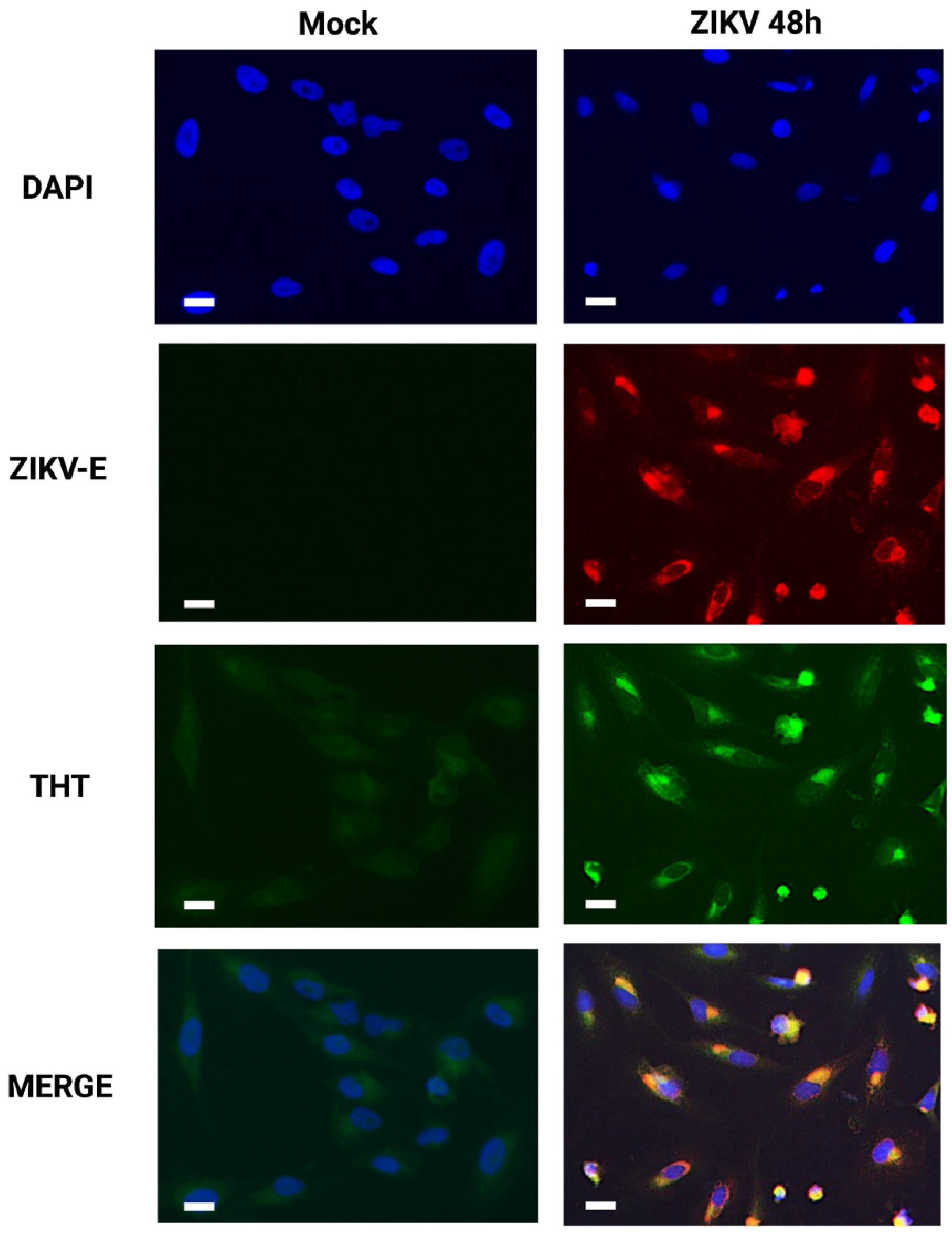
2.3. Immunofluorescence and Thioflavin T Staining
2.4. Glutathione Abundance Measurement
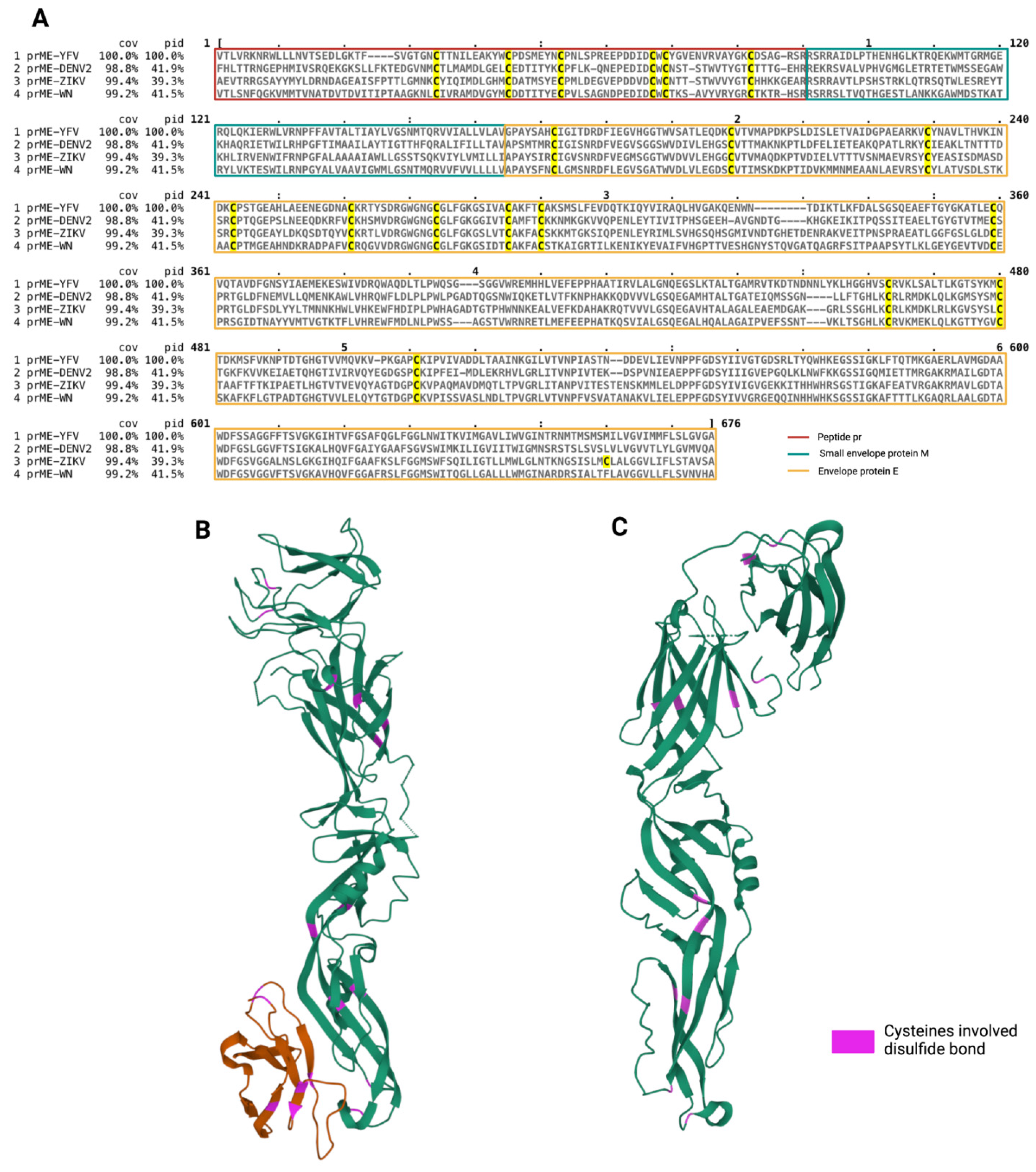
2.5. Protein Sequence Alignment
2.6. Statistical Analyses
3. Results
3.1. Glutathione Abundance in the Course of ZIKV Infection
3.2. Detection of Amyloid-like Structures in ZIKV-Infected Cells
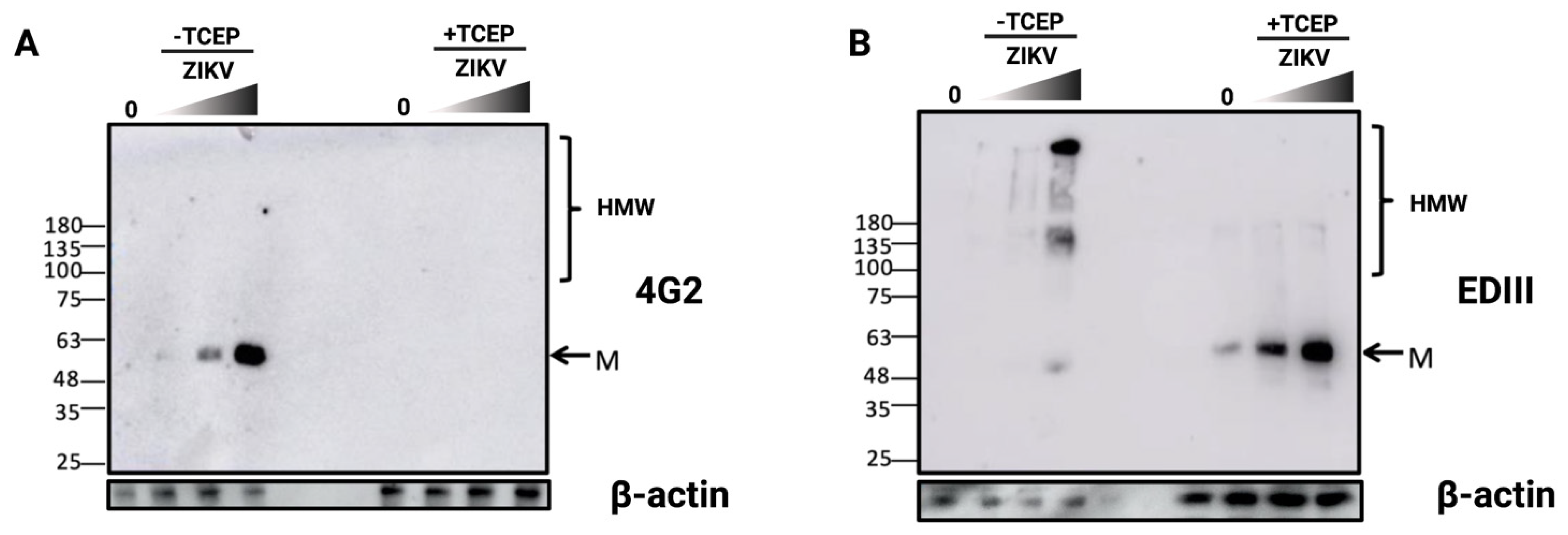
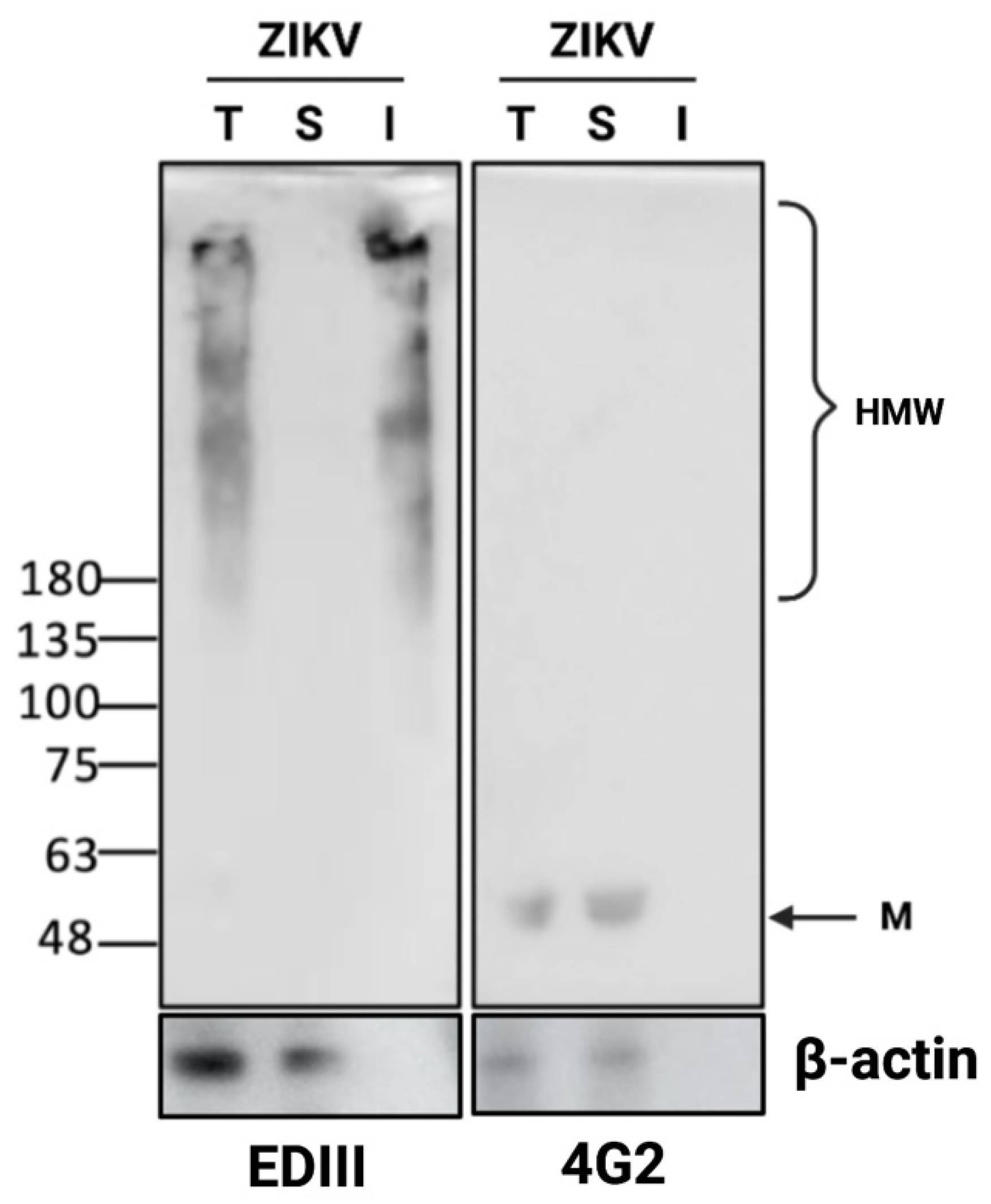
3.3. Detection of Disulfide-Bonded E Oligomers in ZIKV-Infected Cells
3.4. ZIKV E Protein Forms Insoluble Oligomers during Infection
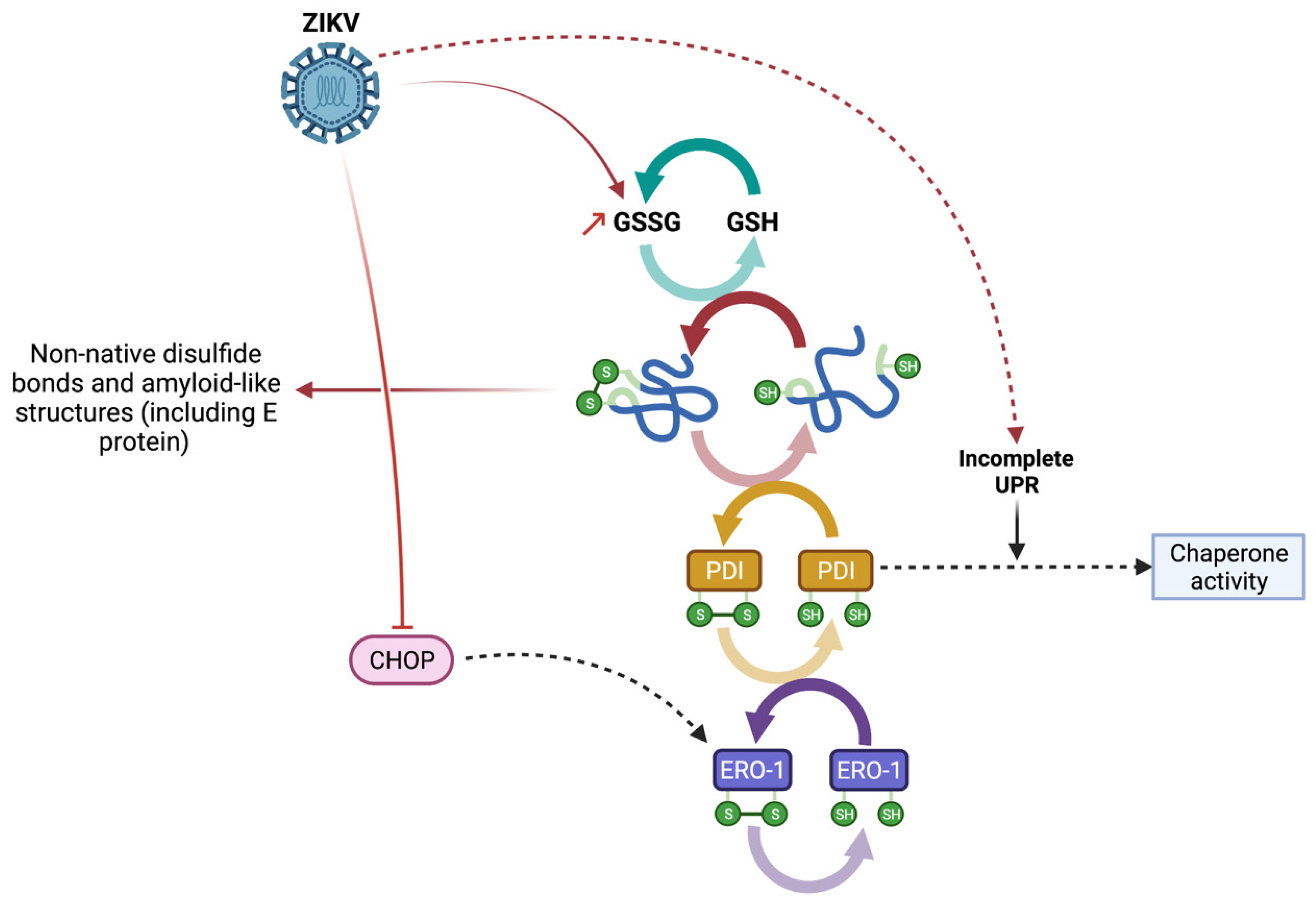
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| CHOP | C/EBP Homologous Protein |
| DENV | Dengue Virus |
| EDIII | Domain III of E protein |
| ER | Endoplasmic Reticulum |
| ERO-1 | ER oxidoreductin 1 |
| GSH | Reduced glutathione |
| GSSG | Oxidized glutathione |
| PDI | Protein Disulfide Isomerase |
| TCEP | Tris (2-carboxyethyl)phosphine |
| THT | Thioflavin T |
| UPR | Unfolded Protein Response |
| WNV | West Nile Virus |
| YFV | Yellow |
| ZIKV | Zika Virus |
| ZIKV-E | Zika Virus envelope protein |
References
- Wen, Z.; Song, H.; Ming, G. How Does Zika Virus Cause Microcephaly? Genes Dev. 2017, 31, 849–861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miranda, J.; Martín-Tapia, D.; Valdespino-Vázquez, Y.; Alarcón, L.; Espejel-Nuñez, A.; Guzmán-Huerta, M.; Muñoz-Medina, J.E.; Shibayama, M.; Chávez-Munguía, B.; Estrada-Gutiérrez, G.; et al. Syncytiotrophoblast of Placentae from Women with Zika Virus Infection Has Altered Tight Junction Protein Expression and Increased Paracellular Permeability. Cells 2019, 8, 1174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cortese, M.; Goellner, S.; Acosta, E.G.; Neufeldt, C.J.; Oleksiuk, O.; Lampe, M.; Haselmann, U.; Funaya, C.; Schieber, N.; Ronchi, P.; et al. Ultrastructural Characterization of Zika Virus Replication Factories. Cell Rep. 2017, 18, 2113–2123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miorin, L.; Romero-Brey, I.; Maiuri, P.; Hoppe, S.; Krijnse-Locker, J.; Bartenschlager, R.; Marcello, A. Three-Dimensional Architecture of Tick-Borne Encephalitis Virus Replication Sites and Trafficking of the Replicated RNA. J. Virol. 2013, 87, 6469–6481. [Google Scholar] [CrossRef] [Green Version]
- Romero-Brey, I.; Bartenschlager, R. Membranous Replication Factories Induced by Plus-Strand RNA Viruses. Viruses 2014, 6, 2826–2857. [Google Scholar] [CrossRef]
- Sun, Z.; Brodsky, J.L. Protein Quality Control in the Secretory Pathway. J. Cell Biol. 2019, 218, 3171–3187. [Google Scholar] [CrossRef] [Green Version]
- Hendershot, L.M. The ER Function BiP Is a Master Regulator of ER Function. Mt. Sinai J. Med. 2004, 71, 289–297. [Google Scholar]
- Xu, C.; Bailly-Maitre, B.; Reed, J.C. Endoplasmic Reticulum Stress: Cell Life and Death Decisions. J. Clin. Investig. 2005, 115, 2656–2664. [Google Scholar] [CrossRef] [Green Version]
- Smith, J.A. A New Paradigm: Innate Immune Sensing of Viruses via the Unfolded Protein Response. Front. Microbiol. 2014, 5, 222. [Google Scholar] [CrossRef]
- Carletti, T.; Zakaria, M.K.; Faoro, V.; Reale, L.; Kazungu, Y.; Licastro, D.; Marcello, A. Viral Priming of Cell Intrinsic Innate Antiviral Signaling by the Unfolded Protein Response. Nat. Commun. 2019, 10, 3889. [Google Scholar] [CrossRef] [Green Version]
- Blázquez, A.-B.; Escribano-Romero, E.; Merino-Ramos, T.; Saiz, J.-C.; Martín-Acebes, M.A. Stress Responses in Fla-Vivirus-Infected Cells: Activation of Unfolded Protein Response and Autophagy. Front. Microbiol. 2014, 5, 266. [Google Scholar] [CrossRef] [PubMed]
- Sano, R.; Reed, J.C. ER Stress-Induced Cell Death Mechanisms. Biochim. Biophys. Acta (BBA)-Mol. Cell Res. 2013, 1833, 3460–3470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gladwyn-Ng, I.; Cordón-Barris, L.; Alfano, C.; Creppe, C.; Couderc, T.; Morelli, G.; Thelen, N.; America, M.; Bessières, B.; Encha-Razavi, F.; et al. Stress-Induced Unfolded Protein Response Contributes to Zika Virus–Associated Microcephaly. Nat. Neurosci. 2018, 21, 63–71. [Google Scholar] [CrossRef] [PubMed]
- Alfano, C.; Gladwyn-Ng, I.; Couderc, T.; Lecuit, M.; Nguyen, L. The Unfolded Protein Response: A Key Player in Zika Virus-Associated Congenital Microcephaly. Front. Cell. Neurosci. 2019, 13, 94. [Google Scholar] [CrossRef]
- Hou, S.; Kumar, A.; Xu, Z.; Airo, A.M.; Stryapunina, I.; Wong, C.P.; Branton, W.; Tchesnokov, E.; Götte, M.; Power, C.; et al. Zika Virus Hijacks Stress Granule Proteins and Modulates the Host Stress Response. J. Virol. 2017, 91, e00474-17. [Google Scholar] [CrossRef] [Green Version]
- Amorim, R.; Temzi, A.; Griffin, B.D.; Mouland, A.J. Zika Virus Inhibits EIF2α-Dependent Stress Granule Assembly. PLoS Negl. Trop. Dis. 2017, 11, e0005775. [Google Scholar] [CrossRef] [Green Version]
- Bonenfant, G.; Williams, N.; Netzband, R.; Schwarz, M.C.; Evans, M.J.; Pager, C.T. Zika Virus Subverts Stress Granules to Promote and Restrict Viral Gene Expression. J. Virol. 2019, 93, e00520-19. [Google Scholar] [CrossRef] [Green Version]
- Turpin, J.; Frumence, E.; Harrabi, W.; Haddad, J.G.; El Kalamouni, C.; Desprès, P.; Krejbich-Trotot, P.; Viranaïcken, W. Zika Virus Subversion of Chaperone GRP78/BiP Expression in A549 Cells during UPR Activation. Biochimie 2020, 175, 99–105. [Google Scholar] [CrossRef]
- Turpin, J.; El-Safadi, D.; Lebeau, G.; Frumence, E.; Desprès, P.; Viranaïcken, W.; Krejbich-Trotot, P. CHOP Pro-Apoptotic Transcriptional Program in Response to ER Stress Is Hacked by Zika Virus. IJMS 2021, 22, 3750. [Google Scholar] [CrossRef]
- Monel, B.; Compton, A.A.; Bruel, T.; Amraoui, S.; Burlaud-Gaillard, J.; Roy, N.; Guivel-Benhassine, F.; Porrot, F.; Génin, P.; Meertens, L.; et al. Zika Virus Induces Massive Cytoplasmic Vacuolization and Paraptosis-like Death in Infected Cells. EMBO J. 2017, 36, 1653–1668. [Google Scholar] [CrossRef]
- Ci, Y.; Liu, Z.-Y.; Zhang, N.-N.; Niu, Y.; Yang, Y.; Xu, C.; Yang, W.; Qin, C.-F.; Shi, L. Zika NS1–Induced ER Remodeling Is Essential for Viral Replication. J. Cell Biol. 2020, 219, e201903062. [Google Scholar] [CrossRef] [PubMed]
- Turpin, J.; El Safadi, D.; Lebeau, G.; Krejbich, M.; Chatelain, C.; Desprès, P.; Viranaïcken, W.; Krejbich-Trotot, P. Apoptosis during ZIKA Virus Infection: Too Soon or Too Late? Int. J. Mol. Sci. 2022, 23, 1287. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Kwon, D.; Kwak, J.-H.; Lee, S.; Lee, Y.-H.; Yun, J.; Son, T.; Jung, Y.-S. Tunicamycin-Induced ER Stress Is Accompanied with Oxidative Stress via Abrogation of Sulfur Amino Acids Metabolism in the Liver. Int. J. Mol. Sci. 2018, 19, 4114. [Google Scholar] [CrossRef] [Green Version]
- Dickinson, D.A.; Forman, H.J. Glutathione in Defense and Signaling: Lessons from a Small Thiol. Ann. N. Y. Acad. Sci. 2002, 973, 488–504. [Google Scholar] [CrossRef] [PubMed]
- Grek, C.; Townsend, D.M. Protein Disulfide Isomerase Superfamily in Disease and the Regulation of Apoptosis. Endoplasmic Reticulum Stress Dis. 2014, 1, 4–17. [Google Scholar] [CrossRef] [PubMed]
- Appenzeller-Herzog, C. Glutathione- and Non-Glutathione-Based Oxidant Control in the Endoplasmic Reticulum. J. Cell Sci. 2011, 124, 847–855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, C.; Sinskey, A.J.; Lodish, H.F. Oxidized Redox State of Glutathione in the Endoplasmic Reticulum. Science 1992, 257, 1496–1502. [Google Scholar] [CrossRef]
- Woycechowsky, K.J.; Raines, R.T. Native Disulfide Bond Formation in Proteins. Curr. Opin. Chem. Biol. 2000, 4, 533–539. [Google Scholar] [CrossRef]
- Frumence, E.; Roche, M.; Krejbich-Trotot, P.; El-Kalamouni, C.; Nativel, B.; Rondeau, P.; Missé, D.; Gadea, G.; Viranaicken, W.; Desprès, P. The South Pacific Epidemic Strain of Zika Virus Replicates Efficiently in Human Epithelial A549 Cells Leading to IFN-β Production and Apoptosis Induction. Virology 2016, 493, 217–226. [Google Scholar] [CrossRef]
- Viranaicken, W.; Nativel, B.; Krejbich-Trotot, P.; Harrabi, W.; Bos, S.; El Kalamouni, C.; Roche, M.; Gadea, G.; Desprès, P. ClearColi BL21(DE3)-Based Expression of Zika Virus Antigens Illustrates a Rapid Method of Antibody Production against Emerging Pathogens. Biochimie 2017, 142, 179–182. [Google Scholar] [CrossRef]
- Mattioli, L.; Anelli, T.; Fagioli, C.; Tacchetti, C.; Sitia, R.; Valetti, C. ER Storage Diseases: A Role for ERGIC-53 in Controlling the Formation and Shape of Russell Bodies. J. Cell Sci. 2006, 119, 2532–2541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Viranaicken, W.; Gasmi, L.; Chaumet, A.; Durieux, C.; Georget, V.; Denoulet, P.; Larcher, J.-C. L-Ilf3 and L-NF90 Traffic to the Nucleolus Granular Component: Alternatively-Spliced Exon 3 Encodes a Nucleolar Localization Motif. PLoS ONE 2011, 6, e22296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beriault, D.R.; Werstuck, G.H. Detection and Quantification of Endoplasmic Reticulum Stress in Living Cells Using the Fluorescent Compound, Thioflavin, T. Biochim. Biophys. Acta 2013, 1833, 2293–2301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Zhang, L.; Zhou, L.; Lei, Y.; Zhang, Y.; Huang, C. Redox Signaling and Unfolded Protein Response Coordinate Cell Fate Decisions under ER Stress. Redox Biol. 2019, 25, 101047. [Google Scholar] [CrossRef] [PubMed]
- Zeeshan, H.; Lee, G.; Kim, H.-R.; Chae, H.-J. Endoplasmic Reticulum Stress and Associated ROS. Int. J. Mol. Sci. 2016, 17, 327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mackenzie, J.M.; Westaway, E.G. Assembly and Maturation of the Flavivirus Kunjin Virus Appear to Occur in the Rough Endoplasmic Reticulum and along the Secretory Pathway, Respectively. J. Virol. 2001, 75, 10787–10799. [Google Scholar] [CrossRef] [Green Version]
- Costin, J.M.; Zaitseva, E.; Kahle, K.M.; Nicholson, C.O.; Rowe, D.K.; Graham, A.S.; Bazzone, L.E.; Hogancamp, G.; Figueroa Sierra, M.; Fong, R.H.; et al. Mechanistic Study of Broadly Neutralizing Human Monoclonal Antibodies against Dengue Virus That Target the Fusion Loop. J. Virol. 2013, 87, 52–66. [Google Scholar] [CrossRef] [Green Version]
- Roehrig, J.T.; Volpe, K.E.; Squires, J.; Hunt, A.R.; Davis, B.S.; Chang, G.-J.J. Contribution of Disulfide Bridging to Epitope Expression of the Dengue Type 2 Virus Envelope Glycoprotein. J. Virol. 2004, 78, 2648–2652. [Google Scholar] [CrossRef] [Green Version]
- Sirohi, D.; Chen, Z.; Sun, L.; Klose, T.; Pierson, T.C.; Rossmann, M.G.; Kuhn, R.J. The 3.8 Å Resolution Cryo-EM Structure of Zika Virus. Science 2016, 352, 467–470. [Google Scholar] [CrossRef] [Green Version]
- Nowak, T.; Wengler, G. Analysis of Disulfides Present in the Membrane Proteins of the West Nile Flavivirus. Virology 1987, 156, 127–137. [Google Scholar] [CrossRef]
- Chiti, F.; Dobson, C.M. Protein Misfolding, Amyloid Formation, and Human Disease: A Summary of Progress Over the Last Decade. Annu. Rev. Biochem. 2017, 86, 27–68. [Google Scholar] [CrossRef] [PubMed]
- Aoyama, K.; Nakaki, T. Glutathione in Cellular Redox Homeostasis: Association with the Excitatory Amino Acid Carrier 1 (EAAC1). Molecules 2015, 20, 8742–8758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merksamer, P.I.; Trusina, A.; Papa, F.R. Real-Time Redox Measurements during Endoplasmic Reticulum Stress Reveal Interlinked Protein Folding Functions. Cell 2008, 135, 933–947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shergalis, A.G.; Hu, S.; Bankhead, A.; Neamati, N. Role of the ERO1-PDI Interaction in Oxidative Protein Folding and Disease. Pharmacol. Ther. 2020, 210, 107525. [Google Scholar] [CrossRef]
- Parakh, S.; Atkin, J.D. Novel Roles for Protein Disulphide Isomerase in Disease States: A Double Edged Sword? Front. Cell Dev. Biol. 2015, 3, 30. [Google Scholar] [CrossRef] [Green Version]
- Lingel, A.; Lin, H.; Gavriel, Y.; Weaver, E.; Polepole, P.; Lopez, V.; Lei, Y.; Petro, T.M.; Solomon, B.; Zhang, C.; et al. Amyloid Precursor Protein Is a Restriction Factor That Protects against Zika Virus Infection in Mammalian Brains. J. Biol. Chem. 2020, 295, 17114–17127. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lebeau, G.; Turpin, J.; Frumence, E.; El Safadi, D.; Harrabi, W.; Desprès, P.; Krejbich-Trotot, P.; Viranaïcken, W. Evidence of RedOX Imbalance during Zika Virus Infection Promoting the Formation of Disulfide-Bond-Dependent Oligomers of the Envelope Protein. Viruses 2022, 14, 1131. https://doi.org/10.3390/v14061131
Lebeau G, Turpin J, Frumence E, El Safadi D, Harrabi W, Desprès P, Krejbich-Trotot P, Viranaïcken W. Evidence of RedOX Imbalance during Zika Virus Infection Promoting the Formation of Disulfide-Bond-Dependent Oligomers of the Envelope Protein. Viruses. 2022; 14(6):1131. https://doi.org/10.3390/v14061131
Chicago/Turabian StyleLebeau, Grégorie, Jonathan Turpin, Etienne Frumence, Daed El Safadi, Wissal Harrabi, Philippe Desprès, Pascale Krejbich-Trotot, and Wildriss Viranaïcken. 2022. "Evidence of RedOX Imbalance during Zika Virus Infection Promoting the Formation of Disulfide-Bond-Dependent Oligomers of the Envelope Protein" Viruses 14, no. 6: 1131. https://doi.org/10.3390/v14061131