
Dynamics of Antibodies to Ebolaviruses in an Eidolon helvum Bat Colony in Cameroon

 , , , , , , , ,
, , , , , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Collection Sites
2.2. Molecular Confirmation of Bat Species
2.3. Screening for Ebolavirus Antibodies
2.4. Nucleic Acid Extraction and PCR Screening for Detection of Filoviruses
2.5. Logistic Regression for Seropositivity
3. Results
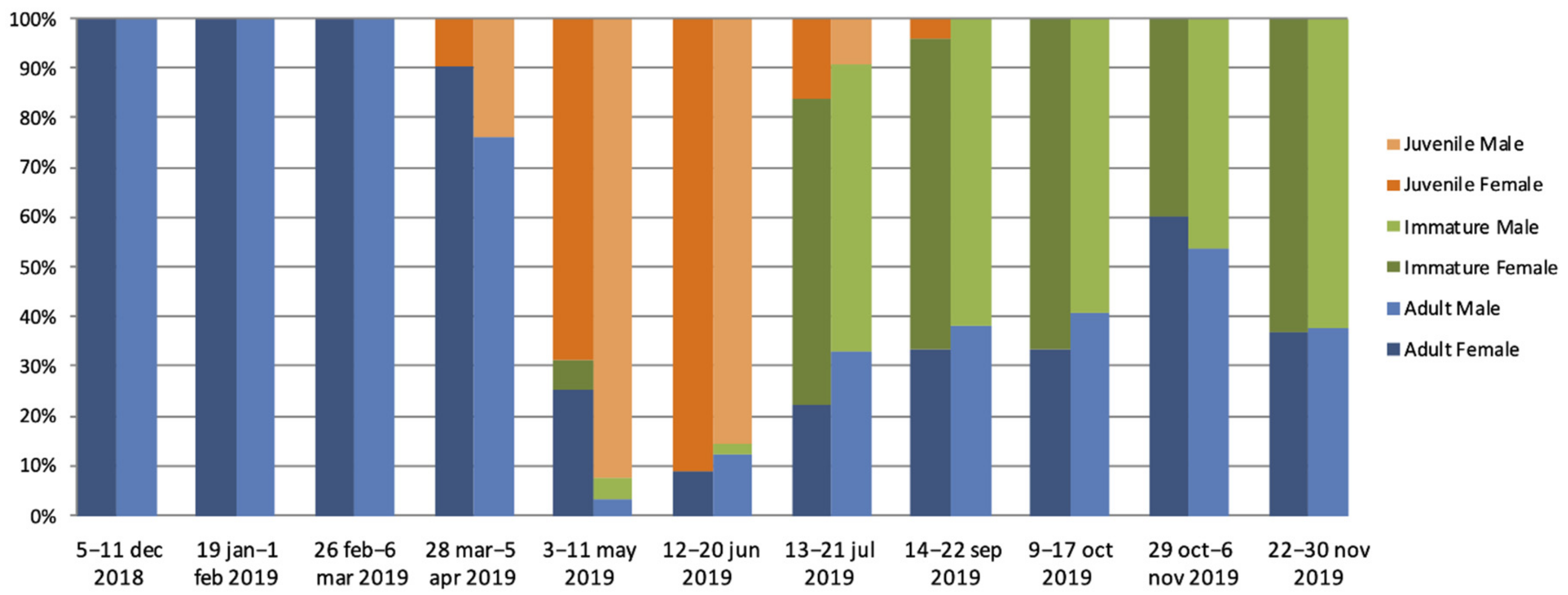
3.1. Eidolon Helvum Population
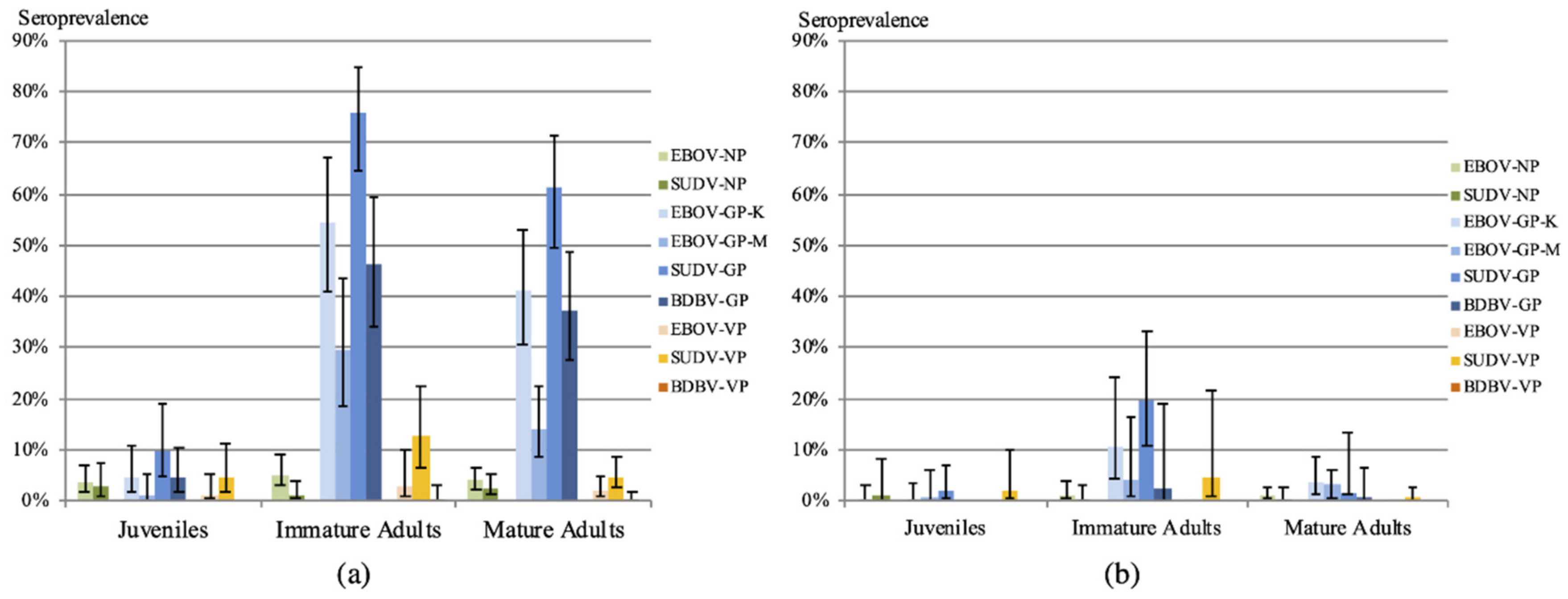
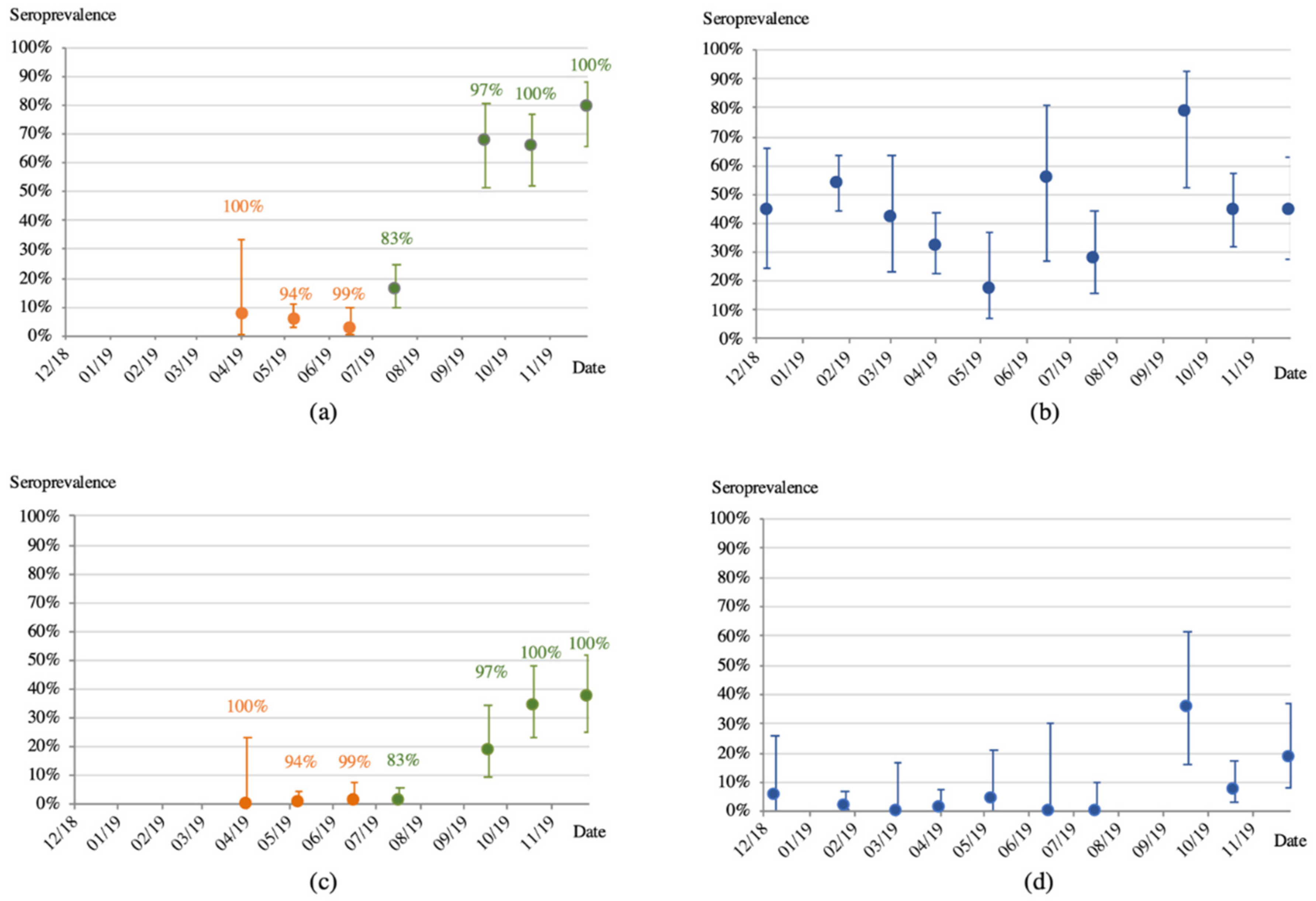
3.2. Antibodies to Ebolaviruses
3.3. Molecular Screening for the Presence of Ebolaviruses
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Malvy, D.; McElroy, A.K.; de Clerck, H.; Günther, S.; van Griensven, J. Ebola virus disease. Lancet 2019, 393, 936–948. [Google Scholar] [CrossRef] [Green Version]
- Center for Disease Control (CDC). History of Ebola Virus Disease Outbreaks. Available online: https://www.cdc.gov/vhf/ebola/history/chronology.html#anchor_1526565058132 (accessed on 6 February 2022).
- LaBrunda, M.; Amin, N. The Emerging Threat of Ebola. In Global Health Security; Springer: Cham, Switzerland, 2020; pp. 103–139. [Google Scholar] [CrossRef] [Green Version]
- Mbala-Kingebeni, P.; Pratt, C.; Mutafali-Ruffin, M.; Pauthner, M.G.; Bile, F.; Nkuba-Ndaye, A.; Black, A.; Kinganda-Lusamaki, E.; Faye, M.; Aziza, A.; et al. Ebola Virus Transmission Initiated by Relapse of Systemic Ebola Virus Disease. N. Engl. J. Med. 2021, 384, 1240–1247. [Google Scholar] [CrossRef] [PubMed]
- Keita, A.K.; Koundouno, F.R.; Faye, M.; Düx, A.; Hinzmann, J.; Diallo, H.; Ayouba, A.; Le Marcis, F.; Soropogui, B.; Ifono, K.; et al. Resurgence of Ebola virus in 2021 in Guinea suggests a new paradigm for outbreaks. Nature 2021, 597, 539–543. [Google Scholar] [CrossRef] [PubMed]
- Pigott, D.M.; Golding, N.; Mylne, A.; Huang, Z.; Henry, A.J.; Weiss, D.J.; Brady, O.; Kraemer, M.U.G.; Smith, D.L.; Moyes, C.; et al. Mapping the zoonotic niche of Ebola virus disease in Africa. eLife 2014, 3, e04395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leroy, E.M.; Kumulungui, B.; Pourrut, X.; Rouquet, P.; Hassanin, A.; Yaba, P.; Délicat, A.; Paweska, J.T.; Gonzalez, J.-P.; Swanepoel, R. Fruit bats as reservoirs of Ebola virus. Nature 2005, 438, 575–576. [Google Scholar] [CrossRef] [PubMed]
- Pourrut, X.; Souris, M.; Towner, J.S.; Rollin, P.E.; Nichol, S.T.; Gonzalez, J.-P.; Leroy, E. Large serological survey showing cocirculation of Ebola and Marburg viruses in Gabonese bat populations, and a high seroprevalence of both viruses in Rousettus aegyptiacus. BMC Infect. Dis. 2009, 9, 159. [Google Scholar] [CrossRef] [Green Version]
- Hayman, D.T.; Yu, M.; Crameri, G.; Wang, L.-F.; Suu-Ire, R.; Wood, J.L.; Cunningham, A.A. Ebola Virus Antibodies in Fruit Bats, Ghana, West Africa. Emerg. Infect. Dis. 2012, 18, 1207–1209. [Google Scholar] [CrossRef]
- Ogawa, H.; Miyamoto, H.; Nakayama, E.; Yoshida, R.; Nakamura, I.; Sawa, H.; Ishii, A.; Thomas, Y.; Nakagawa, E.; Matsuno, K.; et al. Seroepidemiological Prevalence of Multiple Species of Filoviruses in Fruit Bats (Eidolon helvum) Migrating in Africa. J. Infect. Dis. 2015, 212 (Suppl. S2), S101–S108. [Google Scholar] [CrossRef]
- De Nys, H.M.; Kingebeni, P.M.; Keita, A.K.; Butel, C.; Thaurignac, G.; Arenas, C.J.V.; Lemarcis, T.; Geraerts, M.; Al, H.M.D.N.E.; Esteban, A.; et al. Survey of Ebola Viruses in Frugivorous and Insectivorous Bats in Guinea, Cameroon, and the Democratic Republic of the Congo, 2015–2017. Emerg. Infect. Dis. 2018, 24, 2228–2240. [Google Scholar] [CrossRef]
- Leroy, E.M.; Epelboin, A.; Mondonge, V.; Pourrut, X.; Gonzalez, J.-P.; Muyembe-Tamfum, J.-J.; Formenty, P. Human Ebola Outbreak Resulting from Direct Exposure to Fruit Bats in Luebo, Democratic Republic of Congo, 2007. Vector Borne Zoonotic Dis. 2009, 9, 723–728. [Google Scholar] [CrossRef]
- Saéz, A.M.; Weiss, S.; Nowak, K.; Lapeyre, V.; Zimmermann, F.; Düx, A.; Kühl, H.S.; Kaba, M.; Regnaut, S.; Merkel, K.; et al. Investigating the zoonotic origin of the West African Ebola epidemic. EMBO Mol. Med. 2015, 7, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Amman, B.R.; Bird, B.H.; Bakarr, I.A.; Bangura, J.; Schuh, A.J.; Johnny, J.; Sealy, T.K.; Conteh, I.; Koroma, A.H.; Foday, I.; et al. Isolation of Angola-like Marburg virus from Egyptian rousette bats from West Africa. Nat. Commun. 2020, 11, 510. [Google Scholar] [CrossRef] [PubMed]
- Towner, J.S.; Pourrut, X.; Albariño, C.G.; Nkogue, C.N.; Bird, B.H.; Grard, G.; Ksiazek, T.G.; Gonzalez, J.-P.; Nichol, S.T.; Leroy, E.M. Marburg Virus Infection Detected in a Common African Bat. PLoS ONE 2007, 2, e764. [Google Scholar] [CrossRef] [PubMed]
- Towner, J.S.; Amman, B.R.; Sealy, T.K.; Carroll, S.A.R.; Comer, J.A.; Kemp, A.; Swanepoel, R.; Paddock, C.D.; Balinandi, S.; Khristova, M.L.; et al. Isolation of Genetically Diverse Marburg Viruses from Egyptian Fruit Bats. PLoS Pathog. 2009, 5, e1000536. [Google Scholar] [CrossRef]
- Goldstein, T.; Anthony, S.J.; Gbakima, A.; Bird, B.H.; Bangura, J.; Tremeau-Bravard, A.; Belaganahalli, M.N.; Wells, H.L.; Dhanota, J.K.; Liang, E.; et al. The discovery of Bombali virus adds further support for bats as hosts of ebolaviruses. Nat. Microbiol. 2018, 3, 1084–1089. [Google Scholar] [CrossRef]
- Forbes, K.M.; Al, K.M.F.E.; Jääskeläinen, A.J.; Abdurahman, S.; Ogola, J.; Masika, M.M.; Kivistö, I.; Alburkat, H.; Plyusnin, I.; Levanov, L.; et al. Bombali Virus in Mops condylurus Bat, Kenya. Emerg. Infect. Dis. 2019, 25, 955–957. [Google Scholar] [CrossRef] [Green Version]
- Karan, L.S.; Makenov, M.T.; Korneev, M.G.; Sacko, N.; Boumbaly, S.; Yakovlev, S.A.; Kourouma, K.; Bayandin, R.B.; Gladysheva, A.V.; Shipovalov, A.V.; et al. Bombali Virus in Mops condylurus Bats, Guinea. Emerg. Infect. Dis. 2019, 25, 1774–1775. [Google Scholar] [CrossRef] [Green Version]
- Negredo, A.; Palacios, G.; Vázquez-Morón, S.; González, F.; Dopazo, H.; Molero, F.; Juste, J.; Quetglas, J.; Savji, N.; Martínez, M.D.L.C.; et al. Discovery of an Ebolavirus-Like Filovirus in Europe. PLOS Pathog. 2011, 7, e1002304. [Google Scholar] [CrossRef] [Green Version]
- Kemenesi, G.; Kurucz, K.; Dallos, B.; Zana, B.; Földes, F.; Boldogh, S.; Görföl, T.; Carroll, M.W.; Jakab, F. Re-emergence of Lloviu virus in Miniopterus schreibersii bats, Hungary, 2016. Emerg. Microbes Infect. 2018, 7, 1–4. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.-L.; Zhang, Y.-Z.; Jiang, R.-D.; Guo, H.; Zhang, W.; Li, B.; Wang, N.; Wang, L.; Waruhiu, C.; Zhou, J.-H.; et al. Genetically Diverse Filoviruses in Rousettus and Eonycteris spp. Bats, China, 2009 and 2015. Emerg. Infect. Dis. 2017, 23, 482–486. [Google Scholar] [CrossRef]
- Jayme, S.I.; Field, H.E.; de Jong, C.; Olival, K.J.; Marsh, G.; Tagtag, A.M.; Hughes, T.; Bucad, A.C.; Barr, J.; Azul, R.R.; et al. Molecular evidence of Ebola Reston virus infection in Philippine bats. Virol. J. 2015, 12, 107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paweska, J.T.; Van Vuren, P.J.; Masumu, J.; Leman, P.A.; Grobbelaar, A.A.; Birkhead, M.; Clift, S.; Swanepoel, R.; Kemp, A. Virological and Serological Findings in Rousettus aegyptiacus Experimentally Inoculated with Vero Cells-Adapted Hogan Strain of Marburg Virus. PLoS ONE 2012, 7, e45479. [Google Scholar] [CrossRef] [PubMed]
- Amman, B.R.; Jones, M.E.B.; Sealy, T.K.; Uebelhoer, L.S.; Schuh, A.J.; Bird, B.H.; Coleman-McCray, J.D.; Martin, B.E.; Nichol, S.T.; Towner, J.S. Oral shedding of marburg virus in experimentally infected egyptian fruit bats (Rousettus aegyptiacus). J. Wildl. Dis. 2015, 51, 113–124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pourrut, X.; Délicat, A.; Rollin, P.; Ksiazek, T.G.; Gonzalez, J.-P.; Leroy, E.M. Spatial and Temporal Patterns of Zaire ebolavirus Antibody Prevalence in the Possible Reservoir Bat Species. J. Infect. Dis. 2007, 196 (Suppl. S2), S176–S183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baudel, H.; De Nys, H.; Ngole, E.M.; Peeters, M.; Desclaux, A. Understanding Ebola virus and other zoonotic transmission risks through human–bat contacts: Exploratory study on knowledge, attitudes and practices in Southern Cameroon. Zoonoses Public Health 2019, 66, 288–295. [Google Scholar] [CrossRef] [PubMed]
- Kamins, A.; Restif, O.; Ntiamoa-Baidu, Y.; Suu-Ire, R.; Hayman, D.; Cunningham, A.; Wood, J.; Rowcliffe, M. Uncovering the fruit bat bushmeat commodity chain and the true extent of fruit bat hunting in Ghana, West Africa. Biol. Conserv. 2011, 144, 3000–3008. [Google Scholar] [CrossRef] [Green Version]
- Richter, H.V.; Cumming, G.S. First application of satellite telemetry to track African straw-coloured fruit bat migration. J. Zool. 2008, 275, 172–176. [Google Scholar] [CrossRef]
- Hayman, D.T.S.; McCrea, R.; Restif, O.; Suu-Ire, R.; Fooks, A.R.; Wood, J.L.N.; Cunningham, A.A.; Rowcliffe, M. Demography of straw-colored fruit bats in Ghana. J. Mammal. 2012, 93, 1393–1404. [Google Scholar] [CrossRef] [Green Version]
- Amman, B.R.; Carroll, S.A.; Reed, Z.D.; Sealy, T.K.; Balinandi, S.; Swanepoel, R.; Kemp, A.; Erickson, B.R.; Comer, J.A.; Campbell, S.; et al. Seasonal Pulses of Marburg Virus Circulation in Juvenile Rousettus aegyptiacus Bats Coincide with Periods of Increased Risk of Human Infection. PLoS Pathog. 2012, 8, e1002877. [Google Scholar] [CrossRef]
- Montecino-Latorre, D.; Goldstein, T.; Gilardi, K.; Wolking, D.; Van Wormer, E.; Kazwala, R.; Ssebide, B.; Nziza, J.; Sijali, Z.; Cranfield, M.; et al. Reproduction of East-African bats may guide risk mitigation for coronavirus spillover. One Health Outlook 2020, 2, 2. [Google Scholar] [CrossRef] [Green Version]
- Dietrich, M.; Wilkinson, D.A.; Benlali, A.; Lagadec, E.; Ramasindrazana, B.; Dellagi, K.; Tortosa, P. Leptospiraand paramyxovirus infection dynamics in a bat maternity enlightens pathogen maintenance in wildlife. Environ. Microbiol. 2015, 17, 4280–4289. [Google Scholar] [CrossRef] [PubMed]
- Lacroix, A.; Kingebeni, P.M.; Kumugo, S.N.; Lempu, G.; Butel, C.; Serrano, L.; Vidal, N.; Thaurignac, G.; Esteban, A.; Bamuleka, D.M.; et al. Investigating the Circulation of Ebola Viruses in Bats during the Ebola Virus Disease Outbreaks in the Equateur and North Kivu Provinces of the Democratic Republic of Congo from 2018. Pathogens 2021, 10, 557. [Google Scholar] [CrossRef] [PubMed]
- Irwin, D.; Kocher, T.; Wilson, A.C. Evolution of the cytochromeb gene of mammals. J. Mol. Evol. 1991, 32, 128–144. [Google Scholar] [CrossRef] [PubMed]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Ayouba, A.; Touré, A.; Butel, C.; Keita, A.K.; Binetruy, F.; Sow, M.S.; Foulongne, V.; Delaporte, E.; Peeters, M. Development of a Sensitive and Specific Serological Assay Based on Luminex Technology for Detection of Antibodies to Zaire Ebola Virus. J. Clin. Microbiol. 2017, 55, 165–176. [Google Scholar] [CrossRef] [Green Version]
- Mbala-Kingebeni, P.; Arenas, C.J.V.; Vidal, N.; Likofata, J.; Nsio-Mbeta, J.; Makiala-Mandanda, S.; Mukadi, D.; Mukadi, P.; Kumakamba, C.; Djokolo, B.; et al. Rapid Confirmation of the Zaire Ebola Virus in the Outbreak of the Equateur Province in the Democratic Republic of Congo: Implications for Public Health Interventions. Clin. Infect. Dis. 2019, 68, 330–333. [Google Scholar] [CrossRef]
- Becker, D.J.; Crowley, D.E.; Washburne, A.D.; Plowright, R.K. Temporal and spatial limitations in global surveillance for bat filoviruses and henipaviruses. Biol. Lett. 2019, 15, 20190423. [Google Scholar] [CrossRef]
- Hayman, D.T. Bats as Viral Reservoirs. Annu. Rev. Virol. 2016, 3, 77–99. [Google Scholar] [CrossRef]
- Breed, A.C.; Breed, M.F.; Meers, J.; Field, H.E. Evidence of Endemic Hendra Virus Infection in Flying-Foxes (Pteropus conspicillatus)—Implications for Disease Risk Management. PLoS ONE 2011, 6, e28816. [Google Scholar] [CrossRef] [Green Version]
- George, D.B.; Webb, C.T.; Farnsworth, M.L.; O’Shea, T.J.; Bowen, R.A.; Smith, D.L.; Stanley, T.R.; Ellison, L.E.; Rupprecht, C.E. Host and viral ecology determine bat rabies seasonality and maintenance. Proc. Natl. Acad. Sci. USA 2011, 108, 10208–10213. [Google Scholar] [CrossRef] [Green Version]
- Gloza-Rausch, F.; Ipsen, A.; Seebens, A.; Göttsche, M.; Panning, M.; Drexler, J.F.; Petersen, N.; Annan, A.; Grywna, K.; Müller, M.; et al. Detection and Prevalence Patterns of Group I Coronaviruses in Bats, Northern Germany. Emerg. Infect. Dis. 2008, 14, 626–631. [Google Scholar] [CrossRef] [PubMed]
- Plowright, R.K.; Field, H.E.; Smith, C.; Divljan, A.; Palmer, C.; Tabor, G.; Daszak, P.; Foley, J.E. Reproduction and nutritional stress are risk factors for Hendra virus infection in little red flying foxes (Pteropus scapulatus). Proc. R. Soc. B Boil. Sci. 2008, 275, 861–869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brook, C.E.; Ranaivoson, H.C.; Broder, C.C.; Cunningham, A.A.; Héraud, J.; Peel, A.J.; Gibson, L.; Wood, J.L.N.; Metcalf, C.J.; Dobson, A.P. Disentangling serology to elucidate henipa- and filovirus transmission in Madagascar fruit bats. J. Anim. Ecol. 2019, 88, 1001–1016. [Google Scholar] [CrossRef] [PubMed]
- Hayman, D.T.S. Biannual birth pulses allow filoviruses to persist in bat populations. Proc. R. Soc. B Boil. Sci. 2015, 282, 20142591. [Google Scholar] [CrossRef]
- Pawęska, J.T.; Storm, N.; Markotter, W.; Di Paola, N.; Wiley, M.R.; Palacios, G.; Van Vuren, P.J. Shedding of Marburg Virus in Naturally Infected Egyptian Rousette Bats, South Africa, 2017. Emerg. Infect. Dis. 2020, 26, 3051–3055. [Google Scholar] [CrossRef]
- Diallo, M.S.K.; Ayouba, A.; Keita, A.K.; Thaurignac, G.; Sow, M.S.; Kpamou, C.; Barry, T.A.; Msellati, P.; Etard, J.-F.; Peeters, M.; et al. Temporal evolution of the humoral antibody response after Ebola virus disease in Guinea: A 60-month observational prospective cohort study. Lancet Microbe 2021, 2, e676–e684. [Google Scholar] [CrossRef]
- Schuh, A.J.; Amman, B.R.; Sealy, T.S.; Flietstra, T.D.; Guito, J.C.; Nichol, S.T.; Towner, J.S. Comparative analysis of serologic cross-reactivity using convalescent sera from filovirus-experimentally infected fruit bats. Sci. Rep. 2019, 9, 6707. [Google Scholar] [CrossRef] [Green Version]
- Hayman, D.T.S.; Emmerich, P.; Yu, M.; Wang, L.-F.; Suu-Ire, R.; Fooks, A.R.; Cunningham, A.A.; Wood, J.L.N. Long-Term Survival of an Urban Fruit Bat Seropositive for Ebola and Lagos Bat Viruses. PLoS ONE 2010, 5, e11978. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antigen | Stringent Cut-Off | Less Stringent Cut-Off | |||
|---|---|---|---|---|---|
| N Tested = 817 | N Tested = 817 | ||||
| n pos | (%) | n pos | (%) | ||
| EBOV | NP | 8 | (0.97) | 35 | (4.3) |
| EBOV | GP-K | 66 | (8.0) | 282 | (34.4) |
| EBOV | GP-M | 41 | (5.0) | 138 | (16.8) |
| EBOV | VP | 3 | (0.4) | 22 | (2.7) |
| EBOV | GP + NP | 2 | (0.3) | 13 | (1.6) |
| EBOV | GP + VP | 0 | (0.0) | 8 | (1.0) |
| EBOV | NP + GP + VP | 0 | (0.0) | 4 | (0.5) |
| SUDV | NP | 7 | (0.9) | 20 | (2.4) |
| SUDV | GP | 97 | (11.3) | 401 | (48.9) |
| SUDV | VP | 24 | (2.9) | 60 | (7.3) |
| SUDV | GP + NP | 1 | (0.1) | 6 | (0.7) |
| SUDV | GP + VP | 8 | (1.0) | 46 | (5.6) |
| SUDV | NP + GP + VP | 1 | (0.1) | 2 | (0.3) |
| BDBV | GP | 32 | (3.9) | 254 | (30.9) |
| BDBV | VP | 0 | (0.0) | 2 | (0.3) |
| BDBV | GP + VP | 0 | (0.0) | 1 | (0.1) |
| Antigen | Juvenile Females | Juvenile Males | Immature Females | Immature Males | Adult Females | Adult Males | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| N = 108 | N = 112 | N = 103 | N = 122 | N = 191 | N = 181 | |||||||
| n pos | (%) | n pos | (%) | n pos | (%) | n pos | (%) | n pos | (%) | n pos | (%) | |
| EBOV-NP | 1–5 | (2.0–4.6) | 0–3 | (0.0–2.4) | 2–4 | (1.9–3.9) | 1–8 | (0.8–6.5) | 2–5 | (1.0–2.6) | 2–10 | (1.1–5.5) |
| SUDV-NP | 2–3 | (1.8–2.8) | 2–4 | (1.8–3.6) | 0–0 | (0.0–0.0) | 1–3 | (0.8–2.5) | 0–5 | (0.0–2.6) | 2–5 | (1.1–2.7) |
| EBOV-GP-K | 1–5 | (2.0–4.6) | 0–2 | (0.0–1.8) | 22–56 | (21.2–54.3) | 24–60 | (19.7–49.2) | 7–78 | (3.6–40.8) | 12–81 | (6.6–44.8) |
| EBOV-GP-M | 0–1 | (0.0–0.9) | 1–1 | (0.9–0.9) | 17–38 | (16.3–36.9) | 12–37 | (9.8–30.3) | 6–27 | (3.1–14.1) | 5–34 | (2.8–18.8) |
| SUDV-GP | 3–9 | (2.8–8.3) | 0–9 | (0.0–8.0) | 24–74 | (23.1–71.8) | 35–84 | (28.7–68.9) | 15–103 | (10.9–53.9) | 17–122 | (9.4–67.4) |
| BDBV-GP | 0–4 | (0.0–3.7) | 0–3 | (0.0–2.4) | 14–50 | (13.5–48.5) | 13–54 | (10.6–44.3) | 5–71 | (2.6–35.7) | 5–72 | (2.8–39.8) |
| EBOV-VP | 0–3 | (0.0–2.8) | 0–0 | (0.0–0.0) | 2–6 | (1.9–5.8) | 0–5 | (0.0–4.1) | 1–1 | (0.5–0.5) | 0–7 | (0.0–3.8) |
| SUDV-VP | 2–4 | (1.8–3.7) | 1–3 | (0.9–2.4) | 10–16 | (9.6–15.5) | 8–19 | (6.6–15.6) | 1–8 | (0.5–4.2) | 2–10 | (1.1–5.5) |
| BDBV-VP | 0–0 | (0.0–0.0) | 0–0 | (0.0–0.0) | 0–1 | (0.0–0.97) | 0–0 | (0.0–0.0) | 0–0 | (0.0–0.0) | 0–1 | (0.0–0.5) |
| NP | NP | GP | GP | GP | GP | VP | VP | VP | ||
|---|---|---|---|---|---|---|---|---|---|---|
| EBOV | SUDV | EBOV- K | EBOV-M | SUDV | BDBV | EBOV | SUDV | BDBV | ||
| Juveniles vs. Immature Adults | ||||||||||
| Less stringent CO | 0.6825 | 0.4457 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | 0.6247 | 0.1588 | 1.0000 | |
| Stringent CO | 0.6070 | 0.4751 | 0.0076 | 0.2539 | 0.0015 | 0.9882 | 1.0000 | 0.8124 | 1.0000 | |
| Juveniles vs. Mature Adults | ||||||||||
| Less stringent CO | 0.9580 | 0.9641 | <0.0001 | 0.0022 | <0.0001 | <0.0001 | 0.8820 | 0.9966 | 1.0000 | |
| Stringent CO | 0.7190 | 0.5508 | 0.1154 | 0.6942 | 0.1217 | 0.9897 | 1.0000 | 0.4942 | 1.0000 | |
| Immatures vs. Mature Adults | ||||||||||
| Less stringent CO | 0.7837 | 0.4929 | 0.0487 | 0.0007 | 0.0066 | 0.2218 | 0.7007 | 0.0069 | 0.9344 | |
| Stringent CO | 0.9496 | 0.9224 | 0.0021 | 0.0382 | 0.0004 | 0.0280 | 0.5391 | 0.0103 | 1.0000 | |
| Females vs. Males | ||||||||||
| Less stringent CO | 0.2811 | 0.3938 | 0.2028 | 0.5899 | 0.4111 | 0.2235 | 0.9547 | 0.8093 | 0.9672 | |
| Stringent CO | 0.4464 | 0.2838 | 0.8070 | 0.1041 | 0.9348 | 0.1492 | 0.9987 | 0.3327 | 1.0000 | |
| No lactation vs. Lactation | ||||||||||
| Less stringent CO | 0.8367 | 0.8367 | <0.0001 | 0.0045 | <0.0001 | 0.0001 | 0.9963 | 0.9106 | 1.0000 | |
| Stringent CO | 0.6743 | 1.0000 | 0.2571 | 0.9935 | 0.0394 | 0.9936 | 0.9963 | 0.9963 | 1.0000 | |
| Antigen | Gestation | No-Gestation | Lactation | No-Lactation | ||||
|---|---|---|---|---|---|---|---|---|
| N = 13 | N = 178 | N = 68 | N = 123 | |||||
| n pos | (%) | n pos | (%) | n pos | (%) | n pos | (%) | |
| EBOV-NP | 0–0 | (0.0–0.0) | 2–5 | (1.1–2.8) | 1–2 | (1.5–2.9) | 1–3 | (0.8–2.4) |
| SUDV-NP | 0–0 | (0.0–0.0) | 0–5 | (0.0–2.8) | 0–2 | (0.02.9) | 0–3 | (0.0–2.4) |
| EBOV-GP-K | 1–6 | (7.7–46.2) | 6–72 | (3.4–40.4) | 1–13 | (1.5–19.1) | 6–65 | (4.9–52.8) |
| EBOV-GP-M | 1–3 | (7.7–23.1) | 5–24 | (2.8–13.5) | 0–2 | (0.0–2.9) | 6–25 | (4.9–20.3) |
| SUDV-GP | 1–6 | (7.7–46.2) | 14–97 | (7.9–54.5) | 1–21 | (1.5–30.9) | 14–82 | (11.4–66.7) |
| BDBV-GP | 1–5 | (7.7–38.5) | 4–66 | (2.2–37.1) | 0–12 | (0.0–22.1) | 5–59 | (4.1–47.9) |
| EBOV-VP | 0–0 | (0.0–0.0) | 1–1 | (0.6–0.6) | 0–0 | (0.0–0.0) | 1–1 | (0.8–0.8) |
| SUDV-VP | 0–0 | (0.0–0.0) | 1–8 | (0.6–4.5) | 0–3 | (0.0–4.4) | 1–5 | (0.8–4.1) |
| BDBV-VP | 0–0 | (0.0–0.0) | 0–0 | (0.0–0.0) | 0–0 | (0.0–0.0) | 0–0 | (0.0–0.0) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Djomsi, D.M.; Mba Djonzo, F.A.; Ndong Bass, I.; Champagne, M.; Lacroix, A.; Thaurignac, G.; Esteban, A.; De Nys, H.; Bourgarel, M.; Akoachere, J.-F.; et al. Dynamics of Antibodies to Ebolaviruses in an Eidolon helvum Bat Colony in Cameroon. Viruses 2022, 14, 560. https://doi.org/10.3390/v14030560
Djomsi DM, Mba Djonzo FA, Ndong Bass I, Champagne M, Lacroix A, Thaurignac G, Esteban A, De Nys H, Bourgarel M, Akoachere J-F, et al. Dynamics of Antibodies to Ebolaviruses in an Eidolon helvum Bat Colony in Cameroon. Viruses. 2022; 14(3):560. https://doi.org/10.3390/v14030560
Chicago/Turabian StyleDjomsi, Dowbiss Meta, Flaubert Auguste Mba Djonzo, Innocent Ndong Bass, Maëliss Champagne, Audrey Lacroix, Guillaume Thaurignac, Amandine Esteban, Helene De Nys, Mathieu Bourgarel, Jane-Francis Akoachere, and et al. 2022. "Dynamics of Antibodies to Ebolaviruses in an Eidolon helvum Bat Colony in Cameroon" Viruses 14, no. 3: 560. https://doi.org/10.3390/v14030560