
Transient Increases in Inflammation and Proapoptotic Potential Are Associated with the HESN Phenotype Observed in a Subgroup of Kenyan Female Sex Workers


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics
2.2. Study Subjects
2.3. Cell Culture
2.4. Synthesis and Purification of CD4-Binding Aptamer-siRNA Chimeras (CD4-AsiCs)
2.5. Quantitative RT–PCR
2.6. Data Analysis
3. Results
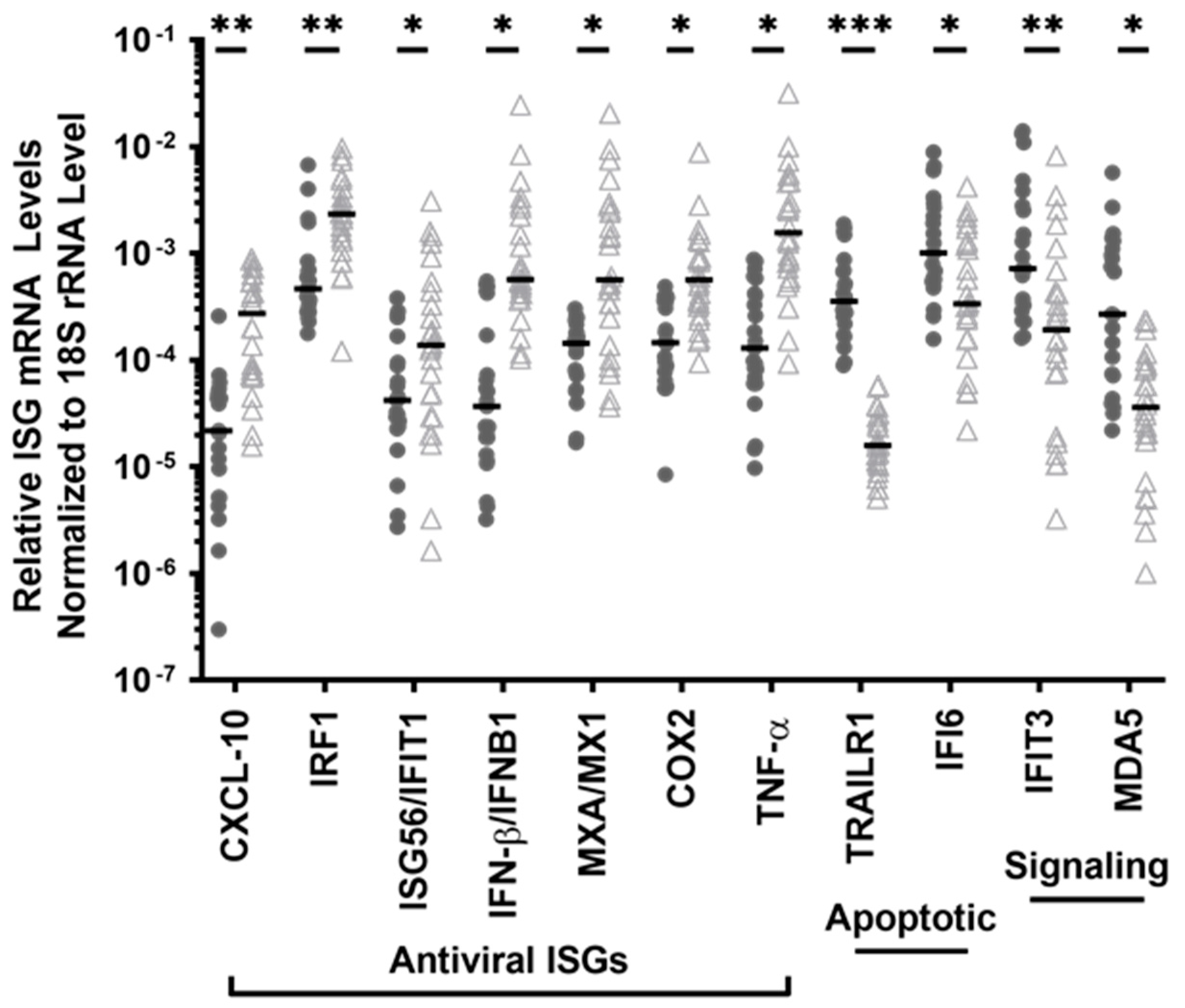
3.1. The PBMC of HESN FSWs Have Low Levels of ISG Transcripts Encoding Proinflammatory Signalling Pathway Proteins but Increased Transcripts of 2 ISGs, Key to the Detection of RNA and DNA Viruses
3.2. The PBMC of HESN FSWs Had Reduced Transcript Levels of Antiapoptotic ISGs and Elevated Proapoptotic TRAILR1 Transcripts
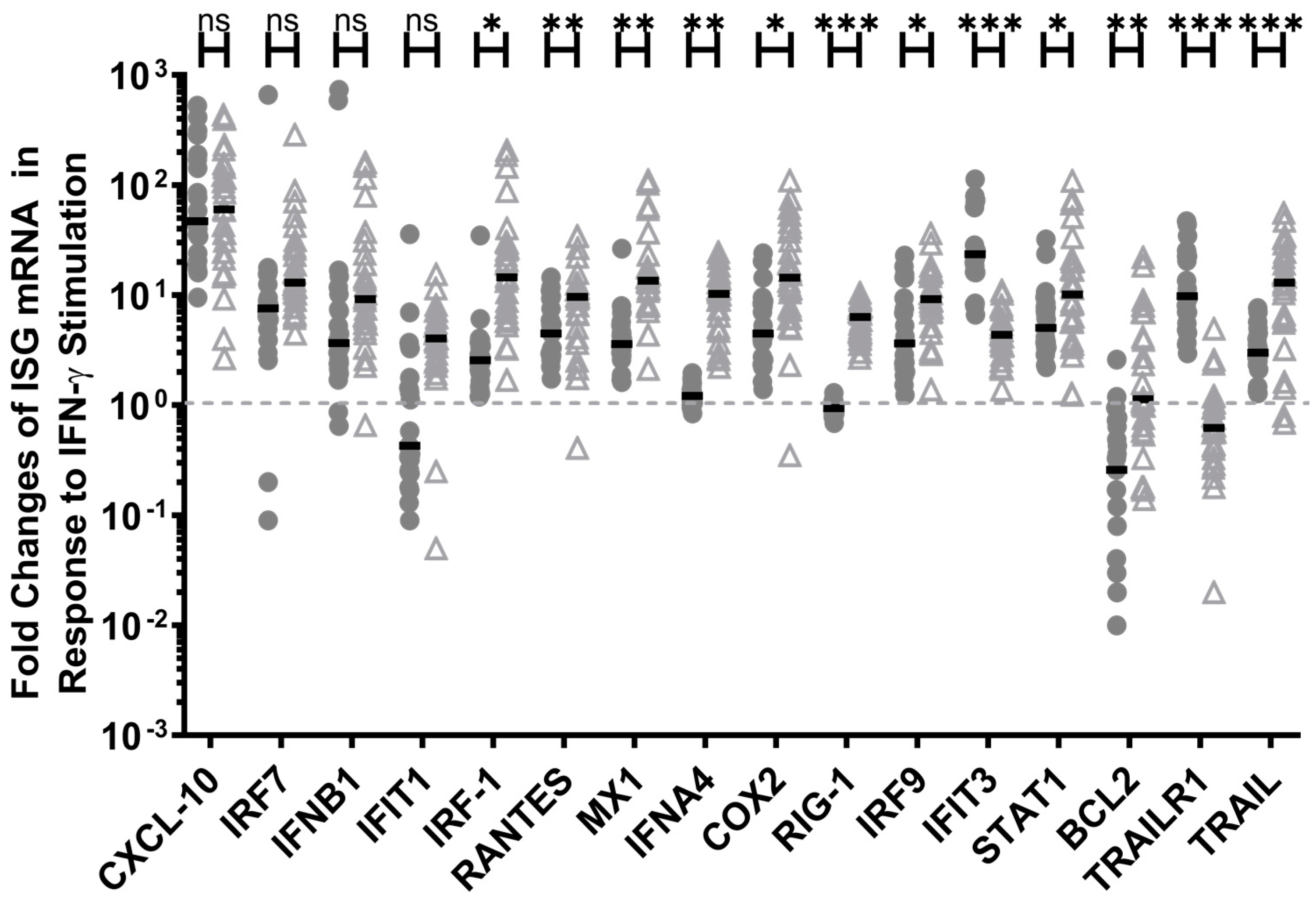
3.3. IFN-γ Induced the Expression of Pro-Inflammatory ISGs in HESN PBMC, Although to a Lesser Extent than in the Non-HESN PBMC, but Greatly Enhanced the Expression of Pro-Apoptosis ISGs
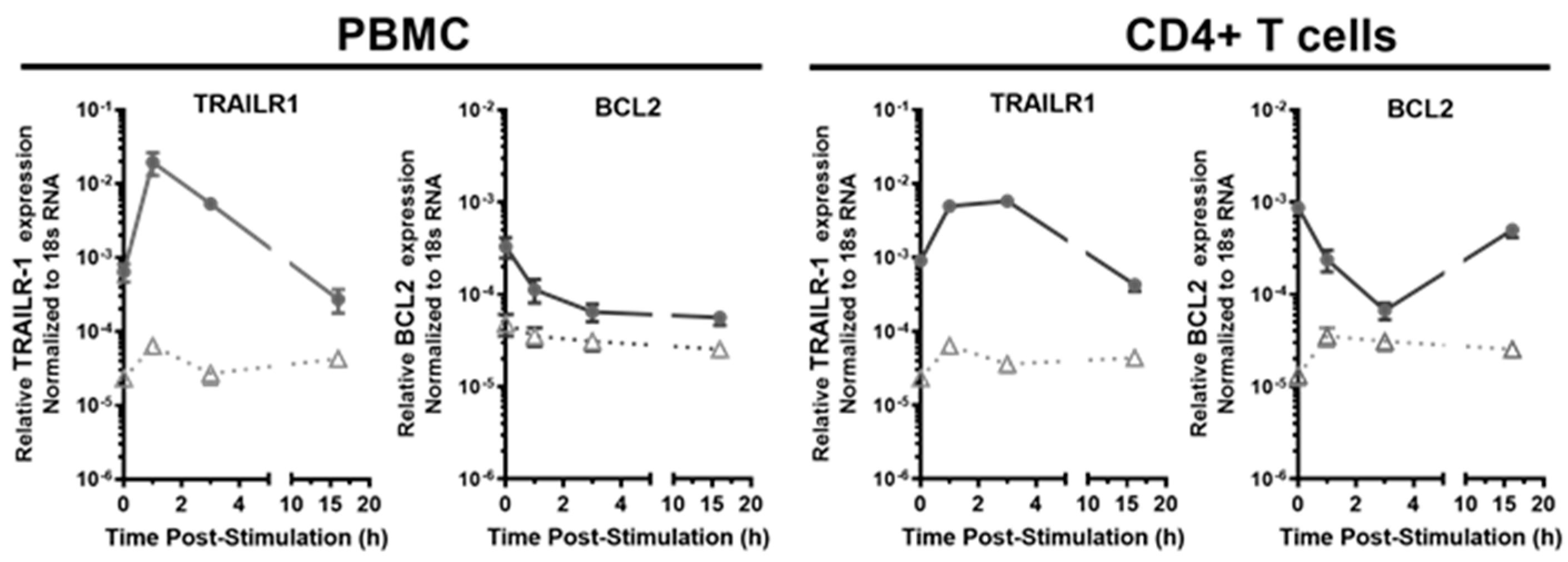
3.4. The Transcriptional Response of Antiviral Effector ISGs Was Transient in the HESN PBMC and CD4+ T Cells, Compared to the Prolonged Response in the Non-HESN Controls
3.5. IFN-γ Transiently Upregulated Proapoptotic Pathway ISG (TRAILR1 and TRAIL) and Downregulated Antiapoptotic ISG, BCL2 in HESN CD4+ T Cells
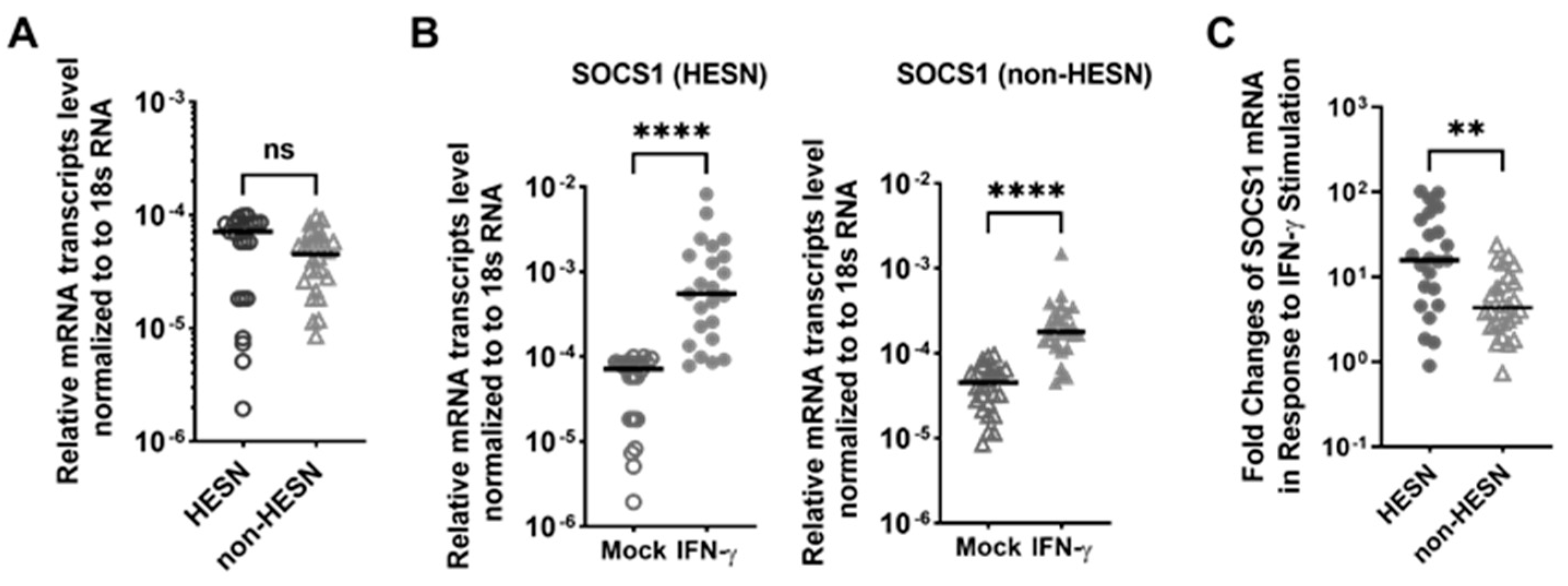
3.6. IFN-γ Induced a Stronger Negative Feedback Regulatory Response in the HESN PBMC, When Compared to Non-HESN Controls
3.7. The Transcriptional Response of Antiviral Effector ISGs and Antiapoptotic ISGs to Exogenous IFN-α2 Stimulation Was Similarly Transient in HESN PBMC and CD4+ T Cells
3.8. Not All Antiviral ISGs in the HESN Cells Exhibited Transient Response to IFN-α2 Treatment
3.9. Reducing IRF-1 Expression in Non-HESN CD4+ Cells Altered the Non-HESN ISG Profile to the ISG Profile, Similar to What Was Observed in the HESN Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- UNAIDS Data; HIV/AIDS (Ed.) UNAIDS: Geneva, Switzerland, 2018; Volume 2018, Available online: https://www.unaids.org/sites/default/files/media_asset/unaids-data-2018_en.pdf (accessed on 30 December 2021).
- UNAIDS Data; HIV/AIDS (Ed.) UNAIDS: Geneva, Switzerland, 2019; Volume 2019, Available online: https://www.unaids.org/sites/default/files/media_asset/2019-UNAIDS-data_en.pdf (accessed on 30 December 2021).
- UNAIDS. GLOBAL HIV STATISTICS; UNAIDS: Geneva, Switzerland, 2020; Volume 2020, Available online: https://www.unaids.org/sites/default/files/media_asset/2020_aids-data-book_en.pdf (accessed on 30 December 2021).
- Izulla, P.; McKinnon, L.R.; Munyao, J.; Karanja, S.; Koima, W.; Parmeres, J.; Kamuti, S.; Kioko, R.; Nagelkerke, N.; Gakii, G.; et al. HIV Postexposure Prophylaxis in an Urban Population of Female Sex Workers in Nairobi, Kenya. JAIDS J. Acquir. Immune Defic. Syndr. 2013, 62, 220–225. [Google Scholar] [CrossRef]
- McKinnon, L.; Gakii, G.; Juno, J.A.; Izulla, P.; Munyao, J.; Ireri, N.; Kariuki, C.W.; Shaw, S.Y.; Nagelkerke, N.J.D.; Gelmon, L.; et al. High HIV risk in a cohort of male sex workers from Nairobi, Kenya. Sex. Transm. Infect. 2013, 90, 237–242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chou, R.; Evans, C.; Hoverman, A.; Sun, C.; Dana, T.; Bougatsos, C.; Grusing, S.; Korthuis, P.T. Preexposure Prophylaxis for the Prevention of HIV Infection: Evidence Report and Systematic Review for the US Preventive Services Task Force. JAMA 2019, 321, 2214–2230. [Google Scholar] [CrossRef] [Green Version]
- Bavinton, B.R.; Rodger, A.J. Undetectable viral load and HIV transmission dynamics on an individual and population level: Where next in the global HIV response? Curr. Opin. Infect. Dis. 2020, 33, 20–27. [Google Scholar] [CrossRef]
- Bor, J.; Musakwa, N.; Onoya, D.; Evans, D. Perceived efficacy of HIV treatment-as-prevention among university students in Johannesburg, South Africa. Sex. Transm. Infect. 2021, 97, 596–600. [Google Scholar] [CrossRef]
- Leis, M.; McDermott, M.; Koziarz, A.; Szadkowski, L.; Kariri, A.; Beattie, T.S.; Kaul, R.; Kimani, J. Intimate partner and client-perpetrated violence are associated with reduced HIV pre-exposure prophylaxis (PrEP) uptake, depression and generalized anxiety in a cross-sectional study of female sex workers from Nairobi, Kenya. J. Int. AIDS Soc. 2021, 24, e25711. [Google Scholar] [CrossRef] [PubMed]
- Shearer, G.; Clerici, M. Historical Perspective on HIV?Exposed Seronegative Individuals: Has Nature Done the Experiment for Us? J. Infect. Dis. 2010, 202 (Suppl. S3), S329–S332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, B.; Jin, L.; Hu, F.; Xiao, J.; Luo, J.; Lu, D.; Zhang, W.; Chu, J.; Du, R.; Geng, Z.; et al. Distribution of Two HIV-1–Resistant Polymorphisms (SDF1-3′A and CCR2-64I) in East Asian and World Populations and Its Implication in AIDS Epidemiology. Am. J. Hum. Genet. 1999, 65, 1047–1053. [Google Scholar] [CrossRef] [Green Version]
- Fowke, K.R.; Kaul, R.; Rosenthal, K.L.; Oyugi, J.; Kimani, J.; Rutherford, W.J.; Nagelkerke, N.J.; Ball, T.B.; Bwayo, J.J.; Simonsen, J.N.; et al. HIV-1-specific cellular immune responses among HIV-1-resistant sex workers. Immunol. Cell Biol. 2000, 78, 586–595. [Google Scholar] [CrossRef] [Green Version]
- Iqbal, S.M.; Ball, T.B.; Kimani, J.; Kiama, P.; Thottingal, P.; Embree, J.E.; Fowke, K.; Plummer, F.A. Elevated T Cell Counts and RANTES Expression in the Genital Mucosa of HIV-1–Resistant Kenyan Commercial Sex Workers. J. Infect. Dis. 2005, 192, 728–738. [Google Scholar] [CrossRef] [Green Version]
- Ball, T.B.; Ji, H.; Kimani, J.; McLaren, P.; Marlin, C.; Hill, A.V.; Plummer, F.A. Polymorphisms in IRF-1 associated with resistance to HIV-1 infection in highly exposed uninfected Kenyan sex workers. AIDS 2007, 21, 1091–1101. [Google Scholar] [CrossRef] [PubMed]
- Burgener, A.; Boutilier, J.; Wachihi, C.; Kimani, J.; Carpenter, M.; Westmacott, G.; Cheng, K.; Ball, T.B.; Plummer, F. Identification of Differentially Expressed Proteins in the Cervical Mucosa of HIV-1-Resistant Sex Workers. J. Proteome Res. 2008, 7, 4446–4454. [Google Scholar] [CrossRef] [PubMed]
- Horton, R.E.; McLaren, P.J.; Fowke, K.; Kimani, J.; Ball, T.B. Cohorts for the Study of HIV-1–Exposed but Uninfected Individuals: Benefits and Limitations. J. Infect. Dis. 2010, 202 (Suppl. S3), S377–S381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guerini, F.R.; Lo Caputo, S.; Gori, A.; Bandera, A.; Mazzotta, F.; Uglietti, A.; Zanzottera, M.; Maserati, R.; Clerici, M. Under representation of the inhibitory KIR3DL1 molecule and the KIR3DL1+/BW4+ complex in HIV exposed seronegative individuals. J. Infect. Dis. 2011, 203, 1235–1239. [Google Scholar] [CrossRef] [Green Version]
- Malhotra, R.; Hu, L.; Song, W.; Brill, I.; Mulenga, J.; Allen, S.; Hunter, E.; Shrestha, S.; Tang, J.; Kaslow, R.A. Association of chemokine receptor gene (CCR2-CCR5) haplotypes with acquisition and control of HIV-1 infection in Zambians. Retrovirology 2011, 8, 22. [Google Scholar] [CrossRef] [Green Version]
- Tomescu, C.; Abdulhaqq, S.; Montaner, L.J. Evidence for the innate immune response as a correlate of protection in human immunodeficiency virus (HIV)-1 highly exposed seronegative subjects (HESN). Clin. Exp. Immunol. 2011, 164, 158–169. [Google Scholar] [CrossRef]
- Wilen, C.B.; Wang, J.; Tilton, J.C.; Miller, J.C.; Kim, K.A.; Rebar, E.J.; Sherrill-Mix, S.A.; Patro, S.C.; Secreto, A.J.; Jordan, A.P.O.; et al. Engineering HIV-Resistant Human CD4+ T Cells with CXCR4-Specific Zinc-Finger Nucleases. PLoS Pathog. 2011, 7, e1002020. [Google Scholar] [CrossRef]
- Ghadially, H.; Keynan, Y.; Kimani, J.; Kimani, M.; Ball, T.B.; Plummer, F.A.; Mandelboim, O.; Meyers, A.F. Altered dendritic cell–natural killer interaction in Kenyan sex workers resistant to HIV-1 infection. AIDS 2012, 26, 429–436. [Google Scholar] [CrossRef]
- Lajoie, J.; Juno, J.; Burgener, A.; Rahman, S.; Mogk, K.; Wachihi, C.; Mwanjewe, J.; Plummer, F.A.; Kimani, J.; Ball, T.B.; et al. A distinct cytokine and chemokine profile at the genital mucosa is associated with HIV-1 protection among HIV-exposed seronegative commercial sex workers. Mucosal Immunol. 2012, 5, 277–287. [Google Scholar] [CrossRef]
- Sironi, M.; Biasin, M.; Cagliani, R.; Forni, D.; De Luca, M.; Saulle, I.; Caputo, S.L.; Mazzotta, F.; Macías, J.; Pineda, J.A.; et al. A Common Polymorphism in TLR3 Confers Natural Resistance to HIV-1 Infection. J. Immunol. 2011, 188, 818–823. [Google Scholar] [CrossRef] [Green Version]
- Songok, E.M.; Luo, M.; Liang, B.; Mclaren, P.; Kaefer, N.; Apidi, W.; Boucher, G.; Kimani, J.; Wachihi, C.; Sekaly, R.; et al. Microarray Analysis of HIV Resistant Female Sex Workers Reveal a Gene Expression Signature Pattern Reminiscent of a Lowered Immune Activation State. PLoS ONE 2012, 7, e30048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jimenez, W.A.; Zapata, W.; Caruz, A.; Rugeles, M.T. High Transcript Levels of Vitamin D Receptor Are Correlated with Higher mRNA Expression of Human Beta Defensins and IL-10 in Mucosa of HIV-1-Exposed Seronegative Individuals. PLoS ONE 2013, 8, e82717. [Google Scholar]
- Card, C.M.; Ball, T.B.; Fowke, K.R. Immune Quiescence: A model of protection against HIV infection. Retrovirology 2013, 10, 141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Sorrentino, A.H.; Sinchi, J.L.; Marinic, K.; López, R.; Iliovich, E. KIR-HLA-A and B alleles of the Bw4 epitope against HIV infection in discordant heterosexual couples in Chaco Argentina. Immunology 2013, 140, 273–279. [Google Scholar] [CrossRef] [PubMed]
- Pattacini, L.; Murnane, P.M.; Kahle, E.M.; Bolton, M.J.; Delrow, J.J.; Lingappa, J.R.; Katabira, E.; Donnell, D.; McElrath, M.J.; Baeten, J.M.; et al. Differential Regulatory T Cell Activity in HIV Type 1-Exposed Seronegative Individuals. AIDS Res. Hum. Retrovir. 2013, 29, 1321–1329. [Google Scholar] [CrossRef] [Green Version]
- Zapata, W.; Jimenez, W.A.; Trujillo, N.P.; Rojas, W.; Estrada, H.; Rugeles, M.T. Influence ofCCR5andCCR2Genetic Variants in the Resistance/Susceptibility to HIV in Serodiscordant Couples from Colombia. AIDS Res. Hum. Retrovir. 2013, 29, 1594–1603. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Liu, T.-J.; Hong, Z.-H. Gene polymorphisms in CCR5, CCR2, SDF1 and RANTES among Chinese Han population with HIV-1 infection. Infect. Genet. Evol. 2014, 24, 99–104. [Google Scholar] [CrossRef]
- Lima, J.F.; Oliveira, L.M.S.; Pereira, N.Z.; Mitsunari, G.E.; Duarte, A.J.S.; Sato, M.N. Distinct Natural Killer Cells in HIV-Exposed Seronegative Subjects with Effector Cytotoxic CD56dim and CD56bright Cells and Memory-Like CD57+NKG2C+CD56dim Cells. JAIDS J. Acquir. Immune Defic. Syndr. 2014, 67, 463–471. [Google Scholar] [CrossRef]
- Shen, R.; Smith, P.D. Mucosal Correlates of Protection in HIV-1-Exposed Sero-negative Persons. Am. J. Reprod. Immunol. 2014, 72, 219–227. [Google Scholar] [CrossRef] [Green Version]
- Sironi, M.; Biasin, M.; Gnudi, F.; Cagliani, R.; Saulle, I.; Forni, D.; Rainone, V.; Trabattoni, D.; Garziano, M.; Mazzotta, F.; et al. A Regulatory Polymorphism in HAVCR2 Modulates Susceptibility to HIV-1 Infection. PLoS ONE 2014, 9, e106442. [Google Scholar] [CrossRef]
- Van Raemdonck, G.; Zegels, G.; Coen, E.; Vuylsteke, B.; Jennes, W.; Van Ostade, X. Increased Serpin A5 levels in the cervicovaginal fluid of HIV-1 exposed seronegatives suggest that a subtle balance between serine proteases and their inhibitors may determine susceptibility to HIV-1 infection. Virology 2014, 458–459, 11–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruiz-Riol, M.; Llano, A.; Ibarrondo, J.; Zamarreño, J.; Yusim, K.; Bach, V.; Mothe, B.; Perez-Alvarez, S.; Fernandez, M.A.; Requena, G.; et al. Alternative effector-function profiling identifies broad HIV-specific T-cell responses in highly HIV-exposed individuals who remain uninfected. J. Infect. Dis. 2014, 211, 936–946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fenizia, C.; Saulle, I.; Clerici, M.; Biasin, M. Genetic and epigenetic regulation of natural resistance to HIV-1 infection: New approaches to unveil the HESN secret. Expert Rev. Clin. Immunol. 2020, 16, 429–445. [Google Scholar] [CrossRef]
- Clerici, M. A role for mucosal immunity in resistance to HIV infection. Immunol. Lett. 1999, 66, 21–25. [Google Scholar] [CrossRef]
- Card, C.M.; McLaren, P.J.; Wachihi, C.; Kimani, J.; Plummer, F.A.; Fowke, K.R. Decreased Immune Activation in Resistance to HIV-1 Infection Is Associated with an Elevated Frequency of CD4+CD25+FOXP3+Regulatory T Cells. J. Infect. Dis. 2009, 199, 1318–1322. [Google Scholar] [CrossRef] [Green Version]
- Sánchez de la Torre, M.; Torres, C.; Nieto, G.; Vergara, S.; Carrero, A.J.; Macías, J.; Pineda, J.A.; Caruz, A.; Fibla, J. Vitamin D receptor gene haplotypes and susceptibility to HIV-1 infection in injection drug users. J. Infect. Dis. 2008, 197, 405–410. [Google Scholar] [CrossRef] [Green Version]
- Torres, C.; Sanchez-De-La-Torre, M.; Garcia-Moruja, C.; Carrero, A.J.; Trujillo, M.D.M.; Fibla, J.; Caruz, A. Immunophenotype of Vitamin D Receptor Polymorphism Associated to Risk of HIV-1 Infection and Rate of Disease Progression. Curr. HIV Res. 2010, 8, 487–492. [Google Scholar] [CrossRef]
- Su, R.-C.; Plesniarski, A.; Ao, Z.; Kimani, J.; Sivro, A.; Jaoko, W.; Plummer, F.A.; Yao, X.; Ball, T.B. Reducing IRF-1 to Levels Observed in HESN Subjects Limits HIV Replication, But Not the Extent of Host Immune Activation. Mol. Ther.-Nucleic Acids 2015, 4, e259. [Google Scholar] [CrossRef]
- Ji, H.; Ball, T.B.; Ao, Z.; Kimani, J.; Yao, X. Reduced HIV-1 long terminal repeat transcription in subjects with protective interferon regulatory factor-1 genotype: A potential mechanism mediating resistance to infection by HIV-1. Scand. J. Infect. Dis. 2010, 42, 389–394. [Google Scholar] [CrossRef]
- Su, R.-C.; Sivro, A.; Kimani, J.; Jaoko, W.; Plummer, F.A.; Ball, T.B. Epigenetic control of IRF1 responses in HIV-exposed seronegative versus HIV-susceptible individuals. Blood 2011, 117, 2649–2657. [Google Scholar] [CrossRef] [Green Version]
- Sivro, A.; Su, R.-C.; Plummer, F.A.; Ball, T.B. HIV and Interferon Regulatory Factor 1: A Story of Manipulation and Control. AIDS Res. Hum. Retrovir. 2013, 29, 1428–1433. [Google Scholar] [CrossRef] [PubMed]
- Schneider, W.M.; Chevillotte, M.D.; Rice, C.M. Interferon-Stimulated Genes: A Complex Web of Host Defenses. Annu. Rev. Immunol. 2014, 32, 513–545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sivro, A.; Su, R.-C.; Plummer, F.A.; Ball, T.B. Interferon responses in HIV infection: From protection to disease. Aids Rev. 2014, 16, 43–51. [Google Scholar] [PubMed]
- JSchoggins, J.W. Interferon-Stimulated Genes: What Do They All Do? Annu. Rev. Virol. 2019, 6, 567–584. [Google Scholar] [CrossRef]
- TNguyen, T.A.; Pang, K.; Masters, S.L. Intercellular communication for innate immunity. Mol. Immunol. 2017, 86, 16–22. [Google Scholar]
- Walker, F.C.; Sridhar, P.R.; Baldridge, M.T. Differential roles of interferons in innate responses to mucosal viral infections. Trends Immunol. 2021, 42, 1009–1023. [Google Scholar] [CrossRef]
- Lederer, S.; Favre, D.; Walters, K.-A.; Proll, S.; Kanwar, B.; Kasakow, Z.; Baskin, C.R.; Palermo, R.; McCune, J.M.; Katze, M.G. Transcriptional Profiling in Pathogenic and Non-Pathogenic SIV Infections Reveals Significant Distinctions in Kinetics and Tissue Compartmentalization. PLoS Pathog. 2009, 5, e1000296. [Google Scholar] [CrossRef] [Green Version]
- Sandler, N.G.; Bosinger, S.E.; Estes, J.D.; Zhu, R.T.R.; Tharp, G.K.; Boritz, E.; Levin, D.; Wijeyesinghe, S.; Makamdop, K.N.; Del Prete, G.Q.; et al. Type I interferon responses in rhesus macaques prevent SIV infection and slow disease progression. Nature 2014, 511, 601–605. [Google Scholar] [CrossRef] [Green Version]
- Dagenais-Lussier, X.; Loucif, H.; Murira, A.; Laulhé, X.; Stäger, S.; Lamarre, A.; Van Grevenynghe, J. Sustained IFN-I Expression during Established Persistent Viral Infection: A “Bad Seed” for Protective Immunity. Viruses 2017, 10, 12. [Google Scholar] [CrossRef] [Green Version]
- Marsili, G.; Remoli, A.L.; Sgarbanti, M.; Battistini, A. Role of Acetylases and Deacetylase Inhibitors in IRF-1-Mediated HIV-1 Long Terminal Repeat Transcription. Ann. N. Y. Acad. Sci. 2004, 1030, 636–643. [Google Scholar] [CrossRef]
- Remoli, A.L.; Marsili, G.; Perrotti, E.; Gallerani, E.; Ilari, R.; Nappi, F.; Cafaro, A.; Ensoli, B.; Gavioli, R.; Battistini, A. Intracellular HIV-1 Tat protein represses constitutive LMP2 transcription increasing proteasome activity by interfering with the binding of IRF-1 to STAT1. Biochem. J. 2006, 396, 371–380. [Google Scholar] [CrossRef] [PubMed]
- Sgarbanti, M.; Remoli, A.L.; Marsili, G.; Ridolfi, B.; Borsetti, A.; Perrotti, E.; Orsatti, R.; Ilari, R.; Sernicola, L.; Stellacci, E.; et al. IRF-1 is required for full NF-kappaB transcriptional activity at the human immunodeficiency virus type 1 long terminal repeat enhancer. J. Virol. 2008, 82, 3632–3641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panda, D.; Gjinaj, E.; Bachu, M.; Squire, E.; Novatt, H.; Ozato, K.; Rabin, R.L. IRF1 Maintains Optimal Constitutive Expression of Antiviral Genes and Regulates the Early Antiviral Response. Front. Immunol. 2019, 10, 1019. [Google Scholar] [CrossRef] [PubMed]
- Feng, H.; Zhang, Y.-B.; Gui, J.-F.; Lemon, S.M.; Yamane, D. Interferon regulatory factor 1 (IRF1) and anti-pathogen innate immune responses. PLoS Pathog. 2021, 17, e1009220. [Google Scholar] [CrossRef] [PubMed]
- Song, R.; Gao, Y.; Dozmorov, I.; Malladi, V.; Saha, I.; McDaniel, M.M.; Parameswaran, S.; Liang, C.; Arana, C.; Zhang, B.; et al. IRF1 governs the differential interferon-stimulated gene responses in human monocytes and macrophages by regulating chromatin accessibility. Cell Rep. 2021, 34, 108891. [Google Scholar] [CrossRef]
- Peteranderl, C.; Herold, S. The Impact of the Interferon/TNF-Related Apoptosis-Inducing Ligand Signaling Axis on Disease Progression in Respiratory Viral Infection and Beyond. Front. Immunol. 2017, 8, 313. [Google Scholar] [CrossRef] [Green Version]
- Echebli, N.; Tchitchek, N.; Dupuy, S.; Bruel, T.; Peireira Bittencourt Passaes, C.; Bosquet, N.; Le Grand, R.; Bourgeois, C.; Favier, B.; Cheynier, R.; et al. Stage-specific IFN-induced and IFN gene expression reveal convergence of type I and type II IFN and highlight their role in both acute and chronic stage of pathogenic SIV infection. PLoS ONE 2018, 13, e0190334. [Google Scholar]
- Fowke, K.; Nagelkerke, N.J.; Kimani, J.; Simonsen, J.N.; Anzala, A.O.; Bwayo, J.J.; MacDonald, K.S.; Ngugi, E.N.; Plummer, F.A. Resistance to HIV-1 infection among persistently seronegative prostitutes in Nairobi, Kenya. Lancet 1996, 348, 1347–1351. [Google Scholar] [CrossRef]
- Cohen, J. AIDS vaccine research. HIV natural resistance field finally overcomes resistance. Science 2009, 326, 1476–1477. [Google Scholar] [CrossRef]
- Tago, A.; McKinnon, L.R.; Wanjiru, T.; Muriuki, F.; Munyao, J.; Gakii, G.; Akolo, M.; Kariri, A.; Reed, N.; Shaw, S.Y.; et al. Declines in HIV prevalence in female sex workers accessing an HIV treatment and prevention programme in Nairobi, Kenya over a 10-year period. AIDS 2021, 35, 317–324. [Google Scholar] [CrossRef]
- Wheeler, L.A.; Trifonova, R.; Vrbanac, V.; Basar, E.; McKernan, S.; Xu, Z.; Seung, E.; Deruaz, M.; Dudek, T.; Einarsson, J.I.; et al. Inhibition of HIV transmission in human cervicovaginal explants and humanized mice using CD4 aptamer-siRNA chimeras. J. Clin. Investig. 2011, 121, 2401–2412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kane, M.; Zang, T.M.; Rihn, S.J.; Zhang, F.; Kueck, T.; Alim, M.; Schoggins, J.; Rice, C.M.; Wilson, S.J.; Bieniasz, P.D. Identification of Interferon-Stimulated Genes with Antiretroviral Activity. Cell Host Microbe 2016, 20, 392–405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, E.; Li, M.M.H. All About the RNA: Interferon-Stimulated Genes That Interfere with Viral RNA Processes. Front. Immunol. 2020, 11, 605024. [Google Scholar] [CrossRef] [PubMed]
- Funabiki, M.; Kato, H.; Miyachi, Y.; Toki, H.; Motegi, H.; Inoue, M.; Minowa, O.; Yoshida, A.; Deguchi, K.; Sato, H.; et al. Autoimmune Disorders Associated with Gain of Function of the Intracellular Sensor MDA5. Immunity 2014, 40, 199–212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lincez, P.J.; Shanina, I.; Horwitz, M.S. Reduced Expression of the MDA5 Gene IFIH1 Prevents Autoimmune Diabetes. Diabetes 2015, 64, 2184–2193. [Google Scholar] [CrossRef] [Green Version]
- Johnson, B.; VanBlargan, L.A.; Xu, W.; White, J.P.; Shan, C.; Shi, P.-Y.; Zhang, R.; Adhikari, J.; Gross, M.L.; Leung, D.W.; et al. Human IFIT3 Modulates IFIT1 RNA Binding Specificity and Protein Stability. Immunity 2018, 48, 487–499.e5. [Google Scholar] [CrossRef] [Green Version]
- Jacquelin, B.; Mayau, V.; Targat, B.; Liovat, A.-S.; Kunkel, D.; Petitjean, G.; Dillies, M.-A.; Roques, P.; Butor, C.; Silvestri, G.; et al. Nonpathogenic SIV infection of African green monkeys induces a strong but rapidly controlled type I IFN response. J. Clin. Investig. 2009, 119, 3544–3555. [Google Scholar] [CrossRef] [Green Version]
- Honke, N.; Shaabani, N.; Merches, K.; Gassa, A.; Kraft, A.; Ehrhardt, K.; Häussinger, D.; Löhning, M.; Dittmer, U.; Hengel, H.; et al. Immunoactivation induced by chronic viral infection inhibits viral replication and drives immunosuppression through sustained IFN-I responses. Eur. J. Immunol. 2015, 46, 372–380. [Google Scholar] [CrossRef]
- BBurke, B.; Brown, H.J.; Marsden, M.D.; Bristol, G.; Vatakis, D.N.; Zack, J.A. Primary Cell Model for Activation-Inducible Human Immunodeficiency Virus. J. Virol. 2007, 81, 7424–7434. [Google Scholar] [CrossRef] [Green Version]
- Vatakis, D.N.; Nixon, C.C.; Bristol, G.; Zack, J.A. Differentially stimulated CD4+ T cells display altered human immunodeficiency virus infection kinetics: Implications for the efficacy of antiviral agents. J. Virol. 2009, 83, 3374–3378. [Google Scholar] [CrossRef] [Green Version]
- Liau, N.P.D.; Laktyushin, A.; Lucet, I.S.; Murphy, J.M.; Yao, S.; Whitlock, E.; Callaghan, K.; Nicola, N.A.; Kershaw, N.J.; Babon, J.J. The molecular basis of JAK/STAT inhibition by SOCS1. Nat. Commun. 2018, 9, 1558. [Google Scholar] [CrossRef] [PubMed]
- Mazewski, C.; Perez, R.E.; Fish, E.N.; Platanias, L.C. Type I Interferon (IFN)-Regulated Activation of Canonical and Non-Canonical Signaling Pathways. Front. Immunol. 2020, 11, 606456. [Google Scholar] [CrossRef]
- Zhao, C.; Denison, C.; Huibregtse, J.M.; Gygi, S.; Krug, R.M. Human ISG15 conjugation targets both IFN-induced and constitutively expressed proteins functioning in diverse cellular pathways. Proc. Natl. Acad. Sci. USA 2005, 102, 10200–10205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Landré, V.; Pion, E.; Narayan, V.; Xirodimas, D.P.; Ball, K.L. DNA-binding regulates site-specific ubiquitination of IRF-1. Biochem. J. 2013, 449, 707–717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Remoli, A.L.; Marsili, G.; Perrotti, E.; Acchioni, C.; Sgarbanti, M.; Borsetti, A.; Hiscott, J.; Battistini, A. HIV-1 Tat Recruits HDM2 E3 Ligase To Target IRF-1 for Ubiquitination and Proteasomal Degradation. mBio 2016, 7, e01528-16. [Google Scholar] [CrossRef] [Green Version]
- Galvin, S.R.; Cohen, M.S. The role of sexually transmitted diseases in HIV transmission. Nat. Rev. Genet. 2004, 2, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Bebell, L.M.; Passmore, J.; Williamson, C.; Mlisana, K.; Iriogbe, I.; Van Loggerenberg, F.; Karim, Q.A.; Karim, S.A. Relationship between Levels of Inflammatory Cytokines in the Genital Tract and CD4+ Cell Counts in Women with Acute HIV-1 Infection. J. Infect. Dis. 2008, 198, 710–714. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaudhuri, A.; Yang, B.; Gendelman, H.E.; Persidsky, Y.; Kanmogne, G.D. STAT1 signaling modulates HIV-1-induced inflammatory responses and leukocyte transmigration across the blood-brain barrier. Blood 2008, 111, 2062–2072. [Google Scholar] [CrossRef] [PubMed]
- Kaul, R.; Prodger, J.; Joag, V.; Shannon, B.; Yegorov, S.; Galiwango, R.; McKinnon, L. Inflammation and HIV Transmission in Sub-Saharan Africa. Curr. HIV/AIDS Rep. 2015, 12, 216–222. [Google Scholar] [CrossRef]
- McKinnon, L.R.; Nyanga, B.; Kim, C.J.; Izulla, P.; Kwatampora, J.; Kimani, M.; Shahabi, K.; Mugo, N.; Smith, J.S.; Anzala, A.O.; et al. Early HIV-1 Infection Is Associated with Reduced Frequencies of Cervical Th17 Cells. JAIDS J. Acquir. Immune Defic. Syndr. 2015, 68, 6–12. [Google Scholar] [CrossRef] [Green Version]
- Mwatelah, R.; McKinnon, L.R.; Baxter, C.; Abdool Karim, Q.; Abdool Karim, S.S. Mechanisms of sexually transmitted infection-induced inflammation in women: Implications for HIV risk. J. Int. AIDS Soc. 2019, 22 (Suppl. S6), e25346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haase, A.T. Population Biology of HIV-1 Infection: Viral and CD4+ T Cell Demographics and Dynamics in Lymphatic Tissues. Annu. Rev. Immunol. 1999, 17, 625–656. [Google Scholar] [CrossRef] [PubMed]
- Koning, F.A.; Otto, S.A.; Hazenberg, M.D.; Dekker, L.; Prins, M.; Miedema, F.; Schuitemaker, H. Low-Level CD4+ T Cell Activation Is Associated with Low Susceptibility to HIV-1 Infection. J. Immunol. 2005, 175, 6117–6122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salgado, M.; Swanson, M.D.; Pohlmeyer, C.W.; Buckheit, R.W.; Wu, J.; Archin, N.M.; Williams, T.M.; Margolis, D.M.; Siliciano, R.F.; Garcia, J.V.; et al. HLA-B*57 Elite Suppressor and Chronic Progressor HIV-1 Isolates Replicate Vigorously and Cause CD4+ T Cell Depletion in Humanized BLT Mice. J. Virol. 2014, 88, 3340–3352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bosinger, S.E.; Li, Q.; Gordon, S.N.; Klatt, N.R.; Duan, L.; Xu, L.; Francella, N.; Sidahmed, A.; Smith, A.J.; Cramer, E.M.; et al. Global genomic analysis reveals rapid control of a robust innate response in SIV-infected sooty mangabeys. J. Clin. Investig. 2009, 119, 3556–3572. [Google Scholar] [CrossRef]
- Cheng, L.; Yu, H.; Li, G.; Li, F.; Ma, J.; Li, J.; Chi, L.; Zhang, L.; Su, L. Type I interferons suppress viral replication but contribute to T cell depletion and dysfunction during chronic HIV-1 infection. JCI Insight 2017, 2, e94366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, B.; Kang, W.; Zuo, J.; Kang, W.; Sun, Y. The Significance of Type-I Interferons in the Pathogenesis and Therapy of Human Immunodeficiency Virus 1 Infection. Front. Immunol. 2017, 8, 1431. [Google Scholar] [CrossRef] [Green Version]
- Alexander, W.S.; Starr, R.; Fenner, J.E.; Scott, C.L.; Handman, E.; Sprigg, N.S.; Corbin, J.E.; Cornish, A.L.; Darwiche, R.; Owczarek, C.M.; et al. SOCS1 is a critical inhibitor of interferon gamma signaling and prevents the potentially fatal neonatal actions of this cytokine. Cell 1999, 98, 597–608. [Google Scholar] [CrossRef] [Green Version]
- Hersberger, M.; Schlaepfer, E.; Bühler, M.; Bochud, P.-Y.; Vernazza, P.; Marti-Jaun, J.; Nemeth, J.; Zwahlen, M.; Schmidlin, K.; Speck, R.F.; et al. Polymorphisms of SOCS-1 Are Associated with a Rapid HIV Progression Rate. JAIDS J. Acquir. Immune Defic. Syndr. 2020, 84, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Sutra Del Galy, A.; Menegatti, S.; Fuentealba, J.; Lucibello, F.; Perrin, L.; Helft, J.; Darbois, A.; Saitakis, M.; Tosello, J.; Rookhuizen, D.; et al. In vivo genome-wide CRISPR screens identify SOCS1 as intrinsic checkpoint of CD4+ TH1 cell response. Sci. Immunol. 2021, 6, eabe8219. [Google Scholar] [CrossRef]
- McKinnon, L.R.; Izulla, P.; Nagelkerke, N.; Munyao, J.; Wanjiru, T.; Shaw, S.Y.; Gichuki, R.; Kariuki, C.; Muriuki, F.; Musyoki, H.; et al. Risk Factors for HIV Acquisition in a Prospective Nairobi-Based Female Sex Worker Cohort. AIDS Behav. 2015, 19, 2204–2213. [Google Scholar] [CrossRef] [PubMed]
- Fragale, A.; Gabriele, L.; Stellacci, E.; Borghi, P.; Perrotti, E.; Ilari, R.; Lanciotti, A.; Remoli, A.L.; Venditti, M.; Belardelli, F.; et al. IFN Regulatory Factor-1 Negatively Regulates CD4+CD25+ Regulatory T Cell Differentiation by Repressing Foxp3 Expression. J. Immunol. 2008, 181, 1673–1682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mostecki, J.; Showalter, B.M.; Rothman, P.B. Early Growth Response-1 Regulates Lipopolysaccharide-induced Suppressor of Cytokine Signaling-1 Transcription. J. Biol. Chem. 2005, 280, 2596–2605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kotredes, K.P.; Gamero, A.M. Interferons as Inducers of Apoptosis in Malignant Cells. J. Interferon Cytokine Res. 2013, 33, 162–170. [Google Scholar] [CrossRef] [Green Version]
- Ji, H.; Ball, T.B.; Kimani, J.; Plummer, F.A. Novel interferon regulatory factor-1 polymorphisms in a Kenyan population revealed by complete gene sequencing. J. Hum. Genet. 2004, 49, 528–535. [Google Scholar] [CrossRef] [PubMed]
- Horton, R.E.; Kaefer, N.; Songok, E.; Guijon, F.B.; Kettaf, N.; Boucher, G.; Sekaly, R.-P.; Ball, T.B.; Plummer, F.A. A Comparative Analysis of Gene Expression Patterns and Cell Phenotypes between Cervical and Peripheral Blood Mononuclear Cells. PLoS ONE 2009, 4, e8293. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | HESN (n = 23) | Non-HESN (n = 26) | p-Value a |
|---|---|---|---|
| Median or Number (Range or %) | |||
| Age (years) | 37 (26–45) | 31 (21–38) | <0.001 |
| Weight (Kg) | 77 (52–98) | 71 (42–96) | n.s. |
| Active in sex work (years) | 12 (7–24) | 2 (0–3) | <10−6 |
| No. clients last week | 9 (0–50) | 12 (0–70) | 0.50 |
| Self-reported HIV+ clients last week | 1 (0–3) | 1 (0–3) | 0.52 |
| Unprotected sex acts in the last 7 days | 2 (0–5) | 2 (0–12) | 0.55 |
| BV b status negative (n, %) | 23, 100% | 26, 100% | n.s. |
| Yeast infection negative (n, %) | 0, 0% | 0, 0% | n.s. |
| Vaginal douching (n, %) c | 23, 100% | 26, 100% | n.s. |
| 35 ISGs Were Detected in Unstimulated Ex Vivo PBMC a |
| ACTB/B-ACTIN, ATM, BAX, BCL2, BclXL/BCL2L1, CCL5/RANTES, CDK4, CDKN2A/p16/p14, CXCL10/IP-10, DPP4/CD26, E2F1, FCGR2B, EIF2AK2/PKR, HDM2, IFI6/G1P3, IFIH1/MDA5, IFIT1/ISG56, IFIT3/ISG60, IFNA2, IFNA4, IFNB1, IL1B, IRF-1, IRF-7, IRF-9, ISG15, MXA, MYD88, PMAIP1/NOXA, PTGS2/COX2, RIGI/DDX58, STAT1, TNF/TNFA, TNFSF10/TRAIL, TNFRSF10A/TRAILR1 |
| 41 ISGs Were NOT Detected in Unstimulated Ex Vivo PBMC a |
| 5LO, ANKRD1, BRCA1, C5, CAR, CASP8, CASP9, CDC25B, CDC25A, CDK1, CDK6, CDK2, CDK4A, CDK7, CIAP2, FAS, FASL, FLIP, FLAP, GADD45, GPT2, IFNa113, IRF4, IRF5, LMP7, LMP10, MCL1, MX2, OAS1, P21cip1, p53/Tp53, p8/NUPR1, PELI1, PSME2, PUMA, RB1, RELA, RSAD2/Viperin TAP1, XIAP, ZFP36L2 |
| HESN Only (0) | HESN & Non-HESN (25) | Non-HESN Only (10) |
|---|---|---|
| ACTB/B-ACTIN, BAX, CXCL10/IP-10, CCL5/RANTES, CDKN2A/p16/p14, DPP4/CD26, EIF2AK2/PKR, E2F1, HDM2, IFI6/G1P3, IFIH1/MDA5, IFIT1/ISG56, IFIT3/ISG60, IFNA4, IFNB1, IRF-1, IRF-7, IRF-9, MX1, MYD88, PTGS2/COX2, TNF/TNFA, TNFSF10/TRAIL1, TNFRSF10A/TRAILR1, STAT1 | ATM, BCL2, BclXL/BCL2L1, CDK4, FCGR2B, IFNA2, IL-1B, ISG15, PMAIP1/NOXA, RIG-1/DDX58 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gluchowski, M.; Yu, X.; Abrenica, B.; Yao, S.; Kimani, J.; Douville, R.N.; Ball, T.B.; Su, R.-C. Transient Increases in Inflammation and Proapoptotic Potential Are Associated with the HESN Phenotype Observed in a Subgroup of Kenyan Female Sex Workers. Viruses 2022, 14, 471. https://doi.org/10.3390/v14030471
Gluchowski M, Yu X, Abrenica B, Yao S, Kimani J, Douville RN, Ball TB, Su R-C. Transient Increases in Inflammation and Proapoptotic Potential Are Associated with the HESN Phenotype Observed in a Subgroup of Kenyan Female Sex Workers. Viruses. 2022; 14(3):471. https://doi.org/10.3390/v14030471
Chicago/Turabian StyleGluchowski, Marcel, Xiaoqiong Yu, Bernard Abrenica, Samantha Yao, Joshua Kimani, Renée N. Douville, Terry Blake Ball, and Ruey-Chyi Su. 2022. "Transient Increases in Inflammation and Proapoptotic Potential Are Associated with the HESN Phenotype Observed in a Subgroup of Kenyan Female Sex Workers" Viruses 14, no. 3: 471. https://doi.org/10.3390/v14030471