
HDV Seroprevalence in HBsAg-Positive Patients in China Occurs in Hotspots and Is Not Associated with HCV Mono-Infection

 ,
,  , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Population
2.2. Serological Tests
2.3. Qualitative Anti-HDV-Ab ELISA
2.4. Quantification of Virus Load
2.5. RT-PCR, PCR, and Nested PCR
2.6. Sanger Sequencing, Sequence Alignment, and Phylogenetic Analysis
2.7. Statistics
3. Results
3.1. HDV Occurs in Geographic Hotspots in China
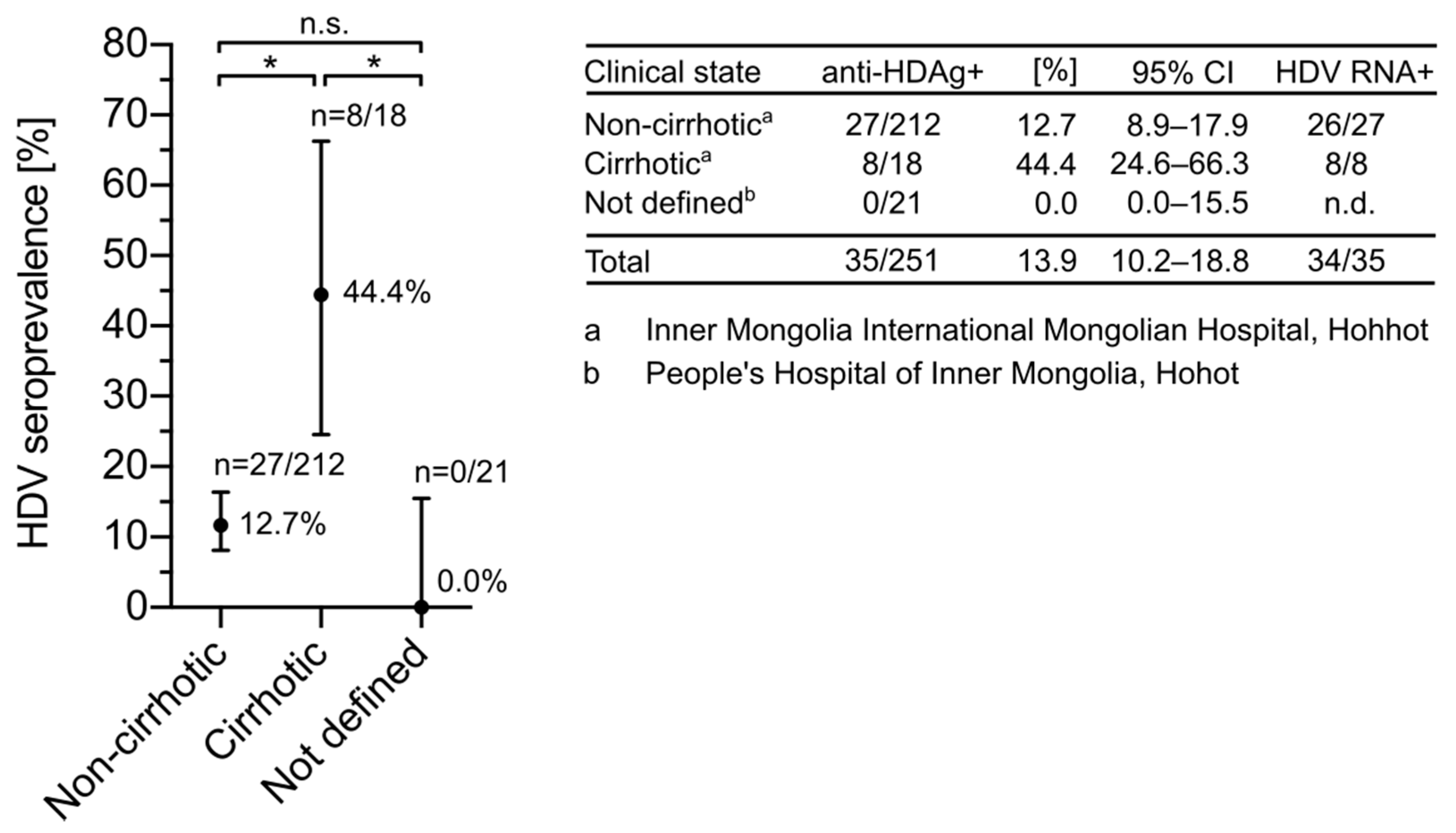
3.2. HDV Seroprevalence Is Increased in Cirrhotic Patients from Inner Mongolia
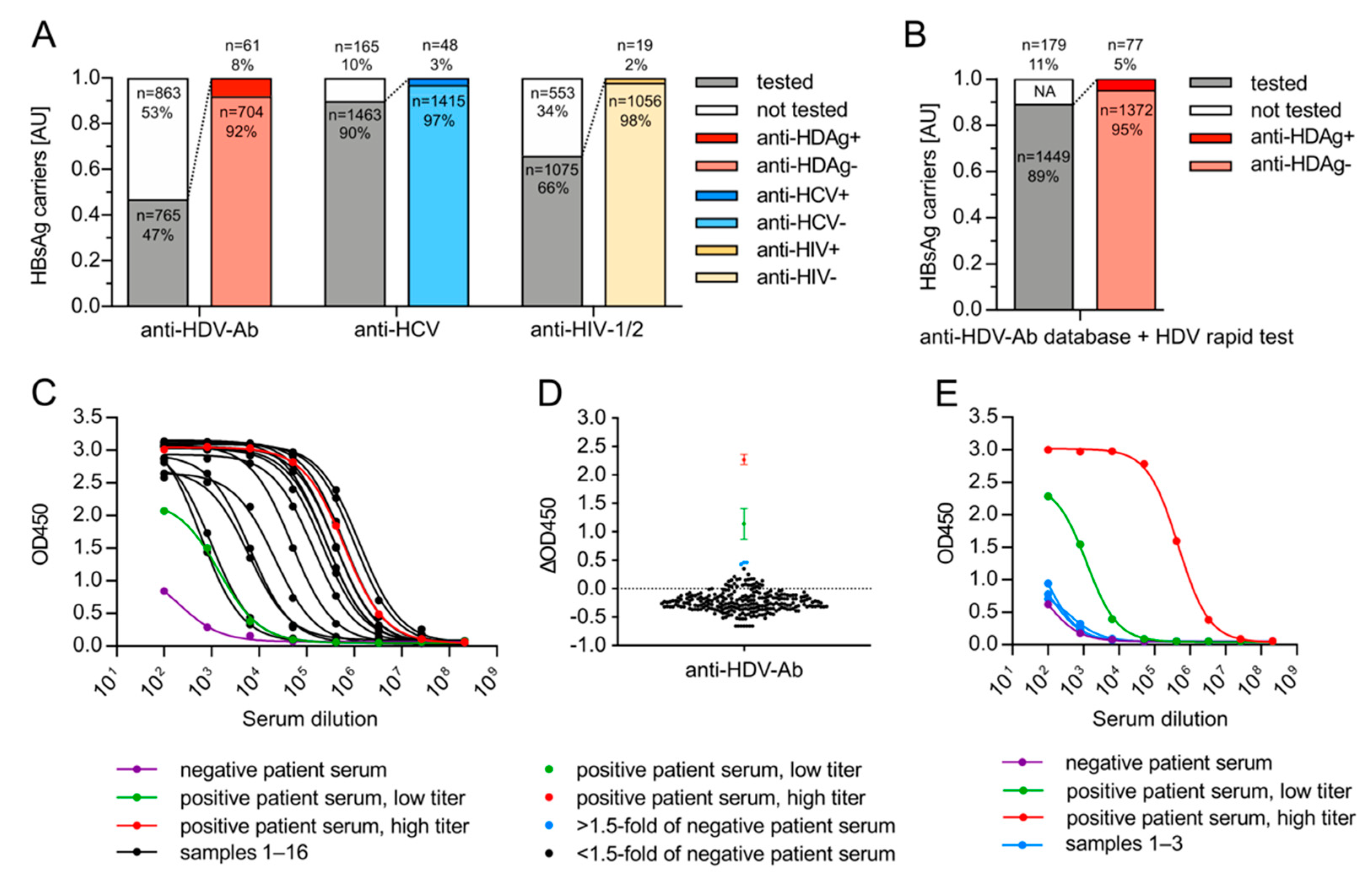
3.3. HDV Associates with HBsAg-Positive but Not with HBsAg-Negative, Anti-HBc-Negative HCV Patients from Germany
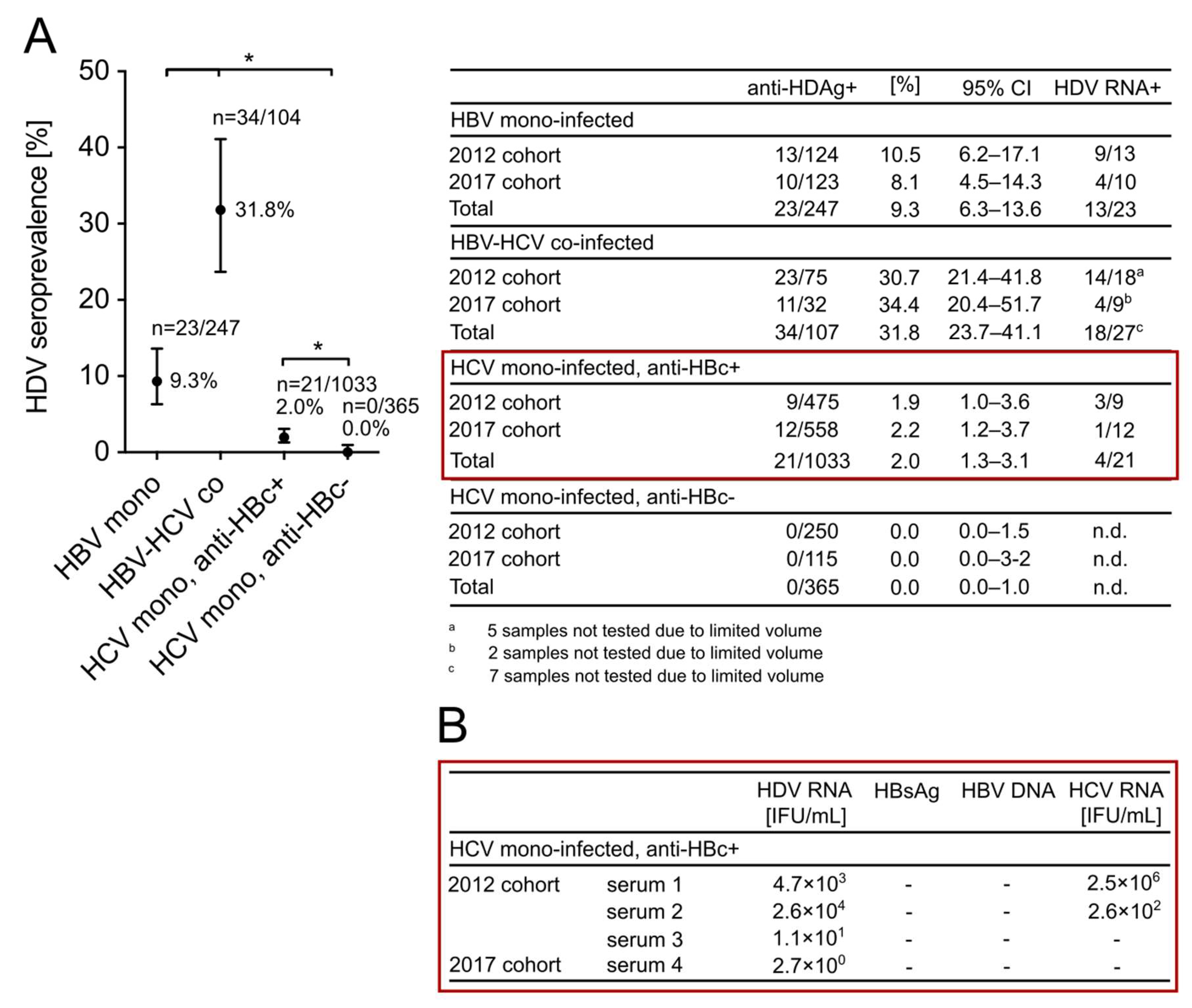
3.4. HDV Is Highly Endemic in Patients with Positive HBV Markers from a High-Risk Needle-Sharing IVDU Population in China and Is Not Associated with HCV Mono-Infection
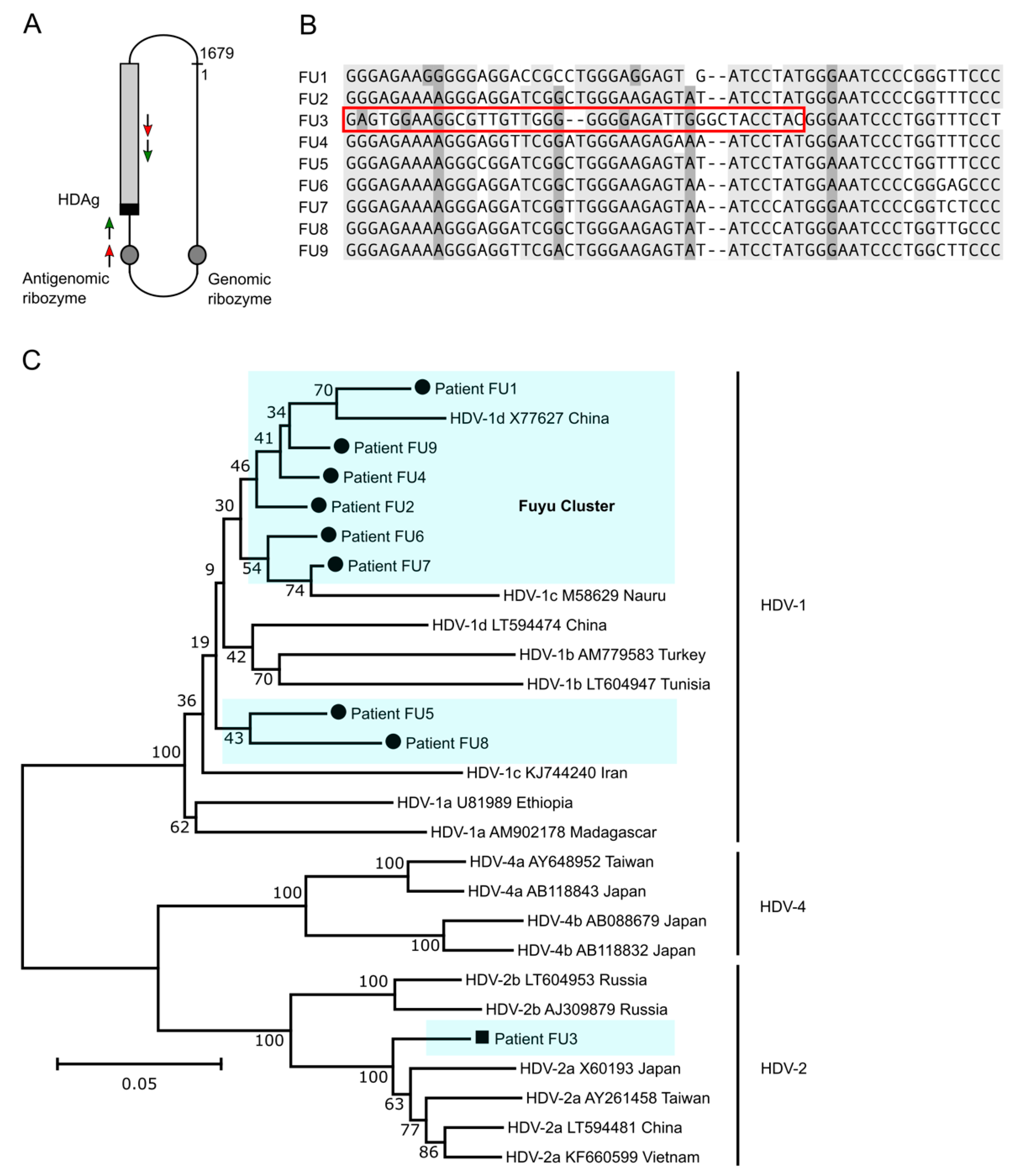
3.5. HDV Genotype 1 Is Most Abundant in Fuyu and Forms a Cluster of Strains but HDV Genotype 2a Is Also Present
4. Discussion
5. Conclusions
6. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ni, Y.; Lempp, F.A.; Mehrle, S.; Nkongolo, S.; Kaufman, C.; Fälth, M.; Stindt, J.; Königer, C.; Nassal, M.; Kubitz, R. Hepatitis B and D viruses exploit sodium taurocholate co-transporting polypeptide for species-specific entry into hepatocytes. Gastroenterology 2014, 146, 1070–1083. [Google Scholar] [CrossRef]
- Rizzetto, M.; Hoyer, B.; Canese, M.G.; Shih, J.; Purcell, R.H.; Gerin, J.L. delta Agent: Association of delta antigen with hepatitis B surface antigen and RNA in serum of delta-infected chimpanzees. Proc. Natl. Acad. Sci. USA 1980, 77, 6124–6128. [Google Scholar] [CrossRef] [Green Version]
- Rizzetto, M.; Canese, M.G.; Arico, S.; Crivelli, O.; Trepo, C.; Bonino, F.; Verme, G. Immunofluorescence detection of new antigen-antibody system (delta/anti-delta) associated to hepatitis B virus in liver and in serum of HBsAg carriers. Gut 1977, 18, 997–1003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.-Y.; Shen, D.-T.; Ji, D.-Z.; Han, P.-C.; Zhang, W.-M.; Ma, J.-F.; Chen, W.-S.; Goyal, H.; Pan, S.; Xu, H.-G. Prevalence and burden of hepatitis D virus infection in the global population: A systematic review and meta-analysis. Gut 2019, 68, 512–521. [Google Scholar] [CrossRef] [PubMed]
- Niro, G.A.; Casey, J.L.; Gravinese, E.; Garrubba, M.; Conoscitore, P.; Sagnelli, E.; Durazzo, M.; Caporaso, N.; Perri, F.; Leandro, G. Intrafamilial transmission of hepatitis delta virus: Molecular evidence. J. Hepatol. 1999, 30, 564–569. [Google Scholar] [CrossRef]
- EASL. Clinical practice guidelines on the management of hepatitis B virus infection. J. Hepatol. 2017, 67, 370–398. [Google Scholar] [CrossRef] [Green Version]
- Terrault, N.A.; Bzowej, N.H.; Chang, K.M.; Hwang, J.P.; Jonas, M.M.; Murad, M.H. AASLD guidelines for treatment of chronic hepatitis B. Hepatology 2016, 63, 261–283. [Google Scholar] [CrossRef]
- Sarin, S.; Kumar, M.; Lau, G.; Abbas, Z.; Chan, H.; Chen, C.; Chen, D.; Chen, H.; Chen, P.; Chien, R. Asian-Pacific clinical practice guidelines on the management of hepatitis B: A 2015 update. Hepatol. Int. 2016, 10, 1–98. [Google Scholar] [CrossRef]
- Safaie, P.; Razeghi, S.; Rouster, S.D.; Privitera, I.; Sherman, K.E. Hepatitis D diagnostics: Utilization and testing in the United States. Virus Res. 2018, 250, 114–117. [Google Scholar] [CrossRef]
- Kang, C.; Syed, Y.Y. Bulevirtide: First Approval. Drugs 2020, 80, 1601–1605. [Google Scholar] [CrossRef]
- Yurdaydin, C.; Keskin, O.; Kalkan, Ç.; Karakaya, F.; Çalişkan, A.; Karatayli, E.; Karatayli, S.; Bozdayi, A.M.; Koh, C.; Heller, T. Optimizing lonafarnib treatment for the management of chronic delta hepatitis: The LOWR HDV-1 study. Hepatology 2018, 67, 1224–1236. [Google Scholar] [CrossRef] [Green Version]
- Al-Mahtab, M.; Bazinet, M.; Vaillant, A. Safety and efficacy of nucleic acid polymers in monotherapy and combined with immunotherapy in treatment-naive Bangladeshi patients with HBeAg+ chronic hepatitis B infection. PLoS ONE 2016, 11, e0156667. [Google Scholar] [CrossRef] [Green Version]
- Wedemeyer, H.; Bogomolov, P.; Blank, A.; Allweiss, L.; Dandri-Petersen, M.; Bremer, B.; Voronkova, N.; Schöneweis, K.; Pathil, A.; Burhenne, J. Final results of a multicenter, open-label phase 2b clinical trial to assess safety and efficacy of Myrcludex B in combination with Tenofovir in patients with chronic HBV/HDV co-infection. J. Hepatol. 2018, 68, 3. [Google Scholar] [CrossRef]
- Noureddin, M.; Gish, R. Hepatitis Delta: Epidemiology, Diagnosis and Management 36 Years after Discovery. Curr. Gastroenterol. Rep. 2014, 16, 365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wedemeyer, H.; Negro, F. Devil hepatitis D: An orphan disease or largely underdiagnosed? Gut 2019, 68, 381–382. [Google Scholar] [CrossRef] [PubMed]
- Miao, Z.; Zhang, S.; Ou, X.; Li, S.; Ma, Z.; Wang, W.; Peppelenbosch, M.P.; Liu, J.; Pan, Q. Estimating the global prevalence, disease progression, and clinical outcome of hepatitis delta virus infection. J. Infect. Dis. 2020, 221, 1677–1687. [Google Scholar] [CrossRef] [PubMed]
- Stockdale, A.J.; Kreuels, B.; Henrion, M.Y.; Giorgi, E.; Kyomuhangi, I.; de Martel, C.; Hutin, Y.; Geretti, A.M. The global prevalence of hepatitis D virus infection: Systematic review and meta-analysis. J. Hepatol. 2020, 73, 523–532. [Google Scholar] [CrossRef]
- Chen, X.; Oidovsambuu, O.; Liu, P.; Grosely, R.; Elazar, M.; Winn, V.D.; Fram, B.; Boa, Z.; Dai, H.; Dashtseren, B. A novel quantitative microarray antibody capture assay identifies an extremely high hepatitis delta virus prevalence among hepatitis B virus–infected mongolians. Hepatology 2017, 66, 1739–1749. [Google Scholar] [CrossRef] [Green Version]
- Tsatsralt-Od, B. Viral hepatitis in Mongolia: Past, present, and future. Euroasian J. Hepatogastroenterol. 2016, 6, 56. [Google Scholar] [CrossRef] [Green Version]
- Scarponi, C.F.d.O.; Silva, R.D.N.d.; Souza Filho, J.A.d.; Guerra, M.R.L.; Pedrosa, M.A.F.; Mol, M.P.G. Hepatitis delta prevalence in South America: A systematic review and meta-analysis. Rev. Soc. Bras. Med. Trop. 2019, 52, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khodjaeva, M.; Ibadullaeva, N.; Khikmatullaeva, A.; Joldasova, E.; Ismoilov, U.; Colombo, M.; Caviglia, G.P.; Rizzetto, M.; Musabaev, E. The medical impact of hepatitis D virus infection in Uzbekistan. Liver Int. 2019, 39, 2077–2081. [Google Scholar] [CrossRef] [PubMed]
- Makuwa, M.; Caron, M.; Souquière, S.; Malonga-Mouelet, G.; Mahé, A.; Kazanji, M. Prevalence and genetic diversity of hepatitis B and delta viruses in pregnant women in Gabon: Molecular evidence that hepatitis delta virus clade 8 originates from and is endemic in central Africa. J. Clin. Microbiol. 2008, 46, 754–756. [Google Scholar] [CrossRef] [Green Version]
- Shen, L.; Gu, Y.; Sun, L.; Yang, Y.; Wang, F.; Li, Y.; Bi, S. Development of a hepatitis delta virus antibody assay for study of the prevalence of HDV among individuals infected with hepatitis B virus in China. J. Med. Virol. 2012, 84, 445–449. [Google Scholar] [CrossRef] [PubMed]
- Kai, M.; Roggendorf, M.; Zhihong, J.; Shisong, X.; Guolong, Z.; Genshan, T.; Deinhardt, F. Epidemiology of hepatitis δ virus infection in China. J. Gastroenterol. Hepatol. 1988, 3, 373–377. [Google Scholar] [CrossRef]
- Gmelin, K.; Theilmann, L.; Bernhardt, R.; Kommereil, B.; Deinhardt, F.; Roggendorf, M.; Lian-jie, H. Antibodies to hepatitis delta virus cannot be detected in carriers of hepatitis B surface antigen in Wuhan area (PR China). J. Tongji Univ. 1986, 6, 198. [Google Scholar] [CrossRef] [PubMed]
- Roggendorf, M.; Mai, K.; Thian, G.; Hu, M.; Zhuang, C.; Gmelin, K.; Bernhardt, R.; Deinhardt, F. Prevalence of hepatitis delta virus infection in different provinces of China. Prog. Clin. Biol. Res. 1987, 234, 487. [Google Scholar]
- Perez-Vargas, J.; Amirache, F.; Boson, B.; Mialon, C.; Freitas, N.; Sureau, C.; Fusil, F.; Cosset, F.-L. Enveloped viruses distinct from HBV induce dissemination of hepatitis D virus in vivo. Nat. Commun. 2019, 10, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Hetzel, U.; Szirovicza, L.; Smura, T.; Prähauser, B.; Vapalahti, O.; Kipar, A.; Hepojoki, J. Identification of a novel deltavirus in Boa Constrictors. MBio 2019, 10, e00014-19. [Google Scholar] [CrossRef] [Green Version]
- Wille, M.; Netter, H.J.; Littlejohn, M.; Yuen, L.; Shi, M.; Eden, J.-S.; Klaassen, M.; Holmes, E.C.; Hurt, A.C. A divergent hepatitis D-like agent in birds. Viruses 2018, 10, 720. [Google Scholar] [CrossRef] [Green Version]
- Chang, W.-S.; Pettersson, J.H.; Le Lay, C.; Shi, M.; Lo, N.; Wille, M.; Eden, J.-S.; Holmes, E.C. Novel hepatitis D-like agents in vertebrates and invertebrates. Virus Evol. 2019, 5, vez021. [Google Scholar] [CrossRef]
- Chemin, I.; Pujol, F.H.; Scholtès, C.; Loureiro, C.L.; Amirache, F.; Levrero, M.; Zoulim, F.; Pérez-Vargas, J.; Cosset, F.L. Preliminary Evidence for Hepatitis Delta Virus Exposure in Patients Who Are Apparently Not Infected With Hepatitis B Virus. Hepatology 2021, 73, 861. [Google Scholar] [CrossRef]
- Cappy, P.; Lucas, Q.; Kankarafou, N.; Sureau, C.; Laperche, S. No Evidence of HCV-Assisted HDV Propagation in a Large Cohort of Hepatitis C Positive Blood Donors. J. Infect. Dis. 2020, 223, 1376–1380. [Google Scholar] [CrossRef]
- Pflüger, L.S.; Schulze zur Wiesch, J.; Polywka, S.; Lütgehetmann, M. Hepatitis delta virus propagation enabled by hepatitis C virus—Scientifically intriguing, but is it relevant to clinical practice? J. Viral Hepat. 2021, 28, 213–216. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Yu, G.; Sun, H.; Lv, J.; Wang, M.; Kong, F.; Zhang, M.; Chi, X.; Wang, X.; Wu, R. Use of parenteral caffeinum natrio-benzoicum: An underestimated risk factor for HCV transmission in China. BMC Public Health 2015, 15, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Lempp, F.A.; Sakin, V.; Nkongolo, S.; Schnitzler, P.; Wedemeyer, H.; Gal, F.; Gordien, E.; Yurdaydin, C.; Urban, S. A rapid point-of-care device for the diagnosis of hepatitis delta virus infection. J. Hepatol. 2019, 70, e32–e33. [Google Scholar] [CrossRef]
- Lempp, F.A.; Schlund, F.; Rieble, L.; Nussbaum, L.; Link, C.; Zhang, Z.; Ni, Y.; Urban, S. Recapitulation of HDV infection in a fully permissive hepatoma cell line allows efficient drug evaluation. Nat. Commun. 2019, 10, 1–11. [Google Scholar] [CrossRef]
- Le Gal, F.; Brichler, S.; Drugan, T.; Alloui, C.; Roulot, D.; Pawlotsky, J.M.; Dény, P.; Gordien, E. Genetic diversity and worldwide distribution of the deltavirus genus: A study of 2,152 clinical strains. Hepatology 2017, 66, 1826–1841. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, V.; Brichler, S.; Khan, E.; Chami, M.; Dény, P.; Kremsdorf, D.; Gordien, E. Large hepatitis delta antigen activates STAT-3 and NF-κB via oxidative stress. J. Viral Hepat. 2012, 19, 744–753. [Google Scholar] [CrossRef] [PubMed]
- Roulot, D.; Brichler, S.; Layese, R.; BenAbdesselam, Z.; Zoulim, F.; Thibault, V.; Scholtes, C.; Roche, B.; Castelnau, C.; Poynard, T.; et al. Origin, HDV genotype and persistent viremia determine outcome and treatment response in patients with chronic hepatitis delta. J. Hepatol. 2020, 73, 1046–1062. [Google Scholar] [CrossRef]
- Gaeta, G.B.; Stornaiuolo, G.; Precone, D.F.; Lobello, S.; Chiaramonte, M.; Stroffolini, T.; Colucci, G.; Rizzetto, M. Epidemiological and clinical burden of chronic hepatitis B virus/hepatitis C virus infection. A multicenter Italian study. J. Hepatol. 2003, 39, 1036–1041. [Google Scholar] [CrossRef]
- Reinheimer, C.; Doerr, H.; Berger, A. Hepatitis delta: On soft paws across Germany. Infection 2012, 40, 621–625. [Google Scholar] [CrossRef] [PubMed]
- Bissinger, A.; Berg, C. Is the HDV seroprevalence in patients admitted to a university hospital representative for all HBV carriers in Germany? Infection 2013, 41, 1031–1032. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Roggenbach, I.; Chi, X.; Lempp, F.A.; Qu, B.; Walter, L.; Wu, R.; Gao, X.; Schnitzler, P.; Ding, Y.; Urban, S.; et al. HDV Seroprevalence in HBsAg-Positive Patients in China Occurs in Hotspots and Is Not Associated with HCV Mono-Infection. Viruses 2021, 13, 1799. https://doi.org/10.3390/v13091799
Roggenbach I, Chi X, Lempp FA, Qu B, Walter L, Wu R, Gao X, Schnitzler P, Ding Y, Urban S, et al. HDV Seroprevalence in HBsAg-Positive Patients in China Occurs in Hotspots and Is Not Associated with HCV Mono-Infection. Viruses. 2021; 13(9):1799. https://doi.org/10.3390/v13091799
Chicago/Turabian StyleRoggenbach, Imme, Xiumei Chi, Florian A. Lempp, Bingqian Qu, Lisa Walter, Ruihong Wu, Xiuzhu Gao, Paul Schnitzler, Yanhua Ding, Stephan Urban, and et al. 2021. "HDV Seroprevalence in HBsAg-Positive Patients in China Occurs in Hotspots and Is Not Associated with HCV Mono-Infection" Viruses 13, no. 9: 1799. https://doi.org/10.3390/v13091799