
Histopathological Analysis of Adrenal Glands after Simian Varicella Virus Infection

 , , ,
, , ,
Abstract
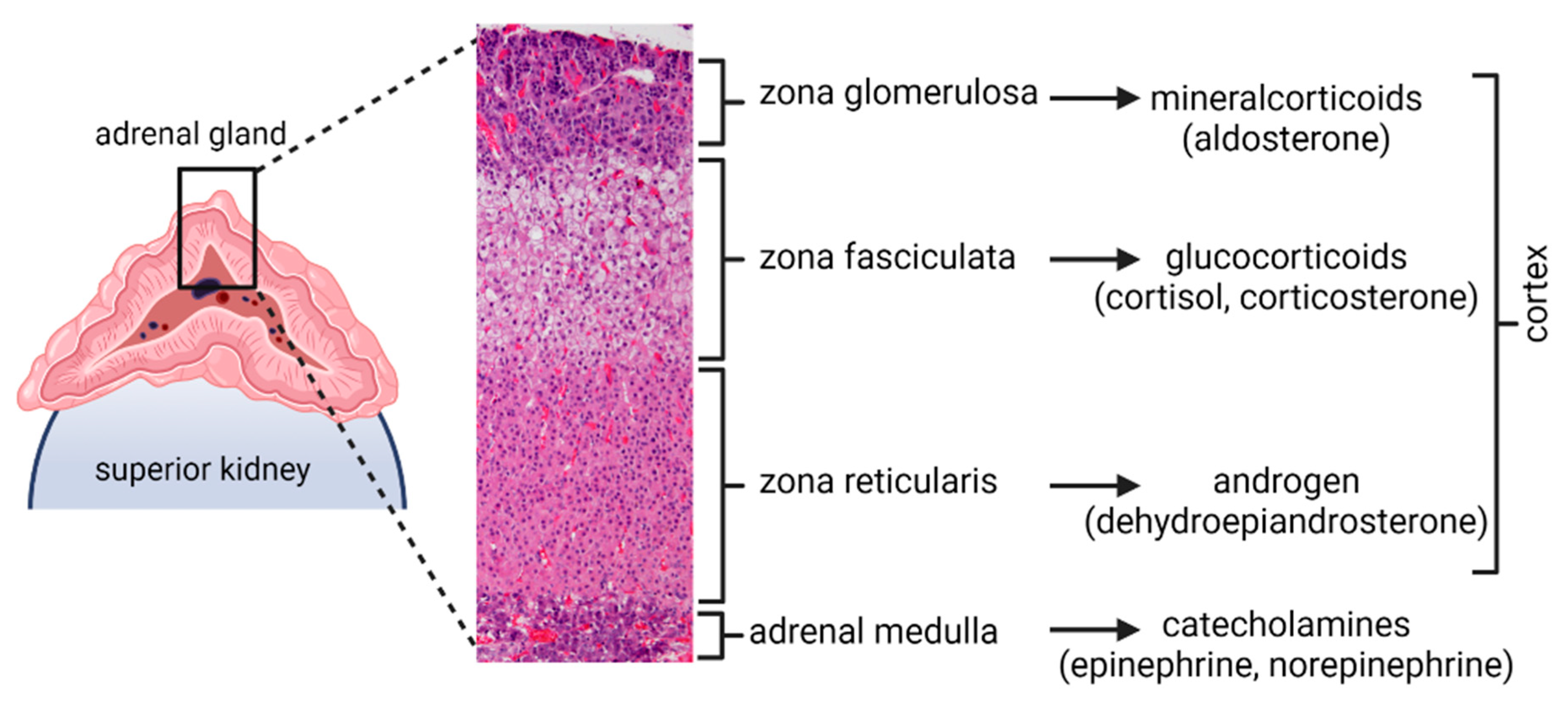
:1. Introduction
2. Materials and Methods
2.1. Animal Infection and Tissue Collection
2.2. Immunohistochemistry
2.3. DNA Extraction and Quantitative PCR
3. Results
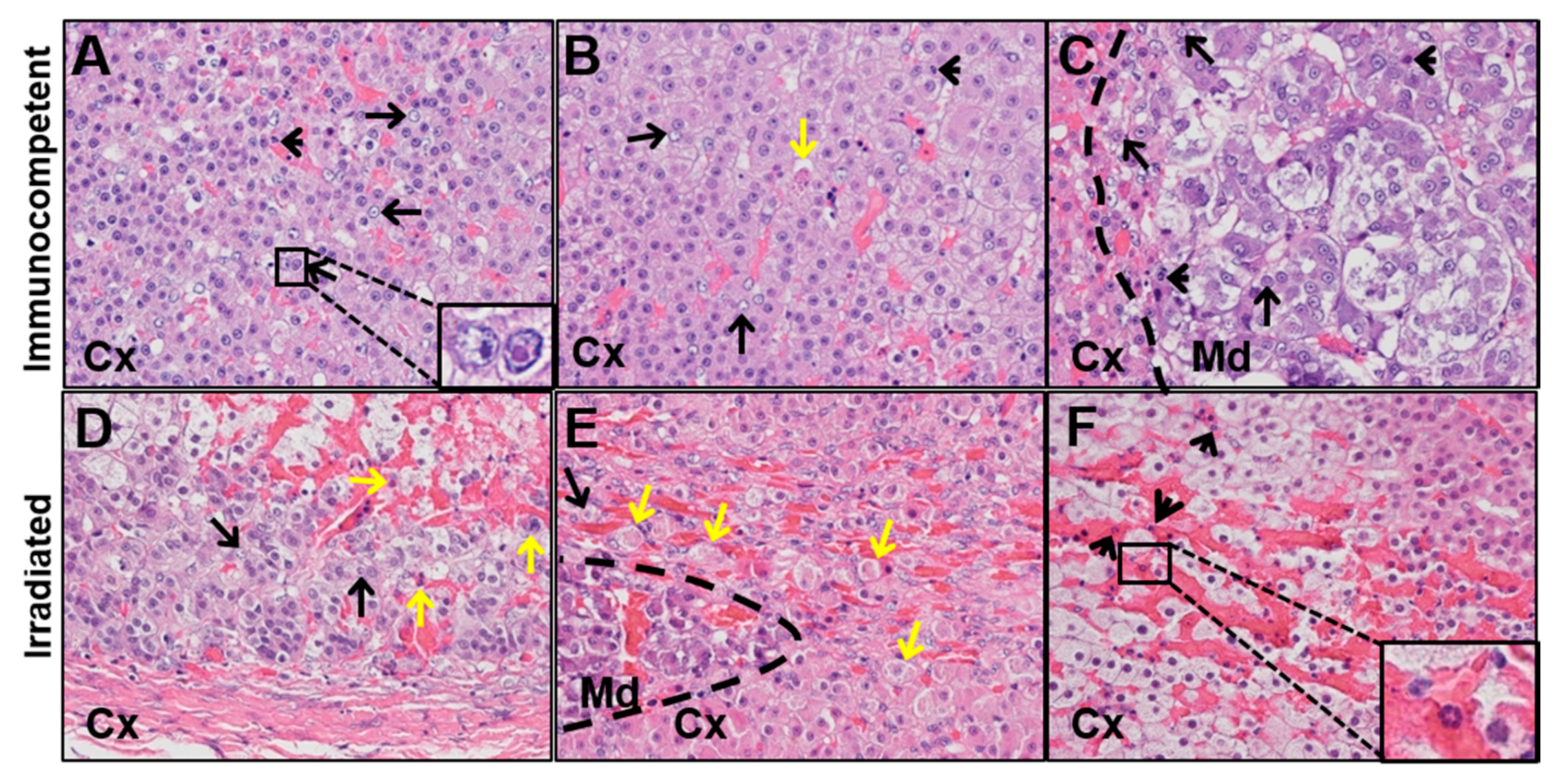
3.1. Adrenal Glands from Macaques with Varicella Contain Cowdry A Inclusion Bodies, Polymorphonuclear Cells (PMNs), Necrotic Cells, and Hemorrhagic Regions
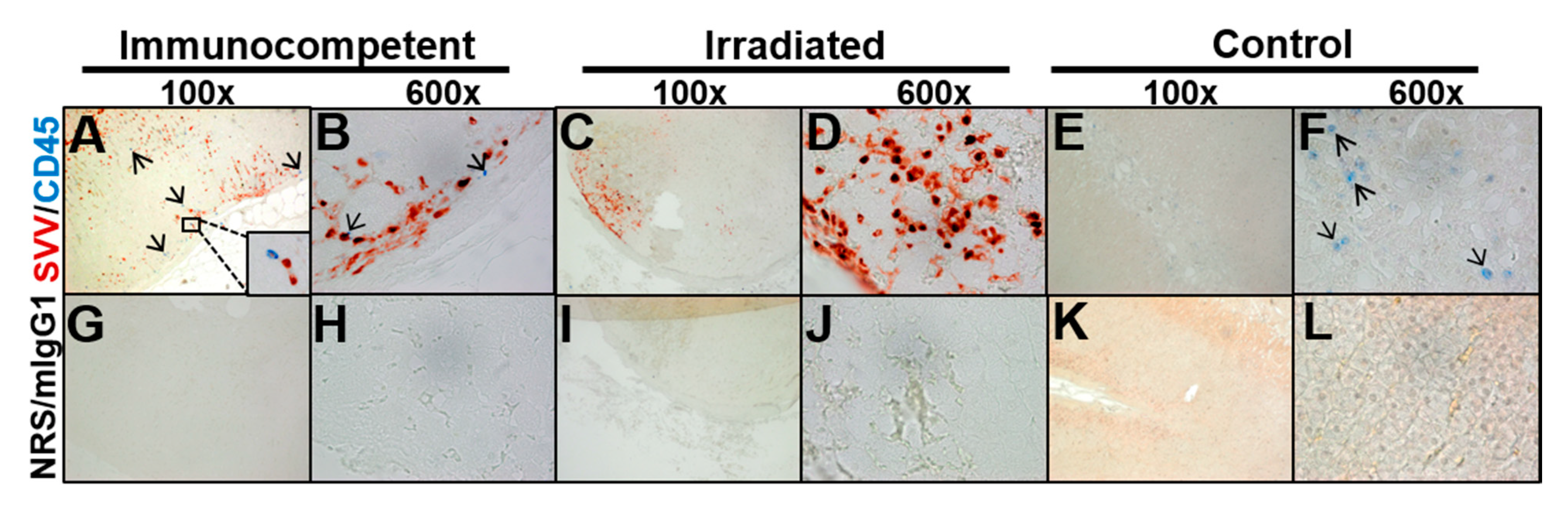
3.2. Adrenal Glands from Macaques with Varicella Contain SVV Antigen Yet Differ in CD45 Expression
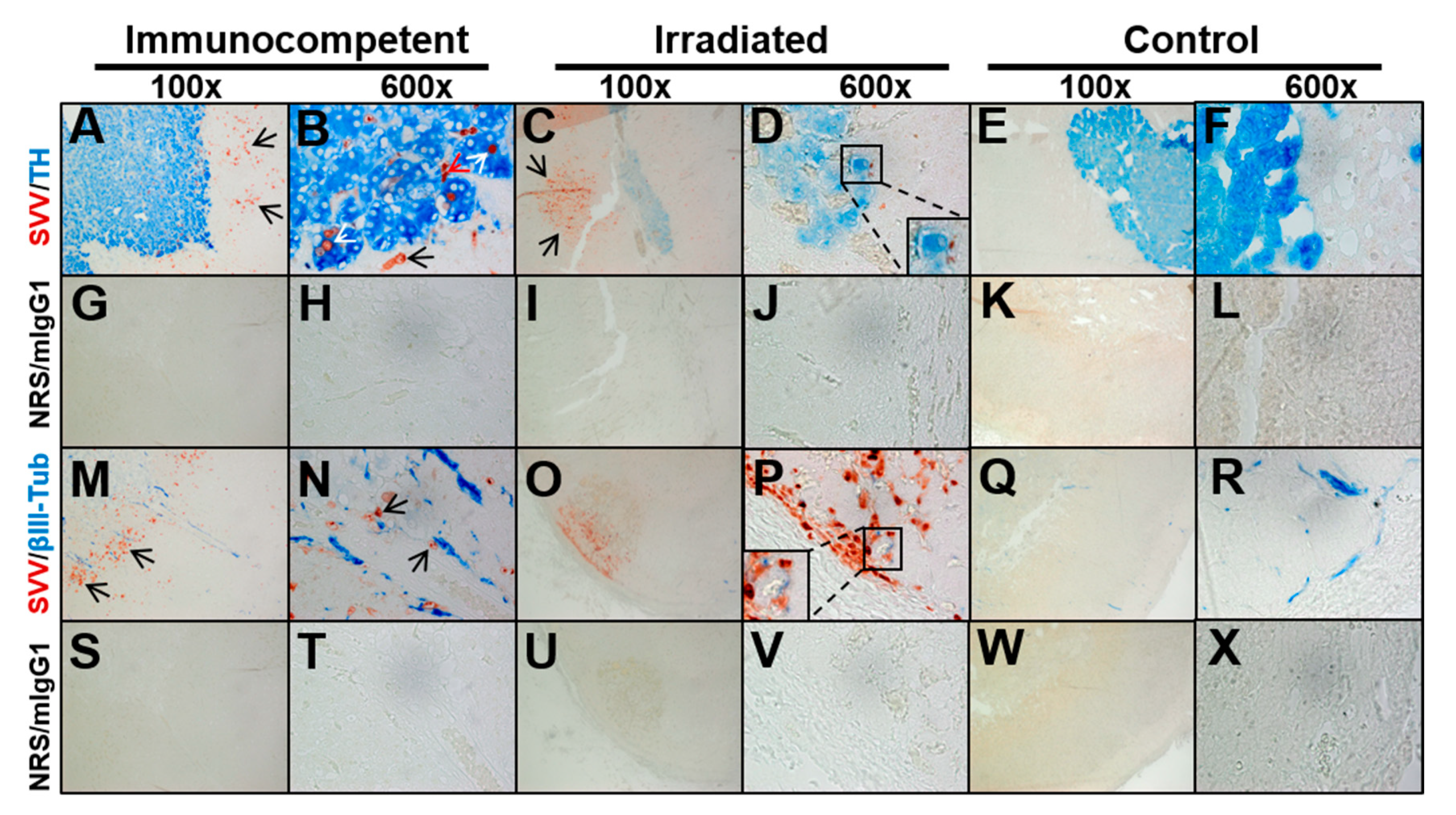
3.3. Adrenal Glands from Macaques with Varicella Have Varying Distributions of SVV Antigen in Cortex and Medulla without a Clear Association with Nerve Fibers
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zerboni, L.; Sen, N.; Oliver, S.L.; Arvin, A.M. Molecular mechanisms of varicella zoster virus pathogenesis. Nat. Rev. Genet. 2014, 12, 197–210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilden, D.H.; Vafai, A.; Shtram, Y.; Becker, Y.; Devlin, M.; Wellish, M. Varicella-zoster virus DNA in human sensory ganglia. Nat. Cell Biol. 1983, 306, 478–480. [Google Scholar] [CrossRef]
- Kennedy, P.G.E.; Grinfeld, E.; Gow, J.W. Latent varicella-zoster virus is located predominantly in neurons in human trigeminal ganglia. Proc. Natl. Acad. Sci. USA 1998, 95, 4658–4662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gershon, A.A.; Chen, J.; Davis, L.; Krinsky, C.; Cowles, R.; Reichard, R.; Gershon, M. Latency of varicella zoster virus in dorsal root, cranial, and enteric ganglia in vaccinated children. Trans. Am. Clin. Clim. Assoc. 2012, 123, 17–35. [Google Scholar]
- Nagel, M.A.; Gilden, D. Neurological complications of VZV reactivation. Curr. Opin. Neurol. 2014, 27, 356–360. [Google Scholar] [CrossRef] [Green Version]
- Badani, H.; White, T.; Schulick, N.; Raeburn, C.D.; Topkaya, I.; Gilden, D.; Nagel, M.A. Frequency of varicella zoster virus DNA in human adrenal glands. J. NeuroVirol. 2016, 22, 400–402. [Google Scholar] [CrossRef] [Green Version]
- Herman, J.P.; Ostrander, M.M.; Mueller, N.K.; Figueiredo, H. Limbic system mechanisms of stress regulation: Hypothala-mo-pituitary-adrenocortical axis. Prog. Neuropsychopharmacol. Biol. Psychiatry 2005, 29, 1201–1213. [Google Scholar] [CrossRef]
- Pariante, C.M.; Lightman, S.L. The HPA axis in major depression: Classical theories and new developments. Trends Neurosci. 2008, 31, 464–468. [Google Scholar] [CrossRef]
- Rudkowski, Z.; Rutowski, R. Zespół Waterhouse-Friderichsena w przebiegu ospy wietrznej u dziecka z toksoplazmoza [Wa-terhouse-Friderichsen syndrome during varicella in a child with toxoplasmosis]. Pediatr. Pol. 1974, 49, 1023–1026. [Google Scholar]
- Montgomery, R.R.; Ólafsson, M. Waterhouse-Friderichsen syndrome in varicella. Ann. Intern. Med. 1960, 53, 576–579. [Google Scholar] [CrossRef] [PubMed]
- Krajewska, B. Two cases of hemorrhage into the suprarerenal glands in the course of varicella. Helv. Paediatr. Acta 1960, 15, 71–73. [Google Scholar]
- Heitz, A.F.N.; Hofstee, H.M.A.; Gelinck, L.B.S.; Puylaert, J.B. A rare case of Waterhouse-Friderichsen syndrome during pri-mary varicella zoster infection. Neth. J. Med. 2017, 75, 351–353. [Google Scholar]
- Soike, K.F.; Rangan, S.R.; Gerone, P.J. Viral disease models in primates. Adv. Vet. Sci. Comp. Med. 1984, 28, 151–199. [Google Scholar] [CrossRef] [PubMed]
- Dueland, A.N.; Martin, J.R.; Devlin, M.E.; Wellish, M.; Mahalingam, R.; Cohrs, R.; Soike, K.F.; Gilden, D.H. Acute simian varicella infection. Clinical, laboratory, pathologic, and virologic features. Lab. Investig. 1992, 66, 762–773. [Google Scholar] [PubMed]
- White, T.M.; Gilden, D.H.; Mahalingam, R. An animal model of varicella virus infection. Brain Pathol. 2001, 11, 475–479. [Google Scholar] [CrossRef] [PubMed]
- Mahalingam, R.; Traina-Dorge, V.; Wellish, M.; Lorino, R.; Sanford, R.; Ribka, E.P.; Alleman, S.J.; Brazeau, E.; Gilden, D.H. Simian varicella virus reactivation in cynomolgus monkeys. Virology 2007, 368, 50–59. [Google Scholar] [CrossRef] [Green Version]
- Messaoudi, I.; Barron, A.; Wellish, M.; Engelmann, F.; Legasse, A.; Planer, S.; Gilden, D.; Nikolich-Zugich, J.; Mahalingam, R. Simian varicella virus infection of rhesus macaques recapitulates essential features of varicella zoster virus infection in humans. PLoS Pathog. 2009, 5, e1000657. [Google Scholar] [CrossRef] [Green Version]
- Mahalingam, R.; Traina-Dorge, V.; Wellish, M.; Deharo, E.; Singletary, M.L.; Ribka, E.P.; Sanford, R.; Gilden, N. Latent simian varicella virus reactivates in monkeys treated with tacrolimus with or without exposure to irradiation. J. NeuroVirol. 2010, 16, 342–354. [Google Scholar] [CrossRef] [Green Version]
- Kolappaswamy, K.; Mahalingam, R.; Traina-Dorge, V.; Shipley, S.T.; Gilden, D.H.; Kleinschmidt-DeMasters, B.K.; McLeod, C.G., Jr.; Hungerford, L.L.; DeTolla, L.J. Disseminated simian varicella virus infection in an irradiated rhesus macaque. J. Virol. 2007, 81, 411–415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahalingam, R.; Clarke, P.; Wellish, M.; Dueland, A.N.; Soike, K.F.; Gilden, D.H.; Cohrs, R. Prevalence and distribution of latent simian varicella virus DNA in monkey ganglia. Virology 1992, 188, 193–197. [Google Scholar] [CrossRef]
- Mahalingam, R.; Wellish, M.; Cohrs, R.; Debrus, S.; Piette, J.; Rentier, B.; Gilden, D.H. Expression of protein encoded by varicella-zoster virus open reading frame 63 in latently infected human ganglionic neurons. Proc. Natl. Acad. Sci. USA 1996, 93, 2122–2124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilden, D.; White, T.; Kandasamy, B.; Lear-Kaul, K.; Holmes, D.B.; Bennett, J.L.; Cohrs, R.J.; Mahalingam, R.; Mandava, N.; Eberhart, C.G.; et al. Prevalence and distribution of VZV in temporal arteries of patients with giant cell arteritis. Neurology 2015, 84, 1948–1955. [Google Scholar] [CrossRef] [Green Version]
- Clayton, N.P.; Khan-Malek, R.C.; Authier, S.; Dangler, C.A.; Zhang, D.; Ascah, A.; Gains, M.; Gardner, B.; Mockbee, C.; Keutzer, J.M.; et al. Sargramostim (rhu GM-CSF) improves survival of non-human primates with severe bone marrow suppression after acute, high-dose, whole-body irradiation. Radiat. Res. 2020, 195, 191–199. [Google Scholar] [CrossRef]
- Potratz, D.; Brake, B.; Dienes, H.P.; Schulz, T.; Hosp, M.; Dierich, M.P.; Falke, D. Herpes simplex virus type 1 and 2 in the adrenal glands: Replication and histopathology. Arch. Virol. 1986, 90, 207–222. [Google Scholar] [CrossRef]
- Wiegand, H.; Dienes, H.P.; Schirmacher, P.; Podlech, J.; Bohl, K.; Bohle, M.; Neumann-Haefelin, D.; Falke, D. Colonization of adrenal glands and ovaries of mice by variants of HSV 1 and 2. Arch. Virol. 1991, 117, 237–249. [Google Scholar] [CrossRef]
- Podlech, J.; Hengerer, F.; Fleck, M.; Walev, I.; Falke, D.; Podlech, J.; Hengerer, F.; Fleck, M.; Walev, I.; Falke, D. Replication of herpes simplex virus type 1 and 2 in the medulla of the adrenal gland after vaginal infection of mice. Arch. Virol. 1996, 141, 1999–2008. [Google Scholar] [CrossRef]
- Nakamura, Y.; Yamamoto, S.; Tanaka, S.; Yano, H.; Nishimura, G.; Saito, Y.; Tanaka, T.; Tanimura, A.; Hirose, F.; Fukuda, S.; et al. Herpes simplex viral infection in human neonates: An immunohistochemical and electron microscopic study. Hum. Pathol. 1985, 16, 1091–1097. [Google Scholar] [CrossRef]
- Su, C.-C.; Lu, J.-J.; Perng, C.-L.; Chen, C.-S.; Lee, C.-C. Adrenal insufficiency combined with gastric cardia ulcer due to herpes simplex virus type 1 infection. J. Formos. Med. Assoc. 2002, 101, 719–721. [Google Scholar]
- Ben-Hur, T.; Conforti, N.; Itzik, A.; Weidenfeld, J. Effects of HSV-1, a neurotropic virus, on the hypothalam-ic-pituitary-adrenocortical axis in rats. Brain Res. 1995, 702, 17–22. [Google Scholar] [CrossRef]
- Ben Hur, T.; Rosenthal, J.; Itzik, A.; Weidenfeld, J. Adrenocortical activation by herpes virus: Involvement of IL-1 beta and central noradrenergic system. NeuroReport 1996, 7, 927–931. [Google Scholar] [PubMed]
- Enquist, L.W.; Husak, P.J.; Banfield, B.W.; Smith, G.A. Infection and spread of alphaherpesviruses in the nervous system. Adv. Virus Res. 1998, 51, 237–347. [Google Scholar] [CrossRef] [PubMed]
- Strack, A.; Sawyer, W.; Hughes, J.; Platt, K.; Loewy, A. A general pattern of CNS innervation of the sympathetic outflow demonstrated by transneuronal pseudorabies viral infections. Brain Res. 1989, 491, 156–162. [Google Scholar] [CrossRef]
- Kerman, I.A.; Enquist, L.W.; Watson, S.J.; Yates, B.J. Brainstem substrates of sympatho-motor circuitry identified using trans-synaptic tracing with pseudorabies virus recombinants. J. Neurosci. 2003, 23, 4657–4666. [Google Scholar] [CrossRef] [PubMed]
- Field, H.J.; Hill, T.J. The pathogenesis of pseudorabies in mice following peripheral inoculation. J. Gen. Virol. 1974, 23, 145–157. [Google Scholar] [CrossRef] [PubMed]
- Sehl, J.; Teifke, J.P. Comparative pathology of pseudorabies in different naturally and experimentally infected species—A review. Pathogens 2020, 9, 633. [Google Scholar] [CrossRef]
- Gotlieb, N.; Albaz, E.; Shaashua, L.; Sorski, L.; Matzner, P.; Rosenne, E.; Amram, B.; Benbenishty, A.; Golomb, E.; Ben-Eliyahu, S. Regeneration of functional adrenal tissue following bilateral adrenalectomy. Endocrinology 2017, 159, 248–259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santana, M.F.; Borba, M.G.S.; Naveca, F.G.; Xavier, M.S.; Monteiro, W.M.; João, G.A.P.; Hajjar, L.A.; Ordi, J.; Lacerda, M.V.G.; Ferreira, L.C.L.; et al. Case report: Adrenal pathology findings in severe COVID-19: An autopsy study. Am. J. Trop. Med. Hyg. 2020, 103, 1604–1607. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Immunocompetent | Irradiated | |||
|---|---|---|---|---|
| Feature | Cortex | Medulla | Cortex | Medulla |
| SVV antigen | ++ | + | ++ | ― |
| Cowdry A inclusions | + | + | + | ― |
| Necrotic cells | + | ― | ++ | ― |
| PMCs | + | + | + | ― |
| Hemorrhage | + | + | ++ | ++ |
| CD45 | + | + | ― | ― |
| Tyrosine hydroxylase | ― | + | ― | + |
| βIII tubulin | ++ | ― | + | ― |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Niemeyer, C.S.; Mescher, T.; Griggs, R.; Orlicky, D.J.; Wilkerson, G.K.; Bubak, A.N.; Hassell, J.E., Jr.; Feia, B.; Mahalingam, R.; Traina-Dorge, V.; et al. Histopathological Analysis of Adrenal Glands after Simian Varicella Virus Infection. Viruses 2021, 13, 1245. https://doi.org/10.3390/v13071245
Niemeyer CS, Mescher T, Griggs R, Orlicky DJ, Wilkerson GK, Bubak AN, Hassell JE Jr., Feia B, Mahalingam R, Traina-Dorge V, et al. Histopathological Analysis of Adrenal Glands after Simian Varicella Virus Infection. Viruses. 2021; 13(7):1245. https://doi.org/10.3390/v13071245
Chicago/Turabian StyleNiemeyer, Christy S., Teresa Mescher, Rocio Griggs, David J. Orlicky, Gregory K. Wilkerson, Andrew N. Bubak, James E. Hassell, Jr., Brittany Feia, Ravi Mahalingam, Vicki Traina-Dorge, and et al. 2021. "Histopathological Analysis of Adrenal Glands after Simian Varicella Virus Infection" Viruses 13, no. 7: 1245. https://doi.org/10.3390/v13071245