
Characterization of a Cell Culture System of Persistent Hepatitis E Virus Infection in the Human HepaRG Hepatic Cell Line


Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Virus
2.3. Virus Inoculation and Passages
2.4. Experimental Infections
2.5. RNA Extraction and Quantification of HEV RNA by TaqMan RT-PCR
2.6. Immunoblot Analysis
2.7. Immunostaining and Fluorescent Microscopy
2.8. HEV Enzyme-Linked Immunosorbent Assay (ELISA)
2.9. Cell Viability Test
2.10. Whole Genome Sequencing and Sequence Analysis
2.11. Phylogenetic Analysis
2.12. Statistical Analyses
3. Results
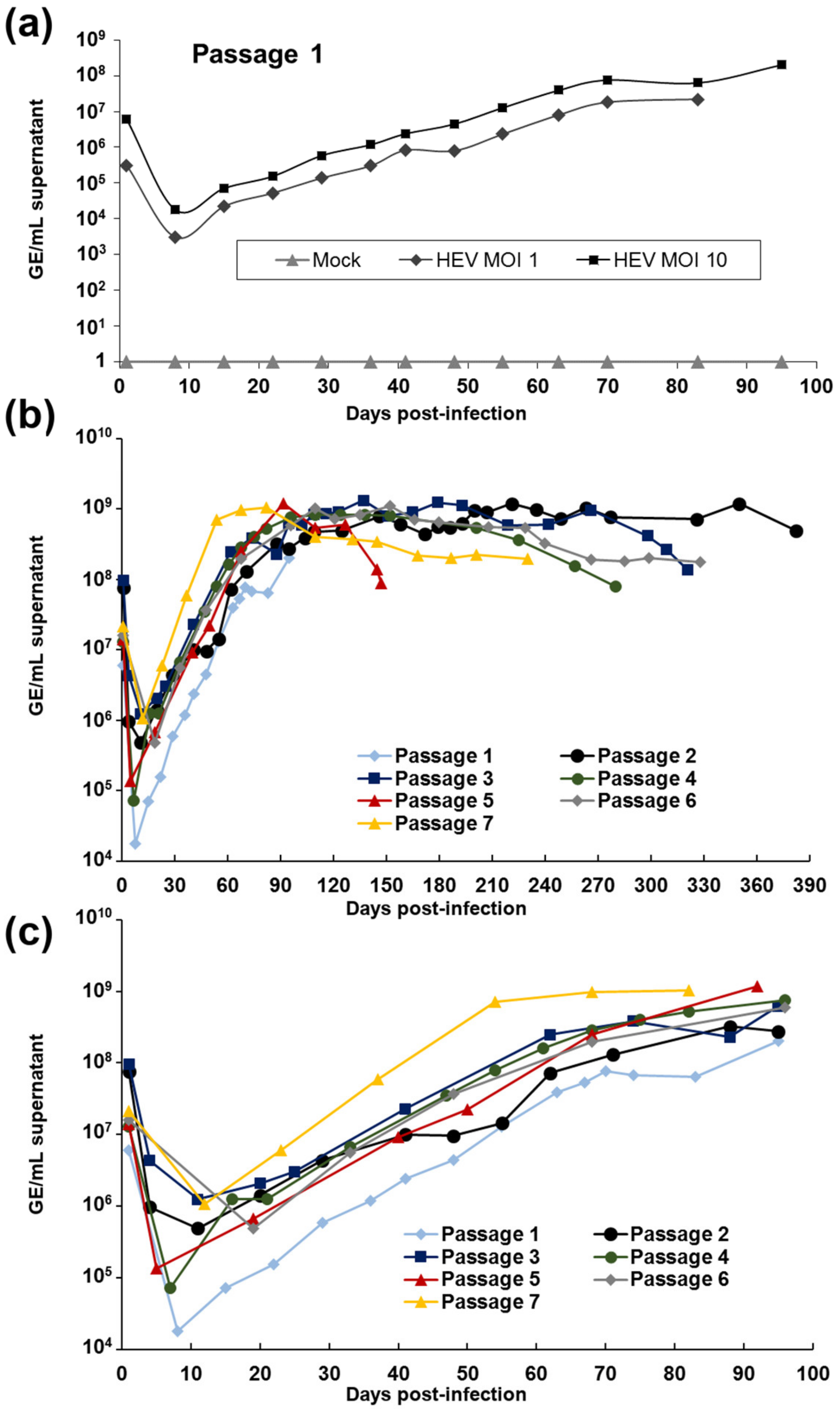
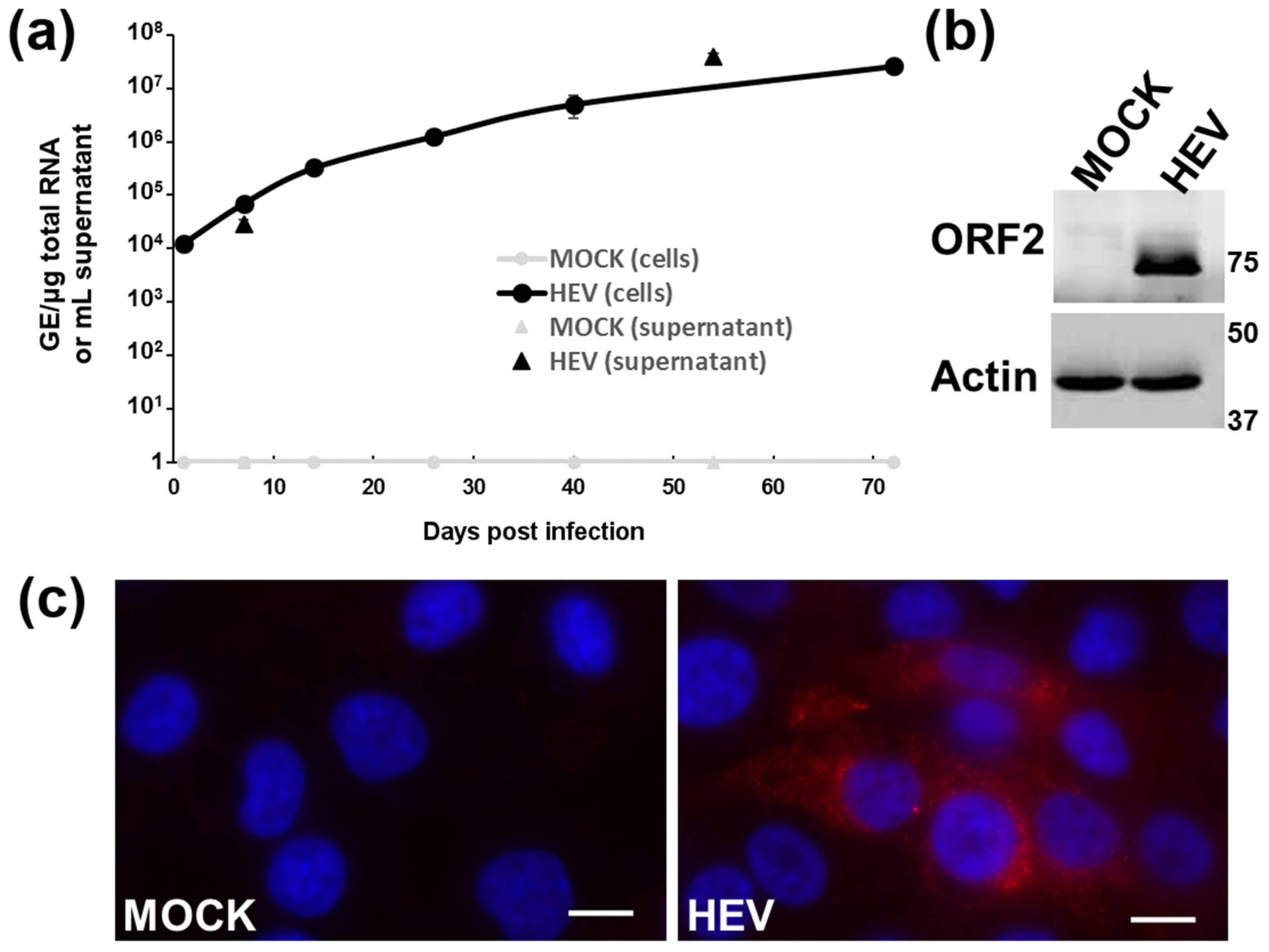
3.1. Culture of a Human HEV-3f Strain in Differentiated HepaRG Cells
3.2. Intracellular Detection of HEV in Differentiated HepaRG Cells
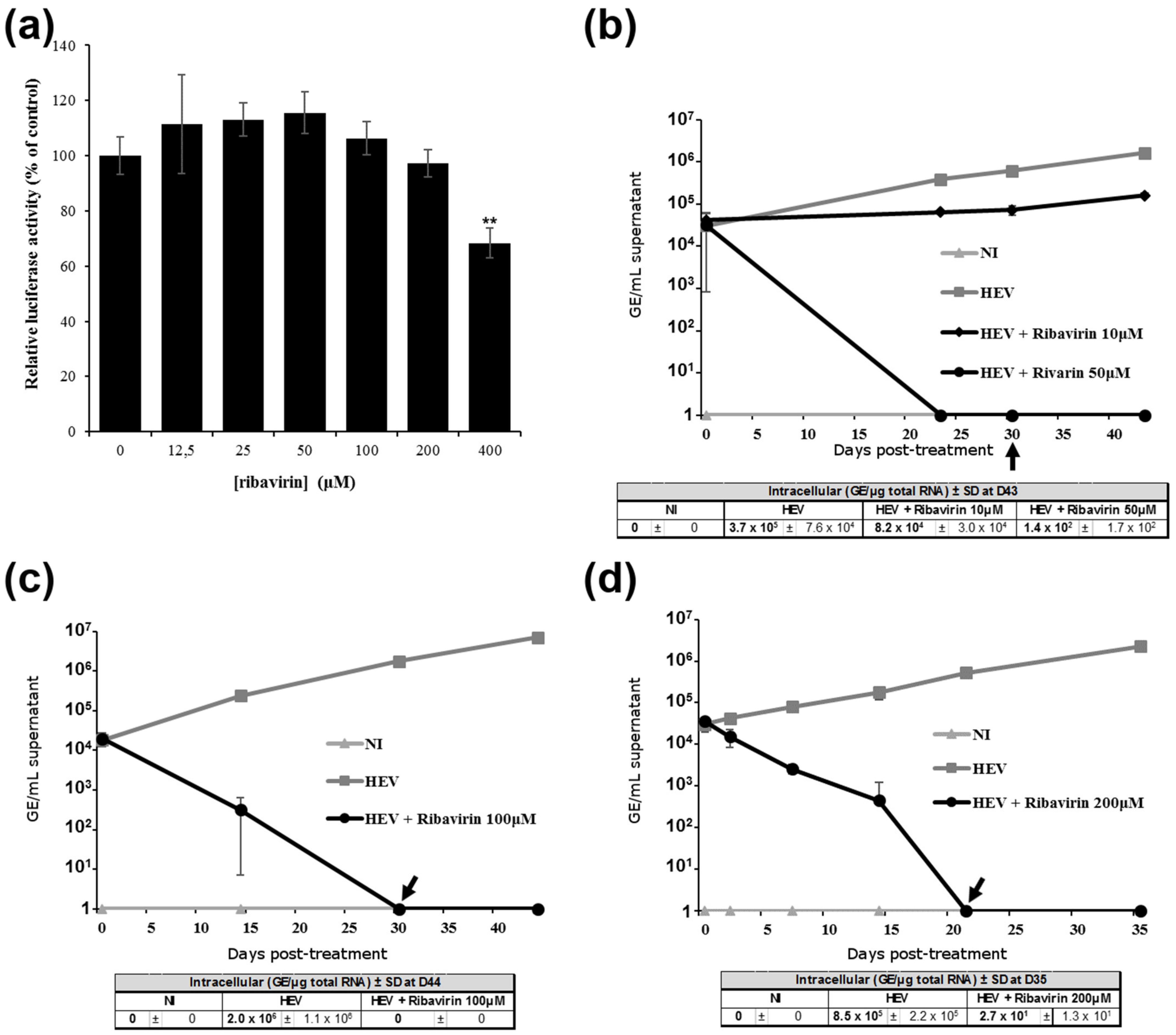
3.3. Ribavirin Inhibits HEV Replication in Differentiated HepaRG Cells
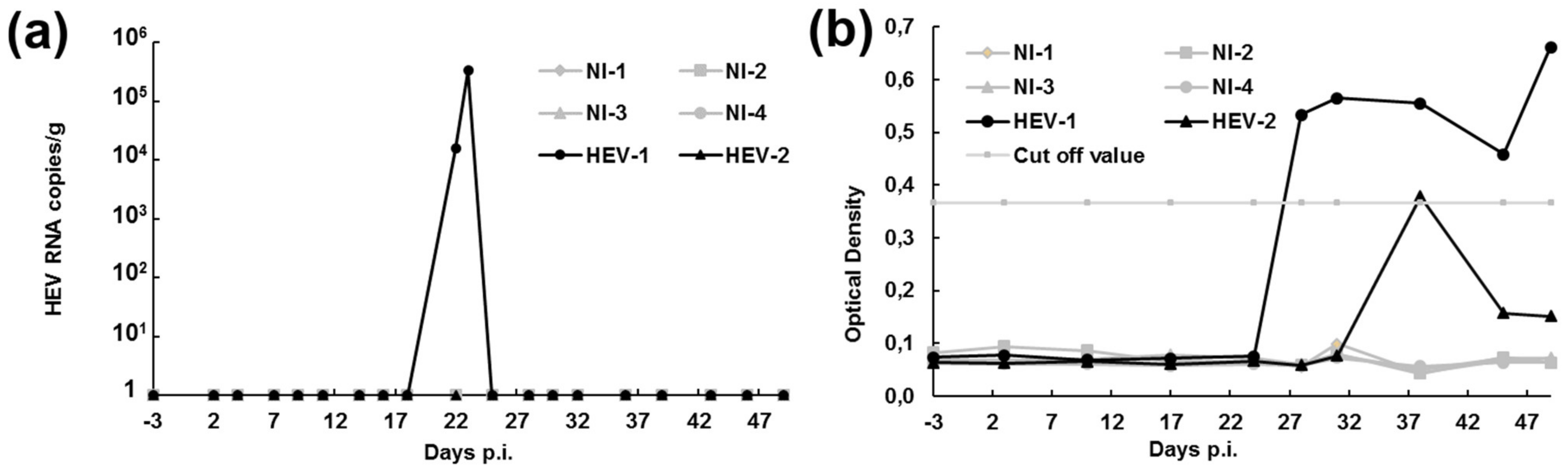
3.4. HEV Remains Infectious In Vivo in Pigs after Several Passages in HumanCcells
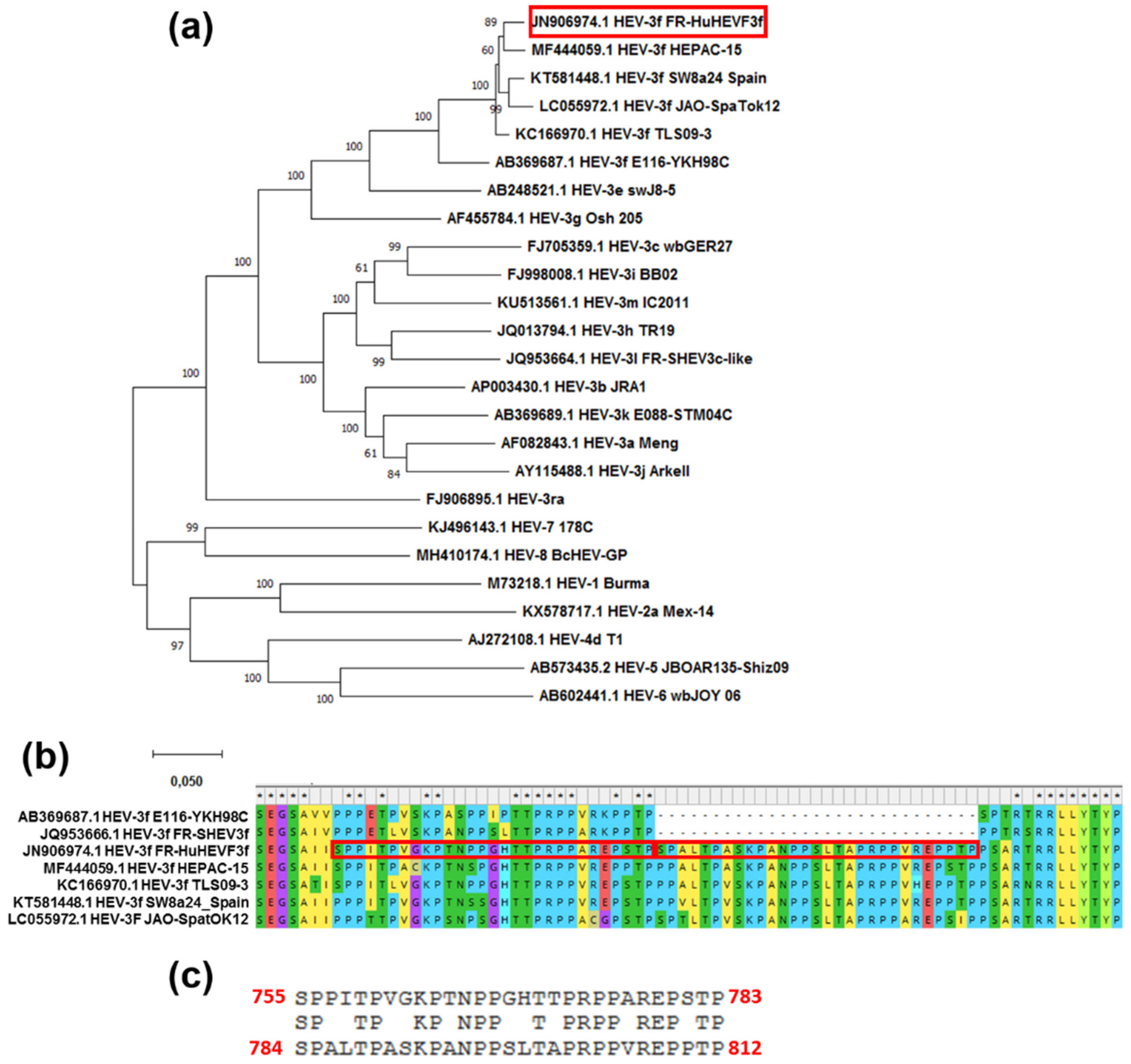
3.5. FR-HuHEVF3f Has an Insertion within the ORF1 HVR
3.6. Evolution of HEV Genome Sequence after Seven Passages in Human Hepatocytes
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Smith, D.B.; Simmonds, P.; Jameel, S.; Emerson, S.U.; Harrison, T.J.; Meng, X.-J.; Okamoto, H.; Van der Poel, W.H.M.; Purdy, M.A. Consensus proposals for classification of the family hepeviridae. J. Gen. Virol. 2014. [Google Scholar] [CrossRef]
- Ahmad, I.; Holla, R.P.; Jameel, S. Molecular Virology of Hepatitis E Virus. Virus Res. 2011, 161, 47–58. [Google Scholar] [CrossRef] [PubMed]
- Koonin, E.V.; Gorbalenya, A.E.; Purdy, M.A.; Rozanov, M.N.; Reyes, G.R.; Bradley, D.W. Computer-Assisted Assignment of Functional Domains in the Nonstructural Polyprotein of Hepatitis E Virus: Delineation of an Additional Group of Positive-Strand RNA Plant and Animal Viruses. Proc. Natl. Acad. Sci. USA 1992, 89, 8259–8263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, X.; Ying, D.; Lhomme, S.; Tang, Z.; Walker, C.M.; Xia, N.; Zheng, Z.; Feng, Z. Origin, Antigenicity, and Function of a Secreted Form of ORF2 in Hepatitis E Virus Infection. Proc. Natl. Acad. Sci. USA 2018, 115, 4773–4778. [Google Scholar] [CrossRef] [Green Version]
- Nair, V.P.; Anang, S.; Subramani, C.; Madhvi, A.; Bakshi, K.; Srivastava, A.; Shalimar, N.; Nayak, B.; Ranjith Kumar, C.T.; Surjit, M. Endoplasmic Reticulum Stress Induced Synthesis of a Novel Viral Factor Mediates Efficient Replication of Genotype-1 Hepatitis E Virus. Plos Pathog. 2016, 12, e1005521. [Google Scholar] [CrossRef] [PubMed]
- Pavio, N.; Meng, X.-J.; Doceul, V. Zoonotic Origin of Hepatitis E. Curr. Opin. Virol. 2015, 10, 34–41. [Google Scholar] [CrossRef]
- Doceul, V.; Bagdassarian, E.; Demange, A.; Pavio, N. Zoonotic Hepatitis E Virus: Classification, Animal Reservoirs and Transmission Routes. Viruses 2016, 8, 270. [Google Scholar] [CrossRef]
- Lee, G.-H.; Tan, B.-H.; Chi-Yuan, T.E.; Lim, S.-G.; Dan, Y.-Y.; Wee, A.; Aw, P.P.K.; Zhu, Y.; Hibberd, M.L.; Tan, C.-K.; et al. Chronic Infection With Camelid Hepatitis E Virus in a Liver Transplant Recipient Who Regularly Consumes Camel Meat and Milk. Gastroenterology 2016, 150, 355–357.e3. [Google Scholar] [CrossRef] [Green Version]
- Dalton, H.R.; Bendall, R.P.; Keane, F.E.; Tedder, R.S.; Ijaz, S. Persistent Carriage of Hepatitis E Virus in Patients with HIV Infection. N. Engl. J. Med. 2009, 361, 1025–1027. [Google Scholar] [CrossRef]
- Kamar, N.; Selves, J.; Mansuy, J.-M.; Ouezzani, L.; Péron, J.-M.; Guitard, J.; Cointault, O.; Esposito, L.; Abravanel, F.; Danjoux, M.; et al. Hepatitis E Virus and Chronic Hepatitis in Organ-Transplant Recipients. N. Engl. J. Med. 2008, 358, 811–817. [Google Scholar] [CrossRef] [Green Version]
- Ollier, L. Chronic Hepatitis After Hepatitis E Virus Infection in a Patient With Non-Hodgkin Lymphoma Taking Rituximab. Ann. Intern. Med. 2009, 150, 430. [Google Scholar] [CrossRef] [PubMed]
- Kamar, N.; Garrouste, C.; Haagsma, E.B.; Garrigue, V.; Pischke, S.; Chauvet, C.; Dumortier, J.; Cannesson, A.; Cassuto-Viguier, E.; Thervet, E.; et al. Factors Associated with Chronic Hepatitis in Patients with Hepatitis E Virus Infection Who Have Received Solid Organ Transplants. Gastroenterology 2011, 140, 1481–1489. [Google Scholar] [CrossRef] [PubMed]
- Kamar, N.; Abravanel, F.; Lhomme, S.; Rostaing, L.; Izopet, J. Hepatitis E Virus: Chronic Infection, Extra-Hepatic Manifestations, and Treatment. Clin. Res. Hepatol. Gastroenterol. 2014. [Google Scholar] [CrossRef] [PubMed]
- Kamar, N.; Izopet, J.; Tripon, S.; Bismuth, M.; Hillaire, S.; Dumortier, J.; Radenne, S.; Coilly, A.; Garrigue, V.; D’Alteroche, L.; et al. Ribavirin for Chronic Hepatitis E Virus Infection in Transplant Recipients. N. Engl. J. Med. 2014, 370, 1111–1120. [Google Scholar] [CrossRef] [Green Version]
- Debing, Y.; Gisa, A.; Dallmeier, K.; Pischke, S.; Bremer, B.; Manns, M.; Wedemeyer, H.; Suneetha, P.V.; Neyts, J. A Mutation in the Hepatitis E Virus RNA Polymerase Promotes Its Replication and Associates With Ribavirin Treatment Failure in Organ Transplant Recipients. Gastroenterology 2014, 147, 1008–1011.e7. [Google Scholar] [CrossRef]
- Todt, D.; Gisa, A.; Radonic, A.; Nitsche, A.; Behrendt, P.; Suneetha, P.V.; Pischke, S.; Bremer, B.; Brown, R.J.P.; Manns, M.P.; et al. In Vivo Evidence for Ribavirin-Induced Mutagenesis of the Hepatitis E Virus Genome. Gut 2016, 65, 1733–1743. [Google Scholar] [CrossRef] [Green Version]
- Kamar, N.; Abravanel, F.; Behrendt, P.; Hofmann, J.; Pageaux, G.P.; Barbet, C.; Moal, V.; Couzi, L.; Horvatits, T.; De Man, R.A.; et al. Ribavirin for Hepatitis E Virus Infection After Organ Transplantation: A Large European Retrospective Multicenter Study. Clin. Infect. Dis. 2020, 71, 1204–1211. [Google Scholar] [CrossRef]
- Debing, Y.; Ramière, C.; Dallmeier, K.; Piorkowski, G.; Trabaud, M.-A.; Lebossé, F.; Scholtès, C.; Roche, M.; Legras-Lachuer, C.; de Lamballerie, X.; et al. Hepatitis E Virus Mutations Associated with Ribavirin Treatment Failure Result in Altered Viral Fitness and Ribavirin Sensitivity. J. Hepatol. 2016, 65, 499–508. [Google Scholar] [CrossRef]
- Debing, Y.; Moradpour, D.; Neyts, J.; Gouttenoire, J. Update on Hepatitis E Virology:Implications for Clinical Practice. J. Hepatol. 2016. [Google Scholar] [CrossRef] [Green Version]
- Meister, T.L.; Bruening, J.; Todt, D.; Steinmann, E. Cell Culture Systems for the Study of Hepatitis E Virus. Antivir. Res. 2019, 163, 34–49. [Google Scholar] [CrossRef]
- Fu, R.M.; Decker, C.C.; Dao Thi, V.L. Cell Culture Models for Hepatitis E Virus. Viruses 2019, 11, 608. [Google Scholar] [CrossRef] [Green Version]
- Marion, O.; Lhomme, S.; Nayrac, M.; Dubois, M.; Pucelle, M.; Requena, M.; Migueres, M.; Abravanel, F.; Peron, J.M.; Carrere, N.; et al. Hepatitis E Virus Replication in Human Intestinal Cells. Gut 2020, 69, 901–910. [Google Scholar] [CrossRef]
- Knegendorf, L.; Drave, S.A.; Dao Thi, V.L.; Debing, Y.; Brown, R.J.P.; Vondran, F.W.R.; Resner, K.; Friesland, M.; Khera, T.; Engelmann, M.; et al. Hepatitis E Virus Replication and Interferon Responses in Human Placental Cells. Hepatol. Commun. 2018, 2, 173–187. [Google Scholar] [CrossRef] [Green Version]
- Capelli, N.; Dubois, M.; Pucelle, M.; Da Silva, I.; Lhomme, S.; Abravanel, F.; Chapuy-Regaud, S.; Izopet, J. Optimized Hepatitis E Virus (HEV) Culture and Its Application to Measurements of HEV Infectivity. Viruses 2020, 12, 139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schemmerer, M.; Johne, R.; Erl, M.; Jilg, W.; Wenzel, J.J. Isolation of Subtype 3c, 3e and 3f-Like Hepatitis E Virus Strains Stably Replicating to High Viral Loads in an Optimized Cell Culture System. Viruses 2019, 11, 483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Todt, D.; Friesland, M.; Moeller, N.; Praditya, D.; Kinast, V.; Brüggemann, Y.; Knegendorf, L.; Burkard, T.; Steinmann, J.; Burm, R.; et al. Robust Hepatitis E Virus Infection and Transcriptional Response in Human Hepatocytes. Proc. Natl. Acad. Sci. USA 2020, 117, 1731–1741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yadav, K.K.; Boley, P.A.; Fritts, Z.; Kenney, S.P. Ectopic Expression of Genotype 1 Hepatitis E Virus ORF4 Increases Genotype 3 HEV Viral Replication in Cell Culture. Viruses 2021, 13, 75. [Google Scholar] [CrossRef]
- Rogée, S.; Talbot, N.; Caperna, T.; Bouquet, J.; Barnaud, E.; Pavio, N. New Models of Hepatitis E Virus Replication in Human and Porcine Hepatocyte Cell Lines. J. Gen. Virol. 2013, 94, 549–558. [Google Scholar] [CrossRef]
- Luangsay, S.; Ait-Goughoulte, M.; Michelet, M.; Floriot, O.; Bonnin, M.; Gruffaz, M.; Rivoire, M.; Fletcher, S.; Javanbakht, H.; Lucifora, J.; et al. Expression and Functionality of Toll- and RIG-like Receptors in HepaRG Cells. J. Hepatol. 2015, 63, 1077–1085. [Google Scholar] [CrossRef] [PubMed]
- Bouquet, J.; Cheval, J.; Rogée, S.; Pavio, N.; Eloit, M. Identical Consensus Sequence and Conserved Genomic Polymorphism of Hepatitis E Virus during Controlled Interspecies Transmission. J. Virol. 2012, 86, 6238–6245. [Google Scholar] [CrossRef] [Green Version]
- Salines, M.; Andraud, M.; Pellerin, M.; Bernard, C.; Grasland, B.; Pavio, N.; Rose, N. Impact of Porcine Circovirus Type 2 (PCV2) Infection on Hepatitis E Virus (HEV) Infection and Transmission under Experimental Conditions. Vet. Microbiol. 2019, 234, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Barnaud, E.; Rogée, S.; Garry, P.; Rose, N.; Pavio, N. Thermal Inactivation of Infectious Hepatitis E Virus in Experimentally Contaminated Food. Appl. Environ. Microbiol. 2012, 78, 5153–5159. [Google Scholar] [CrossRef] [Green Version]
- Jothikumar, N.; Cromeans, T.L.; Robertson, B.H.; Meng, X.J.; Hill, V.R. A Broadly Reactive One-Step Real-Time RT-PCR Assay for Rapid and Sensitive Detection of Hepatitis E Virus. J. Virol Methods 2006, 131, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Garson, J.A.; Ferns, R.B.; Grant, P.R.; Ijaz, S.; Nastouli, E.; Szypulska, R.; Tedder, R.S. Minor Groove Binder Modification of Widely Used TaqMan Probe for Hepatitis E Virus Reduces Risk of False Negative Real-Time PCR Results. J. Virol. Methods 2012, 186, 157–160. [Google Scholar] [CrossRef] [PubMed]
- Salines, M.; Barnaud, E.; Andraud, M.; Eono, F.; Renson, P.; Bourry, O.; Pavio, N.; Rose, N. Hepatitis E Virus Chronic Infection of Swine Co-Infected with Porcine Reproductive and Respiratory Syndrome Virus. Vet. Res. 2015, 46, 55. [Google Scholar] [CrossRef] [Green Version]
- Anderson, D.A.; Li, F.; Riddell, M.; Howard, T.; Seow, H.-F.; Torresi, J.; Perry, G.; Sumarsidi, D.; Shrestha, S.M.; Shrestha, I.L. ELISA for IgG-Class Antibody to Hepatitis E Virus Based on a Highly Conserved, Conformational Epitope Expressed in Eschericia Coli. J. Virol. Methods 1999, 81, 131–142. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A Flexible Trimmer for Illumina Sequence Data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langmead, B.; Salzberg, S.L. Fast Gapped-Read Alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Durbin, R. Fast and Accurate Short Read Alignment with Burrows-Wheeler Transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef] [Green Version]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A New Genome Assembly Algorithm and Its Applications to Single-Cell Sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Ye, W.; Zhang, Y.; Xu, Y. High Speed BLASTN: An Accelerated MegaBLAST Search Tool. Nucleic Acids Res. 2015, 43, 7762–7768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koboldt, D.C.; Chen, K.; Wylie, T.; Larson, D.E.; McLellan, M.D.; Mardis, E.R.; Weinstock, G.M.; Wilson, R.K.; Ding, L. VarScan: Variant Detection in Massively Parallel Sequencing of Individual and Pooled Samples. Bioinformatics 2009, 25, 2283–2285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, D.B.; Izopet, J.; Nicot, F.; Simmonds, P.; Jameel, S.; Meng, X.-J.; Norder, H.; Okamoto, H.; van der Poel, W.H.M.; Reuter, G.; et al. Update: Proposed Reference Sequences for Subtypes of Hepatitis E Virus (Species Orthohepevirus A). J. Gen. Virol. 2020, 101, 692–698. [Google Scholar] [CrossRef] [PubMed]
- Gripon, P.; Rumin, S.; Urban, S.; Le Seyec, J.; Glaise, D.; Cannie, I.; Guyomard, C.; Lucas, J.; Trepo, C.; Guguen-Guillouzo, C. Infection of a Human Hepatoma Cell Line by Hepatitis B Virus. Proc. Natl. Acad. Sci. USA 2002, 99, 15655–15660. [Google Scholar] [CrossRef] [Green Version]
- Debing, Y.; Emerson, S.U.; Wang, Y.; Pan, Q.; Balzarini, J.; Dallmeier, K.; Neyts, J. Ribavirin Inhibits in Vitro Hepatitis E Virus Replication through Depletion of Cellular GTP Pools and Is Moderately Synergistic with Alpha Interferon. Antimicrob. Agents Chemother. 2014, 58, 267–273. [Google Scholar] [CrossRef] [Green Version]
- Todt, D.; François, C.; Anggakusuma, N.; Behrendt, P.; Engelmann, M.; Knegendorf, L.; Vieyres, G.; Wedemeyer, H.; Hartmann, R.; Pietschmann, T.; et al. Antiviral Activities of Different Interferon Types and Subtypes against Hepatitis E Virus Replication. Antimicrob. Agents Chemother. 2016, 60, 2132–2139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lhomme, S.; Abravanel, F.; Dubois, M.; Sandres-Saune, K.; Mansuy, J.-M.; Rostaing, L.; Kamar, N.; Izopet, J. Characterization of the Polyproline Region of the Hepatitis E Virus in Immunocompromised Patients. J. Virol. 2014, 88, 12017–12025. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nicot, F.; Jeanne, N.; Roulet, A.; Lefebvre, C.; Carcenac, R.; Manno, M.; Dubois, M.; Kamar, N.; Lhomme, S.; Abravanel, F.; et al. Diversity of Hepatitis E Virus Genotype 3. Rev. Med. Virol. 2018, 28, e1987. [Google Scholar] [CrossRef] [PubMed]
- MacNab, G.M.; Alexander, J.J.; Lecatsas, G.; Bey, E.M.; Urbanowicz, J.M. Hepatitis B Surface Antigen Produced by a Human Hepatoma Cell Line. Br. J. Cancer 1976, 34, 509–515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marozin, S.; Altomonte, J.; Stadler, F.; Thasler, W.E.; Schmid, R.M.; Ebert, O. Inhibition of the IFN-β Response in Hepatocellular Carcinoma by Alternative Spliced Isoform of IFN Regulatory Factor-3. Mol. Ther. 2008, 16, 1789–1797. [Google Scholar] [CrossRef]
- Yin, X.; Li, X.; Ambardekar, C.; Hu, Z.; Lhomme, S.; Feng, Z. Hepatitis E Virus Persists in the Presence of a Type III Interferon Response. PLoS Pathog. 2017, 13, e1006417. [Google Scholar] [CrossRef] [Green Version]
- Oshiro, Y.; Yasue, H.; Takahashi, K.; Hattori, S.; Ideno, S.; Urayama, T.; Chiba, M.; Osari, S.; Naito, T.; Takeuchi, K.; et al. Mode of Swine Hepatitis E Virus Infection and Replication in Primary Human Hepatocytes. J. Gen. Virol. 2014, 95, 2677–2682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, X.; Dao Thi, V.L.; Liu, P.; Takacs, C.N.; Xiang, K.; Andrus, L.; Gouttenoire, J.; Moradpour, D.; Rice, C.M. Pan-Genotype Hepatitis E Virus Replication in Stem Cell–Derived Hepatocellular Systems. Gastroenterology 2018, 154, 663–674.e7. [Google Scholar] [CrossRef] [PubMed]
- Helsen, N.; Debing, Y.; Paeshuyse, J.; Dallmeier, K.; Boon, R.; Coll, M.; Sancho-Bru, P.; Claes, C.; Neyts, J.; Verfaillie, C.M. Stem Cell-Derived Hepatocytes: A Novel Model for Hepatitis E Virus Replication. J. Hepatol. 2016, 64, 565–573. [Google Scholar] [CrossRef] [Green Version]
- Dao Thi, V.L.; Debing, Y.; Wu, X.; Rice, C.M.; Neyts, J.; Moradpour, D.; Gouttenoire, J. Sofosbuvir Inhibits Hepatitis E Virus Replication in Vitro and Results in an Additive Effect When Combined with Ribavirin. Gastroenterology 2015. [Google Scholar] [CrossRef] [Green Version]
- Parent, R.; Marion, M.-J.; Furio, L.; Trépo, C.; Petit, M.-A. Origin and Characterization of a Human Bipotent Liver Progenitor Cell Line. Gastroenterology 2004, 126, 1147–1156. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Wang, W.; Li, Y.; Zhou, X.; Yin, Y.; Wang, Y.; de Man, R.A.; van der Laan, L.J.W.; Huang, F.; Kamar, N.; et al. RIG-I Is a Key Antiviral Interferon-Stimulated Gene against Hepatitis E Virus Regardless of Interferon Production. Hepatology 2017, 65, 1823–1839. [Google Scholar] [CrossRef] [Green Version]
- Xu, L.; Zhou, X.; Wang, W.; Wang, Y.; Yin, Y.; Laan, L.J.W.; van der Sprengers, D.; Metselaar, H.J.; Peppelenbosch, M.P.; Pan, Q. IFN Regulatory Factor 1 Restricts Hepatitis E Virus Replication by Activating STAT1 to Induce Antiviral IFN-Stimulated Genes. Faseb J. 2016, 30, 3352–3367. [Google Scholar] [CrossRef] [Green Version]
- Lucifora, J.; Xia, Y.; Reisinger, F.; Zhang, K.; Stadler, D.; Cheng, X.; Sprinzl, M.F.; Koppensteiner, H.; Makowska, Z.; Volz, T.; et al. Specific and Nonhepatotoxic Degradation of Nuclear Hepatitis B Virus CccDNA. Science 2014, 343, 1221–1228. [Google Scholar] [CrossRef]
- Ndongo-Thiam, N.; Berthillon, P.; Errazuriz, E.; Bordes, I.; De Sequeira, S.; Trépo, C.; Petit, M.-A. Long-Term Propagation of Serum Hepatitis C Virus (HCV) with Production of Enveloped HCV Particles in Human HepaRG Hepatocytes. Hepatology 2011, 54, 406–417. [Google Scholar] [CrossRef]
- Leite, S.B.; Wilk-Zasadna, I.; Zaldivar, J.M.; Airola, E.; Reis-Fernandes, M.A.; Mennecozzi, M.; Guguen-Guillouzo, C.; Chesne, C.; Guillou, C.; Alves, P.M.; et al. Three-Dimensional HepaRG Model As An Attractive Tool for Toxicity Testing. Toxicol. Sci. 2012, 130, 106–116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ooka, M.; Lynch, C.; Xia, M. Application of In Vitro Metabolism Activation in High-Throughput Screening. Int. J. Mol. Sci. 2020, 21, 8182. [Google Scholar] [CrossRef] [PubMed]
- Tolosa, L.; Gómez-Lechón, M.J.; Jiménez, N.; Hervás, D.; Jover, R.; Donato, M.T. Advantageous Use of HepaRG Cells for the Screening and Mechanistic Study of Drug-Induced Steatosis. Toxicol. Appl. Pharm. 2016, 302, 1–9. [Google Scholar] [CrossRef]
- Kanebratt, K.P.; Andersson, T.B. HepaRG Cells as an in Vitro Model for Evaluation of Cytochrome P450 Induction in Humans. Drug. Metab. Dispos. 2008, 36, 137–145. [Google Scholar] [CrossRef] [Green Version]
- Todt, D.; Moeller, N.; Praditya, D.; Kinast, V.; Friesland, M.; Engelmann, M.; Verhoye, L.; Sayed, I.M.; Behrendt, P.; Dao Thi, V.L.; et al. The Natural Compound Silvestrol Inhibits Hepatitis E Virus (HEV) Replication in Vitro and in Vivo. Antivir. Res. 2018, 157, 151–158. [Google Scholar] [CrossRef]
- Nishiyama, T.; Kobayashi, T.; Jirintai, S.; Kii, I.; Nagashima, S.; Prathiwi Primadharsini, P.; Nishizawa, T.; Okamoto, H. Screening of Novel Drugs for Inhibiting Hepatitis E Virus Replication. J. Virol. Methods 2019, 270, 1–11. [Google Scholar] [CrossRef]
- Purdy, M.A.; Lara, J.; Khudyakov, Y.E. The Hepatitis E Virus Polyproline Region Is Involved in Viral Adaptation. PLoS ONE 2012, 7, e35974. [Google Scholar] [CrossRef] [Green Version]
- Smith, D.B.; Vanek, J.; Ramalingam, S.; Johannessen, I.; Templeton, K.; Simmonds, P. Evolution of the Hepatitis E Virus Hypervariable Region. J. Gen. Virol. 2012, 93, 2408–2418. [Google Scholar] [CrossRef] [Green Version]
- Pudupakam, R.S.; Huang, Y.W.; Opriessnig, T.; Halbur, P.G.; Pierson, F.W.; Meng, X.J. Deletions of the Hypervariable Region (HVR) in Open Reading Frame 1 of Hepatitis E Virus Do Not Abolish Virus Infectivity: Evidence for Attenuation of HVR Deletion Mutants in Vivo. J. Virol. 2009, 83, 384–395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pudupakam, R.S.; Kenney, S.P.; Córdoba, L.; Huang, Y.-W.; Dryman, B.A.; Leroith, T.; Pierson, F.W.; Meng, X.-J. Mutational Analysis of the Hypervariable Region of Hepatitis e Virus Reveals Its Involvement in the Efficiency of Viral RNA Replication. J. Virol. 2011, 85, 10031–10040. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bagdassarian, E.; Doceul, V.; Pellerin, M.; Demange, A.; Meyer, L.; Jouvenet, N.; Pavio, N. The Amino-Terminal Region of Hepatitis E Virus ORF1 Containing a Methyltransferase (Met) and a Papain-Like Cysteine Protease (PCP) Domain Counteracts Type I Interferon Response. Viruses 2018, 10, 726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nan, Y.; Yu, Y.; Ma, Z.; Khattar, S.K.; Fredericksen, B.; Zhang, Y.-J. Hepatitis E Virus Inhibits Type I Interferon Induction by ORF1 Products. J. Virol. 2014, 88, 11924–11932. [Google Scholar] [CrossRef] [Green Version]
- Kim, E.; Myoung, J. Hepatitis E Virus Papain-Like Cysteine Protease Inhibits Type I Interferon Induction by Down-Regulating Melanoma Differentiation-Associated Gene 5. J. Microbiol. Biotechnol. 2018, 28, 1908–1915. [Google Scholar] [CrossRef] [Green Version]
- Karpe, Y.A.; Lole, K.S. Deubiquitination Activity Associated with Hepatitis E Virus Putative Papain-like Cysteine Protease. J. Gen. Virol. 2011, 92, 2088–2092. [Google Scholar] [CrossRef]
- Kanade, G.D.; Pingale, K.D.; Karpe, Y.A. Activities of Thrombin and Factor Xa Are Essential for Replication of Hepatitis E Virus and Are Possibly Implicated in ORF1 Polyprotein Processing. J. Virol. 2018, 92. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, H.T.; Torian, U.; Faulk, K.; Mather, K.; Engle, R.E.; Thompson, E.; Bonkovsky, H.L.; Emerson, S.U. A Naturally Occurring Human/Hepatitis E Recombinant Virus Predominates in Serum but Not in Faeces of a Chronic Hepatitis E Patient and Has a Growth Advantage in Cell Culture. J. Gen. Virol. 2012, 93, 526–530. [Google Scholar] [CrossRef] [PubMed]
- Shukla, P.; Nguyen, H.T.; Torian, U.; Engle, R.E.; Faulk, K.; Dalton, H.R.; Bendall, R.P.; Keane, F.E.; Purcell, R.H.; Emerson, S.U. Cross-Species Infections of Cultured Cells by Hepatitis E Virus and Discovery of an Infectious Virus-Host Recombinant. Proc. Natl. Acad. Sci. USA 2011, 108, 2438–2443. [Google Scholar] [CrossRef] [Green Version]
- Shukla, P.; Nguyen, H.T.; Faulk, K.; Mather, K.; Torian, U.; Engle, R.E.; Emerson, S.U. Adaptation of a Genotype 3 Hepatitis E Virus to Efficient Growth in Cell Culture Depends on an Inserted Human Gene Segment Acquired by Recombination. J. Virol. 2012, 86, 5697–5707. [Google Scholar] [CrossRef] [Green Version]
- Johne, R.; Reetz, J.; Ulrich, R.G.; Machnowska, P.; Sachsenröder, J.; Nickel, P.; Hofmann, J. An ORF1-Rearranged Hepatitis E Virus Derived from a Chronically Infected Patient Efficiently Replicates in Cell Culture. J. Viral Hepat. 2014, 21, 447–456. [Google Scholar] [CrossRef] [PubMed]
- Lorenzo, F.R.; Tanaka, T.; Takahashi, H.; Ichiyama, K.; Hoshino, Y.; Yamada, K.; Inoue, J.; Takahashi, M.; Okamoto, H. Mutational Events during the Primary Propagation and Consecutive Passages of Hepatitis E Virus Strain JE03–1760F in Cell Culture. Virus Res. 2008, 137, 86–96. [Google Scholar] [CrossRef]
- Sari, G.; van de Garde, M.D.B.; van Schoonhoven, A.; Voermans, J.J.C.; van der Eijk, A.A.; de Man, R.A.; Boonstra, A.; Vanwolleghem, T.; Pas, S.D. Hepatitis E Virus Shows More Genomic Alterations in Cell Culture than In Vivo. Pathogens 2019, 8, 255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanaka, T.; Takahashi, M.; Takahashi, H.; Ichiyama, K.; Hoshino, Y.; Nagashima, S.; Mizuo, H.; Okamoto, H. Development and Characterization of a Genotype 4 Hepatitis E Virus Cell Culture System Using a HE-JF5/15F Strain Recovered from a Fulminant Hepatitis Patient. J. Clin. Microbiol. 2009, 47, 1906–1910. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Passage # | Inoculum | MOI (GE/Cell) | Inoculum Added (GE/Well) | Infection Length | Viral Load Around D+21 (GE/mL Supernatant) | Viral Load Around D+100 (GE/mL Supernatant) | Viral Load at the Last Time Point (GE/mL Supernatant) |
|---|---|---|---|---|---|---|---|
| 1 | Human faecal sample | 10 | 2.1 × 107 | D+95 | D+22: 1.55 × 105 | D+95: 2.02 × 108 | D+95: 2.02 × 108 |
| 2 | Supernatant 1st passage (D+91) | 25 | 7.5 × 107 | D+382 | D+20: 1.41 × 106 | D+95: 2.75 × 108 | D+382: 4.93 × 108 |
| 3 | Supernatant 2nd passage (D+84) | 21 | 1 × 108 | D+321 | D+20: 2.06 × 106 | D+95: 6.23 × 108 | D+321: 1.35 × 108 |
| 4 | Supernatant 3rd passage (D+41) | 8 | 1.2 × 107 | D+280 | D+21: 1.25 × 106 | D+96: 7.59 × 108 | D+280: 7.93 × 107 |
| 5 | Supernatant 4th passage (D+133) | 10 | 3.2 × 107 | D+147 | D+19: 6.72 × 105 | D+92: 1.18 × 109 | D+147: 8.73 × 107 |
| 6 | Supernatant 5th passage (D+126) | 10 | 4 × 107 | D+328 | D+33: 5.59 × 106 | D+96: 5.87 × 108 | D+328: 1.77 × 108 |
| 7 | Supernatant 6th passage (D+98) | 10 | 3.8 × 107 | D+230 | D+23: 5.95 × 106 | D+82: 1.04 × 109 | D+230: 1.97 × 108 |
| ORF | Nucleotide Positions | Amino Acid Substitutions | Motif |
|---|---|---|---|
| ORF1 | G 538 T | ||
| T 628 C | |||
| T 925 C | |||
| T 1069 C | |||
| C 1114 T | |||
| G 1499 C | G-493-R | Papain-like cysteine protease (PCP) | |
| G 1534 A | |||
| C 1794 T | A-591-V | PCP | |
| C 1821 T | A-600-V | Un | |
| C 2033 T | H-671-Y | Un | |
| C 2197 T | |||
| T 2246 C | W-742-R | HVR | |
| G 2344 A | |||
| T 2494 C | |||
| C 2630 T | H-870-Y | X | |
| T 2674 C | |||
| A 3974 G | T-1318-A | RNA-dependent RNA polymerase (RdRp) | |
| C 4255 T | |||
| C 4621 T | |||
| T 4837 C | |||
| T 4918 C | |||
| ORF2 | C 6566 T | ||
| T 6740 C | |||
| C 6756 T | L-500-F | ||
| C 7205 T |
| Isolate | Origin (Patient) | Cell Line | Insertion | Point Mutations (Non-Synonymous) | Position of Non-Synonymous Mutation | Reference | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Length | Nature | Position | Cell Culture Adaptation | ORF1 | ORF2 | ORF3 | |||||
| LBPR-0379 (HEV-3a) | Liver transplant Chronic hepatitis E | HepG2/C3A | 117 nt 39 aa | Human ribosomal protein S19 | aa 741 | Selected during passage | Not determined | [76] | |||
| Kernow-C1 (HEV-3a) | HIV patient Chronic hepatitis E | HepG2/C3A | 174 nt 58 aa | Human ribosomal protein S17 | aa 750 | Selected during passage Insertion shown to enhance growth | 64 (16) + insertion of 58 aa in HEV ORF1 after 6 passages | G650A (un) V737A (HVR) N768S (HVR) I770V (HVR) L772S (HVR) P774S (HVR) W846S (X) N957S (X) Q1001H (Hel) F1459L (RdRp) | R2C V66I T483A M652S K653E | M69I | [77,78] |
| 47832 (HEV-3c) | Kidney transplant Chronic hepatitis E | A549 | 186 nt 62 aa | Duplication of adjacent HVR sequence (116 nt) and ORF1 3′ end (70 nt) | aa 750 | - | 25 (8) after 2 passages | F808S (HVR) F809S (HVR) A864V (HVR) D1201A (Hel) T1761I (RdRp) | V12M G73A | F108L | [79] |
| JE03-1706 (HEV-3b) | Acute hepatitis E | PLC/PRF/5 | None | 18 (5) after 10 passages | A189V (Met) W741A (HVR) A1143V (Hel) | N73S P99L | [80] | ||||
| HEV0069 (HEV-3c) | Heart transplant Chronic hepatitis E | A549 | None | 19 (7) after 7 passages | FL462L (PCP) W741R (HVR) | P68S, E270V Y532D D625V | N73S P99L | [81] | |||
| HE-JF5 (HEV-4c) | Fulminant hepatitis E | A549 PLC/PRF/5 | None | 10 (4) after 6 passages | F717L (HVR) V1617I (RdRp) | A565V K653E | [82] | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pellerin, M.; Hirchaud, E.; Blanchard, Y.; Pavio, N.; Doceul, V. Characterization of a Cell Culture System of Persistent Hepatitis E Virus Infection in the Human HepaRG Hepatic Cell Line. Viruses 2021, 13, 406. https://doi.org/10.3390/v13030406
Pellerin M, Hirchaud E, Blanchard Y, Pavio N, Doceul V. Characterization of a Cell Culture System of Persistent Hepatitis E Virus Infection in the Human HepaRG Hepatic Cell Line. Viruses. 2021; 13(3):406. https://doi.org/10.3390/v13030406
Chicago/Turabian StylePellerin, Marie, Edouard Hirchaud, Yannick Blanchard, Nicole Pavio, and Virginie Doceul. 2021. "Characterization of a Cell Culture System of Persistent Hepatitis E Virus Infection in the Human HepaRG Hepatic Cell Line" Viruses 13, no. 3: 406. https://doi.org/10.3390/v13030406