HIV-1 Transcription Inhibitor 1E7-03 Restores LPS-Induced Alteration of Lung Leukocytes’ Infiltration Dynamics and Resolves Inflammation in HIV Transgenic Mice


Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Reagents
2.3. Immunohistochemistry
2.4. Isolation of Lung and Intra-Peritoneal Macrophages
2.5. Generation of Mouse Lung Endothelial Cell Line
2.6. Trans-Endothelial Migration Assay
2.7. Real-Time Polymerase Chain Reaction (RT-PCR)
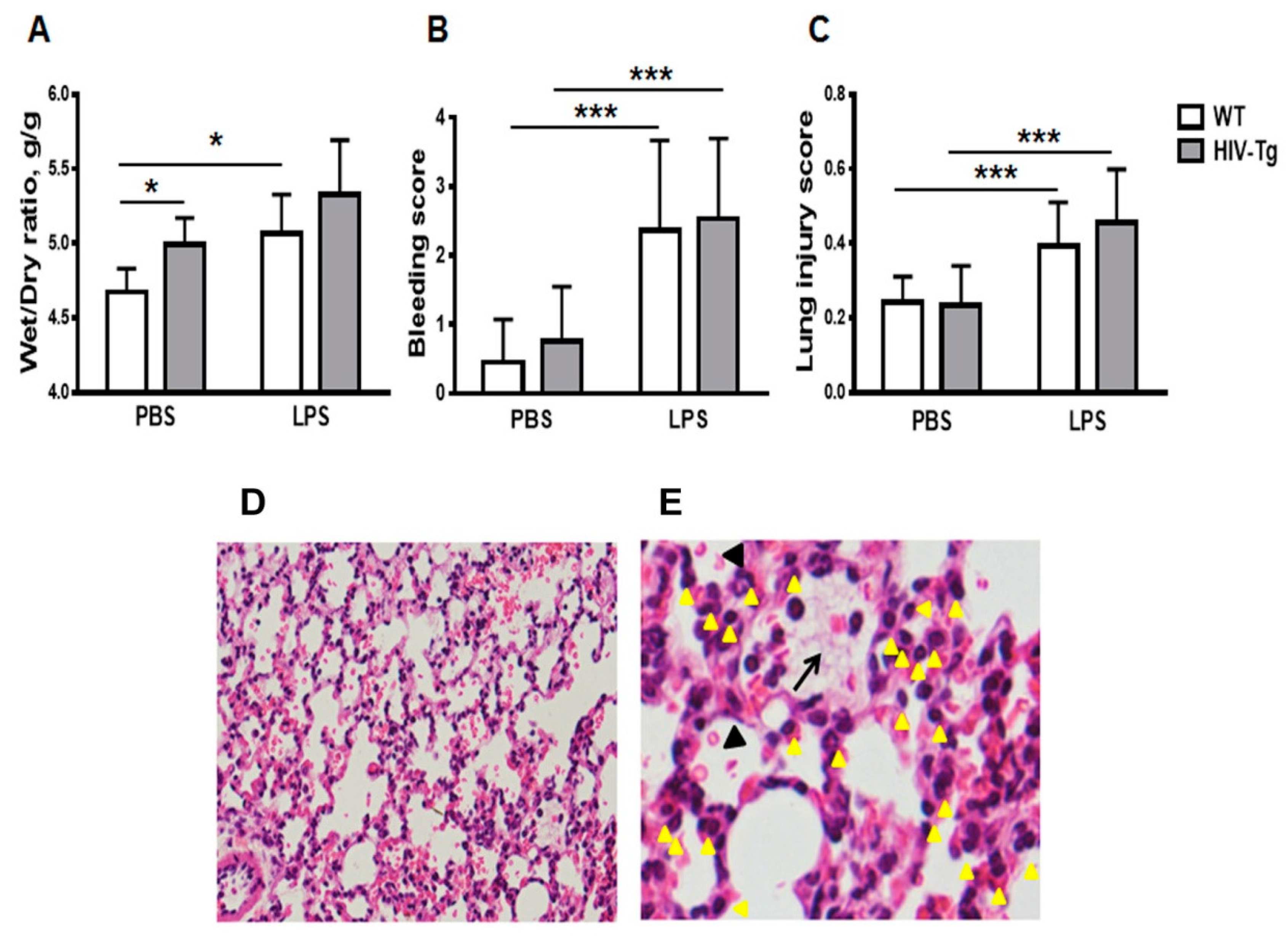
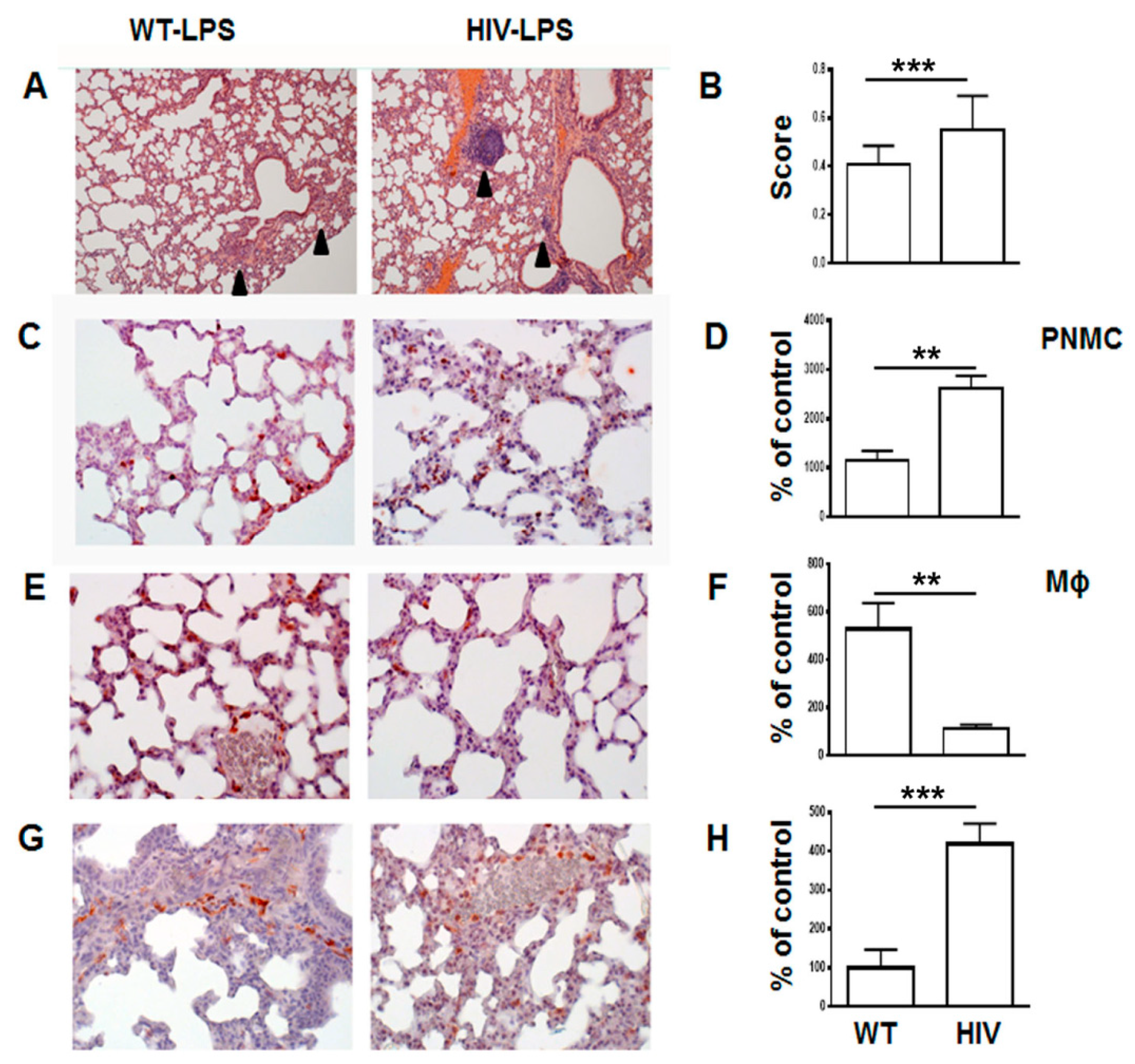
2.8. Lung Injury Scores
2.9. Bleeding Score
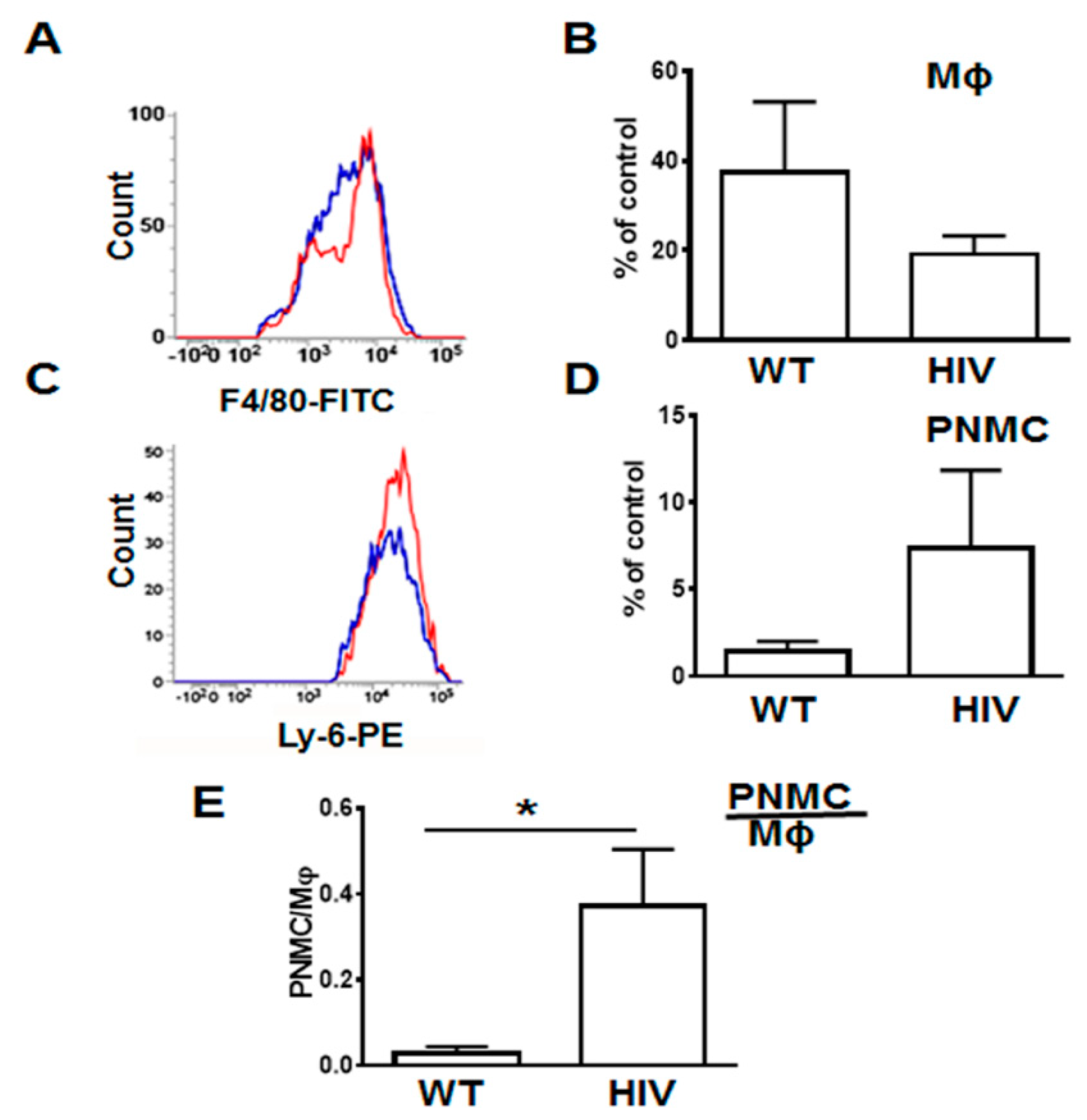
2.10. Flow Cytometry
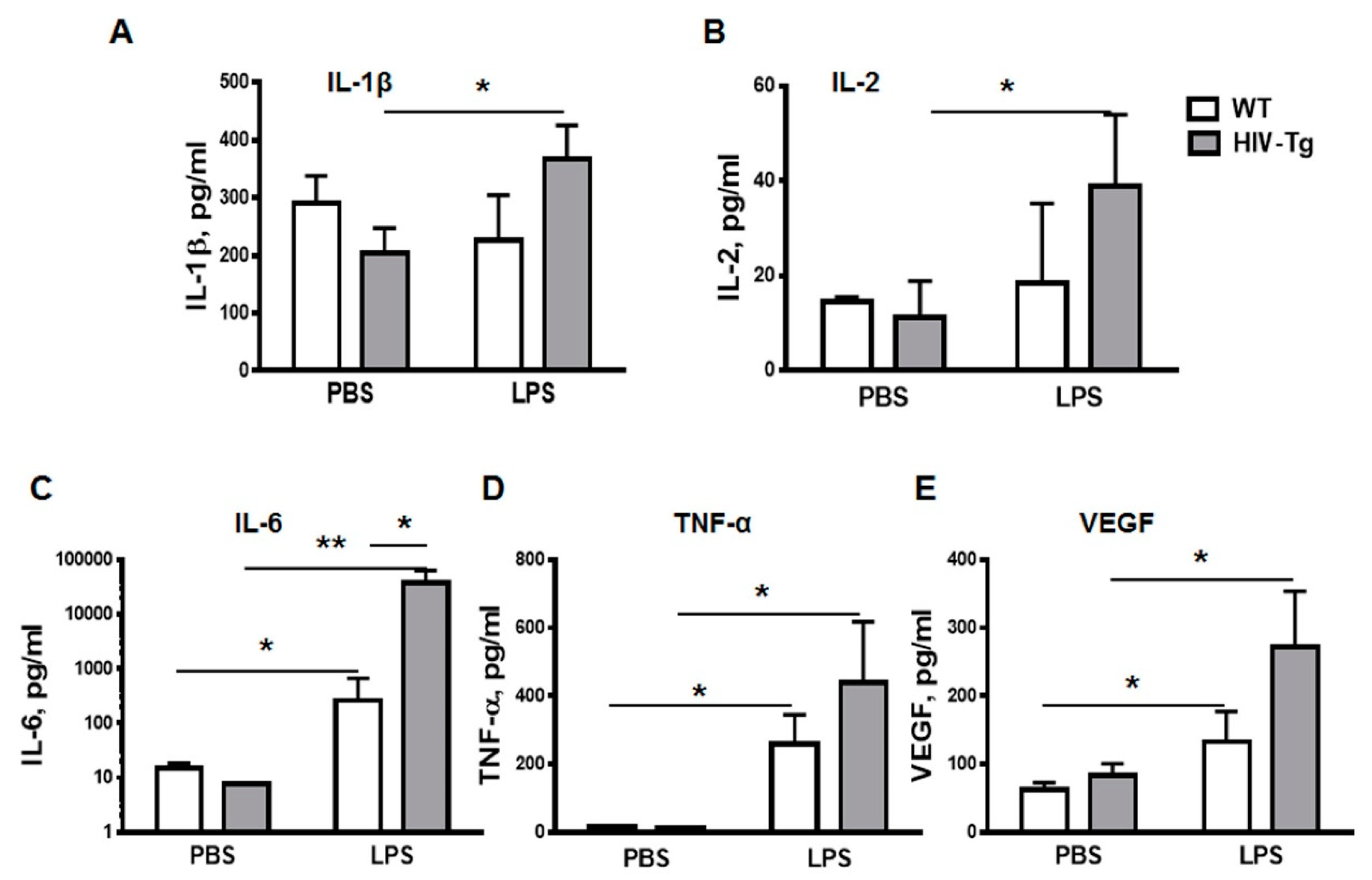
2.11. BioPlex Cytokine Analysis
2.12. Mouse VEGF Enzyme-Linked Immunosorbent Assay (ELISA)
2.13. Statistical Analysis
3. Results
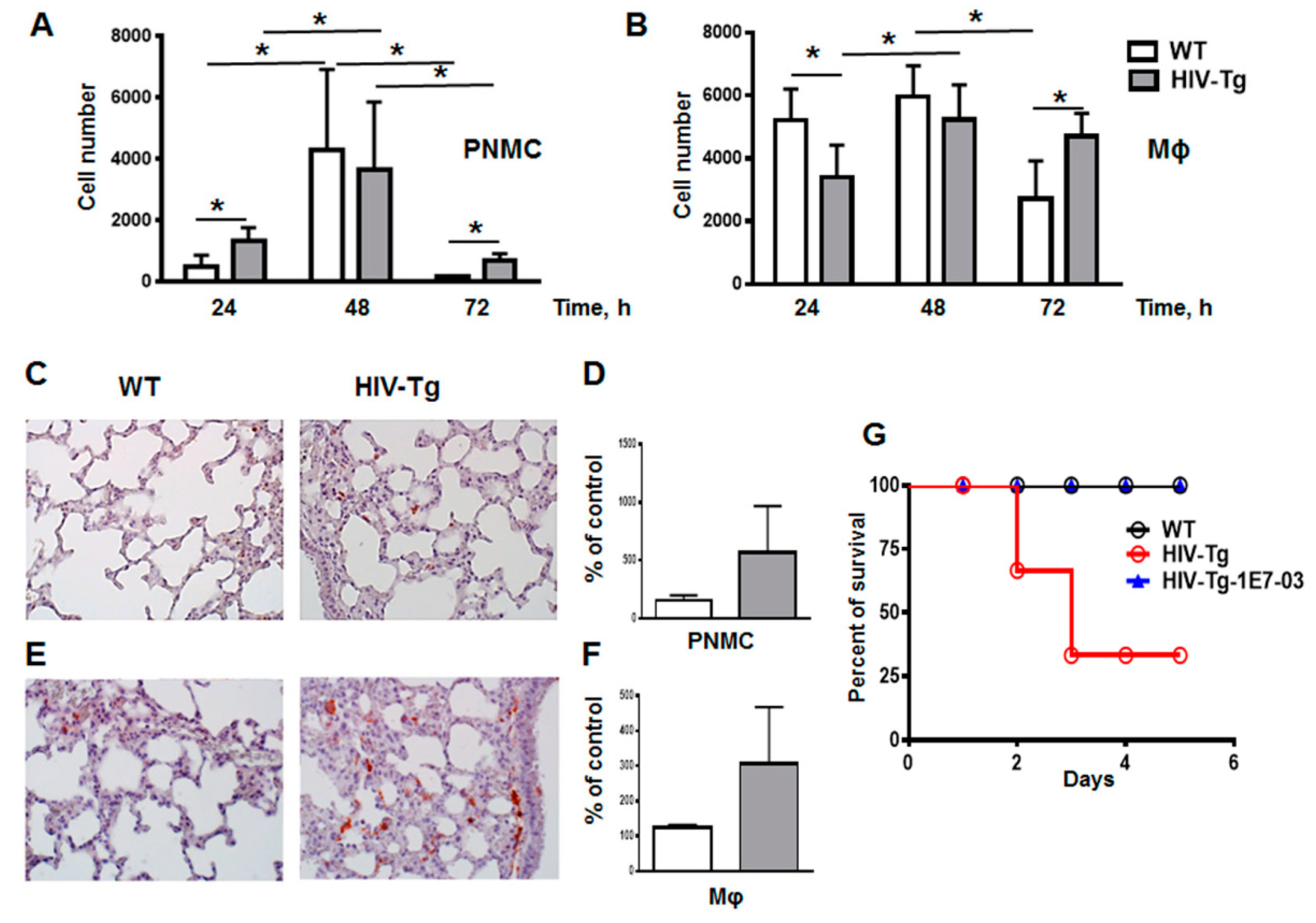
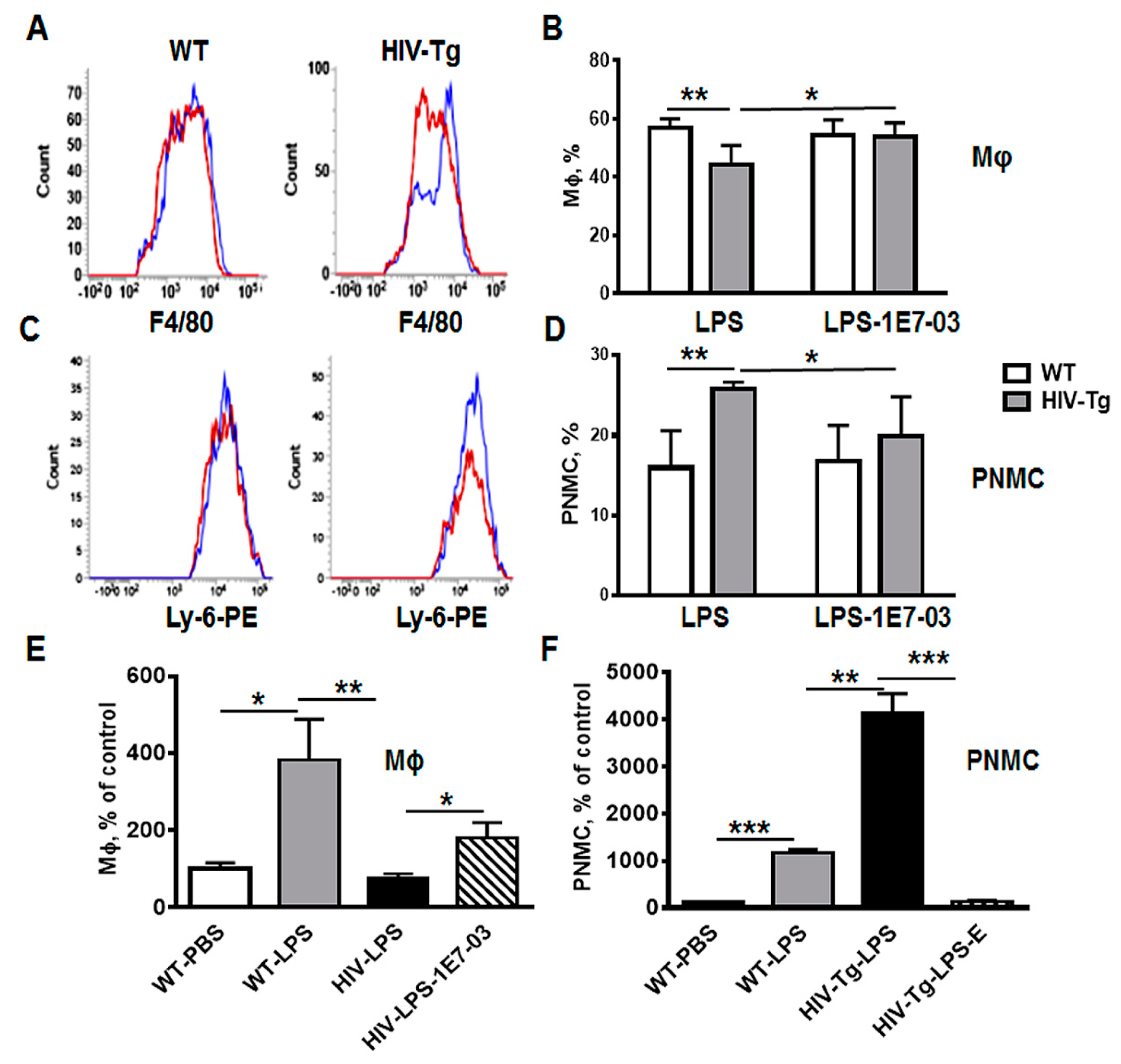
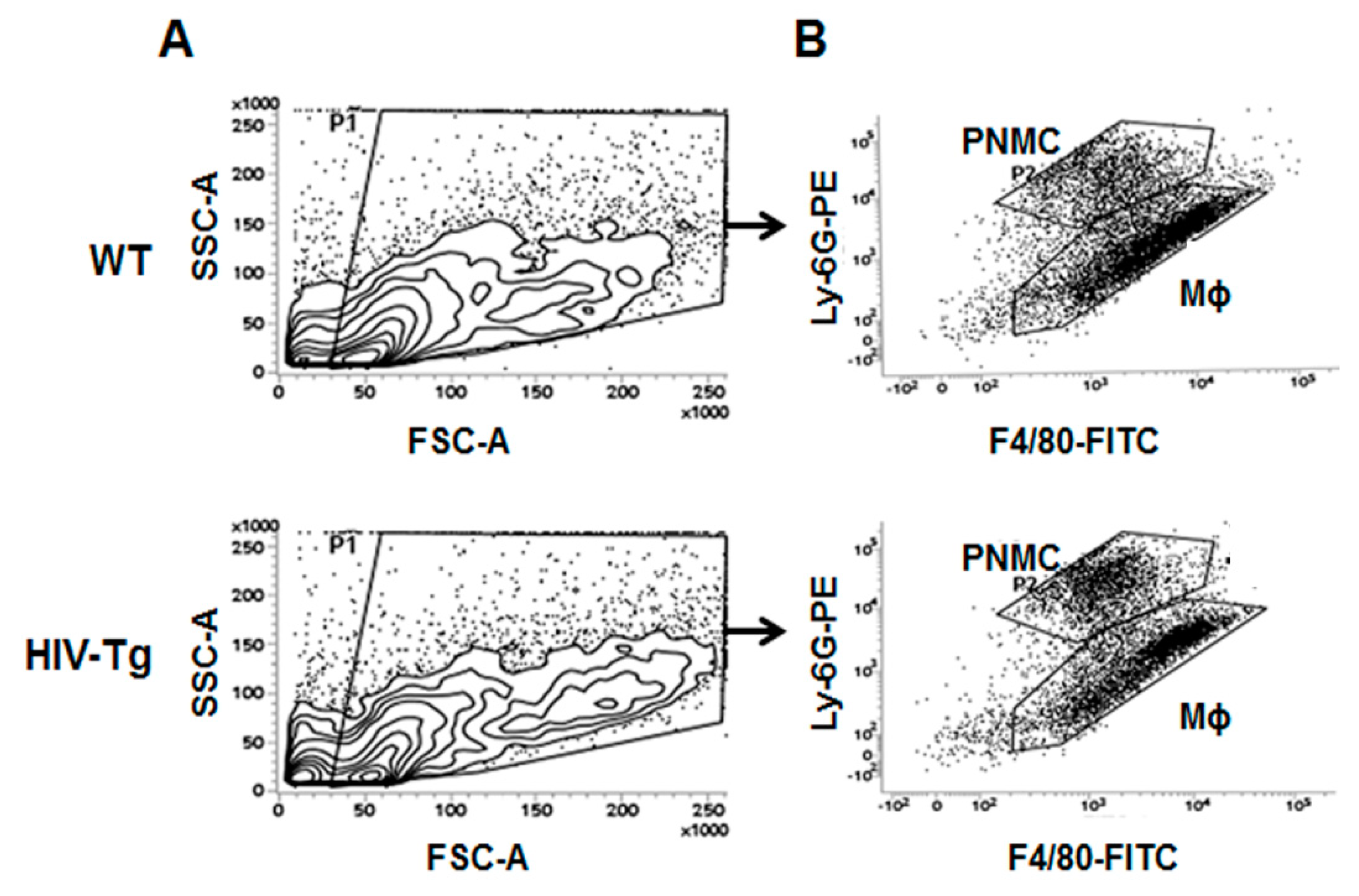
3.1. Increased Neutrophil Infiltration and Decreased Macrophage Infiltration in Lungs of Human Immunodeficiency Virus 1 Transgenic (HIV-Tg) Mice after LPS Administration
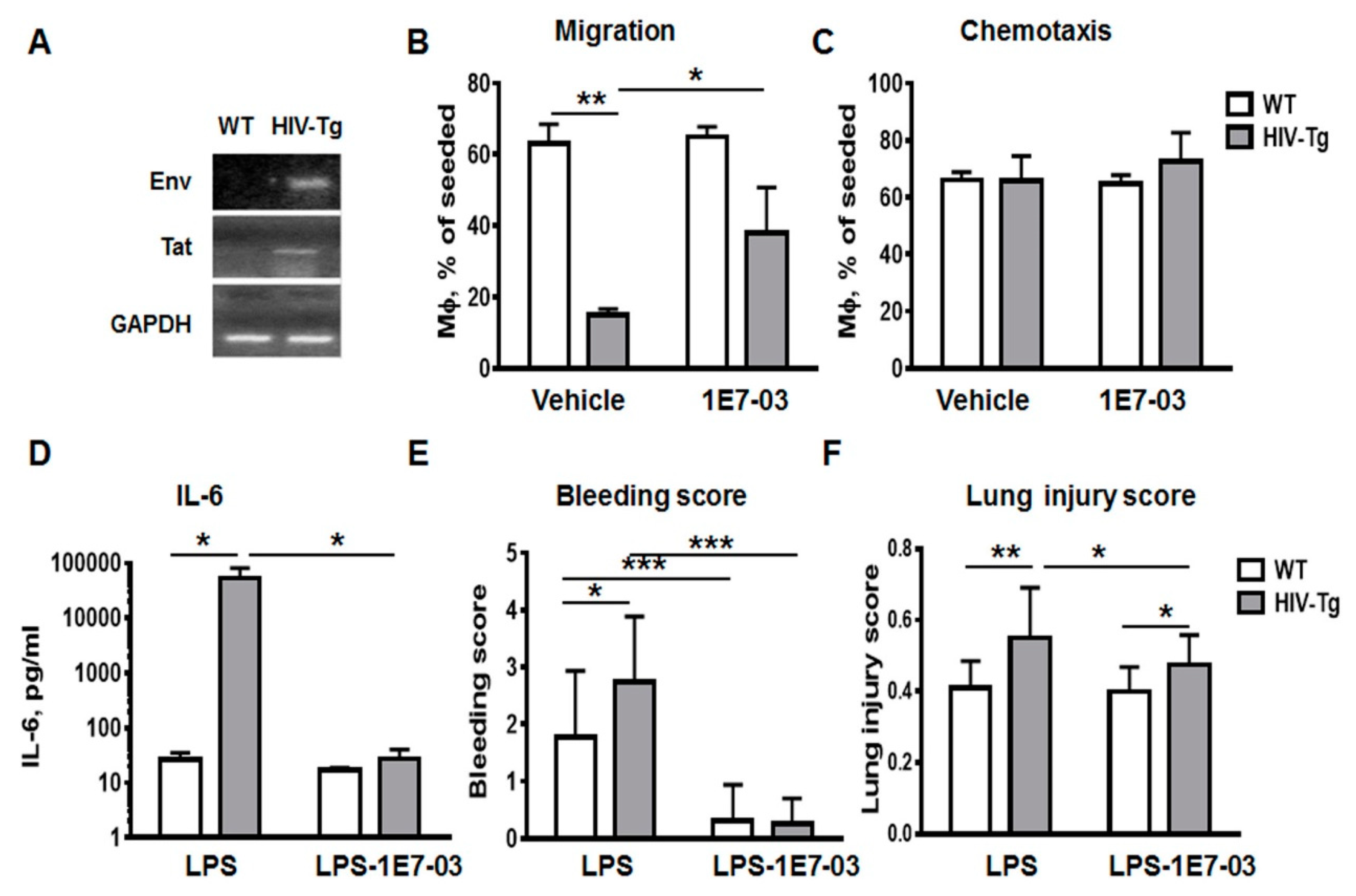
3.2. LPS Injections Induced Higher Levels of Inflammatory Cytokines in HIV-Tg Mice
3.3. Resolution of Lung Leukocytes’ Infiltration after LPS Administration Is Slower in HIV-Tg Mice
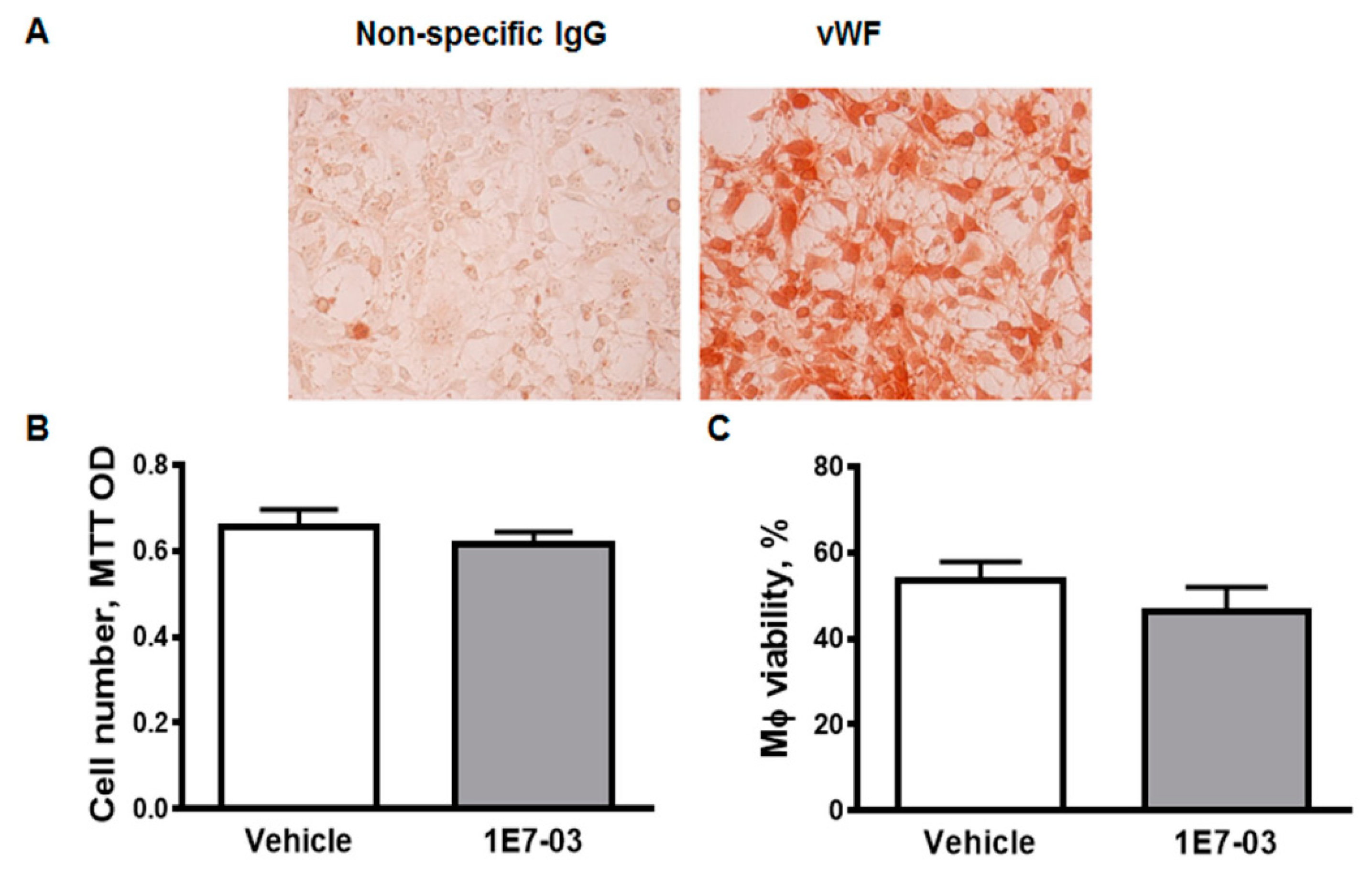
3.4. Trans-Endothelial Migration of Macrophages Isolated from HIV-Tg Mice Are Reduced In Vitro
3.5. Administration of HIV-1 Transcription Inhibitor 1E7-03 Improves Trans-Endothelial Macrophages Migration In Vitro and the Levels of Lung Leukocytes Infiltration in HIV-Tg Mice
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A



References
- Friis-Moller, N.; Sabin, C.A.; Weber, R.; d’Arminio Monforte, A.; El-Sadr, W.M.; Reiss, P.; Thiebaut, R.; Morfeldt, L.; de Wit, S.; Pradier, C.; et al. Data Collection on Adverse Events of Anti, H.I.V.D.S.G., Combination antiretroviral therapy and the risk of myocardial infarction. New Engl. J. Med. 2003, 349, 1993–2003. [Google Scholar]
- Wand, H.; Calmy, A.; Carey, D.L.; Samaras, K.; Carr, A.; Law, M.G.; Cooper, D.A.; Emery, S.; Committee, I.T.I.C. Metabolic syndrome, cardiovascular disease and type 2 diabetes mellitus after initiation of antiretroviral therapy in HIV infection. Aids 2007, 21, 2445–2453. [Google Scholar] [CrossRef] [PubMed]
- Calligaro, G.L.; Gray, D.M. Lung function abnormalities in HIV-infected adults and children. Respirology 2015, 20, 24–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morris, A.; George, M.P.; Crothers, K.; Huang, L.; Lucht, L.; Kessinger, C.; Kleerup, E.C.; Lung, H.I.V.S. HIV and chronic obstructive pulmonary disease: Is it worse and why? Proc. Am. Thorac. Soc. 2011, 8, 320–325. [Google Scholar] [CrossRef]
- Crothers, K.; Thompson, B.W.; Burkhardt, K.; Morris, A.; Flores, S.C.; Diaz, P.T.; Chaisson, R.E.; Kirk, G.D.; Rom, W.N.; Huang, L.; et al. HIV-associated lung infections and complications in the era of combination antiretroviral therapy. Proc. Am. Thorac. Soc. 2011, 8, 275–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agusti, A.; Edwards, L.D.; Rennard, S.I.; MacNee, W.; Tal-Singer, R.; Miller, B.E.; Vestbo, J.; Lomas, D.A.; Calverley, P.M.; Wouters, E.; et al. Persistent systemic inflammation is associated with poor clinical outcomes in COPD: A novel phenotype. PLoS ONE 2012, 7, e37483. [Google Scholar] [CrossRef]
- Decramer, M.; Janssens, W.; Miravitlles, M. Chronic obstructive pulmonary disease. Lancet 2012, 379, 1341–1351. [Google Scholar] [CrossRef]
- Diaz, P.T.; King, M.A.; Pacht, E.R.; Wewers, M.D.; Gadek, J.E.; Nagaraja, H.N.; Drake, J.; Clanton, T.L. Increased susceptibility to pulmonary emphysema among HIV-seropositive smokers. Ann. Intern. Med. 2000, 132, 369–372. [Google Scholar] [CrossRef]
- Pappas, K.; Papaioannou, A.I.; Kostikas, K.; Tzanakis, N. The role of macrophages in obstructive airways disease: Chronic obstructive pulmonary disease and asthma. Cytokine 2013, 64, 613–625. [Google Scholar] [CrossRef]
- Meijer, M.; Rijkers, G.T.; van Overveld, F.J. Neutrophils and emerging targets for treatment in chronic obstructive pulmonary disease. Expert Rev. Clin. Immunol. 2013, 9, 1055–1068. [Google Scholar] [CrossRef]
- Stockley, J.A.; Walton, G.M.; Lord, J.M.; Sapey, E. Aberrant neutrophil functions in stable chronic obstructive pulmonary disease: The neutrophil as an immunotherapeutic target. Int. Immunopharmacol. 2013, 17, 1211–1217. [Google Scholar] [CrossRef]
- Attia, E.F.; Akgun, K.M.; Wongtrakool, C.; Goetz, M.B.; Rodriguez-Barradas, M.C.; Rimland, D.; Brown, S.T.; Soo Hoo, G.W.; Kim, J.; Lee, P.J.; et al. Increased risk of radiographic emphysema in HIV is associated with elevated soluble CD14 and nadir CD4. Chest 2014, 146, 1543–1553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fitzpatrick, M.E.; Singh, V.; Bertolet, M.; Lucht, L.; Kessinger, C.; Michel, J.; Logar, A.; Weinman, R.; McMahon, D.; Norris, K.A.; et al. Relationships of pulmonary function, inflammation, and T-cell activation and senescence in an HIV-infected cohort. Aids 2014, 28, 2505–2515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, S.; Verma, S.K.; Kumar, S.; Ahmad, M.K.; Nischal, A.; Singh, S.K.; Dixit, R.K. Correlation of severity of chronic obstructive pulmonary disease with potential biomarkers. Immunol. Lett. 2018, 196, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Norris, K.A.; Morris, A.; Patil, S.; Fernandes, E. Pneumocystis colonization, airway inflammation, and pulmonary function decline in acquired immunodeficiency syndrome. Immunol. Res. 2006, 36, 175–187. [Google Scholar] [CrossRef]
- Almodovar, S. The complexity of HIV persistence and pathogenesis in the lung under antiretroviral therapy: Challenges beyond AIDS. Viral Immunol. 2014, 27, 186–199. [Google Scholar] [CrossRef] [Green Version]
- Itescu, S.; Simonelli, P.F.; Winchester, R.J.; Ginsberg, H.S. Human immunodeficiency virus type 1 strains in the lungs of infected individuals evolve independently from those in peripheral blood and are highly conserved in the C-terminal region of the envelope V3 loop. Proc. Natl. Acad. Sci. USA 1994, 91, 11378–11382. [Google Scholar] [CrossRef] [Green Version]
- Twigg, H.L.; Schnizlein-Bick, C.T.; Weiden, M.; Valentine, F.; Wheat, J.; Day, R.B.; Rominger, H.; Zheng, L.; Collman, R.G.; Coombs, R.W.; et al. Measurement of antiretroviral drugs in the lungs of HIV-infected patients. HIV Ther. 2010, 4, 247–251. [Google Scholar] [CrossRef] [Green Version]
- Twigg, H.L., 3rd; Iwamoto, G.K.; Soliman, D.M. Role of cytokines in alveolar macrophage accessory cell function in HIV-infected individuals. J. Immunol. 1992, 149, 1462–1469. [Google Scholar]
- Buhl, R.; Jaffe, H.A.; Holroyd, K.J.; Borok, Z.; Roum, J.H.; Mastrangeli, A.; Wells, F.B.; Kirby, M.; Saltini, C.; Crystal, R.G. Activation of alveolar macrophages in asymptomatic HIV-infected individuals. J. Immunol. 1993, 150, 1019–1028. [Google Scholar]
- White, D.A.; Gellene, R.A.; Gupta, S.; Cunningham-Rundles, C.; Stover, D.E. Pulmonary cell populations in the immunosuppressed patient. Bronchoalveolar lavage findings during episodes of pneumonitis. Chest 1985, 88, 352–359. [Google Scholar] [CrossRef] [PubMed]
- Wallace, J.M.; Barbers, R.G.; Oishi, J.S.; Prince, H. Cellular and T-lymphocyte subpopulation profiles in bronchoalveolar lavage fluid from patients with acquired immunodeficiency syndrome and pneumonitis. Am. Rev. Respir. Dis. 1984, 130, 786–790. [Google Scholar] [PubMed]
- Freire, M.O.; van Dyke, T.E. Natural resolution of inflammation. Periodontol. 2000 2013, 63, 149–164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bruggeman, L.A.; Dikman, S.; Meng, C.; Quaggin, S.E.; Coffman, T.M.; Klotman, P.E. Nephropathy in human immunodeficiency virus-1 transgenic mice is due to renal transgene expression. J. Clin. Investig. 1997, 100, 84–92. [Google Scholar] [CrossRef]
- Kopp, J.B.; Klotman, M.E.; Adler, S.H.; Bruggeman, L.A.; Dickie, P.; Marinos, N.J.; Eckhaus, M.; Bryant, J.L.; Notkins, A.L.; Klotman, P.E. Progressive glomerulosclerosis and enhanced renal accumulation of basement membrane components in mice transgenic for human immunodeficiency virus type 1 genes. Proc. Natl. Acad. Sci. USA 1992, 89, 1577–1581. [Google Scholar] [CrossRef] [Green Version]
- Ray, P.E.; Bruggeman, L.A.; Weeks, B.S.; Kopp, J.B.; Bryant, J.L.; Owens, J.W.; Notkins, A.L.; Klotman, P.E. bFGF and its low affinity receptors in the pathogenesis of HIV-associated nephropathy in transgenic mice. Kidney Int. 1994, 46, 759–772. [Google Scholar] [CrossRef] [Green Version]
- Barisoni, L.; Bruggeman, L.A.; Mundel, P.; D’Agati, V.D.; Klotman, P.E. HIV-1 induces renal epithelial dedifferentiation in a transgenic model of HIV-associated nephropathy. Kidney Int. 2000, 58, 173–181. [Google Scholar] [CrossRef] [Green Version]
- Jacob, B.A.; Porter, K.M.; Elms, S.C.; Cheng, P.Y.; Jones, D.P.; Sutliff, R.L. HIV-1-induced pulmonary oxidative and nitrosative stress: Exacerbated response to endotoxin administration in HIV-1 transgenic mouse model. Am. J. Physiol. Lung Cell. Mol. Physiol. 2006, 291, L811-9. [Google Scholar] [CrossRef] [Green Version]
- Bruggeman, L.A.; Thomson, M.M.; Nelson, P.J.; Kopp, J.B.; Rappaport, J.; Klotman, P.E.; Klotman, M.E. Patterns of HIV-1 mRNA expression in transgenic mice are tissue-dependent. Virology 1994, 202, 940–948. [Google Scholar] [CrossRef]
- Leonard, J.; Khillan, J.S.; Gendelman, H.E.; Adachi, A.; Lorenzo, S.; Westphal, H.; Martin, M.A.; Meltzer, M.S. The human immunodeficiency virus long terminal repeat is preferentially expressed in Langerhans cells in transgenic mice. Aids Res. Hum. Retrovir. 1989, 5, 421–430. [Google Scholar] [CrossRef]
- Putatunda, R.; Zhang, Y.; Li, F.; Yang, X.F.; Barbe, M.F.; Hu, W. Adult neurogenic deficits in HIV-1 Tg26 transgenic mice. J. Neuroinflamm. 2018, 15, 287. [Google Scholar] [CrossRef] [Green Version]
- Ammosova, T.; Platonov, M.; Ivanov, A.; Kont, Y.S.; Kumari, N.; Kehn-Hall, K.; Jerebtsova, M.; Kulkarni, A.A.; Uren, A.; Kovalskyy, D.; et al. 1E7-03, a low MW compound targeting host protein phosphatase-1, inhibits HIV-1 transcription. Br. J. Pharmacol. 2014, 171, 5059–5075. [Google Scholar] [PubMed] [Green Version]
- Jerebtsova, M.; Wong, E.; Przygodzki, R.; Tang, P.; Ray, P.E. A novel role of fibroblast growth factor-2 and pentosan polysulfate in the pathogenesis of intestinal bleeding in mice. Am. J. Physiol. Heart Circ. Physiol. 2007, 292, H743-50. [Google Scholar] [CrossRef] [PubMed]
- Heremans, H.; Dillen, C.; Groenen, M.; Matthys, P.; Billiau, A. Role of interferon-gamma and nitric oxide in pulmonary edema and death induced by lipopolysaccharide. Am. J. Respir. Crit. Care Med. 2000, 161, 110–117. [Google Scholar] [CrossRef] [PubMed]
- Brown, L.F.; Detmar, M.; Claffey, K.; Nagy, J.A.; Feng, D.; Dvorak, A.M.; Dvorak, H.F. Vascular permeability factor/vascular endothelial growth factor: A multifunctional angiogenic cytokine. Exs 1997, 79, 233–269. [Google Scholar] [PubMed]
- Bakakos, P.; Patentalakis, G.; Papi, A. Vascular Biomarkers in Asthma and COPD. Curr. Top. Med. Chem. 2016, 16, 1599–1609. [Google Scholar] [CrossRef] [PubMed]
- Kwon, J.; Wang, A.; Burke, D.J.; Boudreau, H.E.; Lekstrom, K.J.; Korzeniowska, A.; Sugamata, R.; Kim, Y.S.; Yi, L.; Ersoy, I.; et al. Peroxiredoxin 6 (Prdx6) supports NADPH oxidase1 (Nox1)-based superoxide generation and cell migration. Free Radic. Biol. Med. 2016, 96, 99–115. [Google Scholar] [CrossRef] [Green Version]
- Mattison, P.C.; Soler-Garcia, A.A.; Das, J.R.; Jerebtsova, M.; Perazzo, S.; Tang, P.; Ray, P.E. Role of circulating fibroblast growth factor-2 in lipopolysaccharide-induced acute kidney injury in mice. Pediatric Nephrol. 2012, 27, 469–483. [Google Scholar] [CrossRef] [Green Version]
- Lin, X.; Kumari, N.; DeMarino, C.; Kont, Y.S.; Ammosova, T.; Kulkarni, A.; Jerebtsova, M.; Vazquez-Meves, G.; Ivanov, A.; Dmytro, K.; et al. Inhibition of HIV-1 infection in humanized mice and metabolic stability of protein phosphatase-1-targeting small molecule 1E7-03. Oncotarget 2017, 8, 76749–76769. [Google Scholar] [CrossRef]
- Verollet, C.; Souriant, S.; Bonnaud, E.; Jolicoeur, P.; Raynaud-Messina, B.; Kinnaer, C.; Fourquaux, I.; Imle, A.; Benichou, S.; Fackler, O.T.; et al. HIV-1 reprograms the migration of macrophages. Blood 2015, 125, 1611–1622. [Google Scholar] [CrossRef]
- Morris, A.; Crothers, K.; Beck, J.M.; Huang, L.; American Thoracic Society Committee on H.I.V.P.D. An official ATS workshop report: Emerging issues and current controversies in HIV-associated pulmonary diseases. Proc. Am. Thorac. Soc. 2011, 8, 17–26. [Google Scholar] [CrossRef] [PubMed]
- Esser, R.; von Briesen, H.; Brugger, M.; Ceska, M.; Glienke, W.; Muller, S.; Rehm, A.; Rubsamen-Waigmann, H.; Andreesen, R. Secretory repertoire of HIV-infected human monocytes/macrophages. Pathobiol.: J. Immunopathol. Mol. Cell. Biol. 1991, 59, 219–222. [Google Scholar] [CrossRef] [PubMed]
- Cobos-Jimenez, V.; Booiman, T.; Hamann, J.; Kootstra, N.A. Macrophages and HIV-1. Curr. Opin. Hiv Aids 2011, 6, 385–390. [Google Scholar] [CrossRef]
- Verani, A.; Sironi, F.; Siccardi, A.G.; Lusso, P.; Vercelli, D. Inhibition of CXCR4-tropic HIV-1 infection by lipopolysaccharide: Evidence of different mechanisms in macrophages and T lymphocytes. J. Immunol. 2002, 168, 6388–6395. [Google Scholar] [CrossRef]
- Simard, S.; Maurais, E.; Gilbert, C.; Tremblay, M.J. LPS reduces HIV-1 replication in primary human macrophages partly through an endogenous production of type I interferons. Clin. Immunol. 2008, 127, 198–205. [Google Scholar] [CrossRef]
- Maus, U.A.; Koay, M.A.; Delbeck, T.; Mack, M.; Ermert, M.; Ermert, L.; Blackwell, T.S.; Christman, J.W.; Schlondorff, D.; Seeger, W.; et al. Role of resident alveolar macrophages in leukocyte traffic into the alveolar air space of intact mice. Am. J. Physiol. Lung Cell. Mol. Physiol. 2002, 282, L1245-52. [Google Scholar] [CrossRef] [Green Version]
- Borregaard, N. Neutrophils, from marrow to microbes. Immunity 2010, 33, 657–670. [Google Scholar] [CrossRef] [Green Version]
- Nathan, C. Neutrophils and immunity: Challenges and opportunities. Nat. Rev. Immunol. 2006, 6, 173–182. [Google Scholar] [CrossRef]
- Bergamini, A.; Faggioli, E.; Bolacchi, F.; Gessani, S.; Cappannoli, L.; Uccella, I.; Demin, F.; Capozzi, M.; Cicconi, R.; Placido, R.; et al. Enhanced production of tumor necrosis factor-alpha and interleukin-6 due to prolonged response to lipopolysaccharide in human macrophages infected in vitro with human immunodeficiency virus type 1. J. Infect. Dis. 1999, 179, 832–842. [Google Scholar] [CrossRef] [Green Version]
- Bierhaus, A.; Chen, J.; Liliensiek, B.; Nawroth, P.P. LPS and cytokine-activated endothelium. Semin. Thromb. Hemost. 2000, 26, 571–587. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Score Per Field | |||
|---|---|---|---|---|
| 0 | 1 | 2 | ||
| I. | Neutrophils in the alveolar space | 0 | 1–5 | >5 |
| II. | Neutrophils in the interstitial space | 0 | 1–5 | >5 |
| III. | Hyaline membranes | 0 | 1 | >1 |
| IV. | Proteinaceous debris filling air spaces | 0 | 1 | >1 |
| V. | Alveolar septal thickening | 0 | 2x–4x | >4x |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jerebtsova, M.; Ahmad, A.; Niu, X.; Rutagarama, O.; Nekhai, S. HIV-1 Transcription Inhibitor 1E7-03 Restores LPS-Induced Alteration of Lung Leukocytes’ Infiltration Dynamics and Resolves Inflammation in HIV Transgenic Mice. Viruses 2020, 12, 204. https://doi.org/10.3390/v12020204
Jerebtsova M, Ahmad A, Niu X, Rutagarama O, Nekhai S. HIV-1 Transcription Inhibitor 1E7-03 Restores LPS-Induced Alteration of Lung Leukocytes’ Infiltration Dynamics and Resolves Inflammation in HIV Transgenic Mice. Viruses. 2020; 12(2):204. https://doi.org/10.3390/v12020204
Chicago/Turabian StyleJerebtsova, Marina, Asrar Ahmad, Xiaomei Niu, Ornela Rutagarama, and Sergei Nekhai. 2020. "HIV-1 Transcription Inhibitor 1E7-03 Restores LPS-Induced Alteration of Lung Leukocytes’ Infiltration Dynamics and Resolves Inflammation in HIV Transgenic Mice" Viruses 12, no. 2: 204. https://doi.org/10.3390/v12020204