Throw out the Map: Neuropathogenesis of the Globally Expanding California Serogroup of Orthobunyaviruses


Laboratory of Persistent Viral Diseases, Rocky Mountain Laboratories, National Institute of Allergy and Infectious Diseases, National Institutes of Health, Hamilton, MT 59840, USA
*
Author to whom correspondence should be addressed.
Viruses 2019, 11(9), 794; https://doi.org/10.3390/v11090794
Submission received: 26 July 2019
/
Revised: 23 August 2019
/
Accepted: 26 August 2019
/
Published: 29 August 2019
(This article belongs to the Special Issue Neuroinvasive Viral Infections)
{kind=link}
{kind=link}
Abstract
:The California serogroup (CSG) comprises 18 serologically and genetically related mosquito-borne orthobunyaviruses. Of these viruses, at least seven have been shown to cause neurological disease in humans, including the leading cause of pediatric arboviral encephalitis in the USA, La Crosse virus. Despite the disease burden from these viruses, much is still unknown about the CSG viruses. This review summarizes our current knowledge of the CSG viruses, including human disease and the mechanisms of neuropathogenesis.
1. Introduction
The California serogroup (CSG) is a closely related group of orthobunyaviruses in the family Peribunyaviridae of the order Bunyavirales. Like all bunyaviruses, the CSG viruses contain tri-segmented, negative sense RNA genomes containing Large (L), Medium (M), and Small (S) genome segments. Classification of CSG viruses has fluctuated over time, as better serological assays and genetic techniques developed. California encephalitis virus (CEV) is the prototype member and was first isolated from mosquitoes in California in 1943 [1]. Currently, there are 18 recognized members of the CSG based on serological and genetic evidence. In addition to CEV, these are La Crosse virus (LACV), Snowshoe hare virus (SSHV), Tahyna virus (TAHV), Jamestown Canyon virus (JCV), Inkoo virus (INKV), Chatanga virus (CHATV), Keystone virus (KEYV), Jerry Slough virus (JSV), Lumbo virus (LUMV), Melao virus (MELV), San Angelo virus (SAV), Serra do Navio virus (SDNV), South River virus (SORV), Trivittatus virus (TVTV), Morro Bay virus (MBV), Achiote virus (ACHOV), and Infirmatus virus (INFV). Guaroa virus was briefly categorized as a CSG virus, however follow-up studies have since indicated it is a distinct virus [2].
All members of the CSG are presumed arboviruses, as all have been isolated from mosquitoes [3,4,5,6,7,8,9,10,11,12,13,14,15]. However, their vector, host ranges, geographic distribution, and ability to cause disease in humans differ, including their ability to cause neuroinvasive disease. This review will summarize our current knowledge of the members of the CSG and their ability to cause neuroinvasive disease in humans.
2. Relationships of the CSG Viruses
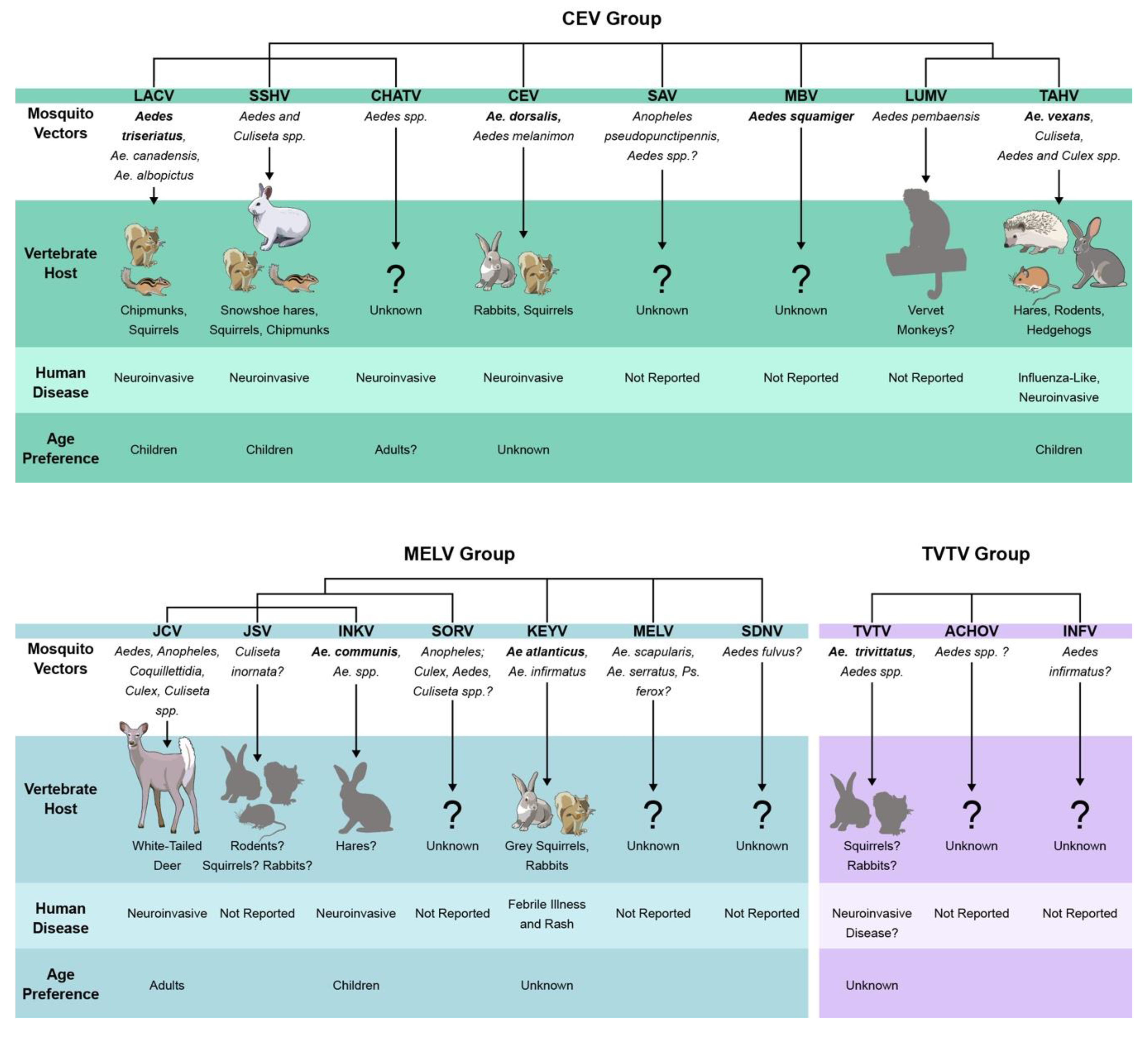
The CSG viruses are antigenically and genetically related, however determining their exact relationships with each other has been challenging. Based on traditional serological tests, three complexes within the CSG have been described: 1) the California encephalitis complex containing CEV, LACV, SSHV, CHATV, SAV, TAHV, LUMV, MBV, and INKV; 2) the Melao complex containing MELV, KEYV, SDNV, SORV, JCV, and JSV; and 3) the Trivittatus complex, containing TVTV and the recent additions of ACHOV and INFV [8,14,16,17]. Genetic characterization of the CSG viruses by phylogenetic analysis has shown that genetic relationships vary by genomic segment, viruses, strains, and methodology used in analyses. In recent years, several CSG phylogenies have been constructed using a variety of methods and virus strains, with some topology discrepancies [8,14,18,19]. However, some general conclusions of genetic relatedness can be made from these studies. Overall, the CSG viruses separate phylogenetically into three distinct groups, similar to but not identical to the serocomplexes: 1) CEV group containing CEV, LACV, SSHV, CHATV, SAV, TAHV, LUMV, and MBV; 2) MELV group containing MELV, KEYV, SDNV, SORV, JCV, JSV, and INKV; 3) TVTV group containing TVTV, ACHOV, and INFV [8,14,18,19]. MBV has only been included in phylogenies based on the S segment and appears to group most closely with CEV, which is how we have classified it in this review [17,19]. However, in one topology, MBV was distinct from the CEV clade [14]. These studies show that antigenic relatedness does not necessarily correlate with genetic relatedness, as INKV is classified in the CEV by serology, but the MELV group by phylogenetic analysis [14,18,20].
Based on results from the phylogenic studies of the CSG viruses in [8,14,18,19], within the CEV clade, SSHV, CHATV, and LACV are most closely related to each other, and LUMV and TAHV are closest to each other. The relationship of CEV and SAV within the CEV clade varies by phylogeny. In the MELV clade, JCV, JSV, and INKV are the most closely related to each other, and these three also form a clade with SORV. The relationship of MELV, SDNV, and KEYV is more variable within the clade. In all phylogenies, TVTV forms a distinct clade, and in the study first describing ACHOV and INFV as members of the CSG, these two viruses always grouped most closely with TVTV, expanding the group to three members [8,14,18,19]. These relationships are visually compiled and summarized in Figure 1.
3. Geographic Distribution
The CSG viruses expand across the globe, however the geographic distribution of each CSG virus differs, likely restricted by the range of their vertebrate hosts and mosquito vectors. Figure 2 summarizes the reported geographic distribution of each CSG virus based on human cases, seroprevalence studies, and virus isolations from mosquitoes. Eleven CSG viruses were detected in the USA, including LACV, SSHV, JCV, KEYV, TVTV, CEV, SORV, MBV, JSV, SAV, and INFV (Figure 2; [3,4,5,9,14,15,22,23,24,25,26,27,28,29,30,31,32,33,34,35,36,37,38,39,40,41,42,43,44,45,46,47,48]. SSHV, JCV, and TVTV have also been detected in Canada, and SORV has also been found in the Yucatan Peninsula of Mexico (Figure 2; [23,49,50]). ACHOV was isolated from mosquitoes in Panama [14], MELV in Trinidad and Brazil [13,51], and SDNV in Brazil [52]. INKV and CHATV are primarily found in Scandinavia and Russia [53,54,55,56], while TAHV is found throughout Europe, Russia, Asia, and Africa [53,57,58,59,60,61,62]. LUMV is the only CSG member found, thus far, exclusively in Africa [12,61]. Some of the CSG viruses have only been isolated a limited number of times, so it is unclear if all 18 viruses are actively circulating. The exact range of the CSG viruses is unclear, as limited studies have been conducted and they are not consistently monitored, so the range of these viruses may be more widespread than reported. Furthermore, as climate change and globalization alters the territories of mosquitoes, the geographic distribution of the CSG viruses is likely to change as well.
4. Vector and Host Range
The CSG viruses have been isolated from a variety of mosquito species, primarily in the Aedes genera. Mosquito taxonomy has been a controversial topic in the 2000s, where many of the Aedes species were reclassified, but for the purpose of this review we will use the mosquito classification proposed by Wilkerson et al. in 2015 and used in Walter Reed’s Systematic Catalog of Culicidae (mosquitocatalog.org) [63]. Many CSG viruses have been isolated from a variety of different species and genera, however, not all mosquitoes that carry a virus are competent vectors. It can be difficult to establish a primary vector for arboviruses, and primary vector species can vary by region even for the same virus. However, for some of the CSG viruses the primary vector is well established. The CSG viruses and their potential vector relationships are summarized in Figure 1. Primary vectors for several of the CSG viruses have been established based on field isolations and transmission studies using suckling mice and other mammals. These include Ae. triseriatus for LACV, Ae. atlanticus for KEYV, Ae. trivittatus for TVTV, Ae. squamiger for MBV, and Ae. dorsalis for CEV (Figure 1), [5,10,11,26,30,42,64,65,66,67,68,69,70,71]). Ae. canadensis and Ae. albopictus have been proposed as additional vectors of LACV [72], [73]. JCV and SSHV have both been isolated from a wide variety of mosquitoes (and horseflies in the case of JCV), and several mosquito species are capable of transmitting these viruses in laboratory experiments (Figure 1) [5,26,30,74,75,76,77,78,79,80,81,82]. It is unclear if JCV and SSHV utilize multiple primary vectors, or if they are simply easily transmittable to suckling mice under laboratory conditions. INKV has nearly exclusively been isolated from Ae. communis, thus it has been assumed to be INKV’s primary vector [7,54,83]. TAHV has been isolated from Aedes, Culiseta, Culex, and Anopheles species, and Ae. vexans and Ae. cantans have been implicated as primary vectors [53,84,85].
Several of the CSG viruses have only been isolated once or a handful of times. ACHOV was isolated from a pool of Aedes species, INFV from Ae. infirmatus, JSV from Cs. inornata, SDNV from Ae. fulvus, and LUMV from Ae. pembaensis. SAV, SORV, and MELV have been isolated from several mosquito species; SAV from An. pseudopunctipennis, Ps. columbiae, Ae. atlanticus, and Ae. infirmatus, SORV from Anopheles, Culex, Aedes, and Culiseta species, and MELV from Ae. scapularis, Ae. serratus, Ps. ferox, and Culex species [5,11,13,48,50,86,87]. CHATV was isolated from pools of unidentified mosquitoes, and positive pools were sequenced for mosquito mitochondrial cytochrome c oxidase to try to identify mosquitoes in the pool. From the pooled sequencing, Ae. cantans, Ae. annulipes, Ae. cinereus, Ae. communis, Ae. punctor were suggested as possible vectors [8]. Because of the limited isolations of these viruses, their primary vectors are still unknown.
Eight of the CSG viruses, LACV, CEV, JCV, TAHV, KEYV, TVTV, SSHV, and MBV are transovarially transmitted in their mosquito vectors, which likely contributes to their survival over winter ([30,67,69,88,89,90,91,92,93,94,95]. SAV has also been shown to be transmitted transovarially in Ae. albopictus, though the virus has not been isolated from this mosquito in nature, so the significance of transovarial transmission in SAV’s natural life cycle remains unclear [96]. Additionally, INKV RNA was detected in Ae. communis larvae, suggesting transovarial transmission of this virus as well [97]. Too little information is available on the other CSG viruses to know if they are transovarially transmitted, however it is possible that transovarial transmission is a common feature among the CSG viruses.
The CSG viruses have been shown to utilize a variety of mammal hosts in their natural life cycle, and suspected host and reservoir species are summarized in Figure 1. Based on serological surveys and the development of viremia after experimental inoculations, small mammals such as squirrels, chipmunks, other rodents, and/or hares have been implicated as hosts for LACV, CEV, SSHV, TAHV, KEYV, and possibly JSV (Figure 1, [1,11,53,65,67,92,98,99,100]). In addition to experimental viremia, SSHV has been isolated from hares in the wild during winter months, suggesting long-term infection of these animals and potentially a second over-wintering strategy for SSHV [11]. Experimental infections with LUMV resulted in viremia in vervet monkeys, but information on natural infections is lacking [12]. JCV, thus far, appears to be the only CSG virus to primarily utilize large mammals as its reservoir. White-tailed deer are its primary host, but other large ungulates such as moose, elk, and bison have also been implicated as amplifying hosts and neutralizing antibody has been detected in domestic animals such as sheep, cattle, and horses [30,38,101,102,103]. Seropositivity to SSHV has been demonstrated in large mammals as well, however, experimental infection of deer with SSHV did not produce viremia [99,101,103,104,105].
For INKV, SAV, and TVTV, vertebrate hosts have been speculated on based solely on serological surveys, however the role of these seropositive animals as reservoirs is unclear. For INKV, high rates of seropositivity have been shown in reindeer, moose, cows, red foxes, and hares; neutralizing antibody (NAb) to SAV has been reported in coyotes, raccoons, and opossums; and NAb to TVTV has been found in rabbits, fox squirrels, opossums, and raccoons [7,11,41]. There is little to no evidence as to the vertebrate hosts of MELV, CHATV, SORV, SDNV, MBV, INFV, and ACHOV.
5. Replication
Few studies have been done specifically looking at the replication cycle of CSG viruses. What is known about the CSG virus genomic organization and replication cycle is based primarily on studies of LACV and SSHV. Additional information about the CSG replication cycle has been inferred from studies of Bunyamwera virus (BUNV), the Orthobunyavirus prototype, and other orthobunyaviruses (reviewed in [106]). The CSG viruses, like all orthobunyaviruses, contain three single-stranded negative sense RNA genome segments. The exact length of each genome segment varies by virus, but typically are about 6.9 kb (L), 4.5 kb (M), and 1.0 kb (S) [106]. Each genome segment is flanked by complementary 5’ and 3’ UTRs, which circularize each genome segment by forming a panhandle structure that is used as the viral promoter [107,108]. Encapsidation of the genome segments is initiated by interactions between the 5’ end of the genomic segments and the Nucleocapsid (N) protein, resulting in ribonucleoprotein (RNP) complexes [109,110,111,112]. These genomic RNPs associate with the L protein in the virion through interactions with the UTRs [106]. The L protein is encoded by the L segment; the M segment encodes the two envelope glycoproteins (Gn [previously G2] and Gc [previously G1]) and a non-structural protein (NSm), and the S segment encodes the N protein and a non-structural protein (NSs) [113,114,115,116]. M encodes a single open-reading frame (ORF) resulting in the glycoprotein precursor (GPC) polyprotein in the order Gn-NSm-Gc [117]. In studies of BUNV, it was shown that GPC is processed into mature proteins by internal signal peptides and host signal peptide peptidase [118]. N and NSs are encoded by overlapping reading frames on the S segment, with NSs translation resulting from leaky ribosomal scanning [119,120].
Much of the process of CSG virus entry into cells is still unknown. The CSG viruses enter cells via receptor-mediated endocytosis initiated by binding of the Gn/Gc heterodimer, likely mediated by Gc, with a currently unknown cell receptor [106,121,122]. DC-SIGN has been implicated as a potential LACV receptor [123]. However, DC-SIGN is likely not the only, or primary, receptor as its expression is limited to certain Dendritic cell and macrophage types and LACV, in the CNS, primarily infects neurons [124,125]. Studies of the related Akabane and Schmallenberg (SBV) orthobunyaviruses have shown that heparan sulfate facilitates viral attachment to cells, however further studies are needed to know if this is an important attachment factor for the CSG viruses as well [126].
Studies of LACV have shown that upon attachment, the virus enters the cell via clathrin-mediated endocytosis [127]. Uncoating of LACV occurs in the early endosome where the change in pH induces conformational changes of the envelope proteins mediated by the Gc fusion peptide, resulting in the release of the RNPs into the cytoplasm [127,128,129,130,131]. LACV transcription is primed via cap-snatching of the 5’ 7-methylguanylate capped ends of host-derived mRNAs, in a process facilitated by an endonuclease domain on the L protein [132,133]. Transcription of LACV, like all Bunyaviruses, requires concurrent host translation to prevent premature termination by the RNA-dependent RNA polymerase (RdRp), and results in viral mRNAs lacking a poly(A) tail [106,134,135,136]. Studies of BUNV and Oropouche (OROV) orthobunyavirus have shown that these viruses form Golgi-derived viral factories for replication [137,138]. Genome replication is primer-independent and is carried out by the RdRp, encoded by the L protein, on full-length anti-sense RNPs [106].
Orthobunyavirus virions assemble, mature, and bud at the membrane of the Golgi [106]. For BUNV, the viral RNPs are trafficked to the glycoprotein-expressing Golgi membrane, and genome packaging is facilitated by the UTRs [106,109]. For OROV, remodeling of the Golgi membranes to create viral factories requires endosomal complex required for transport (ESCRT) machinery, and this process is likely required for virus assembly and budding [138]. Budding is facilitated by the interactions of the RNPs with Gn and Gc, virions are released from the cell via exocytosis, and the glycoproteins undergo a final maturation step upon exit [139,140,141,142]. The CSG viruses likely utilize a similar mechanism, as Gn and Gc of LACV are processed in the Golgi [121,143]. However, further studies specifically on the CSG viruses are needed to elucidate differences in replication within and between the CSG viruses and other orthobunyaviruses.
6. Human Infection and Disease
Several CSG viruses have been shown to cause disease in humans, ranging from mild rashes to severe neurological disease (Figure 1). However, infection and disease rates are difficult to determine as serosurveys have primarily focused on the CSG viruses capable of causing human disease and are complicated by the cross-reactivity among CSG viruses. Moderate to high seroprevalence rates in endemic regions of many CSG viruses suggest that these viruses frequently infect humans, but that symptomatic disease is rare. Depending on the region sampled, reported seroprevalence rates for CSG members are up to 27% for LACV, JCV, CEV, KEYV, LUMV and TVTV, up to 42% for SSHV, 24–50% for INKV and/or CHATV, and up to 80% for TAHV [1,8,12,25,47,53,54,144,145,146,147,148,149,150], Even when clinical cases do arise, it can be difficult to diagnose the specific CSG virus responsible due to the high amount of antibody cross-reactivity within the serogroup. Additionally, historical cases of CSG viral disease may have been misattributed to the wrong CSG virus due to less sensitive diagnostic techniques, lack of inclusion of relevant CSG virus comparisons, or they were performed prior to the identification of other CSG viruses.
Of the 18 CSG viruses, eight have caused confirmed human disease (Figure 1). CEV, LACV, SSHV, JCV, TAHV, INKV, and CHATV have been shown to cause neuroinvasive disease [1,55,84,151,152,153]. KEYV was associated with one case of febrile illness and diffuse rash without CNS involvement [146]. TVTV was retrospectively implicated in one case of human neurological disease in New York in 1981, but no additional cases have been reported since, so it is difficult to say if TVTV causes human disease [43]. Among the CSG viruses that cause neurological disease, the incidence and ages of susceptibility vary.
In North America, LACV, JCV, and SSHV are the main disease-causing CSG viruses. LACV traditionally has caused the most cases of neuroinvasive disease annually, primarily in children, and is the leading cause of pediatric arboviral encephalitis in the USA with an average of ~70 reported neuroinvasive cases per year [154]. In recent years, JCV has been responsible for an increasing number of neuroinvasive cases, which may be due to better arboviral reporting and testing, or it may reflect an emergence of this pathogen. In the USA, the CDC started routine JCV IgM testing in 2013 and since then ~25 neuroinvasive cases are reported in the US annually [22]. JCV neuroinvasive cases also occur in Canada every year [23,155]. Unlike LACV, JCV appears to affect more adults than children [24,43]. SSHV causes several reported cases of neuroinvasive disease annually in Canada, primarily in children [23,152]. Since the first reports of CEV neuroinvasive disease in three people in 1945, only one other confirmed case of CEV has been reported, a 65-year-old man with blurred vision and dizziness in 1996 [1,156]. All four CEV cases occurred in California.
In Europe, INKV, CHATV, and TAHV cause human disease. TAHV disease primarily manifests as a flu-like illness called “Valtice fever”, and only rarely causes neuroinvasive disease [53,157,158]. Like LACV and SSHV, TAHV primarily causes disease in children [53]. Neuroinvasive cases of INKV and CHATV were described in Finland during a retrospective study of serum samples from patients with suspected viral neuroinvasive disease [55]. INKV caused neuroinvasive disease in both children and adults, but children appeared to have more severe disease than adults [55]. There were three suspected and confirmed CHATV cases, all of which were adults (≥40 years old) [55].
CSG virus neuroinvasive disease symptoms include headache, fever, dizziness, vomiting, muscle weakness, stiff neck, seizures, encephalitis, meningoencephalitis, and meningitis [23,55,59,154,159,160]. While most patients with CSG virus neuroinvasive disease fully recover, some cases result in long-term and serious sequelae including seizures, hemiparesis, cognitive deficits, and behavior changes [154,161,162]. LACV has a reported case fatality rate of up to 1.9% in children [35]. Fatalities from the other CSG viruses have not been reported. Because mild illnesses are not included and testing for CSG viruses is not routinely done unless arboviral disease is suspected, disease from the CSG viruses is likely underreported.
7. Neuropathogenesis – Experimental Infections of Animals
Despite the ability of some of the CSG viruses to cause neuroinvasive disease, and new cases occurring every year, our understanding of the pathogenesis of these viruses is still limited. Traditionally, CSG virus pathogenesis was evaluated using a suckling mouse model via intracranial (IC) inoculation or mosquito transmission. KEYV, LUMV, MELV, TVTV, JCV, SSHV, CEV, TAHV, and LACV have all been shown to cause disease and death in suckling mice [3,12,13,30,51,67,68,78,128,163]. Suckling mice were particularly useful for mosquito transmission studies, but because they are so susceptible to CSG virus infection; they are not a very informative model for pathogenesis studies.
Weanling (3–4-week-old) and adult mice are also susceptible to many CSG viruses; however, susceptibility depends on inoculation route and dose. Pathogenicity of viruses that cause neurological disease can be evaluated by a number of parameters. Neuroinvasion describes the ability of the virus to enter the brain from the periphery, regardless of the development of disease. Neuroinvasive disease is evaluated by the ability of the virus to enter the CNS from the periphery and cause disease. Neurovirulence describes the ability of the virus to replicate in the brain following a direct route of inoculation, such as IC or intranasal (IN), and neuropathogenicity is the ability of the virus to cause neurological disease.
Of the CSG viruses that have not been shown to cause human neurological disease, all that have been tested in weanling mice are neurovirulent, however not all are neuroinvasive. Following IC inoculation of weanling mice, LUMV, MELV and KEYV all cause disease indicating neurovirulence [9,12,13]. However, LUMV is not pathogenic following IP inoculation indicating it is not neuroinvasive. Some strains of MELV and SAV can cause disease in weanling mice or hamsters following IP inoculation, indicating these viruses are neuroinvasive and neurovirulent [12,51,164].
Interestingly, similar patterns of pathogenicity are observed in weanling mice inoculated with the CSG viruses known to cause human disease. Weanling mice are susceptible to disease from CEV, JCV, and INKV following IC or IN inoculation, but not following IP inoculation, indicating these viruses are neurovirulent, but not neuroinvasive, in mice [3,21,165]. LACV, SSHV, and TAHV all cause neuroinvasive disease in weanling mice following IP inoculation. Of note, the ability of TAHV to cause neuroinvasive disease varies by strain and requires a high dose, suggesting that it is less virulent than LACV or SSHV [21,125,166,167,168]. Neuroinvasive disease caused by LACV, SSHV, and TAHV is age-dependent, as adult and aged mice are resistant to neuroinvasive disease after IP inoculation [21,163,167]. However, LACV, SSHV, TAHV, and JCV are highly neurovirulent and cause disease in adult and aged mice when inoculated IN. In contrast, INKV caused disease in only about 20-25% of mice when inoculated at a high dose IN in adult mice, suggesting this virus differs in neuropathogenesis compared to the other CSG viruses that cause human disease [21].
The results of the mouse infection studies reflect, to an extent, human disease caused by the CSG viruses. LACV, SSHV, TAHV, and INKV all have been reported to cause more serious neuroinvasive disease in children than adults. LACV causes the most reported neuroinvasive cases annually, followed by SSHV, then TAHV and INKV. At a high dose (105 PFU virus), LACV, SSHV, and TAHV all cause neuroinvasive disease in the majority of weanling mice, however LACV and SSHV, but not TAHV, maintain their pathogenicity at a low dose of 103 PFU [21,167]. Additionally, LACV causes disease in nearly 100% of mice inoculated but SSHV causes disease in ~60–80% of mice [21]. These results may reflect differences in virulence and account for the higher number of LACV neuroinvasive cases reported in humans compared to SSHV and TAHV. In humans, INKV causes few neuroinvasive cases and disease is more severe in children than adults. In mice, INKV did not cause neuroinvasive disease, however, weanling mice had a higher incidence of disease than adult mice when inoculated IN, correlating with reported disease in humans [21]. Because some of the CSG viruses that have not been reported to cause neurological disease in humans can cause disease when inoculated into animals IC, it is possible that neurotropism is a shared trait among the CSG viruses. The ability to enter the brain may determine which CSG viruses cause neuroinvasive disease in humans. It is also possible that some cases of neurological disease in humans have been misattributed to the wrong CSG virus due to the high level of cross-reactivity among the group. The development of more specific diagnostic tests will provide a more accurate account of which CSG viruses cause neurological disease in humans.
In experimental infections of rhesus monkeys inoculated subcutaneously or intramuscularly with LACV, TAHV, or JCV, none of the monkeys developed disease [125,165,168]. However, nearly all monkeys developed high neutralizing antibody titers, indicating they were productively infected [125,165,168]. These results suggest that CSG virus infection in monkeys may be akin to human infections, where infection and NAb responses happen readily, but neuroinvasive disease is a rare event.
8. Neuropathogenesis – Molecular Mechanisms
Much of what is known mechanistically about CSG virus pathogenesis is based on studies of LACV and has focused on the host interferon (IFN) response. The IFN response, primarily type-I IFNs α and β, has been shown to be important in many viral infections, including LACV [169]. IFNs are activated through a variety of pathways in response to RNA viruses. These include the RIG-I-like receptor (RLR) and Toll-like receptor (TLR) pathways. In general, RLR pathways are activated in response to RNA viruses in the cytoplasm, and signal through mitochondrial antiviral-signaling proteins (MAVS, aka IPS-1) and interferon regulatory factor (IRF)3 and IRF7 [170,171,172]. TLRs 3, 7, and 9 are endosomal and can recognize viral RNA within endosomes and use the myeloid differentiation factor-88 (MyD88) adaptor protein in the case of TLR7 and TLR9 [173]. Signaling through these pathways stimulates the production of Type-I IFNs which are released from cells, bind to cellular IFN receptors (IFNAR), then signal through the JAK–STAT (Janus kinase-signal transducer and activator of transcription proteins) pathway to induce the expression of IFN-stimulated genes and effector molecules with antiviral properties [173].
Investigations into the mechanism of age-dependent susceptibility to LACV in mice have shown that the IFN response has an important role. While wild-type adult mice are resistant to neuroinvasive disease, adult Ifnar-/- mice are highly susceptible to neuroinvasive disease following IP inoculation of LACV, indicating that the type I interferon (IFN) response plays a crucial role in protection [169,174,175]. Adult Irf3/Irf7-/-, Mavs-/-, and Unc93b13D (deficient in TLR3/7/9) mice, but not Myd88-/- mice, have an increased susceptibility to neuroinvasive LACV disease, indicating that protection against LACV requires both RLRs and TLRs [167]. In wild-type (WT) mice, a robust peripheral IFNα4 and IFNβ response was observed in adult mice, but not in weanlings, and myeloid DCs were responsible for their expression. In weanling mice, mDCs had little production of type-I IFNS, leading to the susceptibility of young mice to neuroinvasive disease [167]. The IFN effector protein myxovirus resistance protein 1 (MxA) has been shown to be capable of restricting LACV growth in vitro, and MxA transgenic Ifnar-/- mice are less susceptible to neuroinvasive disease, indicating an important role for MxA in protection [176,177]. Additionally, the adaptive immune response also plays a role in age-dependent susceptibility to LACV neuroinvasive disease. In adult mice, depletion of CD4+ helper and CD8+ cytotoxic T cells or depletion of B cells results in increased susceptibility of mice to neurological disease, indicating a protective role of lymphocytes in adult mice [178]. In contrast, when T and B cells are depleted in weanling mice, neuropathogenesis is not affected despite infiltration of these cells into the brain during disease in WT mice [178].
In addition to mediating age-dependent susceptibility to CSG viruses, immune responses may also influence the susceptibility of host species. In studies comparing restriction of LACV, which infects humans but not ruminants, and SBV, which infects ruminants but not humans, by human and sheep restriction factor BST-2/tetherin, LACV was restricted by sheep BST-2, but not human BST-2, and the reverse was true for SBV, suggesting a mechanism of orthobunyavirus host determination [179]. However, whether BST-2 restriction plays a role in age-related CSG virus pathogenesis, or if restriction differs between CSG viruses, has not been investigated.
Dissemination of the CSG viruses from the periphery to the brain likely occurs with the virus first replicating near the site of entry (via mosquito bite or inoculation) in the muscle, lymph nodes, or spleen [125,163,180,181]. Infectious LACV and RNA can be found at low levels in many tissues throughout the body after IP inoculation of weanling and suckling mice, suggesting either a wide tissue tropism for the virus, or representing virus in the blood within these tissues [125,163]. In support of the latter, short viremia is observed in weanling mice, likely contributing to its dissemination and presence in multiple tissues, but viral antigen staining has not been reported for peripheral tissues, suggesting a lack of, or limited, replication in peripheral tissues ([125], unpublished observations). Entry of LACV into the CNS is mediated by blood-brain-barrier (BBB) breakdown via vascular leakage of the capillaries in the olfactory bulb, which allows LACV entry into the brain via a hematogenous route rather than axonal transport in olfactory sensory neurons [182]. Mice deficient in the TAM receptor Mertk, which is involved in innate immune regulation, are more susceptible to LACV neuroinvasive disease [183]. This susceptibility was associated with increased BBB permeability, suggesting TAM receptors may be involved in BBB integrity during LACV infection [183].
Once in the CNS, the primary site of replication for the CSG viruses is neurons, and the CSG viruses readily replicate in neuronal cultures in vitro [21,125,163,166]. Co-cultures of neurons and astrocytes have shown that astrocytes are susceptible to LACV infection in vitro, however this does not appear to be a main target cell in vivo [184]. LACV, SSHV, TAHV, and JCV all replicate extensively throughout the mouse brain, however INKV’s replication is limited [21]. Neurological disease from LACV is the result of neuronal damage and cell death, and this damage is mediated by sterile alpha and TIR-containing motif 1 (SARM1), a MyD88-related protein expressed at high levels in neurons and associated with multiple forms of neuronal death [185], [186]. In vitro, siRNA to Sarm1 reduced LACV-induced death in neuronal cultures, and in vivo, weanling Sarm1-/- mice were less susceptible to neuroinvasive disease following LACV inoculation, despite similar amounts of virus in the brains of WT and Sarm1-/- mice [186]. In neuronal cultures in vitro, SSHV, TAHV, JCV, and INKV also induce cell death, however, depending on cell type, not typically to the same degree as LACV [21]. Further studies are needed to determine if neuronal cell death caused by the other CSG viruses is similarly mediated by SARM1.
In weanling mice inoculated with LACV, infiltrating monocytes/macrophages are associated with areas of infection [125]. Characterization of these cells demonstrated that the majority were inflammatory monocytes, which are defined by their high expression of the cell surface marker Ly6C, as well as CCR2, a chemokine receptor that is involved in monocyte trafficking from the bone marrow to the blood. Surprisingly, unlike other neuroinvasive viral infections, CCR2 is not required for monocyte recruitment to the blood following LACV infection [187]. Within the brain, however, iMO trafficking to sites of infection is CCR2-dependent [187]. Interestingly, CSG viruses vary in their iMO recruitment mechanism. JCV induces a similar CCR2-independent monocyte recruitment mechanism from the bone marrow to the blood [187]. However, iMO recruitment from the bone marrow during TAHV infection is CCR2-dependent [187]. These results are particularly interesting in the respect that LACV and TAHV are more closely related to each other than to JCV; LACV and TAHV are both neuroinvasive in weanling mice and JCV is not, yet LACV and JCV share a monocyte recruitment mechanism distinct from TAHV. This highlights the need for studies to better understand the mechanistic differences of pathogenesis between the CSG viruses.
Studies investigating the virulence factors of the CSG viruses have shown that the M segment, L protein, and NSs all contribute to LACV virulence. In experiments that utilized reassortants between a virulent LACV strain and a lowly virulent TAHV strain (as determined by neuroinvasion in suckling mice), neuroinvasiveness primarily mapped to the M segment [188]. However, some intermediate phenotypes were observed suggesting compensatory mutations could exist in the L and/or S segments [188]. In additional studies using an attenuated CSG virus clone, neurovirulence was primarily determined by the L segment, but again the neurovirulence phenotype also appeared dependent on the other genomic segments [180,189]. More recently, a recombinant virus containing the LACV backbone with the M ORF from JCV (rLACV/JCV) showed that the chimeric virus was highly attenuated and unable to cause disease after either IP or IC inoculation at high doses, a surprising result considering both parental LACV and JCV viruses were highly neurovirulent [190]. Taken together, the results of these studies suggest that interactions between multiple CSG viral proteins encoded from multiple genomic segments determine CSG virus virulence. Individual CSG virus proteins may have unexpected phenotypes when expressed outside of their specific genetic context. This may be an advantageous feature in terms of vaccine development, as the highly attenuated rLACV/JCV virus was highly immunogenic and when mice and monkeys were immunized with it, they were protected from subsequent LACV, TAHV, and JCV challenge [190]. However, if reassortants between the CSG viruses can have unique phenotypes from either parental virus, it is possible that more virulent novel reassortants may arise.
An important virulence factor for LACV is NSs. Utilizing a recombinant LACV virus with an NSs deletion, it was shown that NSs antagonizes the type-I IFN response in vitro and in vivo [191,192]. The NSs deletion mutant strongly induced type-I IFN responses in vitro and in vivo, whereas WT LACV did not induce high levels of type-I IFN [192]. NSs appears to not play a role in pathogenesis outside of IFN antagonism, as no difference in virulence was observed between WT and NSs deletion LACV viruses in cells and mice deficient in type-I IFN responses [192]. NSs antagonizes the IFN response by targeting and degrading host RNA Polymerase II [192,193]. NSs did not affect LACV’s ability to infect neurons in the mouse CNS, and ultimately most mice still succumbed to disease from the NSs deletion virus [192]. The authors speculated that this may be due to a lack of an appropriate IFN effector protein, as the MxA homologue in mice, Mx2, is non-functional in C57BL/6 mice, the strain used for the studies [192]. It remains to be seen whether differences in virulence between the CSG viruses may be due to differences in the ability of their NSs to antagonize host IFN responses.
The majority of pathogenesis studies of the CSG viruses have focused on LACV. Studies that have examined multiple CSG viruses have often found differences between CSG viruses. Further studies are clearly needed to elucidate the mechanisms of pathogenesis for the different neuroinvasive CSG viruses.
9. Conclusions and Future Perspectives
The California serogroup of orthobunyaviruses comprises a large, related group of mosquito-borne viruses with differing geographical distributions, vector and host ranges, and abilities to cause human disease. Despite the continual, and possibly increasing, disease burden from the CSG viruses, much is still unknown. Of particular interest is why only a subset of infections in humans result in neurological disease, and why susceptibility to specific viruses differs with age. Research efforts have primarily focused on LACV and a handful of other CSG viruses. From these studies, we know that the mechanism of LACV pathogenesis can vary from other CSG viruses, highlighting the importance of future research into LACV and other CSG viruses. Additionally, as climate change continues, the geographical range and abundance of arboviral mosquito vectors is also changing [194]. Many of the human disease-causing CSG viruses already exist in overlapping geographical regions and share some vector and host species. As mosquito ranges expand, the potential for coinfections and novel reassortant CSG viruses increases. Laboratory infections of mosquitoes with LACV and SSHV have shown that it is possible to recover reassortant viruses from coinfected mosquitoes, suggesting the possibility of the occurrence of highly virulent novel reassortants in nature [195,196]. Elucidating the differences between the neuroinvasive CSG viruses and the non-neuroinvasive CSG viruses, and differences in virulence between the neuroinvasive CSG viruses, will help us understand how these viruses cause disease. A better understanding of the underlying mechanisms of pathogenesis of different CSG viruses will help in the development of treatments and vaccines.
Author Contributions
Conceptualization: A.E. and K.P.; writing—original draft preparation: A.E.; writing—review and editing: A.E. and K.P.
Funding
This research was supported by the Intramural Research Program of NIH.
Acknowledgments
We thank Austin Athman and Ryan Kissinger for figure construction.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Hammon, W.M.; Reeves, W.C. California Encephalitis Virus—A Newly Described Agent. Calif. Med. 1952, 77, 303–309. [Google Scholar]
- Aguilar, P.V.; Morrison, A.C.; Rocha, C.; Watts, D.M.; Beingolea, L.; Suarez, V.; Vargas, J.; Cruz, C.; Guevara, C.; Montgomery, J.M.; et al. Guaroa Virus Infection among Humans in Bolivia and Peru. Am. J. Trop. Med. Hyg. 2010, 83, 714–721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hammon, W.M.; Reeves, W.C.; Sather, G. California encephalitis virus, a newly described agent. II. Isolations and attempts to identify and characterize the agent. J. Immunol. 1952, 69, 493–510. [Google Scholar] [PubMed]
- Thompson, W.H.; Anslow, R.O.; Hanson, R.P.; DeFoliart, G.R. La Crosse Virus Isolations from Mosquitoes in Wisconsin, 1964–68. Am. J. Trop. Med. Hyg. 1972, 21, 90–96. [Google Scholar] [CrossRef] [PubMed]
- Sudia, W.D.; Chamberlain, R.W.; Calisher, C.H.; Newhouse, V.F. California Group Arboviruses—Isolations from Mosquitos in North-America. Mosq. News 1971, 31, 576–600. [Google Scholar]
- Bardos, V.; Danielova, V. The Tahyna virus—A virus isolated from mosquitoes in Czechoslovakia. J. Hyg. Epidemiol. Microbiol. Immunol. 1959, 3, 264–276. [Google Scholar] [PubMed]
- Brummer-Korvenkontio, M.; Saikku, P.; Korhonen, P.; Ulmanen, I.; Reunala, T.; Karvonen, J. Arboviruses in Finland. IV. Isolation and characterization of Inkoo virus, a Finnish representative of the California group. Am. J. Trop. Med. Hyg. 1973, 22, 404–413. [Google Scholar] [CrossRef]
- Putkuri, N.; Kurkela, S.; Levanov, L.; Huhtamo, E.; Vaheri, A.; Sironen, T.; Vapalahti, O. Isolation and characterization of a California encephalitis serogroup orthobunyavirus from Finnish mosquitoes. Infect. Genet. Evol. 2014, 22, 164–173. [Google Scholar] [CrossRef]
- Bond, J.O.; Hammon, W.M.; Lewis, A.L.; Sather, G.E.; Taylor, D.J. California group arboviruses in Florida and report of a new strain, Keystone virus. Public Health Rep. 1966, 81, 607–613. [Google Scholar] [CrossRef]
- DeFoliart, G.R.; Anslow, R.O.; Thompson, W.H.; Hanson, R.P.; Wright, R.E.; Sather, G.E. Isolations of Trivittatus Virus from Wisconsin Mosquitoes, 1964–1968. J. Med. Èntomol. 1972, 9, 67–70. [Google Scholar] [CrossRef]
- LeDuc, J.W. The ecology of California group viruses. J. Med. Èntomol. 1979, 16, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Kokernot, R.H.; McIntosh, B.M.; Worth, C.B.; Moraist, D.; Weinbren, M.P. Isolation of viruses from mosquitoes collected at Lumbo, Mozambique. I. Lumbo virus, a new virus isolated from Aedes (Skusea) pembaensis Theobald. Am. J. Trop. Med. Hyg. 1962, 11, 678–682. [Google Scholar] [CrossRef] [PubMed]
- Spence, L.; Anderson, C.R.; Aitken, T.H.G.; Downs, W.G. Melao Virus, a New Agent Isolated from Trinidadian Mosquitoes. Am. J. Trop. Med. Hyg. 1962, 11, 687–690. [Google Scholar] [CrossRef] [PubMed]
- Rogers, M.B.; Gulino, K.M.; Tesh, R.B.; Cui, L.; Fitch, A.; Unnasch, T.R.; Popov, V.L.; Da Rosa, A.P.A.T.; Guzman, H.; Carrera, J.-P.; et al. Characterization of five unclassified orthobunyaviruses (Bunyaviridae) from Africa and the Americas. J. Gen. Virol. 2017, 98, 2258–2266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eldridge, B.F.; Lanzaro, G.C.; Campbell, G.L.; Reeves, W.C.; Hardy, J.L. Occurrence and Evolutionary Significance of a California Encephalitis-Like Virus in Aedes squamiger (Diptera: Culicidae). J. Med. Èntomol. 1991, 28, 645–651. [Google Scholar] [CrossRef]
- Huang, C.; Shope, R.E.; Spargo, B.; Campbell, W.P. The S RNA genomic sequences of Inkoo, San Angelo, Serra do Navio, South River and Tahyna bunyaviruses. J. Gen. Virol. 1996, 77, 1761–1768. [Google Scholar] [CrossRef]
- Fulhorst, C.F.; Bowen, M.D.; Hardy, J.L.; Eldridge, B.F.; Chiles, R.E.; Jackson, A.O.; Reeves, W.C. Geographic Distribution and Serologic and Genomic Characterization of Morro Bay Virus, a Newly Recognized Bunyavirus. Am. J. Trop. Med. Hyg. 1996, 54, 563–569. [Google Scholar] [CrossRef]
- Hughes, H.R.; Lanciotti, R.S.; Blair, C.D.; Lambert, A.J. Full genomic characterization of California serogroup viruses, genus Orthobunyavirus, family Peribunyaviridae including phylogenetic relationships. Virology 2017, 512, 201–210. [Google Scholar] [CrossRef]
- Blitvich, B.J.; Staley, M.; Loroño-Pino, M.A.; García-Rejón, J.E.; Farfan-Ale, J.A.; Dorman, K.S. Identification of a novel subtype of South River virus (family Bunyaviridae). Arch. Virol. 2012, 157, 1205–1209. [Google Scholar] [CrossRef] [Green Version]
- Campbell, W.P.; Huang, C. Sequence comparisons of medium RNA segment among 15 California serogroup viruses. Virus Res. 1999, 61, 137–144. [Google Scholar] [CrossRef]
- Evans, A.B.; Winkler, C.W.; Peterson, K.E. Differences in Neuropathogenesis of Encephalitic California Serogroup Viruses. Emerg. Infect. Dis. 2019, 25, 728–738. [Google Scholar] [CrossRef] [PubMed]
- ArboNET, C. ArboNET Diseases Map. Available online: https://wwwn.cdc.gov/arbonet/maps/ADB_Diseases_Map/index.html (accessed on 3 June 2019).
- Drebot, M. Emerging mosquito-borne bunyaviruses in Canada. Can. Commun. Dis. Rep. 2015, 41, 117–123. [Google Scholar] [CrossRef] [PubMed]
- Pastula, D.M.; Hoang Johnson, D.K.; White, J.L.; Dupuis, A.P., 2nd; Fischer, M.; Staples, J.E. Jamestown Canyon Virus Disease in the United States-2000–2013. Am. J. Trop. Med. Hyg. 2015, 93, 384–389. [Google Scholar] [CrossRef] [PubMed]
- Kosoy, O.; Rabe, I.; Geissler, A.; Adjemian, J.; Panella, A.; Laven, J.; Basile, A.J.; Velez, J.; Griffith, K.; Wong, D.; et al. Serological Survey for Antibodies to Mosquito-Borne Bunyaviruses Among US National Park Service and US Forest Service Employees. Vector-Borne Zoonotic Dis. 2016, 16, 191–198. [Google Scholar] [CrossRef] [PubMed]
- Anderson, J.F.; Andreadis, T.G.; Ferrandino, F.J.; Main, A.J.; Armstrong, P.M. Arboviruses in North Dakota, 2003–2006. Am. J. Trop. Med. Hyg. 2015, 92, 377–393. [Google Scholar] [CrossRef] [PubMed]
- Andrews, W.N.; Rowley, W.A.; Wong, Y.W.; Dorsey, D.C.; Hausler, W.J. Isolation of Trivittatus Virus from Larvae and Adults Reared from Field-Collected Larvae of Aedes Trivittatus (Diptera: Gulicidae). J. Med. Èntomol. 1977, 13, 699–701. [Google Scholar] [CrossRef] [PubMed]
- CDC. Arbovirus Catalog: San Angelo Virus. Available online: https://wwwn.cdc.gov/Arbocat/VirusDetails.aspx?ID=414&SID=12 (accessed on 3 June 2019).
- Berry, R.L.; Weigert, B.J.L.; Calisher, C.H.; Parsons, M.A.; Bear, G.T. Evidence for Transovarial Transmission of Jamestown Canyon Virus in Ohio. Mosq. News 1977, 37, 494–496. [Google Scholar]
- Boromisa, R.D.; Grimstad, P.R. Virus-Vector-Host Relationships of Aedes stimulans and Jamestown Canyon Virus in a Northern Indiana Enzootic Focus. Am. J. Trop. Med. Hyg. 1986, 35, 1285–1295. [Google Scholar] [CrossRef]
- Campbell, G.L.; Eldridge, B.F.; Hardy, J.L.; Reeves, W.C.; Jessup, D.A.; Presser, S.B. Prevalence of Neutralizing Antibodies Against California and Bunyamwera Serogroup Viruses in Deer from Mountainous Areas of California. Am. J. Trop. Med. Hyg. 1989, 40, 428–437. [Google Scholar] [CrossRef]
- Clark, G.G.; Crabbs, C.; Watts, D.M.; Bailey, C.L. An Ecological Study of Jamestown Canyon Virus on the Delmarva Peninsula, with Emphasis on its Possible Vector. J. Med. Entomol. 1986, 23, 588–599. [Google Scholar] [CrossRef]
- Eldridge, B.F.; Calisher, C.H.; Fryer, J.L.; Bright, L.; Hobbs, D.J. Serological Evidence of California Serogroup Virus Activity in Oregon. J. Wildl. Dis. 1987, 23, 199–204. [Google Scholar] [CrossRef] [PubMed]
- Farfan-Ale, J.A.; Loroño-Pino, M.A.; García-Rejón, J.E.; Soto, V.; Lin, M.; Staley, M.; Dorman, K.S.; Bartholomay, L.C.; Hovav, E.; Blitvich, B.J. Detection of Flaviviruses and Orthobunyaviruses in Mosquitoes in the Yucatan Peninsula of Mexico in 2008. Vector-Borne Zoonotic Dis. 2010, 10, 777–783. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haddow, A.D.; Odoi, A. The Incidence Risk, Clustering, and Clinical Presentation of La Crosse Virus Infections in the Eastern United States, 2003–2007. PLoS ONE 2009, 4, e6145. [Google Scholar] [CrossRef] [PubMed]
- Heard, P.B.; Zhang, M.B.; Grimstad, P.R. Isolation of Jamestown Canyon virus (California serogroup) from Aedes mosquitoes in an enzootic focus in Michigan. J. Am. Mosq. Control Assoc. 1990, 6, 461–468. [Google Scholar] [PubMed]
- Issel, C.J.; Trainer, D.O.; Thompson, W.H. Serologic Evidence of Infections of White-Tailed Deer in Wisconsin with Three California Group Arboviruses (La Crosse, Trivittatus, and Jamestown Canyon). Am. J. Trop. Med. Hyg. 1972, 21, 985–988. [Google Scholar] [CrossRef] [PubMed]
- Issel, C.J. Isolation of Jamestown Canyon Virus (a California Group Arbovirus) from a White-Tailed Deer. Am. J. Trop. Med. Hyg. 1973, 22, 414–417. [Google Scholar] [CrossRef] [PubMed]
- Le Duc, J.W.; Suyemoto, W.; Eldridge, B.F.; Russell, P.K.; Barr, A.R. Ecology of California Encephalitis Viruses on the Del Mar Va Peninsula. Am. J. Trop. Med. Hyg. 1975, 24, 124–126. [Google Scholar] [CrossRef]
- Lowell, J.; Higgins, D.P.; Drebot, M.; Makowski, K.; Staples, J.E. Human Jamestown Canyon Virus Infection-Montana, 2009 (Reprinted from MMWR, vol 60, pg 652–655, 2011). J. Am. Med. Assoc. 2011, 306, 371–376. [Google Scholar]
- Pinger, R.R.; Rowley, W.A.; Wong, Y.W.; Dorsey, D.C. Trivittatus Virus Infections in Wild Mammals and Sentinel Rabbits in Central Iowa. Am. J. Trop. Med. Hyg. 1975, 24, 1006–1009. [Google Scholar] [CrossRef]
- Roberts, D.R.; Scanlon, J.E. The ecology and behavior of Aedes atlanticus D. & K. and other species with reference to Keystone virus in the Houston area, Texas. J. Med. Entomol. 1975, 12, 537–546. [Google Scholar] [CrossRef]
- Srihongse, S.; Grayson, M.A.; Deibel, R. California Serogroup Viruses in New York State: The Role of Subtypes in Human Infections. Am. J. Trop. Med. Hyg. 1984, 33, 1218–1227. [Google Scholar] [CrossRef] [PubMed]
- Trainer, D.O.; Knowlton, F.F. Serologic Evidence of Diseases in Texas Coyotes. J. Wildl. Manag. 1968, 32, 981–983. [Google Scholar] [CrossRef]
- Rhyan, J.; Tyers, D.; Zimmer, J.; Lewandowski, K.; Hennager, S.; Young, J.; Pappert, R.; Panella, A.; Kosoy, O. Serologic Survey of Snowshoe Hares (Lepus americanus) in the Greater Yellowstone Area for Brucellosis, Tularemia, and Snowshoe Hare Virus. J. Wildl. Dis. 2015, 51, 769–773. [Google Scholar] [CrossRef] [PubMed]
- Walker, E.D.; Grayson, M.A.; Edman, J.D. Isolation of Jamestown Canyon and snowshoe hare viruses (California serogroup) from Aedes mosquitoes in western Massachusetts. J. Am. Mosq. Control Assoc. 1993, 9, 131–134. [Google Scholar] [PubMed]
- Walters, L.L.; Tirrell, S.J.; Shope, R.E. Seroepidemiology of California and Bunyamwera serogroup (Bunyaviridae) virus infections in native populations of Alaska. Am. J. Trop. Med. Hyg. 1999, 60, 806–821. [Google Scholar] [CrossRef] [PubMed]
- Wills, W.; Pidcoe, V.; Carroll, D.F.; Satz, J.E. Isolation of California Group Arboviruses from Pennsylvania Mosquitos—1971, 1972. Mosq. News 1974, 34, 376–381. [Google Scholar]
- Artsob, H.; Spence, L.; Th’Ng, C.; Lampotang, V.; Johnston, D.; MacInnes, C.; Matejka, F.; Voigt, D.; Watt, I. Arbovirus infections in several Ontario mammals, 1975–1980. Can. J. Veter. Res. 1986, 50, 42–46. [Google Scholar]
- Blitvich, B.J.; Saiyasombat, R.; Da Rosa, A.T.; Tesh, R.B.; Calisher, C.H.; García-Rejón, J.E.; Farfan-Ale, J.A.; Loroño, R.E.; Bates, A.; Loroño-Pino, M.A. Orthobunyaviruses, a Common Cause of Infection of Livestock in the Yucatan Peninsula of Mexico. Am. J. Trop. Med. Hyg. 2012, 87, 1132–1139. [Google Scholar] [CrossRef] [Green Version]
- Carvalho, V.L.; Nunes, M.R.T.; Da Silva, E.V.P.; Vieira, C.M.A.; Gomes, M.; Casseb, S.M.; Rodrigues, S.G.; Nunes-Neto, J.P.; Quaresma, J.A.O.; Vasconcelos, P.F.C.; et al. Genetic characterization of orthobunyavirus Melao, strains BE AR633512 and BE AR8033, and experimental infection in golden hamsters (Mesocricetus auratus). J. Gen. Virol. 2009, 90, 223–233. [Google Scholar] [CrossRef]
- CDC. Arbovirus Catalog: Serra do Navio Virus. Available online: https://wwwn.cdc.gov/Arbocat/VirusDetails.aspx?ID=435&SID=3 (accessed on 3 June 2019).
- Hubálek, Z. Mosquito-borne viruses in Europe. Parasitol. Res. 2008, 103, 29–43. [Google Scholar] [CrossRef]
- Putkuri, N.; Vaheri, A.; Vapalahti, O. Prevalence and Protein Specificity of Human Antibodies to Inkoo Virus Infection. Clin. Vaccine Immunol. 2007, 14, 1555–1562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Putkuri, N.; Kantele, A.; Levanov, L.; Kivistö, I.; Brummer-Korvenkontio, M.; Vaheri, A.; Vapalahti, O. Acute Human Inkoo and Chatanga Virus Infections, Finland. Emerg. Infect. Dis. 2016, 22, 810–817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lwande, O.W.; Bucht, G.; Ahlm, C.; Ahlm, K.; Näslund, J.; Evander, M. Mosquito-borne Inkoo virus in northern Sweden—Isolation and whole genome sequencing. Virol. J. 2017, 14, 61. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Fu, S.; Tian, Z.; Lu, Z.; He, Y.; Wang, H.; Wang, J.; Guo, W.; Tao, B.; Liang, G. Distribution of Mosquitoes and Mosquito-Borne Arboviruses in Inner Mongolia, China. Vector-Borne Zoonotic Dis. 2011, 11, 1577–1581. [Google Scholar] [CrossRef] [PubMed]
- Chastel, C.; Bach-Hamba, D.; Launay, H.; Le Lay, G.; Hellal, H.; Beaucournu, J.C. Arbovirus infections in Tunisia: New serological survey of small wild mammals. Bull. Soc. Pathol. Exot. Fil. 1983, 76, 21–33. [Google Scholar]
- Jentes, E.S.; Robinson, J.; Johnson, B.W.; Conde, I.; Sakouvougui, Y.; Iverson, J.; Beecher, S.; Bah, M.A.; Diakite, F.; Coulibaly, M.; et al. Acute Arboviral Infections in Guinea, West Africa, 2006. Am. J. Trop. Med. Hyg. 2010, 83, 388–394. [Google Scholar] [CrossRef] [Green Version]
- Kuniholm, M.H.; Wolfe, N.D.; Tamoufe, U.; Gubler, D.J.; Huang, C.Y.-H.; Burke, D.S.; Mpoudi-Ngole, E. Seroprevalence and distribution of flaviviridae, togaviridae, and bunyaviridae arboviral infections in rural cameroonian adults. Am. J. Trop. Med. Hyg. 2006, 74, 1078–1083. [Google Scholar] [CrossRef]
- Kunz, C.; Buckley, S.M.; Casals, J. Antibodies in Man against Tahyna and Lumbo Viruses Determined by Hemagglutination-Inhibition and Tissue-Culture Neutralization Tests. Am. J. Trop. Med. Hyg. 1964, 13, 738–741. [Google Scholar] [CrossRef]
- Lu, Z.; Fu, S.-H.; Wang, F.-T.; Nasci, R.S.; Tang, Q.; Liang, G.-D. Circulation of Diverse Genotypes of Tahyna Virus in Xinjiang, People’s Republic of China. Am. J. Trop. Med. Hyg. 2011, 85, 442–445. [Google Scholar] [CrossRef]
- Wilkerson, R.C.; Linton, Y.-M.; Fonseca, D.M.; Schultz, T.R.; Price, D.C.; Strickman, D.A. Making Mosquito Taxonomy Useful: A Stable Classification of Tribe Aedini that Balances Utility with Current Knowledge of Evolutionary Relationships. PLoS ONE 2015, 10, e0133602. [Google Scholar] [CrossRef]
- Watts, D.M.; Morris, C.D.; Wright, R.E.; DeFoliart, G.R.; Hanson, R.P. Transmission of Lacrosse Virus (California Encephalitis Group) by the Mosquito Aedes Triseriatus. J. Med. Entomol. 1972, 9, 125–127. [Google Scholar] [CrossRef] [PubMed]
- Pantuwatana, S.; Thompson, W.H.; Watts, D.M.; Hanson, R.P. Experimental Infection of Chipmunks and Squirrels with La Crosse and Trivittatus Viruses and Biological Transmission of La Crosse Virus by Aedes Triseriatus. Am. J. Trop. Med. Hyg. 1972, 21, 476–481. [Google Scholar] [CrossRef] [PubMed]
- Taylor, D.J.; Lewis, A.L.; Edman, J.D.; Jennings, W.L. California group arboviruses in Florida. Host-vector relations. Am. J. Trop. Med. Hyg. 1971, 20, 139–145. [Google Scholar] [CrossRef] [PubMed]
- Watts, D.M.; Bailey, C.L.; Roberts, N.T.; Tammariello, R.F.; Dalrymple, J.M.; Clark, G.C. Maintenance and Transmission of Keystone Virus by Aedes atlanticus (Diptera: Culicidae) and the Gray Squirrel in the Pocomoke Cypress Swamp, Maryland. J. Med. Entomol. 1988, 25, 493–500. [Google Scholar] [CrossRef] [PubMed]
- Watts, D.M.; DeFoliart, G.R.; Yuill, T.M. Experimental Transmission of Trivittatus Virus (California Virus Group) by Aedes Trivittatus. Am. J. Trop. Med. Hyg. 1976, 25, 173–176. [Google Scholar] [CrossRef]
- Kramer, L.D.; Reeves, W.C.; Hardy, J.L.; Presser, S.B.; Eldridge, B.F.; Bowen, M.D. Vector Competence of California Mosquitoes for California Encephalitis and California Encephalitis-Like Viruses. Am. J. Trop. Med. Hyg. 1992, 47, 562–573. [Google Scholar] [CrossRef] [PubMed]
- Hammon, W.M.; Reeves, W.C. California encephalitis virus, a newly described agent. III. Mosquito infection and transmission. J. Immunol. 1952, 69, 511–514. [Google Scholar]
- Turell, M.J.; Hardy, J.L.; Reeves, W.C. Stabilized Infection of California Encephalitis Virus in Aedes dorsalis, and its Implications for Viral Maintenance in Nature. Am. J. Trop. Med. Hyg. 1982, 31, 1252–1259. [Google Scholar] [CrossRef]
- Berry, R.L.; Parsons, M.A.; Lalondeweigert, B.J.; Lebio, J.; Stegmiller, H.; Bear, G.T. Aedes-Canadensis, a Vector of Lacrosse Virus (California Serogroup) in Ohio. J. Am. Mosq. Control 1986, 2, 73–78. [Google Scholar]
- Westby, K.M.; Fritzen, C.; Paulsen, D.; Poindexter, S.; Moncayo, A.C. La Crosse Encephalitis Virus Infection in Field-Collected Aedes albopictus, Aedes japonicus, and Aedes triseriatus in Tennessee. J. Am. Mosq. Control Assoc. 2015, 31, 233–241. [Google Scholar] [CrossRef]
- Andreadis, T.G.; Anderson, J.F.; Armstrong, P.M.; Main, A.J. Isolations of Jamestown Canyon Virus (Bunyaviridae: Orthobunyavirus) from Field-Collected Mosquitoes (Diptera: Culicidae) in Connecticut, USA: A Ten-Year Analysis, 1997–2006. Vector-Borne Zoonotic Dis. 2008, 8, 175–188. [Google Scholar] [CrossRef] [PubMed]
- DeFoliart, G.R.; Anslow, R.O.; Hanson, R.P.; Morris, C.D.; Papadopoulos, O.; Sather, G.E. Isolation of Jamestown Canyon Serotype of California Encephalitis Virus from Naturally Infected Aedes Mosquitoes and Tabanids. Am. J. Trop. Med. Hyg. 1969, 18, 440–447. [Google Scholar] [CrossRef] [PubMed]
- Grimstad, P.R.; Kobayashi, J.F.; Zhang, M.B.; Craig, G.B. Recently introduced Aedes albopictus in the United States: Potential vector of La Crosse virus (Bunyaviridae: California serogroup). J. Am. Mosq. Control Assoc. 1989, 5, 422–427. [Google Scholar]
- Boromisa, R.D.; Grayson, M.A. Oral transmission of Jamestown Canyon virus by Aedes provocans mosquitoes from northeastern New York. J. Am. Mosq. Control Assoc. 1991, 7, 42–47. [Google Scholar] [PubMed]
- Heard, P.B.; Zhang, M.B.; Grimstad, P.R. Laboratory transmission of Jamestown Canyon and snowshoe hare viruses (Bunyaviridae: California serogroup) by several species of mosquitoes. J. Am. Mosq. Control Assoc. 1991, 7, 94–102. [Google Scholar] [PubMed]
- Kramer, L.D.; Bowen, M.D.; Hardy, J.L.; Reeves, W.C.; Presser, S.B.; Eldridge, B.F. Vector Competence of Alpine, Central Valley, and Coastal Mosquitoes (Diptera: Culicidae) from California for Jamestown Canyon Virus. J. Med. Entomol. 1993, 30, 398–406. [Google Scholar] [CrossRef] [PubMed]
- Iversen, J.; Papadopoulos, O.; Morris, C.V.; Hanson, R.P.; DeFoliart, G.R. Isolation of Viruses of the California Encephalitis Virus Group from Boreal Aedes Mosquitoes. Am. J. Trop. Med. Hyg. 1969, 18, 735–742. [Google Scholar] [CrossRef]
- McLean, D.M.; Bergman, S.K.A.; Gould, A.P.; Grass, P.N.; Miller, M.A.; Spratt, E.E. California Encephalitis Virus Prevalence throughout the Yukon Territory, 1971. Am. J. Trop. Med. Hyg. 1975, 24, 676–684. [Google Scholar] [CrossRef]
- Hewlett, M.J.; Clerx, J.P.M.; Haaster, C.M.C.-V.; Chandler, L.J.; McLean, D.M.; Beaty, B.J. Genomic and Biologic Analyses of Snowshoe Hare Virus Field and Laboratory Strains. Am. J. Trop. Med. Hyg. 1992, 46, 524–532. [Google Scholar] [CrossRef]
- Francy, D.B.; Jaenson, T.G.T.; Lundström, J.O.; Schildt, E.-B.; Espmark, Å.; Henriksson, B.; Niklasson, B. Ecologic Studies of Mosquitoes and Birds as Hosts of Ockelbo Virus in Sweden and Isolation of Inkoo and Batai Viruses from Mosquitoes. Am. J. Trop. Med. Hyg. 1989, 41, 355–363. [Google Scholar] [CrossRef]
- Kilian, P.; Valdés, J.J.; Lecina-Casas, D.; Chrudimský, T.; Růžek, D. The variability of the large genomic segment of Ťahyňa orthobunyavirus and an all-atom exploration of its anti-viral drug resistance. Infect. Genet. Evol. 2013, 20, 304–311. [Google Scholar] [CrossRef] [PubMed]
- Betasova, L.; Blazejova, H.; Venclikova, K.; Hubalek, Z.; Sebesta, O.; Pesko, J.; Rudolf, I. Isolation of Tahyna Virus (California Encephalitis Group) From Anopheles hyrcanus (Diptera, Culicidae), a Mosquito Species New to, and Expanding in, Central Europe. J. Med. Entomol. 2014, 51, 1264–1267. [Google Scholar]
- Calisher, C.H.; Lazuick, J.S.; Cropp, C.B.; Mitchell, C.J.; Tesh, R.B.; Sabattini, M.S.; Monath, T.P. A Newly Recognized Vesiculovirus, Calchaqui Virus, and Subtypes of Melao and Maguari Viruses from Argentina, with Serologic Evidence for Infections of Humans and Horses. Am. J. Trop. Med. Hyg. 1987, 36, 114–119. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, C.J.; Daffner, J.F.; Monath, T.P.; Jakob, W.L.; Calisher, C.H.; Cropp, C.B.; Sabattini, M.S.; Darsie, R.F., Jr. Arbovirus Isolations from Mosquitoes Collected during and after the 1982–1983 Epizootic of Western Equine Encephalitis in Argentina. Am. J. Trop. Med. Hyg. 1987, 36, 107–113. [Google Scholar] [CrossRef] [PubMed]
- Watts, D.M.; Pantuwatana, S.; DeFoliart, G.R.; Yuill, T.M.; Thompson, W.H. Transovarial Transmission of LaCrosse Virus (California Encephalitis Group) in the Mosquito, Aedes triseriatus. Science 1973, 182, 1140–1141. [Google Scholar] [CrossRef] [PubMed]
- Miller, B.R.; DeFoliart, G.R.; Yuill, T.M. Vertical Transmission of La Crosse Virus (California Encephalitis Group): Transovarial and Filial Infection Rates in Aedes Triseriatus (Diptera: Culicidae). J. Med. Entomol. 1977, 14, 437–440. [Google Scholar] [CrossRef] [PubMed]
- Turell, M.J.; Reeves, W.C.; Hardy, J.L. Transovarial and Trans-Stadial Transmission of California Encephalitis Virus in Aedes Dorsalis and Aedes Melanimon. Am. J. Trop. Med. Hyg. 1982, 31, 1021–1029. [Google Scholar] [CrossRef]
- Danielova, V.; Ryba, J. Laboratory Demonstration of Transovarial Transmission of Tahyna Virus in Aedes Vexans and the Role of This Mechanism in Overwintering of This Arbovirus. Folia Parasitol. 1979, 26, 361–366. [Google Scholar]
- LeDuc, J.W.; Burger, J.F.; Eldridge, B.F.; Russell, P.K. Ecology of keystone virus, a transovarially maintained arbovirus. Ann. N. Y. Acad. Sci. 1975, 266, 144–151. [Google Scholar] [CrossRef]
- Christensen, B.M.; Rowley, W.A.; Dorsey, D.C.; Wong, Y.W.; Hausler, W.J., Jr. Laboratory Studies of Transovarial Transmission of Trivittatus Virus by Aedes Trivittatus. Am. J. Trop. Med. Hyg. 1978, 27, 184–186. [Google Scholar] [CrossRef]
- Crane, G.T.; Elbel, R.E. Transovarial Transmission of California Encephalitis-Virus in Mosquito Aedes-Dorsalis at Blue Lake, Utah. Mosq. News 1977, 37, 479–482. [Google Scholar]
- Mclintock, J.; Curry, P.S.; Wagner, R.J.; Leung, M.K.; Iversen, J.O. Isolation of Snowshoe Hare Virus from Aedes-Implicatus Larvae in Saskatchewan. Mosq. News 1976, 36, 233–237. [Google Scholar]
- Tesh, R.B.; Shroyer, D.A. The Mechanism of Arbovirus Transovarial Transmission in Mosquitoes: San Angelo Virus in Aedes Albopictus. Am. J. Trop. Med. Hyg. 1980, 29, 1394–1404. [Google Scholar] [CrossRef] [PubMed]
- Tingström, O.; Lwande, O.W.; Näslund, J.; Spyckerelle, I.; Engdahl, C.; Von Schoenberg, P.; Ahlm, C.; Evander, M.; Bucht, G. Detection of Sindbis and Inkoo Virus RNA in Genetically Typed Mosquito Larvae Sampled in Northern Sweden. Vector-Borne Zoonotic Dis. 2016, 16, 461–467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moulton, D.W.; Thompson, W.H. California group virus infections in small, forest-dwelling mammals of Wisconsin. Some ecological considerations. Am. J. Trop. Med. Hyg. 1971, 20, 474–482. [Google Scholar] [CrossRef] [PubMed]
- Newhouse, V.F.; Burgdorfer, W.; Corwin, D. Field and Laboratory Studies on Hosts and Vectors of Snowshoe Hare Strain of California Virus. Mosq. News 1971, 31, 401–408. [Google Scholar]
- LeDuc, J.W. Natural Transmission of Keystone Virus to Sentinel Rabbits on the Delmarva Peninsula. Am. J. Trop. Med. Hyg. 1978, 27, 1041–1044. [Google Scholar] [CrossRef]
- Issel, C.J.; Trainer, D.O.; Thompson, W.H. Experimental Studies with White-Tailed Deer and Four California Group Arboviruses (La Crosse, Trivittatus, Snowshoe Hare, and Jamestown Canyon). Am. J. Trop. Med. Hyg. 1972, 21, 979–984. [Google Scholar] [CrossRef]
- Zarnke, R.L.; Calisher, C.H.; Kerschner, J. Serologic evidence of arbovirus infections in humans and wild animals in Alaska. J. Wildl. Dis. 1983, 19, 175–179. [Google Scholar] [CrossRef]
- Goff, G.; Whitney, H.; Drebot, M.A. Roles of Host Species, Geographic Separation, and Isolation in the Seroprevalence of Jamestown Canyon and Snowshoe Hare Viruses in Newfoundland. Appl. Environ. Microbiol. 2012, 78, 6734–6740. [Google Scholar] [CrossRef] [Green Version]
- McFarlane, B.L.; Embree, J.E.; Embil, J.A.; Rozee, K.R.; Artsob, H. Antibodies to the California group of arboviruses in animal populations of New Brunswick. Can. J. Microbiol. 1982, 28, 200–204. [Google Scholar] [CrossRef] [PubMed]
- McFarlane, B.L.; Embil, J.A.; Artsob, H.; Spence, L.; Rozee, K.R. Antibodies to the California group of arboviruses in the moose (Alces alces americana Clinton) population of Nova Scotia. Can. J. Microbiol. 1981, 27, 1219–1223. [Google Scholar] [CrossRef] [PubMed]
- Elliott, R.M. Orthobunyaviruses: Recent genetic and structural insights. Nat. Rev. Genet. 2014, 12, 673–685. [Google Scholar] [CrossRef] [PubMed]
- Barr, J.N.; Elliott, R.M.; Dunn, E.F.; Wertz, G.W. Segment-specific terminal sequences of Bunyamwera bunyavirus regulate genome replication. Virology 2003, 311, 326–338. [Google Scholar] [CrossRef] [Green Version]
- Barr, J.N.; Wertz, G.W. Role of the Conserved Nucleotide Mismatch within 3′- and 5′-Terminal Regions of Bunyamwera Virus in Signaling Transcription. J. Virol. 2005, 79, 3586–3594. [Google Scholar] [CrossRef] [PubMed]
- Kohl, A.; Lowen, A.C.; Leonard, V.H.J.; Elliott, R.M. Genetic elements regulating packaging of the Bunyamwera orthobunyavirus genome. J. Gen. Virol. 2006, 87, 177–187. [Google Scholar] [CrossRef] [PubMed]
- Osborne, J.C.; Elliott, R.M. RNA Binding Properties of Bunyamwera Virus Nucleocapsid Protein and Selective Binding to an Element in the 5′ Terminus of the Negative-Sense S Segment. J. Virol. 2000, 74, 9946–9952. [Google Scholar] [CrossRef] [PubMed]
- Reguera, J.; Malet, H.; Weber, F.; Cusack, S. Structural basis for encapsidation of genomic RNA by La Crosse Orthobunyavirus nucleoprotein. Proc. Natl. Acad. Sci. USA 2013, 110, 7246–7251. [Google Scholar] [CrossRef] [Green Version]
- Ogg, M.M.; Patterson, J.L. RNA Binding Domain of Jamestown Canyon Virus S Segment RNAs. J. Virol. 2007, 81, 13754–13760. [Google Scholar] [CrossRef] [Green Version]
- Endres, M.J.; Jacoby, D.R.; Janssen, R.S.; Gonzalez-Scarano, F.; Nathanson, N. The Large Viral RNA Segment of California Serogroup Bunyaviruses Encodes the Large Viral Protein. J. Gen. Virol. 1989, 70, 223–228. [Google Scholar] [CrossRef]
- Gentsch, J.R.; Bishop, D.L. M viral RNA segment of bunyaviruses codes for two glycoproteins, G1 and G2. J. Virol. 1979, 30, 767–770. [Google Scholar] [PubMed]
- Fuller, F.; Bishop, D.H. Identification of virus-coded nonstructural polypeptides in bunyavirus-infected cells. J. Virol. 1982, 41, 643–648. [Google Scholar] [PubMed]
- Gentsch, J.R.; Bishop, D.H. Small viral RNA segment of bunyaviruses codes for viral nucleocapsid protein. J. Virol. 1978, 28, 417–419. [Google Scholar] [PubMed]
- Fazakerley, J.; Gonzalezscarano, F.; Strickler, J.; Dietzschold, B.; Karush, F.; Nathanson, N. Organization of the middle RNA segment of snowshoe hare bunyavirus. Virology 1988, 167, 422–432. [Google Scholar] [CrossRef]
- Shi, X.; Botting, C.H.; Li, P.; Niglas, M.; Brennan, B.; Shirran, S.L.; Szemiel, A.M.; Elliott, R.M. Bunyamwera orthobunyavirus glycoprotein precursor is processed by cellular signal peptidase and signal peptide peptidase. Proc. Natl. Acad. Sci. USA 2016, 113, 8825–8830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuller, F.; Bhown, A.S.; Bishop, D.H. Bunyavirus nucleoprotein, N, and a non-structural protein, NSS, are coded by overlapping reading frames in the S RNA. J. Gen. Virol. 1983, 64, 1705–1714. [Google Scholar] [CrossRef] [PubMed]
- Bowen, M.D.; Jackson, A.O.; Bruns, T.D.; Hacker, D.L.; Hardy, J.L. Determination and comparative analysis of the small RNA genomic sequences of California encephalitis, Jamestown Canyon, Jerry Slough, Melao, Keystone and Trivittatus viruses (Bunyaviridae, genus Bunyavirus, California serogroup). J. Gen. Virol. 1995, 76, 559–572. [Google Scholar] [CrossRef] [PubMed]
- Bupp, K.; Stillmock, K.; Gonzalez-Scarano, F. Analysis of the Intracellular Transport Properties of Recombinant La Crosse Virus Glycoproteins. Virology 1996, 220, 485–490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hacker, J.K.; Hardy, J.L. Adsorptive Endocytosis of California Encephalitis Virus into Mosquito and Mammalian Cells: A Role for G1. Virology 1997, 235, 40–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hofmann, H.; Li, X.; Zhang, X.; Liu, W.; Kühl, A.; Kaup, F.; Soldan, S.S.; González-Scarano, F.; Weber, F.; He, Y.; et al. Severe Fever with Thrombocytopenia Virus Glycoproteins Are Targeted by Neutralizing Antibodies and Can Use DC-SIGN as a Receptor for pH-Dependent Entry into Human and Animal Cell Lines. J. Virol. 2013, 87, 4384–4394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia-Vallejo, J.J.; Bloem, K.; Knippels, L.M.; Garssen, J.; van Vliet, S.J.; van Kooyk, Y. The Consequences of Multiple Simultaneous C-Type Lectin-Ligand Interactions: DCIR Alters the Endo-Lysosomal Routing of DC-SIGN. Front. Immunol. 2015, 6, 87. [Google Scholar] [CrossRef] [PubMed]
- Bennett, R.S.; Cress, C.M.; Ward, J.M.; Firestone, C.-Y.; Murphy, B.R.; Whitehead, S.S. La Crosse virus infectivity, pathogenesis, and immunogenicity in mice and monkeys. Virol. J. 2008, 5, 25. [Google Scholar] [CrossRef] [PubMed]
- Murakami, S.; Takenaka-Uema, A.; Kobayashi, T.; Kato, K.; Shimojima, M.; Palmarini, M.; Horimoto, T. Heparan Sulfate Proteoglycan Is an Important Attachment Factor for Cell Entry of Akabane and Schmallenberg Viruses. J. Virol. 2017, 91. [Google Scholar] [CrossRef] [PubMed]
- Hollidge, B.S.; Nedelsky, N.B.; Salzano, M.-V.; Fraser, J.W.; González-Scarano, F.; Soldan, S.S. Orthobunyavirus Entry into Neurons and Other Mammalian Cells Occurs via Clathrin-Mediated Endocytosis and Requires Trafficking into Early Endosomes. J. Virol. 2012, 86, 7988–8001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plassmeyer, M.L.; Soldan, S.S.; Stachelek, K.M.; Martín-García, J.; Gonzalez-Scarano, F. California serogroup Gc (G1) glycoprotein is the principal determinant of pH-dependent cell fusion and entry. Virology 2005, 338, 121–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plassmeyer, M.L.; Soldan, S.S.; Stachelek, K.M.; Roth, S.M.; Martin-Garcia, J.; Gonzalez-Scarano, F. Mutagenesis of the La Crosse Virus glycoprotein supports a role for Gc (1066–1087) as the fusion peptide. Virology 2007, 358, 273–282. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Scarano, F. La Crosse virus G1 glycoprotein undergoes a conformational change at the pH of fusion. Virology 1985, 140, 209–216. [Google Scholar] [CrossRef]
- Pekosz, A.; Gonzalez-Scarano, F. The Extracellular Domain of La Crosse Virus G1 Forms Oligomers and Undergoes pH-Dependent Conformational Changes. Virology 1996, 225, 243–247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reguera, J.; Weber, F.; Cusack, S. Bunyaviridae RNA Polymerases (L-Protein) Have an N-Terminal, Influenza-Like Endonuclease Domain, Essential for Viral Cap-Dependent Transcription. PLoS Pathog. 2010, 6, e1001101. [Google Scholar] [CrossRef]
- Reguera, J.; Gerlach, P.; Rosenthal, M.; Gaudon, S.; Coscia, F.; Günther, S.; Cusack, S. Comparative Structural and Functional Analysis of Bunyavirus and Arenavirus Cap-Snatching Endonucleases. PLoS Pathog. 2016, 12, e1005636. [Google Scholar] [CrossRef]
- Bellocq, C.; Kolakofsky, D. Translational requirement for La Crosse virus S-mRNA synthesis: A possible mechanism. J. Virol. 1987, 61, 3960–3967. [Google Scholar] [PubMed]
- Kolakofsky, D.; Bellocq, C.; Raju, R. The Translational Requirement for La Crosse Virus S-mRNA Synthesis. Cold Spring Harb. Symp. Quant. Biol. 1987, 52, 373–379. [Google Scholar] [CrossRef] [PubMed]
- Barr, J.N. Bunyavirus mRNA synthesis is coupled to translation to prevent premature transcription termination. RNA 2007, 13, 731–736. [Google Scholar] [CrossRef] [PubMed]
- Fontana, J.; Elliott, R.M.; Fernández, J.J.; Risco, C.; López-Montero, N.; Lopez-Montero, N.; López-Montero, N. The unique architecture of Bunyamwera virus factories around the Golgi complex. Cell. Microbiol. 2008, 10, 2012–2028. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, N.S.; Mendonça, L.R.; Dias, M.V.S.; Pontelli, M.C.; Da Silva, E.Z.M.; Criado, M.F.; Da Silva-Januário, M.E.; Schindler, M.; Jamur, M.C.; Oliver, C.; et al. ESCRT machinery components are required for Orthobunyavirus particle production in Golgi compartments. PLoS Pathog. 2018, 14, e1007047. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; Kohl, A.; Li, P.; Elliott, R.M. Role of the Cytoplasmic Tail Domains of Bunyamwera Orthobunyavirus Glycoproteins Gn and Gc in Virus Assembly and Morphogenesis. J. Virol. 2007, 81, 10151–10160. [Google Scholar] [CrossRef] [Green Version]
- Shi, X.; Van Mierlo, J.T.; French, A.; Elliott, R.M. Visualizing the Replication Cycle of Bunyamwera Orthobunyavirus Expressing Fluorescent Protein-Tagged Gc Glycoprotein. J. Virol. 2010, 84, 8460–8469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salanueva, I.J.; Novoa, R.R.; Cabezas, P.; López-Iglesias, C.; Carrascosa, J.L.; Elliott, R.M.; Risco, C. Polymorphism and Structural Maturation of Bunyamwera Virus in Golgi and Post-Golgi Compartments. J. Virol. 2003, 77, 1368–1381. [Google Scholar] [CrossRef] [Green Version]
- Novoa, R.R.; Calderita, G.; Cabezas, P.; Elliott, R.M.; Risco, C. Key Golgi Factors for Structural and Functional Maturation of Bunyamwera Virus. J. Virol. 2005, 79, 10852–10863. [Google Scholar] [CrossRef] [Green Version]
- Madoff, D.H.; Lenard, J. A membrane glycoprotein that accumulates intracellularly: Cellular processing of the large glycoprotein of LaCrosse virus. Cell 1982, 28, 821–829. [Google Scholar] [CrossRef]
- Barrett, C.L.; Humphrey, R.L.; Sinsko, M.J.; Grimstad, P.R. Serologic evidence for widespread infection with la crosse and st. louis encephalitis viruses in the indiana human population. Am. J. Epidemiol. 1984, 119, 913–930. [Google Scholar]
- Grimstad, P.R.; Wentworth, B.B.; Calisher, C.H.; Harroff, R.N. Jamestown Canyon Virus (California Serogroup) is the Etiologic Agent of Widespread Infection in Michigan Humans. Am. J. Trop. Med. Hyg. 1986, 35, 376–386. [Google Scholar] [CrossRef] [PubMed]
- Lednicky, J.A.; White, S.K.; Stephenson, C.J.; Cherabuddi, K.; Loeb, J.C.; Moussatche, N.; Lednicky, A.; Morris, J.G., Jr. Keystone Virus Isolated From a Florida Teenager with Rash and Subjective Fever: Another Endemic Arbovirus in the Southeastern United States? Clin. Infect. Dis. 2019, 68, 143–145. [Google Scholar] [CrossRef] [PubMed]
- Parkin, W.E.; Hammon, W.M.; Sather, G.E. Review of Current Epidemiological Literature on Viruses of the California Arbovirus Group. Am. J. Trop. Med. Hyg. 1972, 21, 964–978. [Google Scholar] [CrossRef] [PubMed]
- Monath, T.P.C.; Nuckolls, J.G.; Berall, J.; Bauer, H.; Coleman, P.H.; Chappell, W.A. Studies on California Encephalitis in Minnesota. Am. J. Epidemiol. 1970, 92, 40–50. [Google Scholar] [CrossRef] [PubMed]
- Sampasa-Kanyinga, H.; Lé Vesque, B.; Anassour-Laouan-Sidi, E.; Cô Té Serhir, B.; Ward, B.J.; Libman, M.D.; Drebot, M.A.; Makowski, K.; Dimitrova, K.; Ndao, M.; et al. Zoonotic Infections in Communities of the James Bay Cree Territory: An Overview of Seroprevalence. Can. J. Infect. Dis. Med. Microbiol. 2013, 24, 79–84. [Google Scholar] [CrossRef] [Green Version]
- Evander, M.; Putkuri, N.; Eliasson, M.; Lwande, O.W.; Vapalahti, O.; Ahlm, C. Seroprevalence and Risk Factors of Inkoo Virus in Northern Sweden. Am. J. Trop. Med. Hyg. 2016, 94, 1103–1106. [Google Scholar] [CrossRef] [Green Version]
- Thompson, W.H.; Kalfayan, B.; Anslow, R.O. Isolation of California Encephalitis Group Virus from a Fatal Human Illness. Am. J. Epidemiol. 1965, 81, 245–253. [Google Scholar] [CrossRef]
- Fauvel, M.; Artsob, H.; Calisher, C.H.; Davignon, L.; Chagnon, A.; Skvorc-Ranko, R.; Belloncik, S. California group virus encephalitis in three children from Quebec: Clinical and serologic findings. Can. Med. Assoc. J. 1980, 122, 60–64. [Google Scholar]
- Grimstad, P.R.; Waldman, R.J.; Calisher, C.H.; Shabino, C.L. A Case of Encephalitis in a Human Associated with a Serologic Rise to Jamestown Canyon Virus. Am. J. Trop. Med. Hyg. 1982, 31, 1238–1244. [Google Scholar] [CrossRef]
- CDC. La Crosse Encephalitis. Available online: https://www.cdc.gov/lac/index.html (accessed on 26 June 2019).
- Webster, D.; Dimitrova, K.; Holloway, K.; Makowski, K.; Safronetz, D.; Drebot, M.A. California Serogroup Virus Infection Associated with Encephalitis and Cognitive Decline, Canada. Emerg. Infect. Dis. 2017, 23, 1423–1424. [Google Scholar] [CrossRef] [PubMed]
- Eldridge, B.F.; Glaser, C.; Pedrin, R.E.; Chiles, R.E. The first reported case of California encephalitis in more than 50 years. Emerg. Infect. Dis. 2001, 7, 451–452. [Google Scholar] [CrossRef] [PubMed]
- Sluka, F.; Simkova, A. Demonstration of human infection in the natural focus of the Valtice fever. Folia Parasitol. 1972, 19, 358. [Google Scholar] [PubMed]
- Demikhov, V.G.; Chaitsev, V.G.; Butenko, A.M.; Nedyalkova, M.S.; Morozova, T.N. California Serogroup Virus Infections in the Ryazan Region of the USSR. Am. J. Trop. Med. Hyg. 1991, 45, 371–376. [Google Scholar] [CrossRef] [PubMed]
- Gaensbauer, J.T.; Lindsey, N.P.; Messacar, K.; Staples, J.E.; Fischer, M. Neuroinvasive Arboviral Disease in the United States: 2003 to 2012. Pediatrics 2014, 134, e642–e650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teleron, A.L.A.; Rose, B.K.; Williams, D.M.; Kemper, S.E.; McJunkin, J.E. La Crosse Encephalitis: An Adult Case Series. Am. J. Med. 2016, 129, 881–884. [Google Scholar] [CrossRef] [PubMed]
- Byrd, B.D.; Williams, C.J.; Staples, J.E.; Burkhalter, K.L.; Savage, H.M.; Doyle, M.S. Notes from the Field: Spatially Associated Coincident and Noncoincident Cases of La Crosse Encephalitis—North Carolina, 2002–2017. MMWR Morb. Mortal. Wkly. Rep. 2018, 67, 1104–1105. [Google Scholar] [CrossRef] [PubMed]
- Campbell, W.; Grady, L.; Kirouac, I.; Huang, C.; LaForce, F.M. Diagnosis of Jamestown Canyon Encephalitis by Polymerase Chain Reaction. Clin. Infect. Dis. 1999, 28, 1294–1297. [Google Scholar] [Green Version]
- Johnson, R.T. Pathogenesis of La Crosse virus in mice. Prog. Clin. Biol. Res. 1983, 123, 139–144. [Google Scholar]
- Smee, D.F.; Alaghamandan, H.A.; Cottam, H.B.; Sharma, B.S.; Jolley, W.B.; Robins, R.K. Broad-spectrum in vivo antiviral activity of 7-thia-8-oxoguanosine, a novel immunopotentiating agent. Antimicrob. Agents Chemother. 1989, 33, 1487–1492. [Google Scholar] [CrossRef] [Green Version]
- Bennett, R.S.; Nelson, J.T.; Gresko, A.K.; Murphy, B.R.; Whitehead, S.S. The full genome sequence of three strains of Jamestown Canyon virus and their pathogenesis in mice or monkeys. Virol. J. 2011, 8, 136. [Google Scholar] [CrossRef] [PubMed]
- Janssen, R.; Gonzalez-Scarano, F.; Nathanson, N. Mechanisms of bunyavirus virulence. Comparative pathogenesis of a virulent strain of La Crosse and an avirulent strain of Tahyna virus. Lab. Investig. 1984, 50, 447–455. [Google Scholar] [PubMed]
- Taylor, K.G.; Woods, T.A.; Winkler, C.W.; Carmody, A.B.; Peterson, K.E. Age-Dependent Myeloid Dendritic Cell Responses Mediate Resistance to La Crosse Virus-Induced Neurological Disease. J. Virol. 2014, 88, 11070–11079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bennett, R.S.; Gresko, A.K.; Murphy, B.R.; Whitehead, S.S. Tahyna virus genetics, infectivity, and immunogenicity in mice and monkeys. Virol. J. 2011, 8, 135. [Google Scholar] [CrossRef] [PubMed]
- Taylor, K.G.; Peterson, K.E. Innate immune response to La Crosse virus infection. J. Neurovirol. 2014, 20, 150–156. [Google Scholar] [CrossRef] [PubMed]
- Yoneyama, M.; Kikuchi, M.; Natsukawa, T.; Shinobu, N.; Imaizumi, T.; Miyagishi, M.; Taira, K.; Akira, S.; Fujita, T. The RNA helicase RIG-I has an essential function in double-stranded RNA-induced innate antiviral responses. Nat. Immunol. 2004, 5, 730–737. [Google Scholar] [CrossRef]
- Kato, H.; Sato, S.; Yoneyama, M.; Yamamoto, M.; Uematsu, S.; Matsui, K.; Tsujimura, T.; Takeda, K.; Fujita, T.; Takeuchi, O.; et al. Cell Type-Specific Involvement of RIG-I in Antiviral Response. Immunity 2005, 23, 19–28. [Google Scholar] [CrossRef] [Green Version]
- Kawai, T.; Takahashi, K.; Sato, S.; Coban, C.; Kumar, H.; Kato, H.; Ishii, K.J.; Takeuchi, O.; Akira, S. IPS-1, an adaptor triggering RIG-I- and Mda5-mediated type I interferon induction. Nat. Immunol. 2005, 6, 981–988. [Google Scholar] [CrossRef]
- Perry, A.K.; Chen, G.; Zheng, D.; Tang, H.; Cheng, G. The host type I interferon response to viral and bacterial infections. Cell Res. 2005, 15, 407–422. [Google Scholar] [CrossRef] [Green Version]
- Schuh, T.; Schultz, J.; Moelling, K.; Pavlovic, J. DNA-based vaccine against La Crosse virus: Protective immune response mediated by neutralizing antibodies and CD4+ T cells. Hum. Gene Ther. 1999, 10, 1649–1658. [Google Scholar] [CrossRef]
- Pavlovic, J.; Schultz, J.; Hefti, H.; Schuh, T.; Mölling, K. DNA Vaccination against La Crosse Virus. Intervirology 2000, 43, 312–321. [Google Scholar] [CrossRef] [PubMed]
- Frese, M.; Kochs, G.; Feldmann, H.; Hertkorn, C.; Haller, O. Inhibition of bunyaviruses, phleboviruses, and hantaviruses by human MxA protein. J. Virol. 1996, 70, 915–923. [Google Scholar] [PubMed]
- Hefti, H.P.; Frese, M.; Landis, H.; Di Paolo, C.; Aguzzi, A.; Haller, O.; Pavlovic, J. Human MxA Protein Protects Mice Lacking a Functional Alpha/Beta Interferon System against La Crosse Virus and Other Lethal Viral Infections. J. Virol. 1999, 73, 6984–6991. [Google Scholar] [PubMed]
- Winkler, C.W.; Myers, L.M.; Woods, T.A.; Carmody, A.B.; Taylor, K.G.; Peterson, K.E. Lymphocytes have a role in protection, but not in pathogenesis, during La Crosse Virus infection in mice. J. Neuroinflamm. 2017, 14, 62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varela, M.; Piras, I.M.; Mullan, C.; Shi, X.; Tilston-Lunel, N.L.; Pinto, R.M.; Taggart, A.; Welch, S.R.; Neil, S.J.; Kreher, F.; et al. Sensitivity to BST-2 restriction correlates with Orthobunyavirus host range. Virology 2017, 509, 121–130. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Scarano, F.; Jacoby, D.; Griot, C.; Nathanson, N. Genetics, infectivity and virulence of California serogroup viruses. Virus Res. 1992, 24, 123–135. [Google Scholar] [CrossRef]
- Griot, C.; Pekosz, A.; Davidson, R.; Stillmock, K.; Hoek, M.; Lukac, D.; Schmeidler, D.; Cobbinah, I.; Gonzalez-Scarano, F.; Nathanson, N. Replication in Cultured C2C12 Muscle Cells Correlates with the Neuroinvasiveness of California Serogroup Bunyaviruses. Virology 1994, 201, 399–403. [Google Scholar] [CrossRef]
- Winkler, C.W.; Race, B.; Phillips, K.; Peterson, K.E. Capillaries in the olfactory bulb but not the cortex are highly susceptible to virus-induced vascular leak and promote viral neuroinvasion. Acta Neuropathol. 2015, 130, 233–245. [Google Scholar] [CrossRef] [PubMed]
- Miner, J.J.; Daniels, B.P.; Shrestha, B.; Proenca-Modena, J.L.; Lew, E.D.; LaZear, H.M.; Gorman, M.J.; Lemke, G.; Klein, R.S.; Diamond, M.S. The TAM receptor Mertk protects against neuroinvasive viral infection by maintaining blood-brain barrier integrity. Nat. Med. 2015, 21, 1464–1472. [Google Scholar] [CrossRef] [Green Version]
- Dawes, B.E.; Gao, J.; Atkins, C.; Nelson, J.T.; Johnson, K.; Wu, P.; Freiberg, A.N. Human neural stem cell-derived neuron/astrocyte co-cultures respond to La Crosse virus infection with proinflammatory cytokines and chemokines. J. Neuroinflamm. 2018, 15, 315. [Google Scholar] [CrossRef]
- Pekosz, A.; Phillips, J.; Pleasure, D.; Merry, D.; Gonzalez-Scarano, F. Induction of apoptosis by La Crosse virus infection and role of neuronal differentiation and human bcl-2 expression in its prevention. J. Virol. 1996, 70, 5329–5335. [Google Scholar] [PubMed]
- Mukherjee, P.; Woods, T.A.; Moore, R.A.; Peterson, K.E. Activation of the Innate Signaling Molecule MAVS by Bunyavirus Infection Upregulates the Adaptor Protein SARM1, Leading to Neuronal Death. Immunity 2013, 38, 705–716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winkler, C.W.; Woods, T.A.; Robertson, S.J.; McNally, K.L.; Carmody, A.B.; Best, S.M.; Peterson, K.E. Cutting Edge: CCR2 Is Not Required for Ly6C(hi) Monocyte Egress from the Bone Marrow but Is Necessary for Migration within the Brain in La Crosse Virus Encephalitis. J. Immunol. 2018, 200, 471–476. [Google Scholar] [CrossRef] [PubMed]
- Janssen, R.S.; Nathanson, N.; Endres, M.J.; Gonzalez-Scarano, F. Virulence of La Crosse virus is under polygenic control. J. Virol. 1986, 59, 1–7. [Google Scholar] [PubMed]
- Endres, M.J.; Griot, C.; Gonzalez-Scarano, F.; Nathanson, N. Neuroattenuation of an avirulent bunyavirus variant maps to the L RNA segment. J. Virol. 1991, 65, 5465–5470. [Google Scholar] [PubMed]
- Bennett, R.S.; Gresko, A.K.; Nelson, J.T.; Murphy, B.R.; Whitehead, S.S. A recombinant chimeric La Crosse virus expressing the surface glycoproteins of Jamestown Canyon virus is immunogenic and protective against challenge with either parental virus in mice or monkeys. J. Virol. 2012, 86, 420–426. [Google Scholar] [CrossRef] [PubMed]
- Blakqori, G.; Weber, F. Efficient cDNA-Based Rescue of La Crosse Bunyaviruses Expressing or Lacking the Nonstructural Protein NSs. J. Virol. 2005, 79, 10420–10428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blakqori, G.; Delhaye, S.; Habjan, M.; Blair, C.D.; Sánchez-Vargas, I.; Olson, K.E.; Attarzadeh-Yazdi, G.; Fragkoudis, R.; Kohl, A.; Kalinke, U.; et al. La Crosse Bunyavirus Nonstructural Protein NSs Serves To Suppress the Type I Interferon System of Mammalian Hosts. J. Virol. 2007, 81, 4991–4999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verbruggen, P.; Ruf, M.; Blakqori, G.; Overby, A.K.; Heidemann, M.; Eick, D.; Weber, F. Interferon antagonist NSs of La Crosse virus triggers a DNA damage response-like degradation of transcribing RNA polymerase II. J. Biol. Chem. 2011, 286, 3681–3692. [Google Scholar] [CrossRef] [PubMed]
- Elbers, A.; Koenraadt, C.J.M.; Meiswinkel, R. Mosquitoes and Culicoides biting midges: Vector range and the influence of climate change. Rev. Sci. Tech. 2015, 34, 123–137. [Google Scholar] [CrossRef] [PubMed]
- Beaty, B.; Rozhon, E.; Gensemer, P.; Bishop, D. Formation of reassortant bunyaviruses in dually infected mosquitoes. Virology 1981, 111, 662–665. [Google Scholar] [CrossRef]
- Borucki, M.K.; Beaty, B.J.; Blair, C.D.; Parker, B.M.; Chandler, L.J. Bunyavirus superinfection and segment reassortment in transovarially infected mosquitoes. J. Gen. Virol. 1999, 80, 3173–3179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
Summary of the genetic relationships, mosquito vectors, vertebrate hosts, and human disease for the California serogroup (CSG) viruses, expanded from Figure 1B in [21]. Groups are divided based on phylogenetic classification. Relationships reflect the compilation and summary of several phylogenetic analyses [8,14,18,19,20]. Monophyletic groups are shown and reflect that these were consistent in the majority of studies, and ungrouped vertical lines indicate relationship discrepancies of those viruses between phylogenetic studies. Mosquito genera and species from which CSG viruses have been isolated from are listed, and species in bold indicate primary vectors as determined by frequent virus isolations and laboratory transmission studies. Confirmed and likely vertebrate hosts are designated with colored icons and are based on evidence of both serological evidence in the field, and experimental viremias or virus isolations from animals in the field. Grayed icons indicate possible hosts based on either serological evidence or experimental viremias.
Figure 1.
Summary of the genetic relationships, mosquito vectors, vertebrate hosts, and human disease for the California serogroup (CSG) viruses, expanded from Figure 1B in [21]. Groups are divided based on phylogenetic classification. Relationships reflect the compilation and summary of several phylogenetic analyses [8,14,18,19,20]. Monophyletic groups are shown and reflect that these were consistent in the majority of studies, and ungrouped vertical lines indicate relationship discrepancies of those viruses between phylogenetic studies. Mosquito genera and species from which CSG viruses have been isolated from are listed, and species in bold indicate primary vectors as determined by frequent virus isolations and laboratory transmission studies. Confirmed and likely vertebrate hosts are designated with colored icons and are based on evidence of both serological evidence in the field, and experimental viremias or virus isolations from animals in the field. Grayed icons indicate possible hosts based on either serological evidence or experimental viremias.

Figure 2.
Map of the geographical distribution of the California serogroup viruses, expanded from Figure 1A in [21]. Geographic distribution is based on positive serology of humans and animals, mosquito isolations of virus, and probable and confirmed human cases.
Figure 2.
Map of the geographical distribution of the California serogroup viruses, expanded from Figure 1A in [21]. Geographic distribution is based on positive serology of humans and animals, mosquito isolations of virus, and probable and confirmed human cases.

© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Evans, A.B.; Peterson, K.E. Throw out the Map: Neuropathogenesis of the Globally Expanding California Serogroup of Orthobunyaviruses. Viruses 2019, 11, 794. https://doi.org/10.3390/v11090794
AMA Style
Evans AB, Peterson KE. Throw out the Map: Neuropathogenesis of the Globally Expanding California Serogroup of Orthobunyaviruses. Viruses. 2019; 11(9):794. https://doi.org/10.3390/v11090794
Chicago/Turabian StyleEvans, Alyssa B., and Karin E. Peterson. 2019. "Throw out the Map: Neuropathogenesis of the Globally Expanding California Serogroup of Orthobunyaviruses" Viruses 11, no. 9: 794. https://doi.org/10.3390/v11090794
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.