
High Incidence of Lysogeny in the Oxygen Minimum Zones of the Arabian Sea (Southwest Coast of India)


 ,
, 
Abstract
:1. Introduction
2. Material and Methods
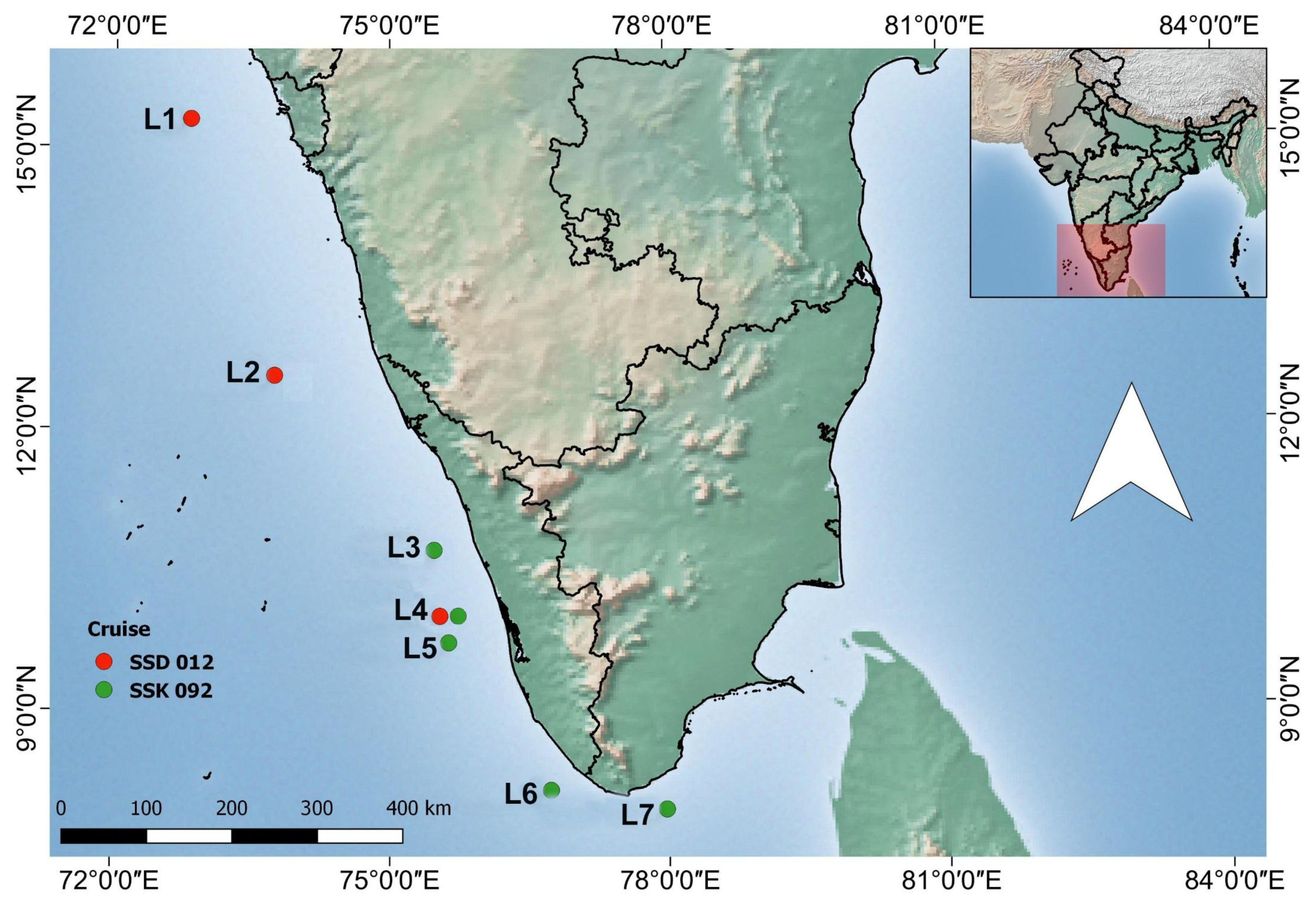
2.1. Study Sites
2.2. Sampling and Physicochemical Characteristics
2.3. Abundances of Viruses and Prokaryotes Using Epifluorescence Microscopy
2.4. Viral Lytic Infection
2.5. Viral Production (VP)
2.6. Induction Assays for Lysogenic Prokaryotes
2.7. Statistical Analysis
3. Results and Discussion
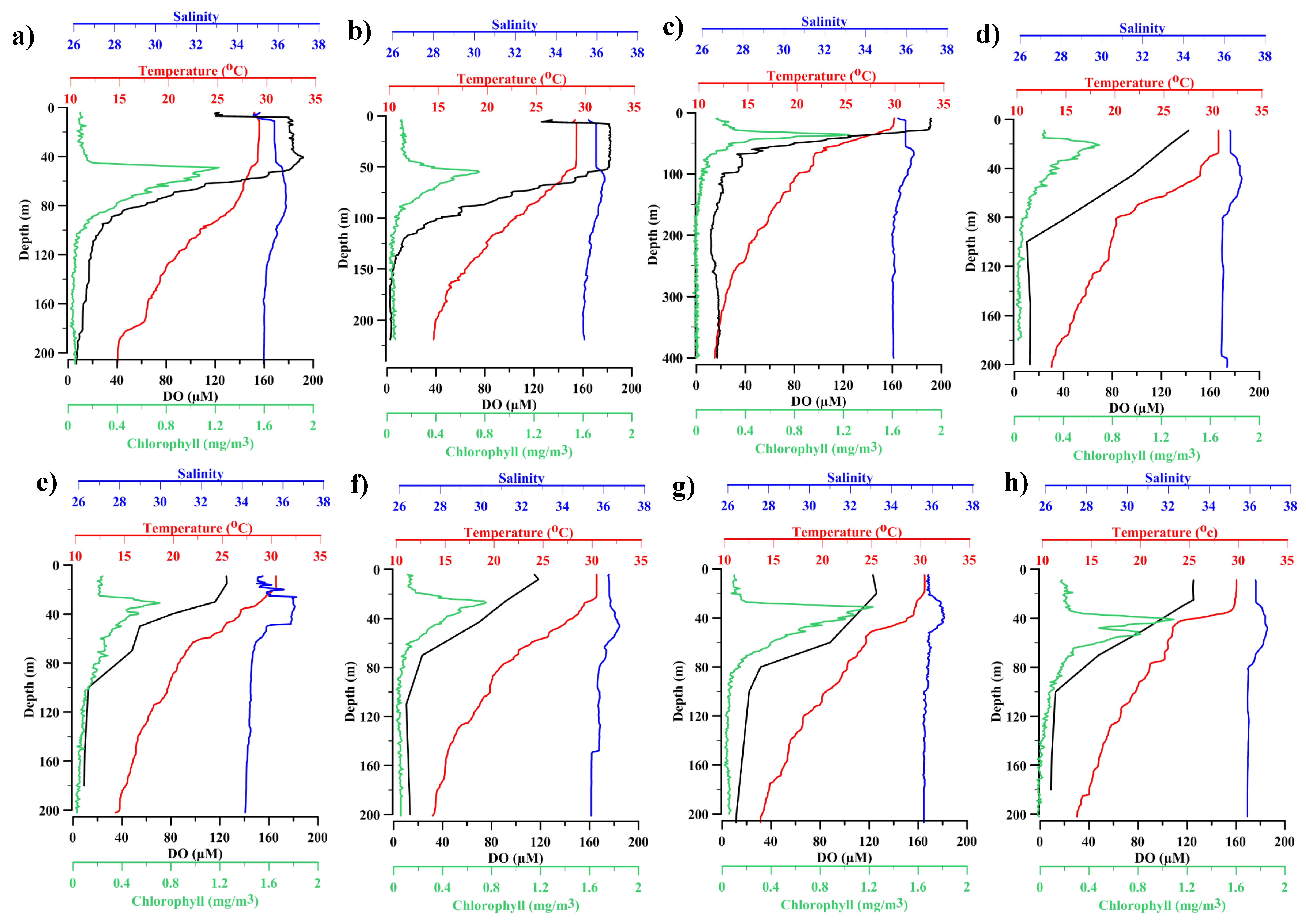
3.1. Physicochemical Characteristics of the Study Area
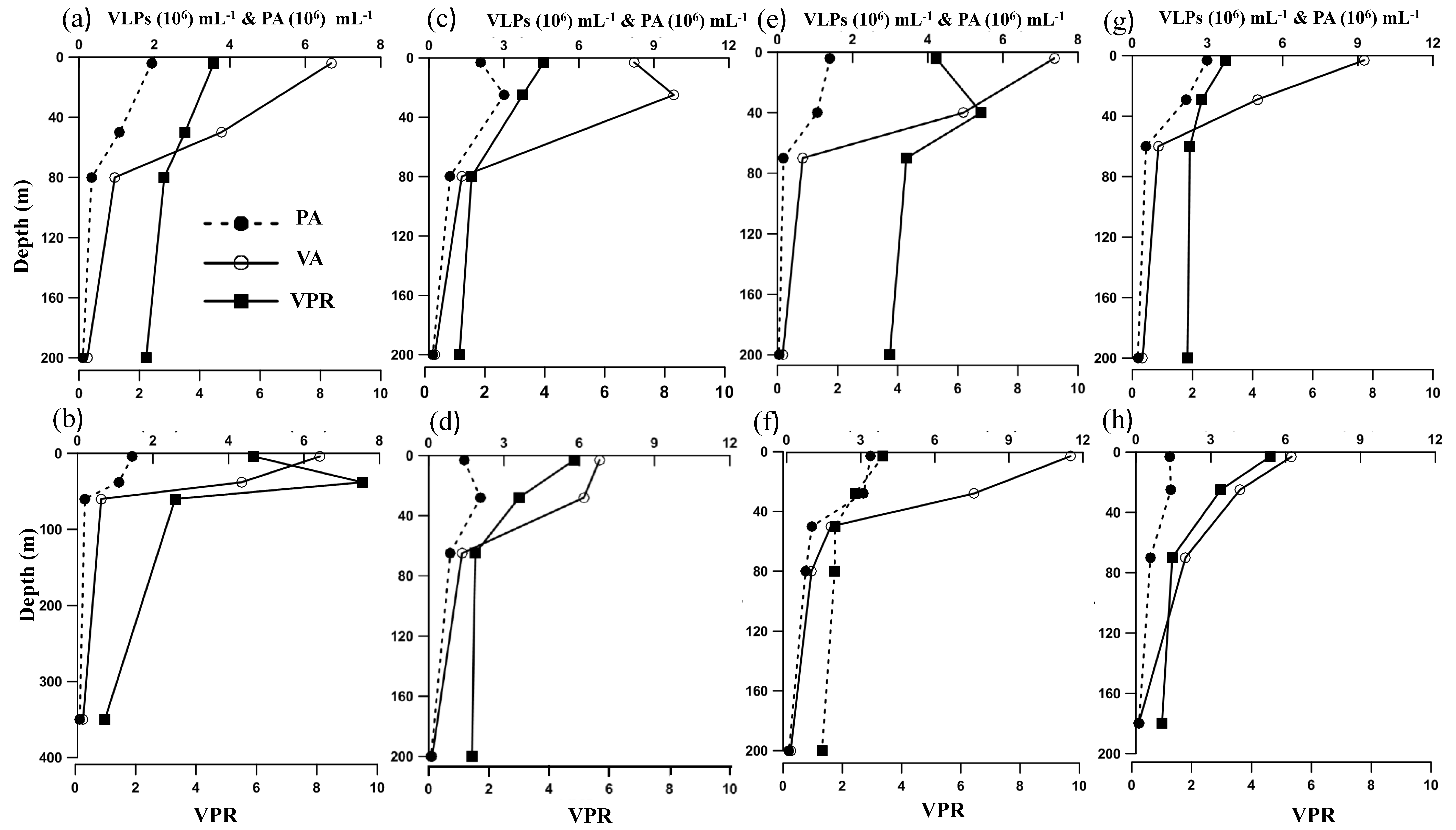
3.2. Viral and Prokaryotic Abundance
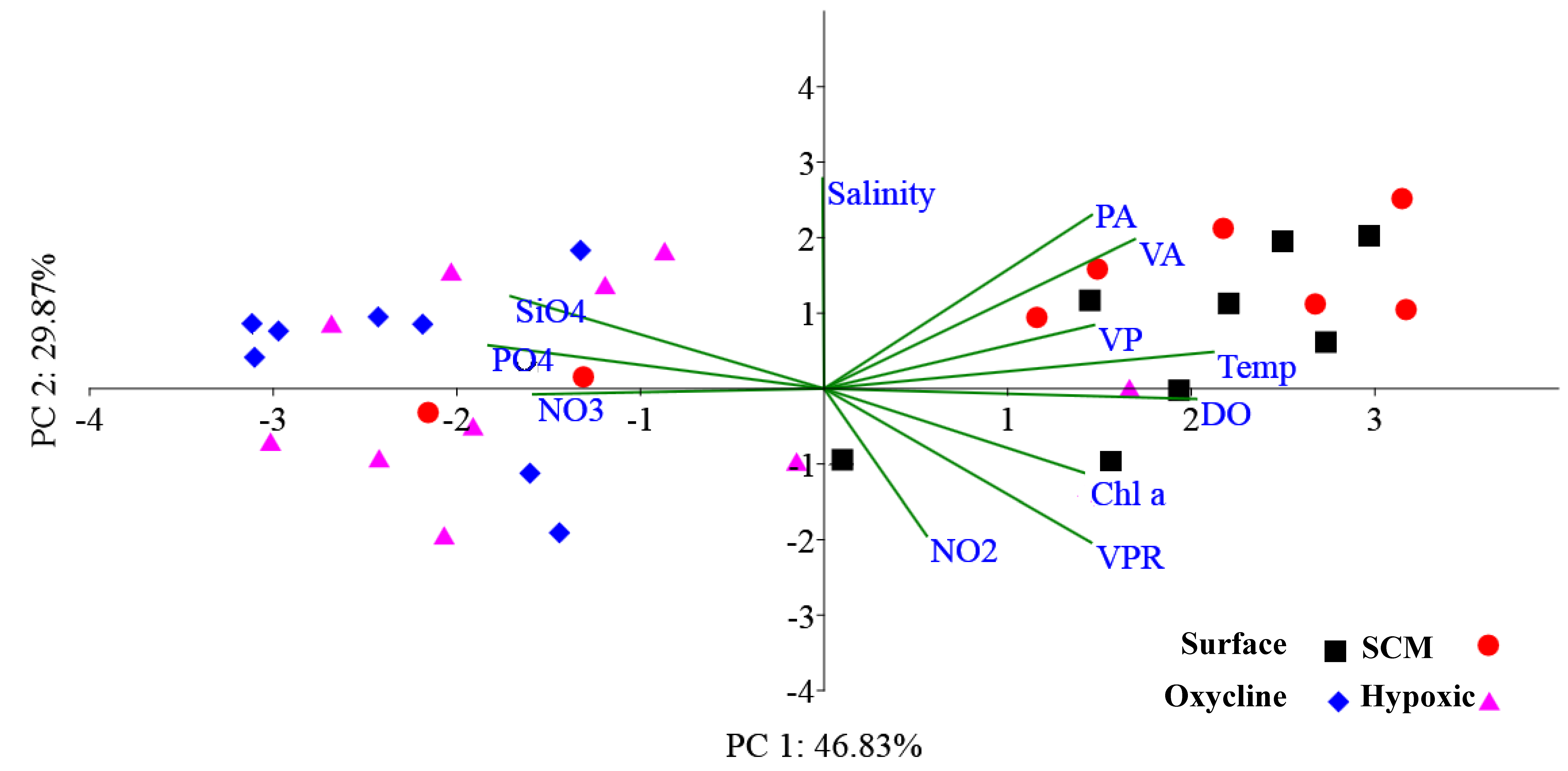
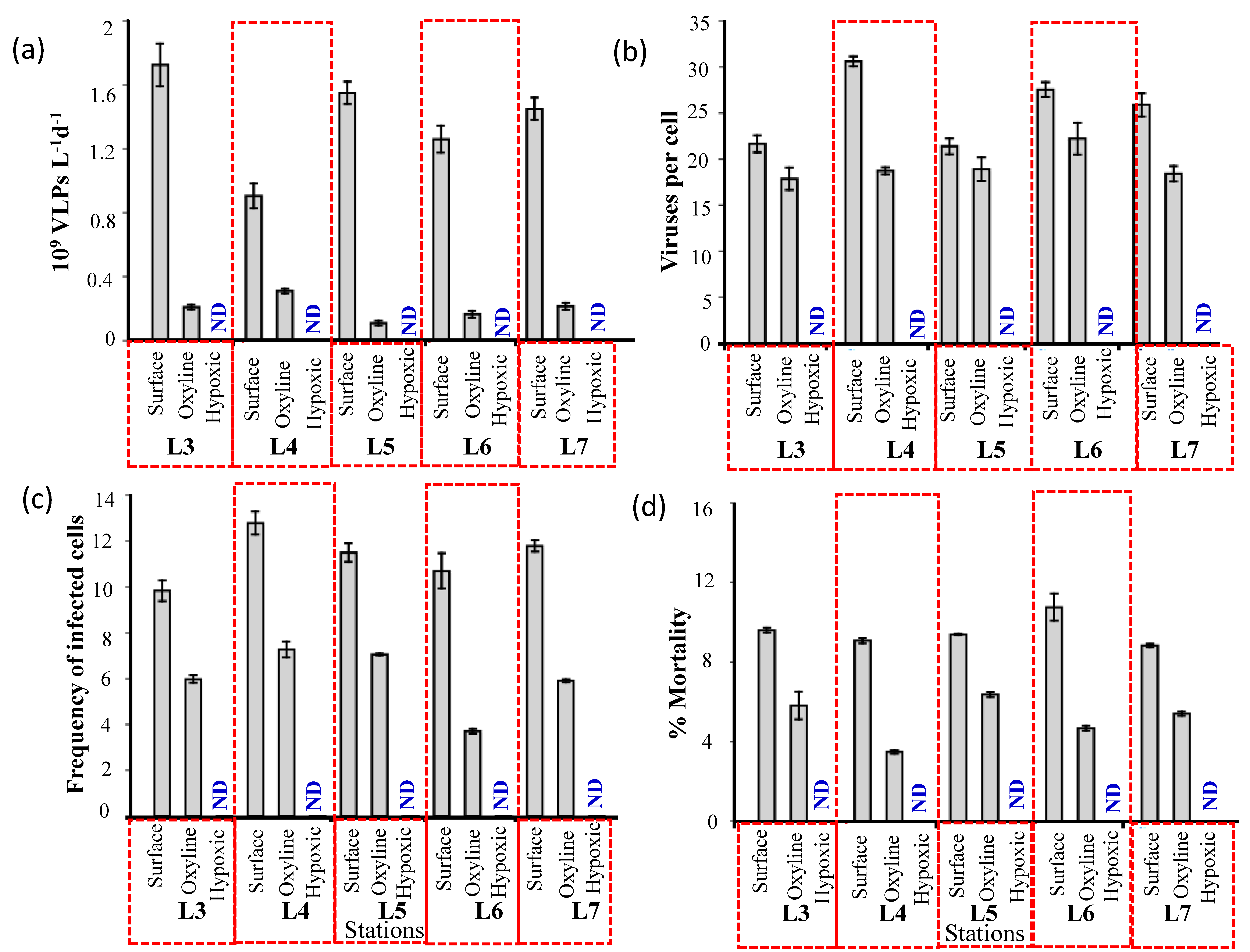
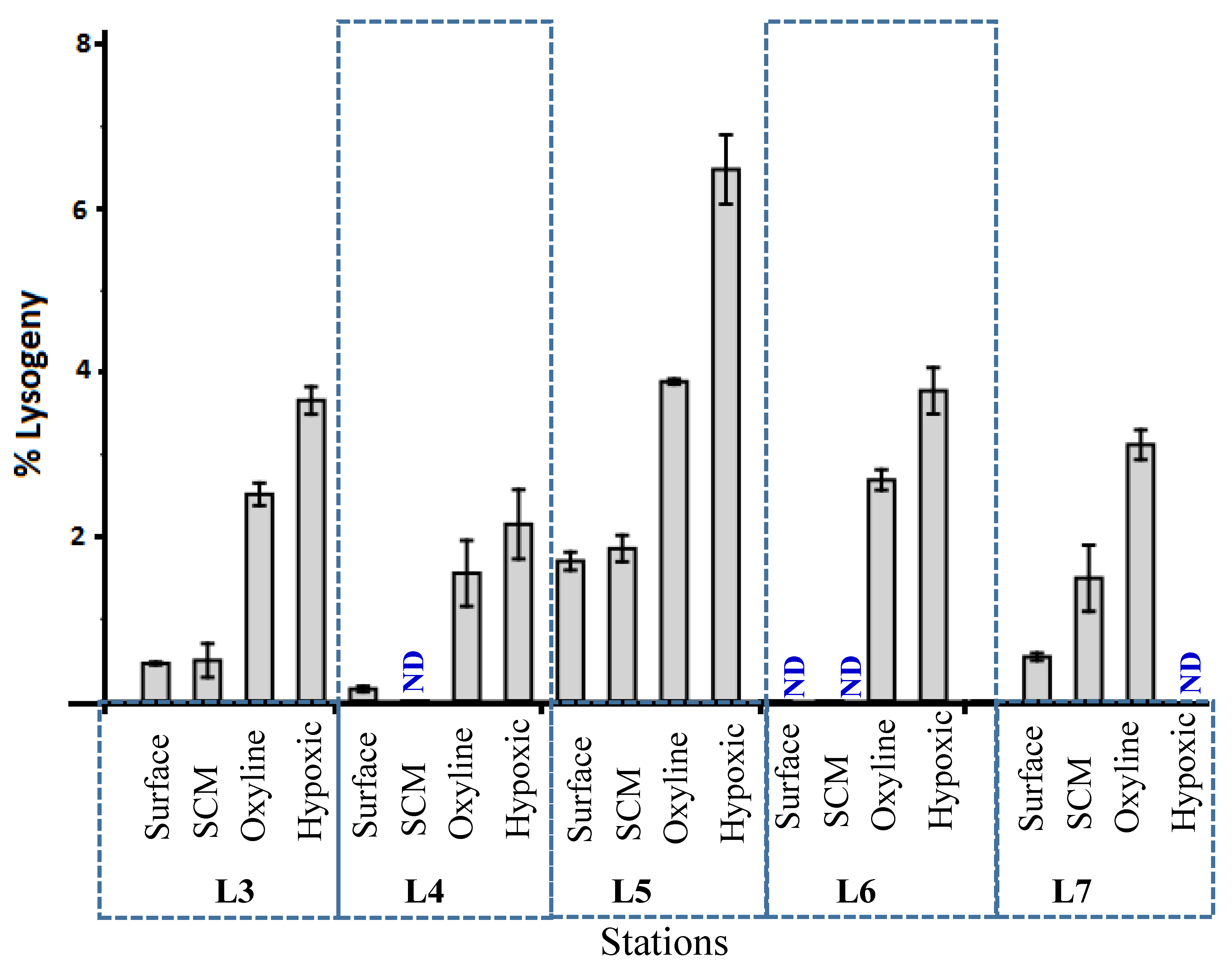
3.3. Lytic Viral Production, Viral-Mediated Prokaryote Mortality, and Lysogenic Induction
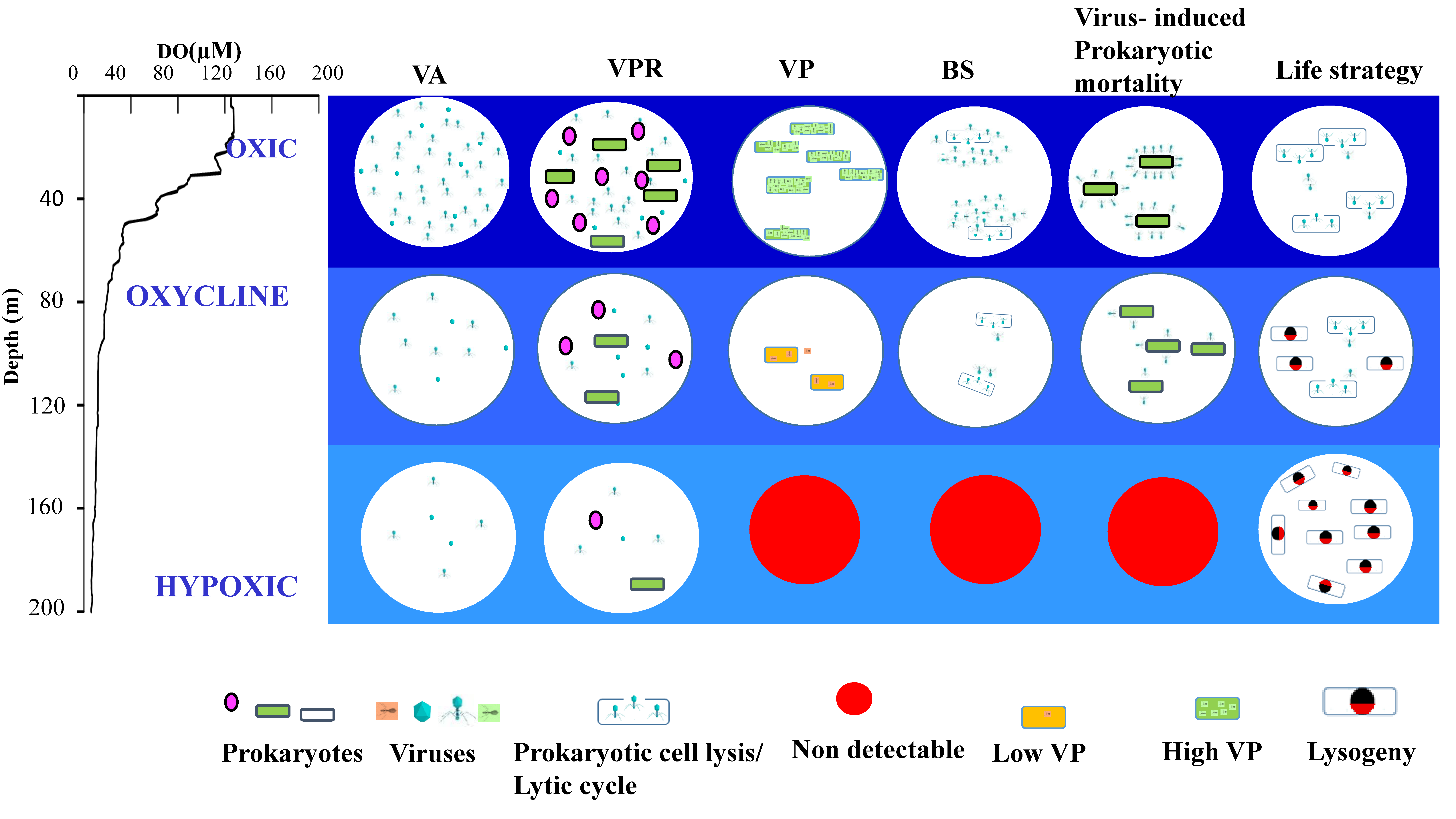
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ulloa, O.; Pantoja, S. The oxygen minimum zone of the eastern South Pacific. Deep Sea Res. Part II Top. Stud. Oceanogr. 2009, 56, 987–991. [Google Scholar] [CrossRef]
- Paulmier, A.; Ruiz-Pino, D. Oxygen Minimum Zones (OMZs) in the modern ocean. Prog. Oceanogr. 2008, 80, 113–118. [Google Scholar] [CrossRef]
- McCreary, J.P.; Yu, Z.; Hood, R.R.; Vinaychandran, P.; Furue, R.; Ishida, A.; Richards, K.J. Dynamics of the Indian-Ocean oxygen minimum zones. Prog. Oceanogr. 2013, 112, 15–37. [Google Scholar] [CrossRef]
- Morrison, J.M.; Codispoti, L.A.; Gaurin, S.; Jones, B.; Magnhnani, V.; Zheng, Z. Seasonal variation of hydrographic and nutrient fields during the US JGOFS Arabian Sea Process Study. Deep Sea Res. Part II 1998, 45, 2053–2102. [Google Scholar] [CrossRef]
- Naqvi, S.W.A.; Muffett, J.W.; Gauns, M.U.; Narvekar, P.V.; Pratihary, A.K.; Naik, H.; Shenoy, D.M.; Jayakumar, D.A.; Geopfert, T.J.; Patra, P.K.; et al. The Arabian Sea as a high-nutrient, low-chlorophyll region during the late Southwest Monsoon. Biogeosciences 2010, 7, 2091–2100. [Google Scholar] [CrossRef] [Green Version]
- Jayakumar, A.; O’Mullan, G.D.; Naqvi, S.W.A.; Ward, B.B. Dentrifying bacterial community composition changes associated with stages of denitrification in oxygen minimum zones. Microb. Ecol. 2009, 58, 350–362. [Google Scholar] [CrossRef] [PubMed]
- Wright, J.J.; Konwar, K.M.; Hallam, S.J. Microbial ecology of expanding oxygen minimum zones. Nat. Rev. Microbiol. 2012, 10, 381–394. [Google Scholar] [CrossRef] [PubMed]
- Kalvelage, T.; Lavik, G.; Lam, P.; Contreras, S.; Arteaga, L.; Loscher, C.R.; Oschlies, A.; Paulmier, A.; Stramma, L.; Kuypers, M.M.M. Nitrogen cycling driven by organic matter export in the South Pacific Oxygen Minimum Zone. Nat. Geosci. 2013, 6, 228–234. [Google Scholar] [CrossRef] [Green Version]
- Ulloa, O.; Wright, J.J.; Belmar, L.; Hallam, S.J. Pelagic Oxygen Minimum Zone Microbial Communities in the Prokaryotes-Prokaryotic Communities and Ecophysiology; Rosenberg, E., Ed.; Springer: Berlin, Germany, 2013; pp. 113–120. [Google Scholar]
- Löscher, C.R.; Bourbonnais, A.; Dekaezemacker, J.; Charoenpong, C.N.; ALtabet, M.A.; Bange, H.W.; Czeschel, R.; Hoffmann, C.; Schmitz, R. N2 fixation in eddies of the eastern tropical South Pacific Ocean. Biogeosciences 2016, 13, 2889–2899. [Google Scholar] [CrossRef] [Green Version]
- Pitcher, A.; Villanueva, L.; Hopmans, E.C.; Schouten, S.; Reichart, G.J.; Sinninghe Damste, J.S. Niche segregation of ammonia-oxidizing archaea and anammox bacteria in the Arabian Sea oxygen minimum zone. ISME J. 2011, 1896–1904. [Google Scholar] [CrossRef] [PubMed]
- Lüke, C.; Speth, D.R.; Kox, M.A.R.; Villanueva, L.; Jetten, M.S.M. Metagenomic analysis of nitrogen and methane cycling in the Arabian Sea oxygen minimum zone. Peer J. 2016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weitz, J.S.; Wilhelm, S.W. Ocean viruses and their effects on microbial communities and biogeochemical cycles. F1000 Biol. Rep. 2012, 4, 17. [Google Scholar] [CrossRef] [PubMed]
- Cassman, N.; Prieto-Davo, A.; Walsh, K.; Silva, G.G.; Angly, F.; Akhter, S.; Barott, K.; Busch, J.; McDote, T.; Haggerty, J.M.; et al. Oxygen minimum zones harbor novel viral communities with low diversity. Environ. Microb. 2012, 14, 3043–3065. [Google Scholar] [CrossRef] [PubMed]
- Mojica, K.D.A.; Brussaard, C.P.D. Factor’s affecting virus dynamics and microbial host-virus interactions in marine environments. FEMS Microbiol. Ecol. 2014, 3, 495–515. [Google Scholar] [CrossRef] [PubMed]
- Finke, J.F.; Hunt, B.P.V.; Winter, C.; Carmack, E.C.; Suttle, C.A. Nutrients and other environmental factors influence virus abundances across oxic and hypoxic marine environments. Viruses 2017, 152. [Google Scholar] [CrossRef] [PubMed]
- Anderson, R.; Winter, C.; Jürgens, K. Protist grazing and viral lysis as prokaryotic mortality factors at Baltic Sea oxic-anoxic interfaces. Mar. Ecol. Prog. Ser. 2012, 467, 1–14. [Google Scholar] [CrossRef]
- Weinbauer, M.G.; Brettar, C.; Höfle, M.G. Lysogeny and virus-induced mortality of bacterioplankton in surface, deep and anoxic marine waters. Limnol. Oceanogr. 2003, 48, 1457–1465. [Google Scholar] [CrossRef]
- Köstner, N.; Scharnreitner, L.; Jurgens, K.; Labrenz, M.; Herndl, G.J.; Winter, C. High viral abundance as a consequence of low viral decay in the Baltic Sea redoxcline. PLoS ONE 2017, 12. [Google Scholar] [CrossRef]
- Roux, S.; Hawley, A.K.; Torres Beltran, M.; Scofield, M.; Schwientek, P.; Stepanauskas, R.; Woyke, T.; Hallam, S.J.; Sullivan, M.B. Ecology and evolution of viruses infecting uncultivated SUPO5 bacteria as revealed by single cell and metagenomics. eLife 2014, 2, 303125. [Google Scholar] [CrossRef]
- Rastelli, E.; Corinaldesi, C.; Petani, B.; Dell’Anno, A.; Ciglenečki, I.; Danovaro, R. Enhanced viral activity and dark CO2 fixation rates under oxygen depletion: The case study of the marine Lake Rogoznica. Environ. Microbiol. 2016, 18, 4511–4522. [Google Scholar] [CrossRef] [PubMed]
- Paul, J.H. Prophages in marine bacteria: Dangerous molecular time bombs or the key to survival in the seas? ISME J. 2008, 2, 579–589. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.C.; Paul, J.H. Significance of Lysogeny in the Marine Environment: Studies with Isolates and a Model of Lysogenic Phage Production. Microb. Ecol. 1998, 35, 235–243. [Google Scholar] [CrossRef] [PubMed]
- Howard-Varona, C.; Hargreaves, K.R.; Abedon, S.T.; Sullivan, M.B. Lysogeny in nature: Mechanisms, impact and ecology of temperate phages. ISME J. 2017, 11, 1511–1520. [Google Scholar] [CrossRef] [PubMed]
- McDaniel, L.; Paul, J.H. Effect of nutrient addition and environmental factors on prophage induction in natural populations of marine Synechococcus species. Appl. Environ. Microbiol. 2005, 71, 842–850. [Google Scholar] [CrossRef] [PubMed]
- Long, A.; McDaniel, L.D.; Mobberley, J.; Paul, J.H. Comparison of lysogeny (prophage induction) in heterotrophic bacterial and Synechococcus populations in the Gulf of Mexico and Mississippi river plume. ISME J. 2008, 2. [Google Scholar] [CrossRef] [PubMed]
- Brum, J.R.; Hurwitz, B.L.; Schofield, O.; Ducklow, H.W.; Sullivan, M.B. Seasonal time bombs: Dominant temperate viruses affect Southern Ocean microbial dynamics. ISME J. 2016, 10, 437. [Google Scholar] [CrossRef] [PubMed]
- Knowles, B.; Silveira, C.B.; Bailey, B.A.; Barott, K.; Cantu, V.A.; Cobián-Güemes, A.G.; George, E.E. Lytic to temperate switching of viral communities. Nature 2016, 531, 466. [Google Scholar] [CrossRef] [PubMed]
- Wigington, C.H.; Sonderegger, D.; Brussaard, C.P.; Buchan, A.; Finke, J.F.; Fuhrman, J.A. Re-examination of the relationship between marine virus and microbial cell abundances. Nat. Microbiol. 2016, 1, 15024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parikka, K.J.; Le Romancer, M.; Wauters, N.; Jacquet, S. Deciphering the virus-to-prokaryote ratio (VPR): Insights into virus–host relationships in a variety of ecosystems. Biol. Rev. 2017, 92, 1081–1100. [Google Scholar] [CrossRef] [PubMed]
- Weitz, J.S.; Beckett, S.J.; Brum, J.R.; Cael, B.B.; Dushoff, J. Lysis, lysogeny and virus-microbe ratios. Nature 2017, 549, E1–E3. [Google Scholar] [CrossRef] [PubMed]
- Jain, A.; Bandekar, M.; Gomes, J.; Shenoy, D.M.; Meena, R.M.; Naik, H.; Khandeparkar, R.; Ramaiah, N. Temporally invariable bacterial community structure in the Arabian Sea oxygen minimum zone. Aquat. Microb. Ecol. 2014, 73, 51–67. [Google Scholar] [CrossRef]
- Grasshoff, K.; Ehrhardt, M.K.K. Methods of Seawater Analysis; Grasshoff, K., Ehrhardt, M., Kremling, K., Eds.; Verlag Chemie: Weinheim, Germany, 1983; pp. 89–224. [Google Scholar]
- Patel, A.; Noble, R.T.; Steele, J.A.; Schwalbach, M.S.; Hewson, I.; Fuhrman, J.A. Virus and prokaryote enumeration from planktonic aquatic environments by epifluorescence microscopy with SYBR Green I. Nat. Protoc. 2007, 2, 269–276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pradeep Ram, A.S.; Sime-Ngando, T. Functional responses of prokaryotes and viruses to grazer effects and nutrient additions in freshwater microcosms. ISME J. 2008, 2, 498–509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Binder, B. Reconsidering the relationship between virally induced bacterial mortality and frequency of infected cells. Aquat. Microb. Ecol. 1999, 18, 207–215. [Google Scholar] [CrossRef] [Green Version]
- Wilhelm, S.W.; Brigden, S.; Suttle, C. A dilution technique for the direct measurement of viral production. A comparison in stratified and tidally mixed coastal waters. Microb. Ecol. 2002, 43, 168–173. [Google Scholar] [CrossRef] [PubMed]
- Brettar, I.; Rheinheimer, G. Denitification at the oxic-anoxic interface. Mar. Ecol. Prog. Ser. 1991, 77, 157–169. [Google Scholar] [CrossRef]
- Hewson, I.; Fuhrman, J.A. Covariation of viral parameters with bacterial assemblage richness and diversity in the water column and sediments. Deep Sea Res. Part I Oceanogr. Res. 2007, 54, 811–830. [Google Scholar] [CrossRef]
- Winter, C.; Herndl, G.; Weinbauer, M.G. Diel cycles in viral infection of bacterioplankton in the North Sea. Aquat. Microb. Ecol. 2004, 35, 207–216. [Google Scholar] [CrossRef] [Green Version]
- Paul, J.H.; Jiang, S.C. Lysogeny and Transduction. In Marine Microbiology (Methods in Microbiology); Pauls, J.H., Ed.; Academic Press: Cambridge, MA, USA, 2001; p. 666. [Google Scholar]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological Statistics Software Package for education and data analysis. Palaeontol. Electron. 2001, 4, 4–9. [Google Scholar]
- McArdle, B.H.; Marti, J. Anderson. Fitting multivariate models to community data: A comment on distance-based redundancy analysis. Ecology 2001, 82, 290–297. [Google Scholar] [CrossRef]
- Prasanna, K.S.; Madhupratap, M.; Dileep, K.M.; Muraleedharan, P.M.; De Souza, S.N.; Gauns, M.; Sarma, V.V.S.S. High biological productivity in the central Arabian Sea during the summer monsoon driven by Ekman pumping and lateral advection. Curr. Sci. 2001, 81, 1633–1638. [Google Scholar]
- Madhupratap, M.; Kumar, S.P.; Bhattathiri, P.M.A.; Kumar, M.D.; Raghukumar, S.; Nair, K.K.C.; Ramaiah, N. Mechanisms of the biological response to winter cooling in the northeastern Arabian Sea. Nature 1996, 384, 349–352. [Google Scholar] [CrossRef]
- Rao, A.O.; Madhu, J.; Indu, J.; Mahapatra, D.K.; Babu, S.V. Modelling of coastal upwelling along the southwest coast of India using POM. Proc. METOC 2004, 2004, 183–188. [Google Scholar]
- Muni Krishna, K. Coastal upwelling along the southwest coast of India-ENSO modulation. Ocean Sci. Discuss. 2008, 5, 123–134. [Google Scholar] [CrossRef]
- McCreary, J.P.; Kundu, P.K.; Molinari, R.L. A numerical investigation of dynamics, thermodynamics and mixed-layer process in the Indian Ocean. Prog. Oceanogr. 1993, 31, 181–244. [Google Scholar] [CrossRef]
- Kuypers, M.M.; Lavik, G.; Woebken, D.; Schmid, M.; Fuchs, B.M.; Amann, R.; Jørgensen, B.B.; Jetten, M.S.M. Massive nitrogen loss from the Benguela upwelling system through anaerobic ammonium oxidation. Proc. Natl. Acad. Sci. USA 2005, 102, 6478–6483. [Google Scholar] [CrossRef] [PubMed]
- Hara, S.; Koike, I.; Terauchi, K.; Kamiya, H.; Tanoue, E. Abundance of viruses in deep oceanic waters. Mar. Ecol. Prog. Ser. 1996, 145, 269–277. [Google Scholar] [CrossRef] [Green Version]
- Weinbauer, M.G.; Fuks, D.; Puskaric, S.; Peduzzi, P. Diel, seasonal and depth-related variability of viruses and dissolved DNA in the northern Adriatic Sea. Microb. Ecol. 1995, 30, 25–41. [Google Scholar] [CrossRef] [PubMed]
- Tuomi, P.; Torsvik, T.; Heldal, M.; Bratbak, G. Bacterial population dynamics in a meromitic lake. Appl. Environ. Microbiol. 1997, 63, 2181–2188. [Google Scholar] [PubMed]
- Clasen, J.L.; Brigden, S.M.; Payet, J.P.; Suttle, C.A. Evidence that viral abundance across oceans and lakes is driven by different biological factors. Freshw. Biol. 2008, 53, 1090–1100. [Google Scholar] [CrossRef]
- Parada, V.; Herndl, J.; Weinbauer, M.G. Viral burst size of heterotrophic prokaryotes in aquatic systems. J. Mar. Biol. Assoc. 2006, 86, 613–621. [Google Scholar] [CrossRef]
- Ward, B.B.; Devol, A.H.; Rich, J.J.; Chang, B.X.; Bulow, S.E.; Naik, H.; Pratihary, A.; Jayakumar, A. Denitrification as the dominant nitrogen loss process in the Arabian Sea. Nature 2009, 7260, 78. [Google Scholar] [CrossRef] [PubMed]
- Beman, J.M.; Carolan, M.T. Deoxygenation alters bacterial diversity and community composition in the ocean’s largest oxygen minimum zone. Nat. Commun. 2013, 4, 2705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bettarel, Y.; Sime-Ngando, T.; Amblard, C.; Dolan, J. Viral activity in two contrasting lake ecosystems. Appl. Environ. Microbiol. 2004, 70, 2941–2951. [Google Scholar] [CrossRef] [PubMed]
- Steward, C.C.; Pinckney, J.; Piceno, Y.; Lovell, C.R. Bacterial numbers and activity, microalgal biomass and productivity and meiofaunal distribution in sediments naturally contaminated with biogenic bromophenols. Mar. Ecol. Prog. Ser. 1992, 90, 61–72. [Google Scholar] [CrossRef]
- Weinbauer, M.G.; Peduzzi, P. Frequency, size and distribution of bacteriophages in different marine bacterial morphotypes. Mar. Ecol. Prog. Ser. 1994, 108, 11–20. [Google Scholar] [CrossRef]
- Suttle, C.A.; Chan, A.M. Dynamics and distribution of cyanophages and their effect on marine Synechococcus spp. Appl. Environ. Microbiol. 1994, 60, 3167–3174. [Google Scholar] [PubMed]
- Pradeep Ram, A.S.; Sime-Ngando, T. Resources drive trade-off between viral lifestyles in the plankton: Evidence from freshwater microbial microcosms. Environ. Microbiol. 2010, 12, 467–479. [Google Scholar] [CrossRef] [PubMed]
- Rastelli, E.; Corinaldesi, C.; Dell’Anno, A.; Tangherlini, M.; Martorelli, E.; Ingrassia, M. High potential for temperate viruses to drive carbon cycling in chemoautotrophy-dominated shallow-water hydrothermal vents. Environ. Microbiol. 2017, 19, 4432–4446. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.C.; Paul, J.H. Gene transfer by transduction in the marine environment. Appl. Environ. Microbiol. 1998, 64, 2780–2787. [Google Scholar] [PubMed]
- Weinbauer, M.G.; Suttle, C.A. Lysogeny and prophage induction in coastal and offshore bacterial communities. Aquat. Microb. Ecol. 1999, 18, 217–225. [Google Scholar] [CrossRef] [Green Version]
- Maurice, C.F.; Bouvier, T.; Comte, J.; Guillemette, F.; Del Giorgio, P.A. Seasonal variations of phage life strategies and bacterial physiological states in three northern temperate lakes. Environ. Microbiol. 2010, 12, 628–641. [Google Scholar] [CrossRef] [PubMed]
- Maurice, C.; Bouvier, C.; De Wit, R.; Bouvier, T. Linking the lytic and lysogenic bacteriophage cycles to environmental conditions, host physiology and their variability in coastal lagoons. Environ. Microbiol. 2013, 15, 2463–2475. [Google Scholar] [CrossRef] [PubMed]
- Payet, J.P.; Suttle, C.A. To kill or not to kill: The balance between lytic and lysogenic viral infection is driven by trophic status. Limnol. Oceanogr. 2013, 58, 465–474. [Google Scholar] [CrossRef] [Green Version]
- Brum, J.R.; Ignacio-Espinoza, J.C.; Roux, S.; Doulcier, G.; Acinas, S.G.; Alberti, A. Patterns and ecological drivers of ocean viral communities. Science 2015, 348, 1261498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pradeep Ram, A.S.; Boucher, D.; Sime-Ngando, T.; Debroas, D.; Romagoux, J.C. Phage bacteriolysis, protistan bacterivory potential, and bacterial production in a freshwater reservoir: Coupling with temperature. Microb. Ecol. 2005, 50, 64–72. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sl. No | Latitude | Longitude | Location Name | Depth (m) |
|---|---|---|---|---|
| RV Sindhu Sadhana (2015) | ||||
| 1 | 15.3005 N | 72.82274 E | L1 | 200 m |
| 2 | 12.56856 N | 73.75 E | L2 | 400 m |
| 3 | 9.956254 N | 75.54024 E | L4 | 200 m |
| RV Sindhu Sankalp (2016) | ||||
| 5 | 10.7028 N | 75.5691 E | L3 | 200 m |
| 6 | 9.956254 N | 75.54024 E | L4 | 200 m |
| 7 | 9.76388 N | 75.61861 E | L5 | 200 m |
| 8 | 8.13944 N | 76.7325 E | L6 | 200 m |
| 9 | 7.93388 N | 77.9722 E | L7 | 200 m |
| Parameters | Surface | SCM | Oxyline | Hypoxic |
|---|---|---|---|---|
| Depth (m) | 5.0 ± 2.4 | 29.0 ± 12.1 | 64.0 ± 10.2 | 200 ± 10.1 |
| Temperature (°C) | 30.1 ± 0.4 | 29.3 ± 1.21 | 20.8 ± 2.0 | 14.6 ± 2.0 |
| Salinity | 35.5 ± 2.4 | 36 ± 2.4 | 35.3 ± 2.49 | 35.2 ± 2.6 |
| DO (µM) | 146.3 ± 29.0 | 137.6 ± 28.2 | 69. 8 ± 12.5 | 12.5 ± 3.4 |
| NO2 (µM) | 0.09 ±0.10 | 0.24 ± 0.23 | 0.11 ± 0.10 | 0.08 ± 0.11 |
| NO3 (µM) | 1 ± 1.8 | 0.6 ± 0.8 | 6.2 ± 5.7 | 7.8 ± 3.7 |
| PO4 (µM) | 0.4 ± 0.5 | 0.4 ± 0.3 | 1.0 ± 0.5 | 1.7 ± 0.7 |
| SiO4 (µM) | 3.3 ± 4.5 | 3.2 ± 2.8 | 10.1 ± 6.3 | 17.8 ± 8.6 |
| Chl a (mg/m3) | 0.3 ± 0.3 | 0.6 ± 0.4 | 0.1 ± 0.03 | 0.03 ± 0.03 |
| PA (105 Cells/mL) | 1.6 ± 1.1 | 1.6 ± 1.1 | 0.6 ± 0.3 | 0.2 ± 0.2 |
| VA (106 VLPs/mL) | 6.1 ± 3.4 | 4.7 ± 2.9 | 1.2 ± 0.5 | 0.3 ± 0.3 |
| VPR | 4.4 ± 0.8 | 4.2 ± 2.4 | 2.3 ± 1.0 | 1.8 ± 0.8 |
| VP (109 VLPs/L/d) | 1.1 ± 0.4 | 0.2 ± 0.1 | ND | ND |
| Variable | SS | FP | Pc | Prop. |
|---|---|---|---|---|
| DO | 6738.6 | 86.388 | 0.001 * | 0.74867 |
| NO2 | 549.65 | 1.8862 | 0.156 | 6.11E-02 |
| NO3 | 5234.5 | 40.306 | 0.089 | 0.58157 |
| PO4 | 4901.2 | 34.671 | 0.079 | 0.54453 |
| SiO4 | 4381.9 | 27.512 | 0.002 | 0.48684 |
| Temp | 6421.8 | 72.214 | 0.001 * | 0.71348 |
| Salinity | 617.62 | 2.1366 | 0.136 | 6.86E-02 |
| PA | 3237.7 | 16.293 | 0.001 * | 0.35972 |
| Chl a | 4187.8 | 25.233 | 0.001 * | 0.46527 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Parvathi, A.; Jasna, V.; Aparna, S.; Pradeep Ram, A.S.; Aswathy, V.K.; Balachandran, K.K.; Muraleedharan, K.R.; Mathew, D.; Sime-Ngando, T. High Incidence of Lysogeny in the Oxygen Minimum Zones of the Arabian Sea (Southwest Coast of India). Viruses 2018, 10, 588. https://doi.org/10.3390/v10110588
Parvathi A, Jasna V, Aparna S, Pradeep Ram AS, Aswathy VK, Balachandran KK, Muraleedharan KR, Mathew D, Sime-Ngando T. High Incidence of Lysogeny in the Oxygen Minimum Zones of the Arabian Sea (Southwest Coast of India). Viruses. 2018; 10(11):588. https://doi.org/10.3390/v10110588
Chicago/Turabian StyleParvathi, Ammini, Vijayan Jasna, Sreekumar Aparna, Angia Sriram Pradeep Ram, Vijaya Krishna Aswathy, Kizhakkeppat K. Balachandran, Kallungal Ravunnikutty Muraleedharan, Dayana Mathew, and Telesphore Sime-Ngando. 2018. "High Incidence of Lysogeny in the Oxygen Minimum Zones of the Arabian Sea (Southwest Coast of India)" Viruses 10, no. 11: 588. https://doi.org/10.3390/v10110588