
Anti-Leishmania amazonensis Activity, Cytotoxic Features, and Chemical Profile of Allium sativum (Garlic) Essential Oil

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemical Reagents and Culture Media
2.2. Parasites and Cell Cultures
2.3. ASEO Phytochemical Analysis
2.4. Viability Assay for Parasites
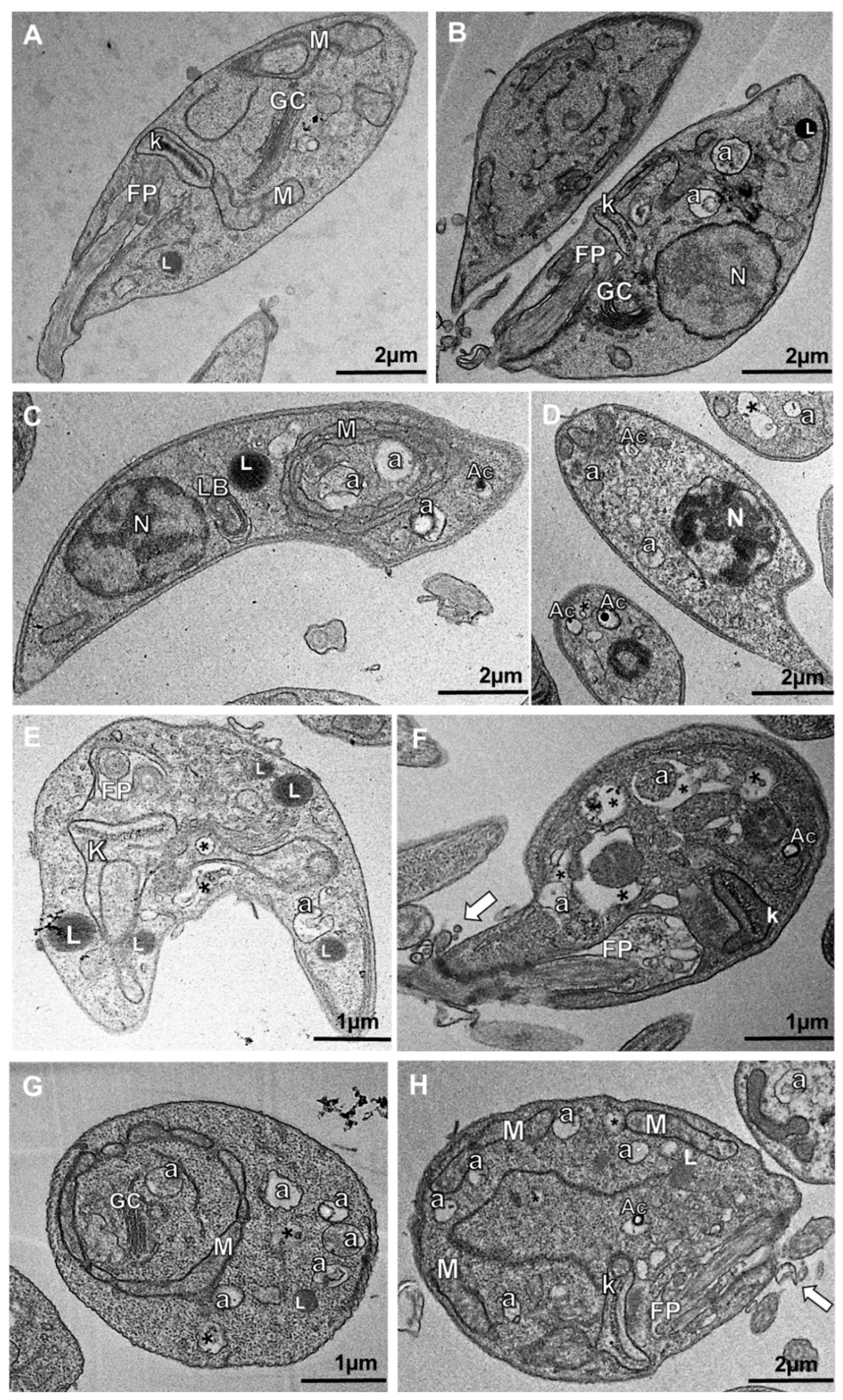
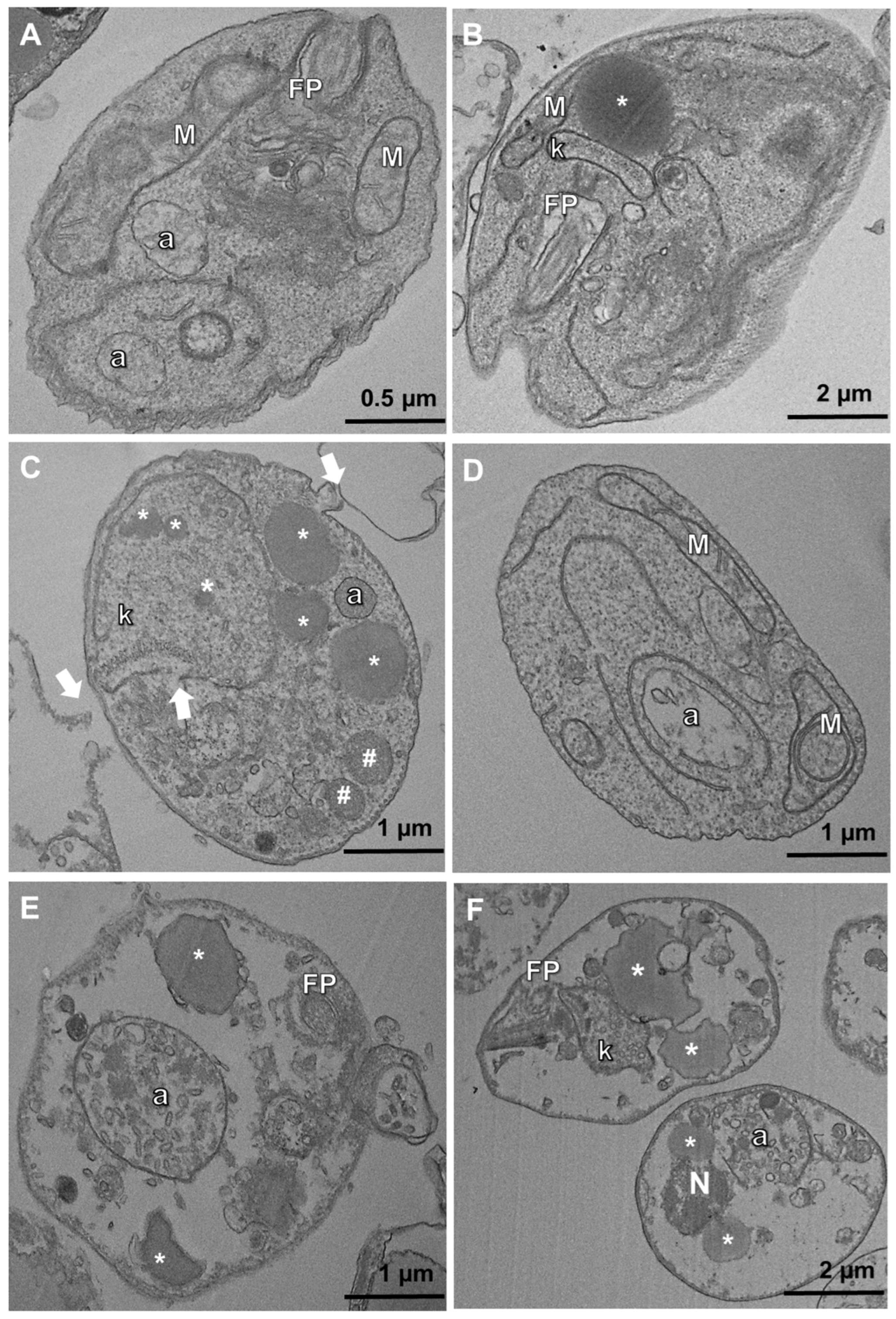
2.5. Transmission Electron Microscopy (TEM)
2.6. Viability Assay for Mammalian Cells and Selectivity
2.7. Hemolytic Assay
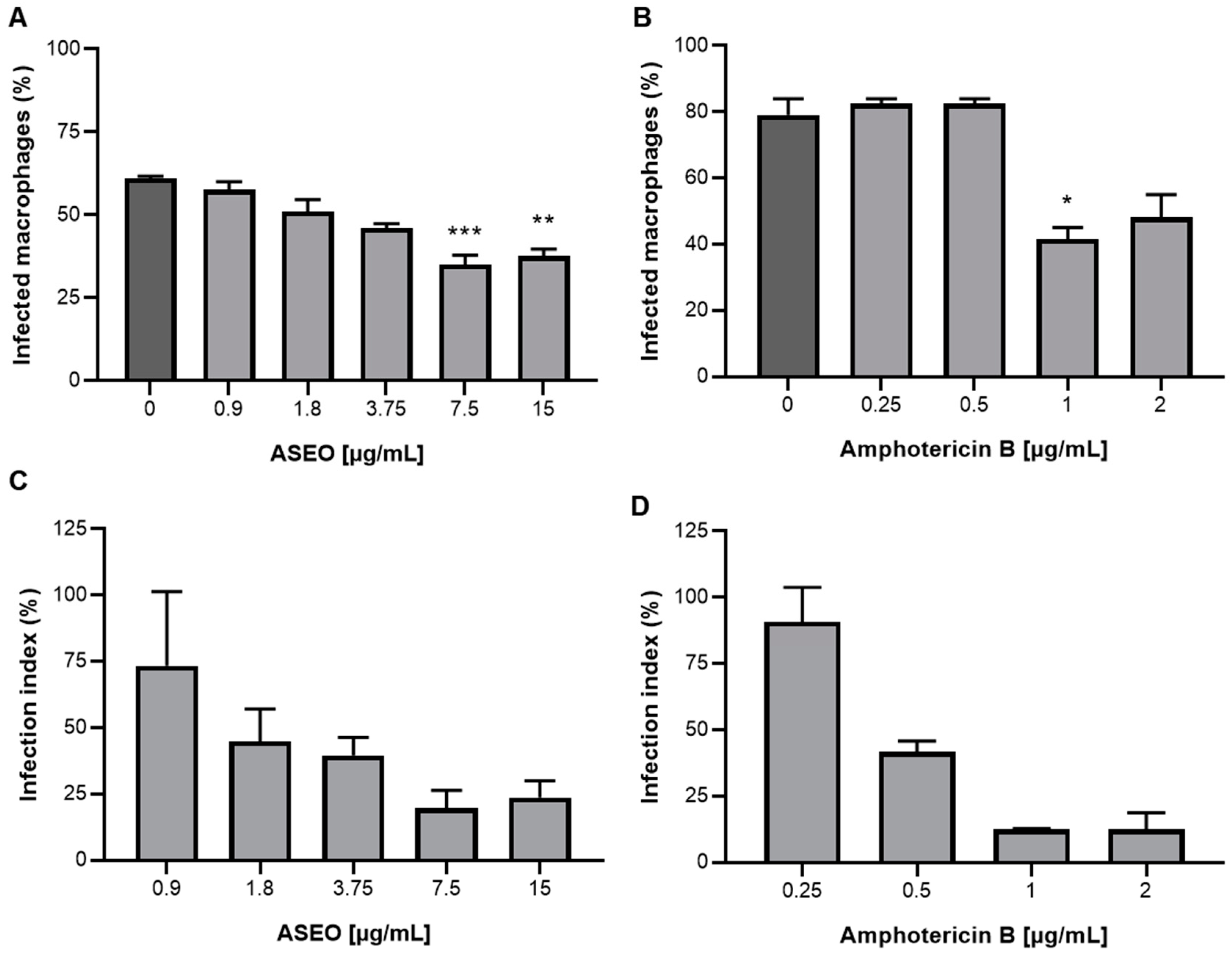
2.8. Infection of Primary Macrophages and Anti-Intracellular Amastigote Assay
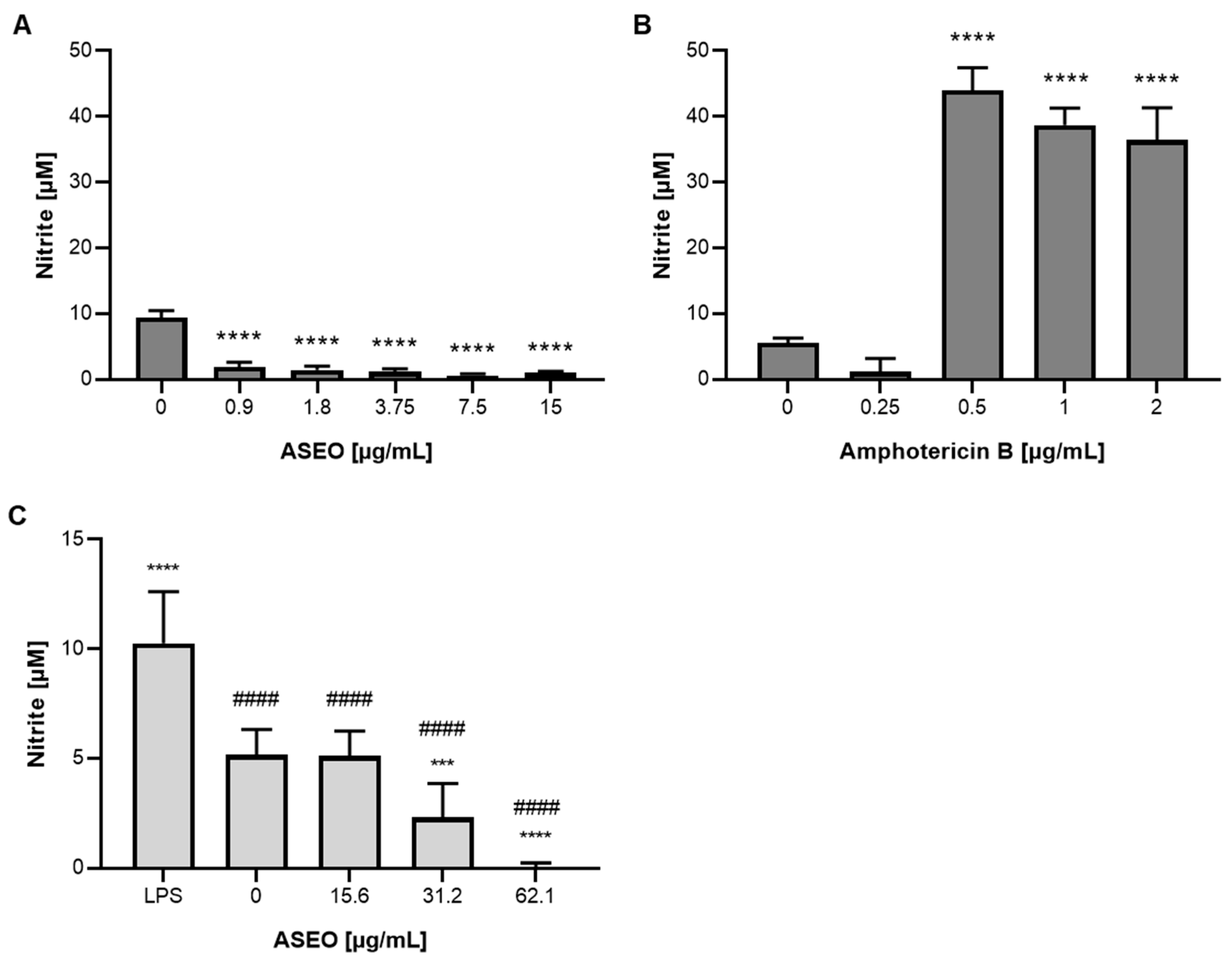
2.9. Determination of Nitric Oxide
2.10. Mitochondrial Membrane Potential (ΔΨm)
2.11. Determination of Intracellular Reactive Oxygen Species (ROS)
2.12. Autophagic Activity
2.13. In Silico Analysis
2.14. Statistical Analyses
3. Results and Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Brindha, J.; Balamurali, M.M.; Chanda, K. An Overview on the therapeutics of neglected infectious diseases-leishmaniasis and Chagas diseases. Front. Chem. 2021, 9, 622286. [Google Scholar] [CrossRef]
- WHO. World Health Organization. 2022. Available online: https://www.who.int/health-topics/leishmaniasis#tab=tab_1 (accessed on 9 November 2022).
- Elmahallawy, E.K.; Martinez, A.S.; Rodriguez-Granger, J.; Hoyos-Mallecot, Y.; Agil, A.; Mari, J.M.N.; Fernandez, J.G. Diagnosis of leishmaniasis. J. Infect. Dev. Ctries. 2014, 8, 961–972. [Google Scholar] [CrossRef] [Green Version]
- David, C.V.; Craft, N. Cutaneous and mucocutaneous leishmaniasis. Dermatol. Ther. 2009, 22, 491–502. [Google Scholar] [CrossRef] [PubMed]
- Carvalho-Silva, R.; Ribeiro-da-Silva, R.C.; Cruz, L.N.P.D.; Oliveira, M.S.; Amoedo, P.M.; Rebêlo, J.M.M.; Guimarães-E-Silva, A.S.; Pinheiro, V.C.S. The predominance of Leishmania (Leishmania) amazonensis DNA in Lutzomyia longipalpis sand flies (Diptera: Psychodidae) from an endemic area for leishmaniasis in Northeastern Brazil. Rev. Inst. Med. Trop. Sao Paulo 2022, 64, e32. [Google Scholar] [CrossRef] [PubMed]
- Marchi, M.N.A.; Caldart, E.T.; Martins, F.D.C.; Freire, R.L. Spatial analysis of leishmaniasis in Brazil: A systematized review. Rev. Inst. Med. Trop. Sao Paulo 2019, 61, e68. [Google Scholar] [CrossRef] [PubMed]
- Chakravarty, J.; Sundar, S. Current and emerging medications for the treatment of leishmaniasis. Expert Opin. Pharmacother. 2019, 20, 1251–1265. [Google Scholar] [CrossRef]
- Mrabet, N.; Khaled, A.; Zeglaoui, F.; Kharfi, M.; Fazaa, B.; Kamoun, M.R. Side effects of meglumine antimoniate in cutaneous leishmaniasis: 15 cases. Tunis Med. 2010, 88, 9–11. [Google Scholar]
- Torres, D.C.; Ribeiro-Alves, M.; Romero, G.A.S.; Dávila, A.M.R.; Cupolillo, E. Assessment of drug resistance related genes as candidate markers for treatment outcome prediction of cutaneous leishmaniasis in Brazil. Acta Trop. 2013, 126, 132–141. [Google Scholar] [CrossRef]
- Briones Nieva, C.A.; Cid, A.G.; Romero, A.I.; García-Bustos, M.F.; Villegas, M.; Bermúdez, J.M. An appraisal of the scientific current situation and new perspectives in the treatment of cutaneous leishmaniasis. Acta Trop. 2021, 221, 105988. [Google Scholar] [CrossRef]
- Ware, J.M.; O’Connell, E.M.; Brown, T.; Wetzler, L.; Talaat, K.R.; Nutman, T.B.; Nash, T.E. Efficacy and tolerability of miltefosine in the treatment of cutaneous leishmaniasis. Clin. Infect. Dis. 2021, 73, e2457–e2562. [Google Scholar] [CrossRef]
- Petrovska, B.B.; Cekovska, S. Extracts from the history and medical properties of garlic. Pharmacogn. Rev. 2010, 4, 106–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Saber Batiha, G.; Magdy Beshbishy, A.; Wasef, L.G.; Elewa, Y.H.A.; Al-Sagan, A.A.; Abd El-Hack, M.E.; Taha, A.E.; Abd-Elhakim, Y.M.; Prasad Devkota, H. Chemical constituents and pharmacological activities of garlic (Allium sativum L.): A review. Nutrients 2020, 12, 872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corzo-Martínez, M.; Corzo, N.; Villamiel, M. Biological properties of onions and garlic. Trends Food Sci. Technol. 2007, 18, 609–625. [Google Scholar] [CrossRef]
- Foroutan-Rad, M.; Tappeh, K.H.; Khademvatan, S. Antileishmanial and immunomodulatory activity of Allium sativum (Garlic): A Review. J. Evid. Based Complement. Altern. Med. 2017, 22, 141–155. [Google Scholar] [CrossRef] [PubMed]
- Bai, H.; Ni, X.; Han, J.; Luo, D.; Hu, Y.; Jin, C.; Li, Z. Phytochemical profiling and allelopathic effect of garlic essential oil on barnyard grass (Echinochloa crusgalli L.). PLoS ONE 2023, 18, e0272842. [Google Scholar] [CrossRef]
- Verma, T.; Aggarwal, A.; Dey, P.; Chauhan, A.K.; Rashid, S.; Chen, K.T.; Sharma, R. Medicinal and therapeutic properties of garlic, garlic essential oil, and garlic-based snack food: An updated review. Front. Nutr. 2023, 10, 1120377. [Google Scholar] [CrossRef]
- Garcia, A.R.; Amaral, A.C.F.; Maria, A.C.B.; Paz, M.M.; Amorim, M.M.B.; Chaves, F.C.M.; Vermelho, A.B.; Nico, D.; Rodrigues, I.A. Antileishmanial screening, cytotoxicity, and chemical composition of essential oils: A special focus on Piper callosum essential oil. Chem. Biodivers. 2023, 20, e202200689. [Google Scholar] [CrossRef]
- Rolón, M.; Vega, C.; Escario, J.A.; Gómez-Barrio, A. Development of resazurin microtiter assay for drug sensibility testing of Trypanosoma cruzi epimastigotes. Parasitol. Res. 2006, 99, 103–107. [Google Scholar] [CrossRef]
- Jordan, J.P.; Hand, C.M.; Markowitz, R.S.; Black, P. Test for chemotherapeutic sensitivity of cerebral gliomas: Use of colorimetric MTT assay. J. Neurooncol. 1992, 14, 19–35. [Google Scholar] [CrossRef]
- Brochot, E.; François, C.; Castelain, S.; Helle, F.; Van-Nhien, A.N.; Duchaussoy, I.; Capron, D.; Nguyen-Khac, E.; Duverlie, G. A new tool to study ribavirin-induced haemolysis. Antivir. Ther. 2012, 17, 1311–1317. [Google Scholar] [CrossRef] [Green Version]
- Garcia, A.R.; Amaral, A.C.F.; Azevedo, M.M.B.; Corte-Real, S.; Lopes, R.C.; Alviano, C.S.; Pinheiro, A.S.; Vermelho, A.B.; Rodrigues, I.A. Cytotoxicity and anti-Leishmania amazonensis activity of Citrus sinensis leaf extracts. Pharm. Biol. 2017, 55, 1780–1786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cyrino, L.T.; Araújo, A.P.; Joazeiro, P.P.; Vicente, C.P.; Giorgio, S. In vivo and in vitro Leishmania amazonensis infection induces autophagy in macrophages. Tissue Cell 2012, 44, 401–408. [Google Scholar] [CrossRef] [PubMed]
- Green, L.C.; Wagner, D.A.; Glogowski, J.; Skipper, P.L.; Wishnok, J.S.; Tannenbaum, S.R. Analysis of nitrate, nitrite, and [15N]nitrate in biological fluids. Anal Biochem. 1982, 126, 131–138. [Google Scholar] [CrossRef] [PubMed]
- Machado, P.A.; Gomes, P.S.; Midlej, V.; Coimbra, E.S.; Guedes, H.L.M. PF-429242, a Subtilisin Inhibitor, Is Effective in vitro Against Leishmania infantum. Front Microbiol. 2021, 12, 583834. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.F.; Ouyang, S.H.; Tu, L.F.; Wang, X.; Yuan, W.L.; Wang, G.E.; Wu, Y.P.; Duan, W.J.; Yu, H.M.; Fang, Z.Z.; et al. Caffeine protects skin from oxidative stress-induced senescence through the activation of autophagy. Theranostics 2018, 8, 5713–5730. [Google Scholar] [CrossRef]
- Garcia, A.R.; Silva-Luiz, Y.P.G.; Alviano, C.S.; Alviano, D.S.; Vermelho, A.B.; Rodrigues, I.A. The Natural Alkaloid Tryptanthrin Induces Apoptosis-like Death in Leishmania spp. Trop. Med. Infect. Dis. 2022, 7, 112. [Google Scholar] [CrossRef]
- Tocmo, R.; Liang, D.; Lin, Y.; Huang, D. Chemical and biochemical mechanisms underlying the cardioprotective roles of dietary organopolysulfides. Front. Nutr. 2015, 2, 1. [Google Scholar] [CrossRef] [Green Version]
- Marcinkowska, M.A.; Jeleń, H.H. Role of Sulfur Compounds in Vegetable and Mushroom Aroma. Molecules 2022, 27, 6116. [Google Scholar] [CrossRef]
- Fukushima-Sakuno, E. Bioactive small secondary metabolites from the mushrooms Lentinula edodes and Flammulina velutipes. J. Antibiotics. 2020, 73, 687–696. [Google Scholar] [CrossRef]
- Morita, K.; Kobayashi, S. Isolation, structure, and synthesis of lenthionine and its analogs. Chem. Pharm. Bull. 1967, 15, 988–993. [Google Scholar] [CrossRef] [Green Version]
- Battal, M.; Kartal, A.; Citgez, B.; Yilmaz, B.; Akcakaya, A.; Karatepe, O. Impact of allyl disulfide on oxidative damage and liver regeneration in an experimental hepatectomy model. Chirurgia 2015, 110, 117–122. [Google Scholar] [PubMed]
- Chung, L.Y. The antioxidant properties of garlic compounds: Allyl cysteine, alliin, allicin, and allyl disulfide. J. Med. Food. 2006, 9, 205–213. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.X.; Liu, H.; Wang, S.; Zhang, C.L.; Guo, F.F.; Zeng, T. Diallyl disulfide ameliorates ethanol-induced liver steatosis and inflammation by maintaining the fatty acid catabolism and regulating the gut-liver axis. Food Chem. Toxicol. 2022, 164, 113108. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Wang, Y.; Zhang, J.; Liu, B.; Li, G.; Xin, S.; Xu, K. Diallyl disulfide attenuates non-alcoholic steatohepatitis by suppressing key regulators of lipid metabolism, lipid peroxidation and inflammation in mice. Mol. Med. Rep. 2019, 20, 1363–1372. [Google Scholar] [CrossRef]
- Zhang, X.N.; Zhao, N.; Guo, F.F.; Wang, Y.R.; Liu, S.X.; Zeng, T. Diallyl disulfide suppresses the lipopolysaccharide-driven inflammatory response of macrophages by activating the Nrf2 pathway. Food Chem. Toxicol. 2022, 59, 112760. [Google Scholar] [CrossRef]
- Alnomasy, S.F. In vitro and in vivo anti-Toxoplasma effects of Allium sativum essential oil against Toxoplasma gondii RH strain. Infect. Drug Resist. 2021, 14, 5057–5068. [Google Scholar] [CrossRef]
- Wabwoba, B.W.; Anjili, C.O.; Ngeiywa, M.M.; Ngure, P.K.; Kigondu, E.M.; Ingonga, J.; Makwali, J. Experimental chemotherapy with Allium sativum (Liliaceae) methanolic extract in rodents infected with Leishmania major and Leishmania donovani. J. Vector Borne Dis. 2010, 47, 160–167. [Google Scholar]
- Mahmoudvand, H.; Sepahvand, P.; Jahanbakhsh, S.; Azadpour, M. Evaluation of the antileishmanial and cytotoxic effects of various extracts of garlic (Allium sativum) on Leishmania tropica. J. Parasit. Dis. 2016, 40, 423–426. [Google Scholar] [CrossRef] [Green Version]
- Krstin, S.; Sobeh, M.; Braun, M.S.; Wink, M. Anti-parasitic activities of Allium sativum and Allium cepa against Trypanosoma b. brucei and Leishmania tarentolae. Medicines 2018, 5, 37. [Google Scholar] [CrossRef] [Green Version]
- Ghazanfari, T.; Hassan, Z.M.; Khamesipour, A. Enhancement of peritoneal macrophages phagocytic activity against Leishmania major by garlic (Allium sativum) treatment. J. Ethnopharmacol. 2006, 103, 333–337. [Google Scholar] [CrossRef]
- Gamboa-León, M.R.; Aranda-González, I.; Mut-Martín, M.; García-Miss, M.R.; Dumonteil, E. In vivo and in vitro control of Leishmania mexicana due to garlic-induced NO production. Scand. J. Immunol. 2007, 66, 508–514. [Google Scholar] [CrossRef]
- Rodrigues, I.A.; Garcia, A.R.; Paz, M.M.; Grilo Junior, R.G.D.; Amaral, A.C.F.; Pinheiro, A.S. Polyamine and trypanothione pathways as targets for novel antileishmanial drugs. In Antiprotozoal Drug Development and Delivery. Topics in Medicinal Chemistry; Vermelho, A.B., Supuran, C.T., Eds.; Springer: Cham, Switzerland, 2021; Volume 39. [Google Scholar] [CrossRef]
- Shin, I.S.; Hong, J.; Jeon, C.M.; Shin, N.R.; Kwon, O.K.; Kim, H.S.; Kim, J.C.; Oh, S.R.; Ahn, K.S. Diallyl-disulfide, an organosulfur compound of garlic, attenuates airway inflammation via activation of the Nrf-2/HO-1 pathway and NF-kappaB suppression. Food Chem. Toxicol. 2013, 62, 506–513. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.P.; Chen, Y.H. Differential effects of organosulfur compounds from garlic oil on nitric oxide and prostaglandin E2 in stimulated macrophages. Nutrition 2005, 21, 530–536. [Google Scholar] [CrossRef] [PubMed]
- Miranda-Sapla, M.M.; Tomiotto-Pellissier, F.; Assolini, J.P.; Carloto, A.C.M.; Bortoleti, B.T.D.S.; Gonçalves, M.D.; Tavares, E.R.; Rodrigues, J.H.D.S.; Simão, A.N.C.; Yamauchi, L.M.; et al. Trans-Chalcone modulates Leishmania amazonensis infection in vitro by Nrf2 overexpression affecting iron availability. Eur. J. Pharmacol. 2019, 853, 275–288. [Google Scholar] [CrossRef] [PubMed]
- Cascajosa-Lira, A.; Andreo-Martínez, P.; Prieto, A.I.; Banos, A.; Guillamón, E.; Jos, E.; Cameán, A.M. In vitro toxicity studies of bioactive organosulfur compounds from Allium spp. with potential application in the agri-food industry: A review. Foods 2022, 11, 2620. [Google Scholar] [CrossRef] [PubMed]
- Munday, R.; Munday, J.S.; Munday, C.M. Comparative effects of mono-, di-, tri-, and tetrasulfides derived from plants of the Allium family: Redox cycling in vitro and hemolytic activity and Phase 2 enzyme induction in vivo. Free Radic. Biol. Med. 2003, 34, 1200–1211. [Google Scholar] [CrossRef]
- Katsuno, K.; Burrows, J.N.; Duncan, K.; Hooft Van Huijsduijnen, R.; Kaneko, T.; Kita, K.; Mowbray, C.E.; Schmatz, D.; Warner, P.; Slingsby, B.T. Hit and lead criteria in drug discovery for infectious diseases of the developing world. Nat Rev. Drug Discov. 2015, 14, 751–758. [Google Scholar] [CrossRef]
- Loset, J.R.; Brun, R.; Wenzler, T.; Kaiser, M.; Yardley, V. Drug Screening for Kinetoplastid Diseases: A Training Manual for Screening in Neglected Deseases. DNDi and Pan-Asian Screening Network. 2009. Available online: https://dndi.org/wp-content/uploads/2009/04/kinetoplastid_drug_screening_manual_final.pdf (accessed on 22 November 2022).
- Vannier-Santos, M.A.; Martiny, A.; Lins, U.; Urbina, J.A.; Borges, V.M.; de Souza, W. Impairment of sterol biosynthesis leads to phosphorus and calcium accumulation in Leishmania acidocalcisomes. Microbiology 1999, 145, 3213–3220. [Google Scholar] [CrossRef] [Green Version]
- Kessler, R.L.; Soares, M.J.; Probst, C.M.; Krieger, M.A. Trypanosoma cruzi response to sterol biosynthesis inhibitors: Morphophysiological alterations leading to cell death. PLoS ONE 2013, 8, e55497. [Google Scholar] [CrossRef] [Green Version]
- De Medeiros, M.D.; Da Silva, A.C.; Citó, A.M.G.L.; Borges, A.R.; De Lima, S.G.; Lopes, J.A.D.; Figueiredo, R.C.B.Q. In vitro antileishmanial activity and cytotoxicity of essential oil from Lippia sidoides Cham. Parasitol. Int. 2011, 60, 237–241. [Google Scholar] [CrossRef]
- Ledezma, E.; Jorquera, A.; Bendezú, H.; Vivas, J.; Pérez, G. Antiproliferative and leishmanicidal effect of ajoene on various Leishmania species: Ultrastructural study. Parasitol. Res. 2002, 88, 748–753. [Google Scholar] [CrossRef] [PubMed]
- Machado, M.; Dinis, A.M.; Santos-Rosa, M.; Alves, V.; Salgueiro, L.; Cavaleiro, C.; Sousa, M.C. Activity of Thymus capitellatus volatile extract, 1,8-cineole and borneol against Leishmania species. Vet. Parasitol. 2014, 200, 39–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Demarchi, I.G.; Thomazella, M.V.; Terron, M.S.; Lopes, L.; Gazim, Z.C.; Cortez, D.A.G.; Donatti, L.; Aristides, S.M.A.; Silveira, T.G.V.; Lonardoni, M.V.C. Antileishmanial activity of essential oil and 6,7-dehydroroyleanone isolated from Tetradenia riparia. Exp. Parasitol. 2015, 157, 128–137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Machado, M.; Pires, P.; Dinis, A.M.; Santos-Rosa, M.; Alves, V.; Salgueiro, L.; Cavaleiro, C.; Sousa, M.C. Monoterpenic aldehydes as potential anti-Leishmania agents: Activity of Cymbopogon citratus and citral on L. infantum, L. tropica and L. major. Exp. Parasitol. 2012, 130, 223–231. [Google Scholar] [CrossRef]
- Niemann, A.; Baltes, J.; Elsasser, H.P. Fluorescence properties and staining behavior of monodansylpentane, a structural a homologue of the lysosomotropic agent monodansylcadaverine. J. Histochem. Cytochem. 2001, 49, 177–185. [Google Scholar] [CrossRef] [Green Version]
- Jiménez-Ruiz, A.; Alzate, J.F.; Macleod, E.T.; Lüder, C.G.; Fasel, N.; Hurd, H. Apoptotic markers in protozoan parasites. Parasit. Vectors 2010, 3, 104. [Google Scholar] [CrossRef] [Green Version]
- Sen, N.; Das, B.B.; Ganguly, A.; Mukherjee, T.; Tripathi, G.; Bandyopadhyay, S.; Rakshit, S.; Sen, T.; Majumder, H.K. Camptothecin induced mitochondrial dysfunction leading to programmed cell death in unicellular hemoflagellate Leishmania donovani. Cell Death Differ. 2004, 11, 924–936. [Google Scholar] [CrossRef] [Green Version]
- Perry, S.W.; Norman, J.P.; Barbieri, J.; Brown, E.B.; Gelbard, H.A. Mitochondrial membrane potential probes and the proton gradient: A practical usage guide. Biotechniques 2011, 50, 98–115. [Google Scholar] [CrossRef]
- Corral, M.J.; Benito-Peña, E.; Jiménez-Antón, M.D.; Cuevas, L.; Moreno-Bondi, M.C.; Alunda, J.M. Allicin induces calcium and mitochondrial dysregulation causing necrotic death in Leishmania. PLoS Negl. Trop. Dis. 2016, 10, e0004525. [Google Scholar] [CrossRef] [Green Version]
- Manikandan, P.; Nagini, S. Cytochrome P450 structure, function and clinical significance: A review. Curr. Drug Targets 2018, 19, 38–54. [Google Scholar] [CrossRef]
- Lu, Y.; Zhang, M.; Huang, D. Dietary organosulfur-containing compounds and their health-promotion mechanisms. Annu. Rev. Food Sci. Technol. 2022, 13, 287–313. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
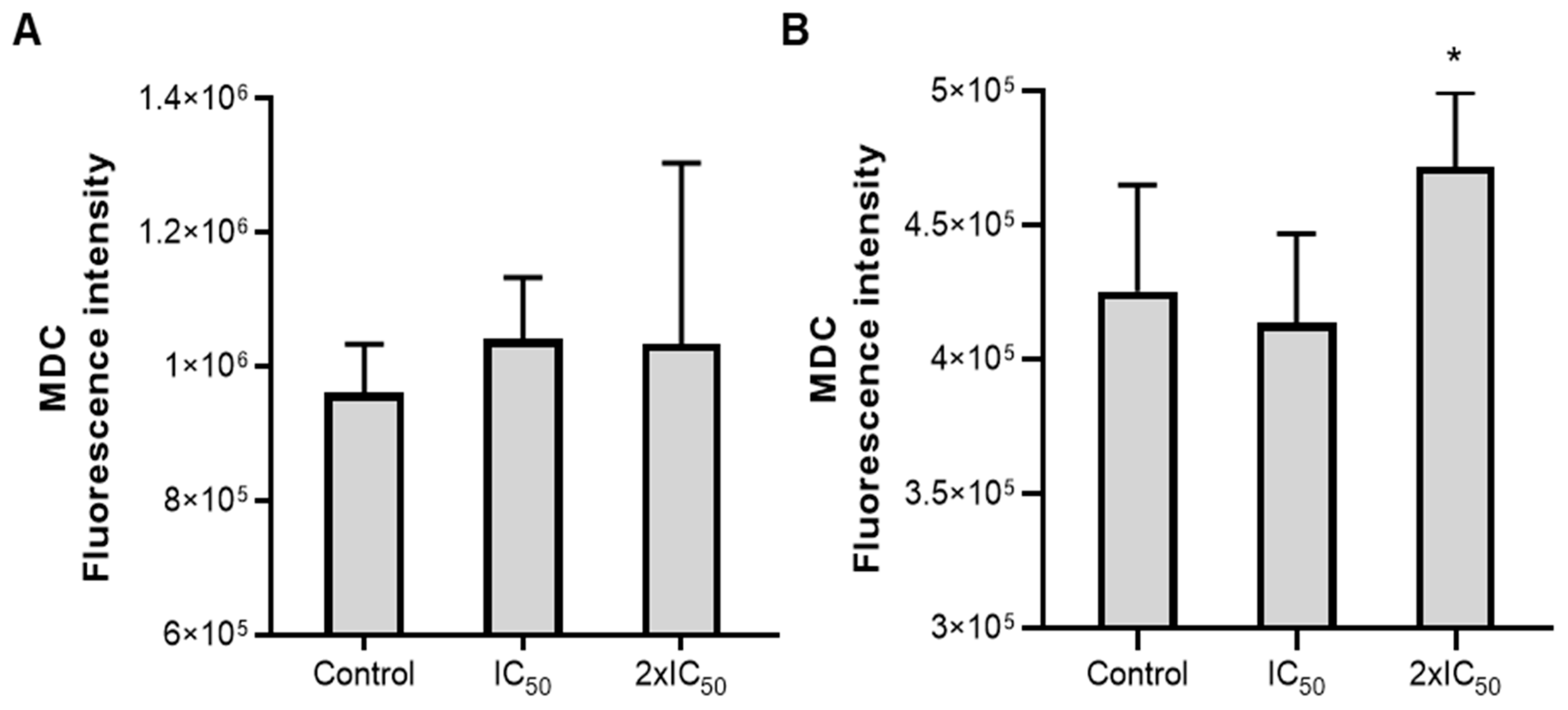{kind=link}
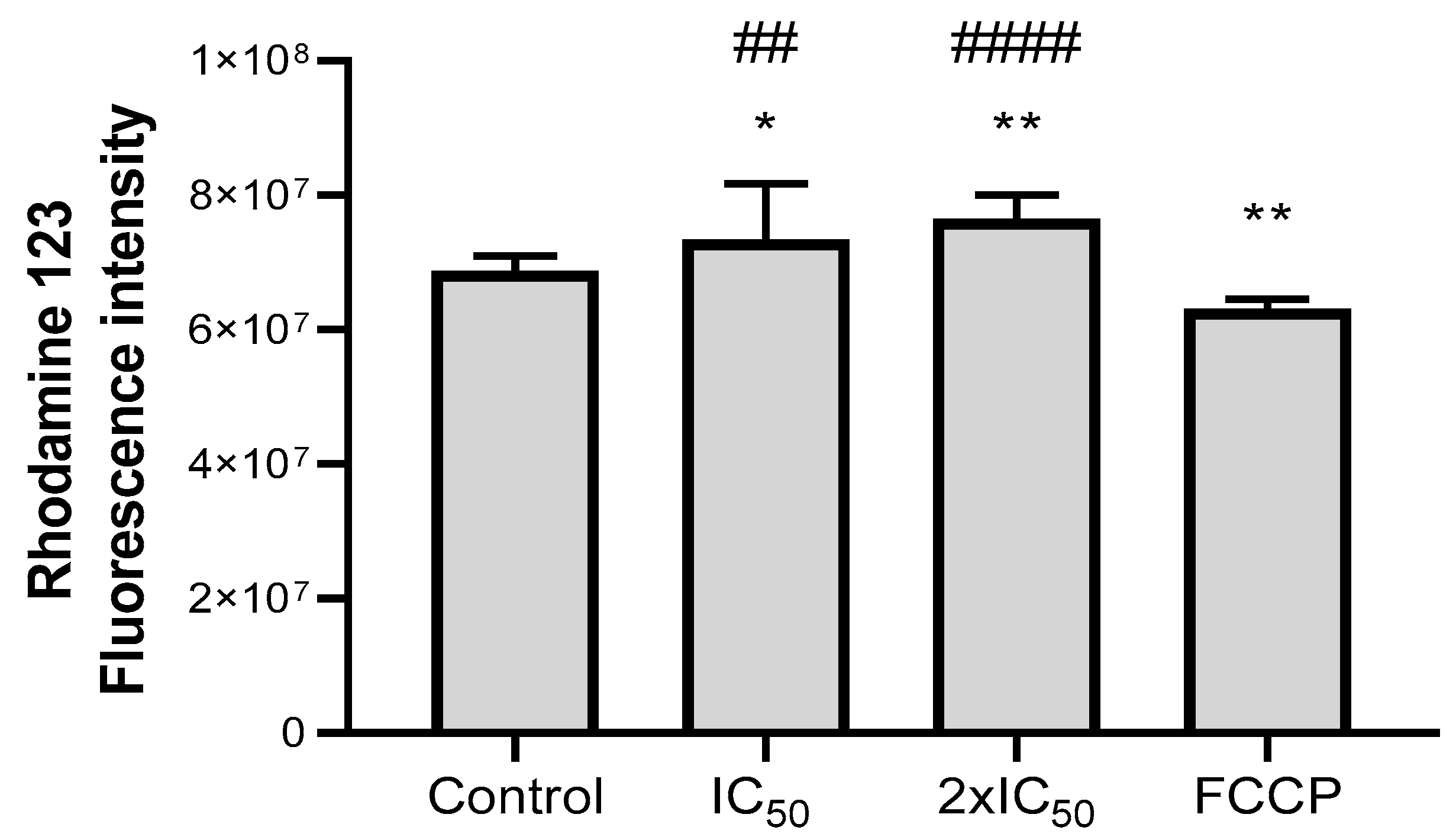{kind=link}
| Compound | RT (min) | Area (%) |
|---|---|---|
| Diallyl sulfide | 10.103 | 4 |
| N, N’-dimethylthiourea | 14.434 | 8.7 |
| Diallyl disulfide | 19.909 | 16.75 |
| Diallyl methyl trisulfide | 22.353 | 3.61 |
| 1,3,5-Trithiane | 23.148 | 8.7 |
| Hexahydro-1,2,4,5-tetrazine-3,6-dithione | 23.541 | 2.24 |
| 3-vinyl-1,2-dithiocyclohex-4-ene | 24.455 | 1.2 |
| 1,3-Dithiolane-2-thione | 24.93 | 7.15 |
| Diallyl trisulfide | 28.667 | 1.4 |
| 1,2,4,6-tetrathiepane | 31.438 | 24.84 |
| 3,5-Diethyl-1,2,4-trithiolane | 33.251 | 0.9 |
| Isobutyl isothiocyanate | 33.622 | 0.2 |
| Propenylpropytrisulfide | 33.875 | 2.4 |
| 2-(2-thia-4-pentenyl)-1-thia-cyclohex-5-ene | 36.177 | 2 |
| 3H-1,2,4-Triazole-3-thione, 4,5-dihydro-4-methy | 38.22 | 1.92 |
| n-Butyl isothiocyanate | 38.532 | 4.84 |
| 4,5-dimethyl thiazole | 43.465 | 1.23 |
| Total | - | 92.08 |
| Drugs | Promastigote | Amastigoteaxe | Amastigoteint |
|---|---|---|---|
| ASEO | 1.76 ± 0.37 a,A | 3.46 ± 0.43 a,A | 3.77 ± 0.74 a,A |
| AmB | 0.6 ± 0.01 a,B | 1.3 ± 0.5 a,A | 0.75 ± 0.03 a,B |
| Mammalian Cells | ASEO | Amphotericin B | ||||
|---|---|---|---|---|---|---|
| CC50 [µg/mL] | SI Amaaxe | SI Amaint | CC50 [µg/mL] | SI Amaaxe | SI Amaint | |
| MØ | 48.94 ± 3.78 | 14.14 | 13 | 18 ± 4.5 | 13.84 | 24 |
| RAW 264.7 | 99 ± 9.7 | 28.6 | 26.25 | n.d. | n.d. | n.d. |
| VERO | >2000 | >500 | >500 | n.d. | n.d. | n.d. |
| hFB | 557.4 ± 89.3 | 161 | 147.8 | 83.05 ± 0.65 | 63.88 | 110.7 |
| RBC | >2000 | >500 | >500 | n.d. | n.d. | n.d. |
| 1,2,4,6-Tetrathiepane | Diallyl Disulfide | Miltefosine | |
|---|---|---|---|
| SP (logKp, cm/h) | −1.3 | −1.15 | −0.73 |
| S (mg/L) | 37.93 | 170.86 | 270.27 |
| BS (mg/L) | 0.001 | 24 | 25.41 |
| PPB (%) | 86.41 | 98 | 86.2 |
| Pgp inhibition | non | inhibitor | inhibitor |
| MDCK (nm/s) | 0.416 | 1.08 | 54.6 |
| HIA (%) | 98 | 98.17 | 98.35 |
| CYP inhibition | 3A4, 2C9, 2C19 | 2C9 | 2D6 |
| CYP substrate | 9A4 (weakly) | 3A4, 2D6 | 3A4 |
| ICP (nm/s) | 56.24 | 22.01 | 21.7 |
| BBB (brain/blood) | 0.93 | 24.72 | 0.23 |
| Rules of 5 | suitable | suitable | suitable |
| Ames test | mutagen | mutagen | non |
| CarM | negative | positive | negative |
| CarR | positive | positive | positive |
| hERG inhibition | medium risk | medium risk | low risk |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Garcia, A.R.; Amorim, M.M.B.; Amaral, A.C.F.; da Cruz, J.D.; Vermelho, A.B.; Nico, D.; Rodrigues, I.A. Anti-Leishmania amazonensis Activity, Cytotoxic Features, and Chemical Profile of Allium sativum (Garlic) Essential Oil. Trop. Med. Infect. Dis. 2023, 8, 375. https://doi.org/10.3390/tropicalmed8070375
Garcia AR, Amorim MMB, Amaral ACF, da Cruz JD, Vermelho AB, Nico D, Rodrigues IA. Anti-Leishmania amazonensis Activity, Cytotoxic Features, and Chemical Profile of Allium sativum (Garlic) Essential Oil. Tropical Medicine and Infectious Disease. 2023; 8(7):375. https://doi.org/10.3390/tropicalmed8070375
Chicago/Turabian StyleGarcia, Andreza R., Mariana M. B. Amorim, Ana Claudia F. Amaral, Jefferson D. da Cruz, Alane B. Vermelho, Dirlei Nico, and Igor A. Rodrigues. 2023. "Anti-Leishmania amazonensis Activity, Cytotoxic Features, and Chemical Profile of Allium sativum (Garlic) Essential Oil" Tropical Medicine and Infectious Disease 8, no. 7: 375. https://doi.org/10.3390/tropicalmed8070375