Harzianic Acid Activity against Staphylococcus aureus and Its Role in Calcium Regulation

 ,
,  , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Results and Discussion
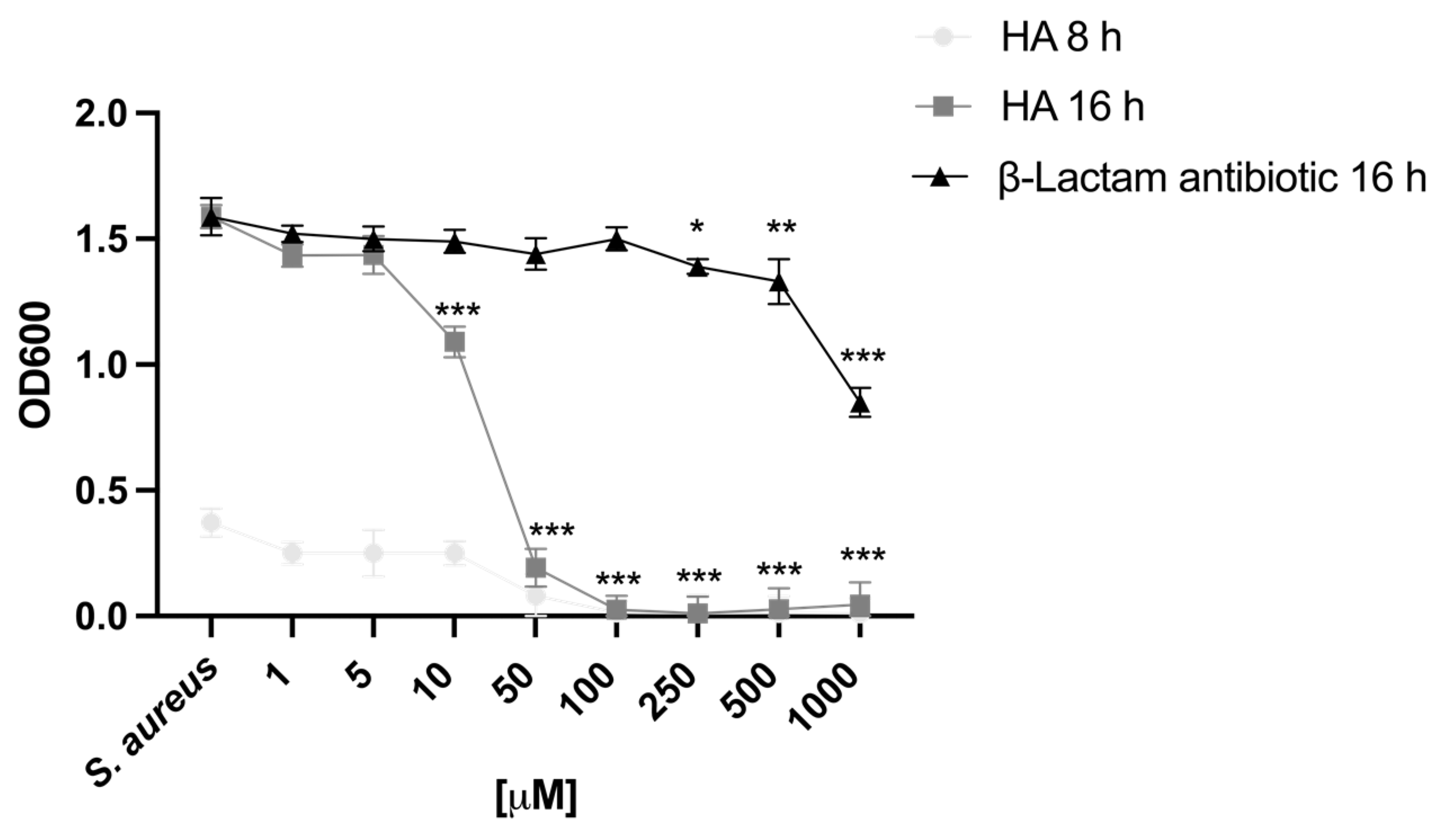
2.1. Harzianic Acid Inhibits Staphylococcus aureus Growth
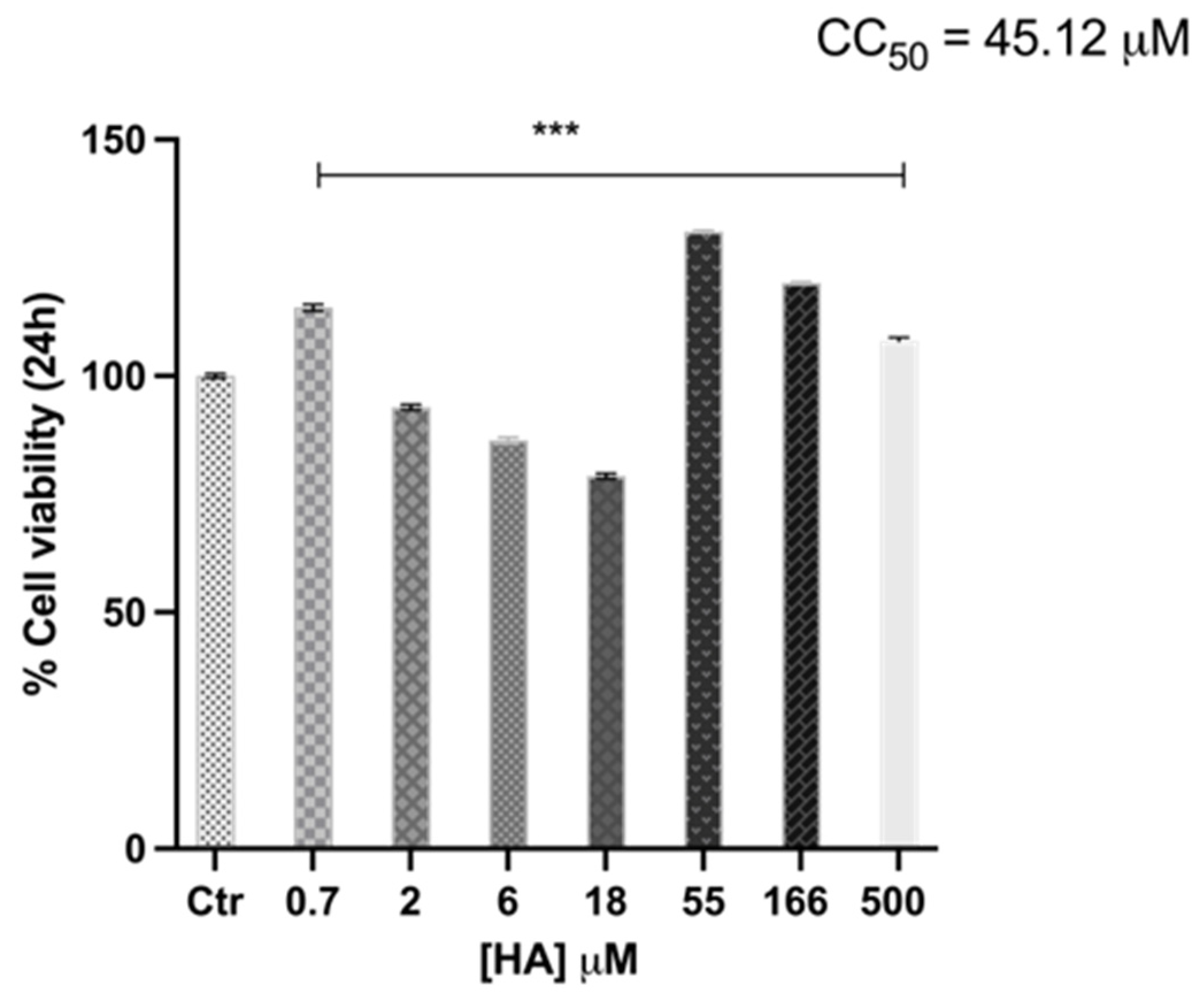
2.2. Harzianic Acid Does Not Alter HaCaT Cell Viability
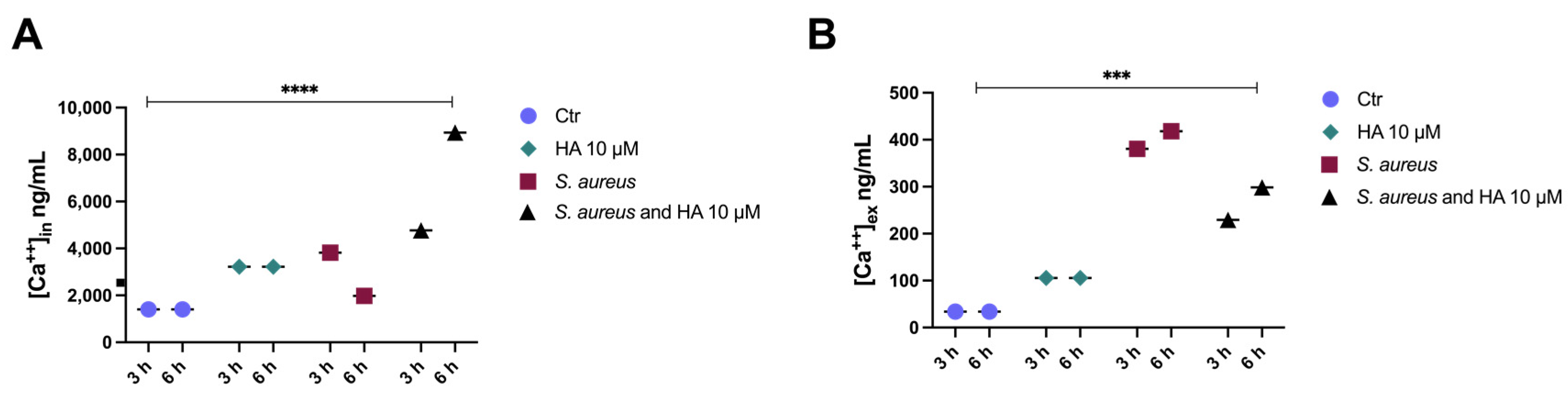
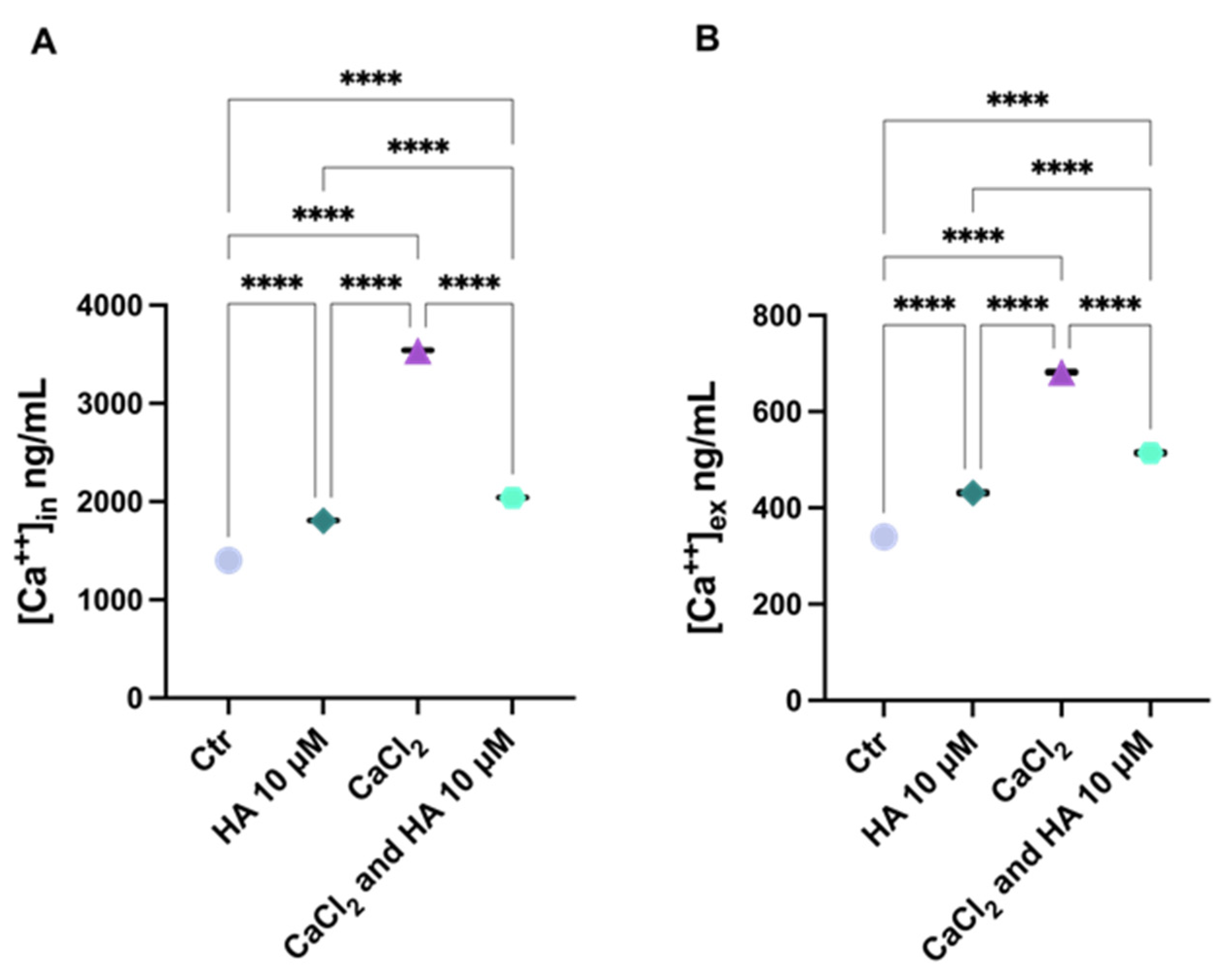
2.3. Harzianic Acid Controls the Cell Host Ca2+ Movements
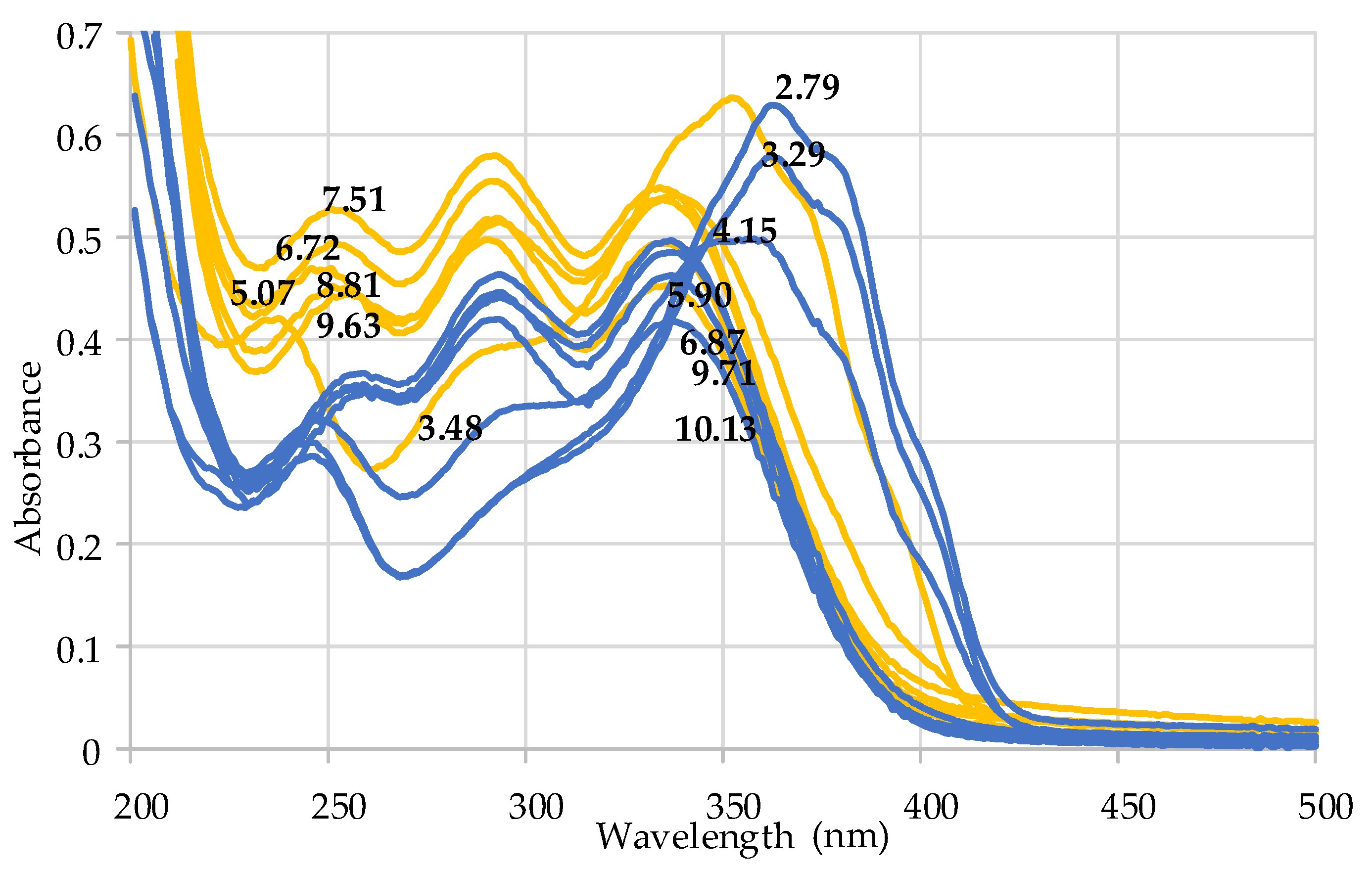
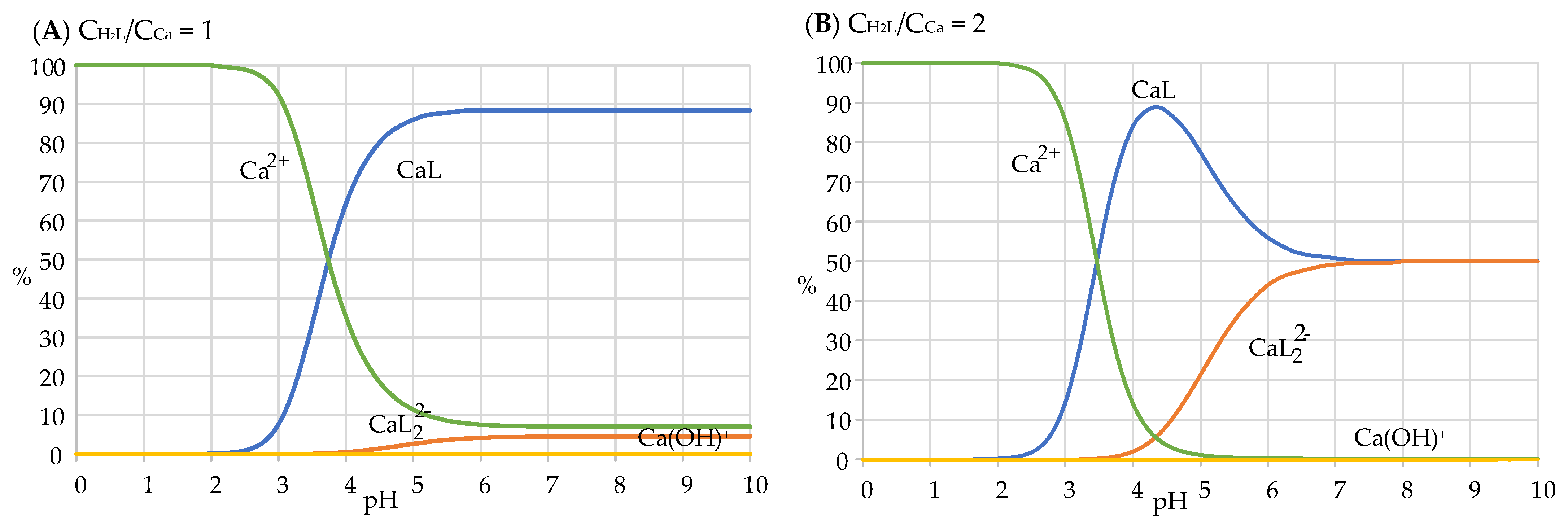
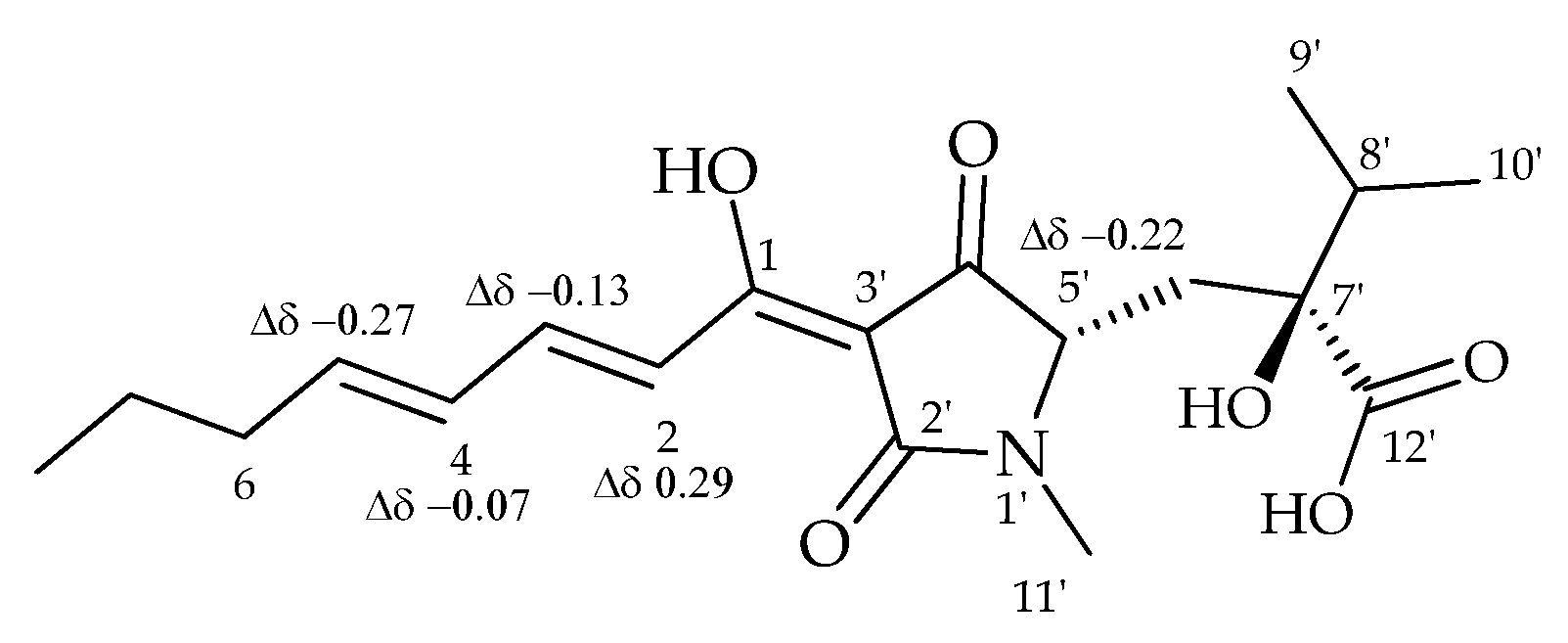
2.4. Coordination Properties of Harzianic Acid toward Ca2+
3. Conclusions
4. Materials and Methods
4.1. Harzianic Acid Production
4.2. Cell Culture Conditions
4.3. Staphylococcus aureus Growth Conditions
4.4. Antibacterial Activity
4.5. Cell Viability Assay
4.6. Infection by Staphylococcus aureus of Eukaryotic Cells
4.7. Extracellular Ca2+ Supplementation
4.8. Ca2+ Measurement by Atomic Adsorption Spectroscopy
4.9. Ca2+ Measurement by Colorimetric Method
4.10. Reagents and Their Analysis
4.11. HPLC-ESI-Q-TOF Analysis
4.12. Preparation of Test Solutions for UV–Vis Spectrophotometric Measurements
4.13. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Berridge, M.J.; Lipp, P.; Bootman, M.D. The versatility and universality of calcium signaling. Nat. Rev. Mol. Cell 2000, 1, 11–21. [Google Scholar] [CrossRef]
- Edel, K.H.; Kudla, J. Increasing complexity and versatility: How the calcium signaling toolkit was shaped during plant land colonization. Cell Calcium 2015, 57, 231–246. [Google Scholar] [CrossRef]
- Permyakov, E.A.; Kretsinger, R.H. Cell signaling, beyond cytosolic calcium in eukaryotes. J. Inorg. Biochem. 2009, 103, 77–86. [Google Scholar] [CrossRef]
- Clapham, D.E. Calcium signaling. Cell 1995, 80, 259–268. [Google Scholar] [CrossRef] [Green Version]
- Bose, J.; Pottosin, I.I.; Shabala, S.S.; Palmgren, M.G.; Shabala, S. Calcium efflux systems in stress signaling and adaptation in plants. Front. Plant Sci. 2011, 2, 85. [Google Scholar] [CrossRef] [Green Version]
- Weiss, N.; Koschak, A. (Eds.) Pathologies of Calcium Channels; Springer: Berlin, Germany, 2014. [Google Scholar]
- Tran Van Nhieu, G.; Dupont, G.; Combettes, L. Ca2+ signals triggered by bacterial pathogens and microdomains. Biochim. Biophys. Acta Mol. Cell Res. 2018, 1865 Pt B, 1838–1845. [Google Scholar] [CrossRef]
- David, M.Z.; Daum, R.S. Treatment of Staphylococcus aureus Infections. In Staphylococcus aureus. Current Topics in Microbiology and Immunology; Bagnoli, F., Rappuoli, R., Grandi, G., Eds.; Springer: Berlin, Germany, 2017; Volume 409, pp. 325–383. [Google Scholar] [CrossRef]
- Tong, S.Y.; Davis, J.S.; Eichenberger, E.; Holland, T.L.; Fowler, V.G., Jr. Staphylococcus aureus infections: Epidemiology, pathophysiology, clinical manifestations, and management. Clin. Microbiol. Rev. 2015, 28, 603–661. [Google Scholar] [CrossRef] [Green Version]
- Olaniyi, R.; Pozzi, C.; Grimaldi, L.; Bagnoli, F. Staphylococcus aureus-Associated Skin and Soft Tissue Infections: Anatomical Localization, Epidemiology, Therapy and Potential Prophylaxis. In Staphylococcus aureus. Current Topics in Microbiology and Immunology; Bagnoli, F., Rappuoli, R., Grandi, G., Eds.; Springer: Berlin, Germany, 2016; Volume 409, pp. 199–227. [Google Scholar] [CrossRef]
- Peraro, M.; van der Goot, F. Pore-forming toxins: Ancient, but never really out of fashion. Nat. Rev. Microbiol. 2016, 14, 77–92. [Google Scholar] [CrossRef]
- Marchi, S.; Morroni, G.; Pinton, P.; Galluzzi, L. Control of host mitochondria by bacterial pathogens. Trends Microbiol. 2022, 30, 452–465. [Google Scholar] [CrossRef]
- Eichstaedt, S.; Gäbler, K.; Below, S.; Müller, C.; Kohler, C.; Engelmann, S.; Hildebrandt, P.; Völker, U.; Hecker, M.; Hildebrandt, J.P. Effects of Staphylococcus aureus-hemolysin A on calcium signaling in immortalized human airway epithelial cells. Cell Calcium 2009, 45, 165–176. [Google Scholar] [CrossRef]
- Singh, R.; Kumar, M.; Mittal, A.; Mehta, P.K. Microbial metabolites in nutrition, healthcare and agriculture. 3 Biotech 2017, 7, 15. [Google Scholar] [CrossRef] [Green Version]
- Ramírez-Rendon, D.; Passari, A.K.; Ruiz-Villafán, B.; Rodríguez-Sanoja, R.; Sánchez, S.; Demain, A.L. Impact of novel microbial secondary metabolites on the pharma industry. Appl. Microbiol. Biotechnol. 2022, 106, 1855–1878. [Google Scholar] [CrossRef]
- Atanasov, A.G.; Zotchev, S.B.; Dirsch, V.M.; International Natural Product Sciences Taskforce; Supuran, C.T. Natural products in drug discovery: Advances and opportunities. Nat. Rev. Drug Discov. 2021, 20, 200–216. [Google Scholar] [CrossRef]
- Lucke, M.; Correa, M.G.; Levy, A. The Role of Secretion Systems, Effectors, and Secondary Metabolites of Beneficial Rhizobacteria in Interactions with Plants and Microbes. Front. Plant Sci. 2020, 11, 589416. [Google Scholar] [CrossRef]
- Sinno, M.; Ranesi, M.; Di Lelio, I.; Iacomino, G.; Becchimanzi, A.; Barra, E.; Molisso, D.; Pennacchio, F.; Digilio, M.C.; Vitale, S.; et al. Selection of Endophytic Beauveria bassiana as a Dual Biocontrol Agent of Tomato Pathogens and Pests. Pathogens 2021, 10, 1242. [Google Scholar] [CrossRef]
- Harman, G.E.; Howell, C.R.; Viterbo, A.; Chet, I.; Lorito, M. Trichoderma species—Opportunistic, avirulent plant symbionts. Nat. Rev. Microbiol. 2004, 2, 43–56. [Google Scholar] [CrossRef]
- Vinale, F.; Sivasithamparam, K.; Ghisalberti, E.L.; Marra, R.; Woo, S.L.; Lorito, M. Trichoderma–plant–pathogen interactions. Soil Biol. Biochem. 2008, 40, 1–10. [Google Scholar] [CrossRef]
- Keswani, C.; Mishra, S.; Sarma, B.K.; Singh, S.P.; Singh, H.B. Unraveling the efficient applications of secondary metabolites of various Trichoderma spp. Appl. Microbiol. Biotechnol. 2014, 98, 533–544. [Google Scholar] [CrossRef]
- Vinale, F.; Sivasithamparam, K. Beneficial effects of Trichoderma secondary metabolites on crops. Phytother. Res. 2020, 34, 2835–2842. [Google Scholar] [CrossRef]
- Ramírez-Valdespino, C.A.; Casas-Flores, S.; Olmedo-Monfil, V. Trichoderma as a Model to Study Effector-Like Molecules. Front. Microbiol. 2019, 10, 1030. [Google Scholar] [CrossRef]
- Vinale, F.; Nigro, M.; Sivasithamparam, K.; Flematti, G.; Ghisalberti, E.L.; Ruocco, M.; Varlese, R.; Marra, R.; Lanzuise, S.; Eid, A.; et al. Harzianic acid: A novel siderophore from Trichoderma harzianum. FEMS Microbiol. Lett. 2013, 347, 123–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vinale, F.; Flematti, G.; Sivasithamparam, K.; Lorito, M.; Marra, R.; Skelton, B.W.; Ghisalberti, E.L. Harzianic acid, an antifungal and plant growth promoting metabolite from Trichoderma harzianum. J. Nat. Prod. 2009, 72, 2032–2035. [Google Scholar] [CrossRef] [PubMed]
- Dini, I.; Pascale, M.; Staropoli, A.; Marra, R.; Vinale, F. Effect of Selected Trichoderma Strains and Metabolites on Olive Drupes. Appl. Sci. 2021, 11, 8710. [Google Scholar] [CrossRef]
- Manganiello, G.; Sacco, A.; Ercolano, M.R.; Vinale, F.; Lanzuise, S.; Pascale, A.; Napolitano, M.; Lombardi, N.; Lorito, M.; Woo, S.L. Modulation of tomato response to Rhizoctonia solani by Trichoderma harzianum and its secondary metabolite harzianic acid. Front. Microbiol. 2018, 9, 1966. [Google Scholar] [CrossRef]
- Dini, I.; Graziani, G.; Fedele, F.L.; Sicari, A.; Vinale, F.; Castaldo, L.; Ritieni, A. Effects of Trichoderma Biostimulation on the Phenolic Profile of Extra-Virgin Olive Oil and Olive Oil By-Products. Antioxidants 2020, 9, 284. [Google Scholar] [CrossRef] [Green Version]
- De Filippis, A.; Nocera, F.P.; Tafuri, S.; Ciani, F.; Staropoli, A.; Comite, E.; Bottiglieri, A.; Gioia, L.; Lorito, M.; Woo, S.L.; et al. Antimicrobial activity of harzianic acid against Staphylococcus pseudintermedius. Nat. Prod. Res. 2021, 35, 5440–5445. [Google Scholar] [CrossRef]
- De Tommaso, G.; Salvatore, M.M.; Nicoletti, R.; DellaGreca, M.; Vinale, F.; Bottiglieri, A.; Staropoli, A.; Salvatore, F.; Lorito, M.; Iuliano, M.; et al. Bivalent Metal-Chelating Properties of Harzianic Acid Produced by Trichoderma pleuroticola Associated to the Gastropod Melarhaphe neritoides. Molecules 2020, 25, 2147. [Google Scholar] [CrossRef]
- De Tommaso, G.; Salvatore, M.M.; Nicoletti, R.; DellaGreca, M.; Vinale, F.; Staropoli, A.; Salvatore, F.; Lorito, M.; Iuliano, M.; Andolfi, A. Coordination Properties of the Fungal Metabolite Harzianic Acid Toward Toxic Heavy Metals. Toxics 2021, 9, 19. [Google Scholar] [CrossRef]
- Ouyang, X.; Hoeksma, J.; Beenker, W.A.G.; van der Beek, S.; den Hertog, J. Harzianic Acid Has Multi-Target Antimicrobial Activity against Gram-Positive Bacteria; Institute Biology Leiden, Leiden University: Leiden, The Netherlands, 2021; to be submitted. [Google Scholar]
- Yendapally, R.; Hurdle, J.G.; Carson, E.I.; Lee, R.B.; Lee, R.E. N-substituted 3-acetyltetramic acid derivatives as antibacterial agents. J. Med. Chem. 2008, 51, 1487–1491. [Google Scholar] [CrossRef]
- King, M.M.; Kayastha, B.B.; Franklin, M.J.; Patrauchan, M.A. Calcium Regulation of Bacterial Virulence. In Calcium Signaling. Advances in Experimental Medicine and Biology; Islam, M., Ed.; Springer: Cham, Switzerland, 2020; Volume 1131, pp. 827–855. [Google Scholar] [CrossRef]
- McCaig, L.F.; McDonald, L.C.; Mandal, S.; Jernigan, D.B. Staphylococcus aureus-associated skin and soft tissue infections in ambulatory care. Emerg. Infect. Dis. 2006, 12, 1715–1723. [Google Scholar] [CrossRef] [Green Version]
- Stelzner, K.; Winkler, A.C.; Liang, C.; Boyny, A.; Ade, C.P.; Dandekar, T.; Fraunholz, M.J.; Rudel, T. Intracellular Staphylococcus aureus perturbs the host cell Ca2+ homeostasis to promote cell death. mBio 2020, 11, e02250-20. [Google Scholar] [CrossRef] [PubMed]
- Cheung, G.Y.C.; Bae, J.S.; Otto, M. Pathogenicity and virulence of Staphylococcus aureus. Virulence 2021, 12, 547–569. [Google Scholar] [CrossRef] [PubMed]
- Menestrina, G.; Dalla Serra, M.; Comai, M.; Coraiola, M.; Viero, G.; Werner, S.; Colin, D.A.; Monteil, H.; Prévost, G. Ion channels and bacterial infection: The case of beta-barrel pore-forming protein toxins of Staphylococcus aureus. FEBS Lett. 2003, 552, 54–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tengholm, A.; Hellman, B.; Gylfe, E. Mobilization of Ca2+ stores in individual pancreatic β-cells permeabilized or not with digitonin or α-toxin. Cell Calcium 2000, 27, 43–51. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Yang, L. Calcium and magnesium ions are membrane-active against stationary-phase Staphylococcus aureus with high specificity. Sci. Rep. 2016, 6, 20628. [Google Scholar] [CrossRef] [Green Version]
- Büchau, A.S.; Gallo, R.L. Innate immunity and antimicrobial defense systems in psoriasis. Clin. Dermatol. 2007, 25, 616–624. [Google Scholar] [CrossRef] [Green Version]
- Rossol, M.; Pierer, M.; Raulien, N.; Quandt, D.; Meusch, U.; Rothe, K.; Schubert, K.; Schöneberg, T.; Schaefer, M.; Krügel, U.; et al. Extracellular Ca2+ is a danger signal activating the NLRP3 inflammasome through G protein-coupled calcium sensing receptors. Nat. Commun. 2012, 3, 1329. [Google Scholar] [CrossRef] [Green Version]
- De Tommaso, G.; Salvatore, M.M.; Siciliano, A.; Staropoli, A.; Vinale, F.; Nicoletti, R.; DellaGreca, M.; Guida, M.; Salvatore, F.; Iuliano, M.; et al. Interaction of the Fungal Metabolite Harzianic Acid with Rare-Earth Cations (La3+, Nd3+, Sm3+, Gd3+). Molecules 2022, 27, 1959. [Google Scholar] [CrossRef]
- Salvatore, M.M.; Siciliano, A.; Staropoli, A.; Vinale, F.; Nicoletti, R.; DellaGreca, M.; Guida, M.; Salvatore, F.; Iuliano, M.; Andolfi, A.; et al. Interaction of the Fungal Metabolite Harzianic Acid with Rare-Earth Cations (Pr3+, Eu3+, Ho3+, Tm3+). Molecules 2022, 27, 6468. [Google Scholar] [CrossRef]
- Gans, P.; Sabatini, A.; Vacca, A. Investigation of equilibria in solution. Determination of equilibrium constants with the HYPERQUAD suite of programs. Talanta 1996, 43, 1739–1753. [Google Scholar] [CrossRef]
- Balouiri, M.; Sadiki, M.; Ibnsouda, S.K. Methods for in vitro evaluating antimicrobial activity: A review. J. Pharm. Anal. 2016, 6, 71–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cuomo, P.; Medaglia, C.; Allocca, I.; Montone, A.M.I.; Guerra, F.; Cabaro, S.; Mollo, E.; Eletto, D.; Papaianni, M.; Capparelli, R. Caulerpin Mitigates Helicobacter pylori-Induced Inflammation via Formyl Peptide Receptors. Int. J. Mol. Sci. 2021, 22, 13154. [Google Scholar] [CrossRef] [PubMed]
- Fiorito, F.; Irace, C.; Nocera, F.P.; Piccolo, M.; Ferraro, M.G.; Ciampaglia, R.; Tenore, G.C.; Santamaria, R.; De Martino, L. MG-132 interferes with iron cellular homeostasis and alters virulence of bovine herpesvirus 1. Res. Vet. Sci. 2021, 137, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Kolthoff, I.M.; Elving, P.J.; Meehan, E.J. Treatise on Analytical Chemistry; Wiley: Hoboken, NJ, USA, 1978; ISBN 978-0-471-80647-9. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ion | Experimental Mass of Main Isotopic Peak (Da) | Formula | Exact Mass (Da) |
|---|---|---|---|
| Harzianic acid + CaCl2 | |||
| [H2L + H]+ | 366.1929 | C19H28NO6 | 366.1917 |
| [H2L + Na]+ | 388.1750 | C19H27NO6Na | 388.1736 |
| [H2L + K]+ | 404.1404 | C19H27NO6K | 404.1475 |
| [2H2L − H + Ca]+ | 769.3243 | C38H53N2O12Ca | 769.3224 |
| [3H2L − H + Ca]+ | 1134.5079 | C57H80N3O18Ca | 1134.5063 |
| Equilibria | log (Formation Constant) ± 3σ |
|---|---|
| Ca2+ + L2− = CaL Ca2+ + 2L2− = CaL22− | 6.3 ± 0.1 10.2 ± 0.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Staropoli, A.; Cuomo, P.; Salvatore, M.M.; De Tommaso, G.; Iuliano, M.; Andolfi, A.; Tenore, G.C.; Capparelli, R.; Vinale, F. Harzianic Acid Activity against Staphylococcus aureus and Its Role in Calcium Regulation. Toxins 2023, 15, 237. https://doi.org/10.3390/toxins15040237
Staropoli A, Cuomo P, Salvatore MM, De Tommaso G, Iuliano M, Andolfi A, Tenore GC, Capparelli R, Vinale F. Harzianic Acid Activity against Staphylococcus aureus and Its Role in Calcium Regulation. Toxins. 2023; 15(4):237. https://doi.org/10.3390/toxins15040237
Chicago/Turabian StyleStaropoli, Alessia, Paola Cuomo, Maria Michela Salvatore, Gaetano De Tommaso, Mauro Iuliano, Anna Andolfi, Gian Carlo Tenore, Rosanna Capparelli, and Francesco Vinale. 2023. "Harzianic Acid Activity against Staphylococcus aureus and Its Role in Calcium Regulation" Toxins 15, no. 4: 237. https://doi.org/10.3390/toxins15040237