2-Amino-3-Methylimidazo[4,5-f]quinoline Triggering Liver Damage by Inhibiting Autophagy and Inducing Endoplasmic Reticulum Stress in Zebrafish (Danio rerio)


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. IQ Induced Histological Damage in Zebrafish Livers
2.2. IQ Induced Ultrastructural Abnormalities in Zebrafish Livers
2.3. IQ Induced ERS in Zebrafish Livers
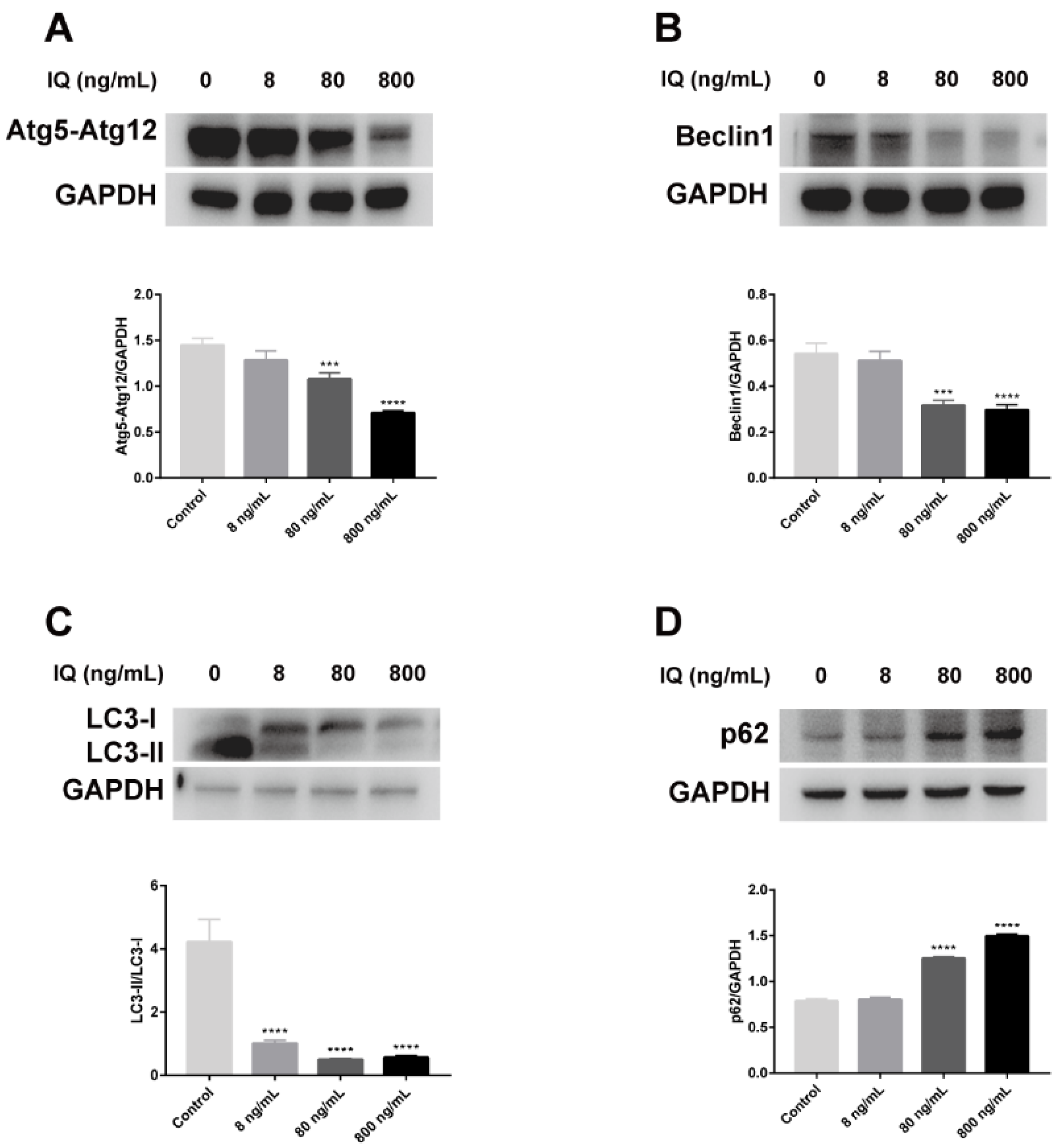
2.4. IQ Inhibited Autophagic Formation in Zebrafish Livers
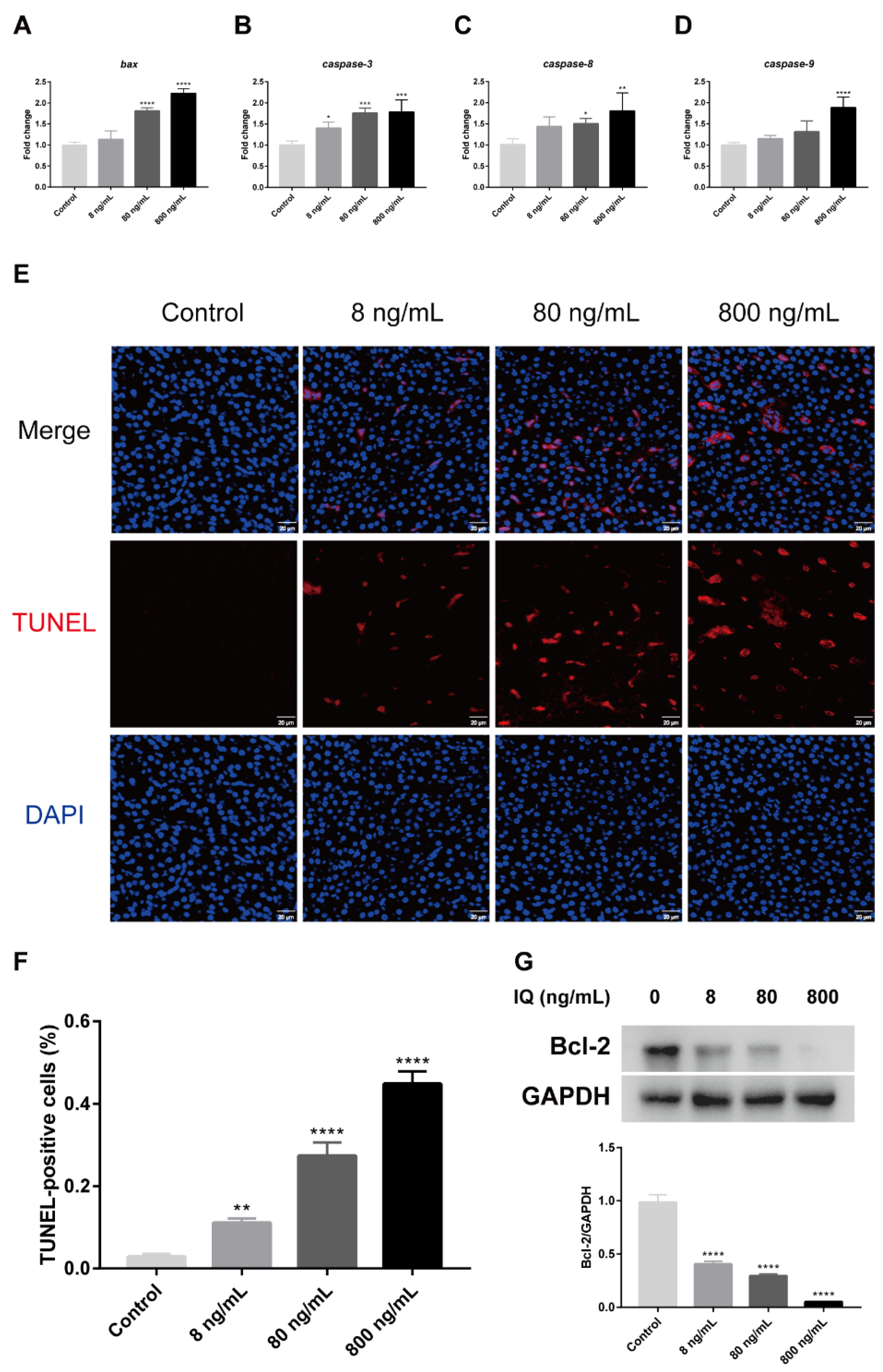
2.5. IQ Induced Hepatocyte Apoptosis in Zebrafish Livers
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Animal Treatment
5.2. Histology
5.3. Transmission Electron Microscopy (TEM)
5.4. Quantitative Real-Time Polymerase Chain Reaction (qPCR)
5.5. Western Blot
5.6. Terminal Deoxynucleotidyl Transferase-Mediated dUTP Nick End Labeling (TUNEL) Assay
5.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Schut, H.A.; Snyderwine, E.G. DNA adducts of heterocyclic amine food mutagens: Implications for mutagenesis and carcinogenesis. Carcinogenesis 1999, 20, 353–368. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Cao, P.; Meng, H.; Li, D.; Zhang, Y.; Li, Y.; Wang, S. Long-term exposure to 2-amino-3-methylimidazo[4,5-f]quinoline can trigger a potential risk of Parkinson’s disease. J. Hazard. Mater. 2021, 412, 125230. [Google Scholar] [CrossRef] [PubMed]
- Immonen, E.; Kummu, M.; Petsalo, A.; Pihlaja, T.; Mathiesen, L.; Nielsen, J.K.; Knudsen, L.E.; Vahakangas, K.; Myllynen, P. Toxicokinetics of the food-toxin IQ in human placental perfusion is not affected by ABCG2 or xenobiotic metabolism. Placenta 2010, 31, 641–648. [Google Scholar] [CrossRef]
- Barzegar, F.; Kamankesh, M.; Mohammadi, A. Heterocyclic aromatic amines in cooked food: A review on formation, health risk-toxicology and their analytical techniques. Food Chem. 2019, 280, 240–254. [Google Scholar] [CrossRef]
- Gibis, M. Heterocyclic Aromatic Amines in Cooked Meat Products: Causes, Formation, Occurrence, and Risk Assessment. Compr. Rev. Food Sci. Food Saf. 2016, 15, 269–302. [Google Scholar] [CrossRef]
- Cruz-Hernandez, A.; Agim, Z.S.; Montenegro, P.C.; McCabe, G.P.; Rochet, J.-C.; Cannon, J.R. Selective dopaminergic neurotoxicity of three heterocyclic amine subclasses in primary rat midbrain neurons. Neurotoxicology 2018, 65, 68–84. [Google Scholar] [CrossRef]
- Li, R.; Tian, J.; Li, W.; Xie, J. Effects of 2-amino-1-methyl-6-phenylimidazo [4, 5-b] pyridine (PhIP) on histopathology, oxidative stress, and expression of c-fos, c-jun and p16 in rat stomachs. Food Chem. Toxicol. 2013, 55, 182–191. [Google Scholar] [CrossRef]
- Zhao, X.; Liu, H.; Zhou, X.; Chen, X.; Hu, N.; Zhang, Y.; Wang, S. 2-Amino-1-methyl-6-phenylimidazo[4,5-b]pyridine Induced Colon Injury by Disrupting the Intestinal Bacterial Composition and Lipid Metabolic Pathways in Rats. J. Agric. Food Chem. 2021, 69, 437–446. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Li, Z.; Qiu, C.; Peng, B.; Zhang, Y.; Sun, H.; Wang, S. 2-Amino-3-methylimidazo[4,5-f]quinoline induced oxidative stress and inflammation via TLR4/MAPK and TLR4/NF-kappaB signaling pathway in zebrafish (Danio rerio) livers. Food Chem. Toxicol. 2021, 157, 112583. [Google Scholar] [CrossRef]
- An, M.Y.; Lee, S.R.; Hwang, H.J.; Yoon, J.G.; Lee, H.J.; Cho, J.A. Antioxidant and Anti-Inflammatory Effects of Korean Black Ginseng Extract through ER Stress Pathway. Antioxidants 2021, 10, 62. [Google Scholar] [CrossRef]
- Chen, H.G.; Han, H.Z.; Li, Y.; Yu, Y.H.; Xie, K.L. Hydrogen alleviated organ injury and dysfunction in sepsis: The role of cross-talk between autophagy and endoplasmic reticulum stress: Experimental research. Int. Immunopharmacol. 2020, 78, 106049. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.M.; Ma, J.Q.; Sun, J.M.; Feng, Z.J.; Cheng, C.; Yang, W.; Jiang, H. Association of changes in ER stress-mediated signaling pathway with lead-induced insulin resistance and apoptosis in rats and their prevention by A-type dimeric epigallocatechin-3-gallate. Food Chem. Toxicol. 2017, 110, 325–332. [Google Scholar] [CrossRef]
- Jia, H.; Liu, N.; Zhang, Y.; Wang, C.; Yang, Y.; Wu, Z. 3-Acetyldeoxynivalenol induces cell death through endoplasmic reticulum stress in mouse liver. Environ. Pollut 2021, 286, 117238. [Google Scholar] [CrossRef]
- Pang, X.; Qiao, Q.; Vonglorkham, S.; Feng, Z.; Pang, L.; Chen, S.; Wang, D.; Lao, L.; Lin, X.; Wei, J. Asiatic acid ameliorates acute hepatic injury by reducing endoplasmic reticulum stress and triggering hepatocyte autophagy. Biomed. Pharm. 2020, 129, 110375. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Fan, Y.; Liu, M.; Zhang, Q.; Guan, S. 1,3-dichloro-2-propanol induced lipid accumulation by blocking autophagy flux in HepG2 cells. Toxicology 2021, 454, 152716. [Google Scholar] [CrossRef] [PubMed]
- Qian, H.; Chao, X.; Williams, J.; Fulte, S.; Li, T.; Yang, L.; Ding, W.X. Autophagy in liver diseases: A review. Mol. Asp. Med. 2021, 100973. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Duan, J.; Yu, Y.; Li, Y.; Liu, X.; Zhou, X.; Ho, K.F.; Tian, L.; Sun, Z. Silica nanoparticles induce autophagy and autophagic cell death in HepG2 cells triggered by reactive oxygen species. J. Hazard. Mater. 2014, 270, 176–186. [Google Scholar] [CrossRef]
- Lei, X.; Liu, Q.; Liu, Q.; Cao, Z.; Zhang, J.; Kuang, T.; Fang, Y.; Liu, G.; Qian, K.; Fu, J.; et al. Camellia oil (Camellia oleifera Abel.) attenuates CCl4-induced liver fibrosis via suppressing hepatocyte apoptosis in mice. Food Funct. 2020, 11, 4582–4590. [Google Scholar] [CrossRef]
- Liu, H.; Lai, W.; Liu, X.; Yang, H.; Fang, Y.; Tian, L.; Li, K.; Nie, H.; Zhang, W.; Shi, Y.; et al. Exposure to copper oxide nanoparticles triggers oxidative stress and endoplasmic reticulum (ER)-stress induced toxicology and apoptosis in male rat liver and BRL-3A cell. J. Hazard. Mater. 2021, 401, 123349. [Google Scholar] [CrossRef]
- Zhao, Y.; Li, Y.; Wang, J.; Manthari, R.K.; Wang, J. Fluoride induces apoptosis and autophagy through the IL-17 signaling pathway in mice hepatocytes. Arch. Toxicol. 2018, 92, 3277–3289. [Google Scholar] [CrossRef]
- Yang, Y.; Dong, F.; Liu, X.; Xu, J.; Wu, X.; Liu, W.; Zheng, Y. Crosstalk of oxidative damage, apoptosis, and autophagy under endoplasmic reticulum (ER) stress involved in thifluzamide-induced liver damage in zebrafish (Danio rerio). Environ. Pollut 2018, 243, 1904–1911. [Google Scholar] [CrossRef]
- Zhang, Y.; Cen, J.; Jia, Z.; Wang, R.; Jiang, Z.; Hsiao, C.-D.; Xia, Q.; Zhang, L.; Liu, K. Hepatotoxicity Induced by Isoniazid-Lipopolysaccharide through Endoplasmic Reticulum Stress, Autophagy, and Apoptosis Pathways in Zebrafish. Antimicrob. Agents Chemother. 2019, 63, e01639-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Cai, Y.; Zhang, S.R.; Li, C.Y.; Jiang, L.L.; Wei, P.; He, M.F. Mechanism of hepatotoxicity of first-line tyrosine kinase inhibitors: Gefitinib and afatinib. Toxicol. Lett. 2021, 343, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Gu, Z.; Jia, R.; He, Q.; Cao, L.; Du, J.; Feng, W.; Jeney, G.; Xu, P.; Yin, G. Alteration of lipid metabolism, autophagy, apoptosis and immune response in the liver of common carp (Cyprinus carpio) after long-term exposure to bisphenol A. Ecotoxicol. Environ. Saf. 2021, 211, 111923. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Li, S.; Li, J.; Han, L.; He, Q.; Wang, R.; Wang, X.; Liu, K. Developmental toxicity induced by PM2.5 through endoplasmic reticulum stress and autophagy pathway in zebrafish embryos. Chemosphere 2018, 197, 611–621. [Google Scholar] [CrossRef]
- Meng, X.; Dong, H.; Pan, Y.; Ma, L.; Liu, C.; Man, S.; Gao, W. Diosgenyl Saponin Inducing Endoplasmic Reticulum Stress and Mitochondria-Mediated Apoptotic Pathways in Liver Cancer Cells. J. Agric. Food Chem. 2019, 67, 11428–11435. [Google Scholar] [CrossRef] [PubMed]
- Qin, J.; Ru, S.; Wang, W.; Hao, L.; Ru, Y.; Wang, J.; Zhang, X. Long-term bisphenol S exposure aggravates non-alcoholic fatty liver by regulating lipid metabolism and inducing endoplasmic reticulum stress response with activation of unfolded protein response in male zebrafish. Environ. Pollut 2020, 263, 114535. [Google Scholar] [CrossRef] [PubMed]
- Ma, M.; Song, L.; Yan, H.; Liu, M.; Zhang, L.; Ma, Y.; Yuan, J.; Hu, J.; Ji, Z.; Zhang, R.; et al. Low dose tunicamycin enhances atherosclerotic plaque stability by inducing autophagy. Biochem. Pharm. 2016, 100, 51–60. [Google Scholar] [CrossRef]
- Bian, M.; Chen, X.; Zhang, C.; Jin, H.; Wang, F.; Shao, J.; Chen, A.; Zhang, F.; Zheng, S. Magnesium isoglycyrrhizinate promotes the activated hepatic stellate cells apoptosis via endoplasmic reticulum stress and ameliorates fibrogenesis in vitro and in vivo. Biofactors 2017, 43, 836–846. [Google Scholar] [CrossRef]
- Li, Z.; Zhang, L.; Gao, M.; Han, M.; Liu, K.; Zhang, Z.; Gong, Z.; Xing, L.; Shi, X.; Lu, K.; et al. Endoplasmic reticulum stress triggers Xanthoangelol-induced protective autophagy via activation of JNK/c-Jun Axis in hepatocellular carcinoma. J. Exp. Clin. Cancer Res. 2019, 38, 8. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Duan, L.X.; Xu, Z.S.; Wang, J.G.; Xi, S.M. The protective effect of the earthworm active ingredients on hepatocellular injury induced by endoplasmic reticulum stress. Biomed. Pharm. 2016, 82, 304–311. [Google Scholar] [CrossRef] [PubMed]
- Ahmadi, A.; Hayes, A.W.; Karimi, G. Resveratrol and endoplasmic reticulum stress: A review of the potential protective mechanisms of the polyphenol. Phytother Res. 2021. [Google Scholar] [CrossRef] [PubMed]
- Zhan, F.; Zhao, G.; Li, X.; Yang, S.; Yang, W.; Zhou, S.; Zhang, F. Inositol-requiring enzyme 1 alpha endoribonuclease specific inhibitor STF-083010 protects the liver from thioacetamide-induced oxidative stress, inflammation and injury by triggering hepatocyte autophagy. Int. Immunopharmacol. 2019, 73, 261–269. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Xu, W.; Wu, L.; Dong, B.; Jin, J.; Han, D.; Zhu, X.; Yang, Y.; Liu, H.; Xie, S. Differential regulation of endoplasmic reticulum stress-induced autophagy and apoptosis in two strains of gibel carp (Carassius gibelio) exposed to acute waterborne cadmium. Aquat. Toxicol. 2021, 231, 105721. [Google Scholar] [CrossRef]
- Hong, Z.; Minghua, W.; Bo, N.; Chaoyue, Y.; Haiyang, Y.; Haiqing, Y.; Chunyu, X.; Yan, Z.; Yuan, Y. Rosmarinic acid attenuates acrylamide induced apoptosis of BRL-3A cells by inhibiting oxidative stress and endoplasmic reticulum stress. Food Chem. Toxicol. 2021, 151, 112156. [Google Scholar] [CrossRef]
- Kim, E.H.; Park, P.H. Globular adiponectin protects rat hepatocytes against acetaminophen-induced cell death via modulation of the inflammasome activation and ER stress: Critical role of autophagy induction. Biochem. Pharm. 2018, 154, 278–292. [Google Scholar] [CrossRef]
- Korovila, I.; Hohn, A.; Jung, T.; Grune, T.; Ott, C. Reduced Liver Autophagy in High-Fat Diet Induced Liver Steatosis in New Zealand Obese Mice. Antioxidants 2021, 10, 501. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Li, X.; Liu, D.; Zhang, S.; Tan, N.; Yokota, H.; Zhang, P. Phosphorylation of eIF2α signaling pathway attenuates obesity-induced non-alcoholic fatty liver disease in an ER stress and autophagy-dependent manner. Cell Death Dis. 2020, 11, 1069. [Google Scholar] [CrossRef]
- Yu, Q.; Zou, L.; Yuan, X.; Fang, F.; Xu, F. Dexmedetomidine Protects Against Septic Liver Injury by Enhancing Autophagy Through Activation of the AMPK/SIRT1 Signaling Pathway. Front. Pharm. 2021, 12, 658677. [Google Scholar] [CrossRef]
- Zou, H.; Wang, T.; Yuan, J.; Sun, J.; Yuan, Y.; Gu, J.; Liu, X.; Bian, J.; Liu, Z. Cadmium-induced cytotoxicity in mouse liver cells is associated with the disruption of autophagic flux via inhibiting the fusion of autophagosomes and lysosomes. Toxicol. Lett. 2020, 321, 32–43. [Google Scholar] [CrossRef]
- Li, H.; Wang, B.; Zhang, H.; Katsube, T.; Xie, Y.; Gan, L. Apoptosis Induction by Iron Radiation via Inhibition of Autophagy in Trp53(+/−) Mouse Testes: Is Chronic Restraint-Induced Stress a Modifying Factor? Int. J. Biol. Sci. 2018, 14, 1109–1121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, H.I.; Kim, S.J.; Choi, J.W.; Lee, S.M. Genipin alleviates sepsis-induced liver injury by restoring autophagy. Br. J. Pharm. 2016, 173, 980–991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, W.; Liao, J.; Yang, F.; Zhang, H.; Chang, X.; Yang, Y.; Bilal, R.M.; Wei, G.; Liang, W.; Guo, J.; et al. Chronic tribasic copper chloride exposure induces rat liver damage by disrupting the mitophagy and apoptosis pathways. Ecotoxicol. Environ. Saf. 2021, 212, 111968. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.H.; Chen, B.; Zhang, J.P. Intracellular Insulin and Impaired Autophagy in a Zebrafish model and a Cell Model of Type 2 diabetes. Int. J. Biol. Sci. 2017, 13, 985–995. [Google Scholar] [CrossRef] [Green Version]
- Baek, A.R.; Hong, J.; Song, K.S.; Jang, A.S.; Kim, D.J.; Chin, S.S.; Park, S.W. Spermidine attenuates bleomycin-induced lung fibrosis by inducing autophagy and inhibiting endoplasmic reticulum stress (ERS)-induced cell death in mice. Exp. Mol. Med. 2020, 52, 2034–2045. [Google Scholar] [CrossRef]
- Cheng, K.C.; Wang, C.J.; Chang, Y.C.; Hung, T.W.; Lai, C.J.; Kuo, C.W.; Huang, H.P. Mulberry fruits extracts induce apoptosis and autophagy of liver cancer cell and prevent hepatocarcinogenesis in vivo. J. Food Drug Anal. 2020, 28, 84–93. [Google Scholar] [CrossRef] [Green Version]
- Liang, Y.; Zhang, Z.; Tu, J.; Wang, Z.; Gao, X.; Deng, K.; El-Samahy, M.A.; You, P.; Fan, Y.; Wang, F. γ-Linolenic Acid Prevents Lipid Metabolism Disorder in Palmitic Acid-Treated Alpha Mouse Liver-12 Cells by Balancing Autophagy and Apoptosis via the LKB1-AMPK-mTOR Pathway. J. Agric Food Chem. 2021, 69, 8257–8267. [Google Scholar] [CrossRef]
- Wu, N.; Li, J.; Luo, H.; Wang, D.; Bai, X. Hydroxysafflor yellow A promotes apoptosis via blocking autophagic flux in liver cancer. Biomed. Pharm. 2021, 136, 111227. [Google Scholar] [CrossRef]
- Dong, G.; Zhang, Z.; Duan, K.; Shi, W.; Huang, R.; Wang, B.; Luo, L.; Zhang, Y.; Ruan, H.; Huang, H. Beclin 1 deficiency causes hepatic cell apoptosis via endoplasmic reticulum stress in zebrafish larvae. Febs. Lett. 2020, 594, 1155–1165. [Google Scholar] [CrossRef]
- Zhan, C.; Liu, W.; Zhang, F.; Zhang, X. Microcystin-LR triggers different endoplasmic reticulum stress pathways in the liver, ovary, and offspring of zebrafish (Danio rerio). J. Hazard. Mater. 2020, 386, 121939. [Google Scholar] [CrossRef]
- Li, X.; Lv, Z.; Chen, J.; Nepovimova, E.; Long, M.; Wu, W.; Kuca, K. Bacillus amyloliquefaciens B10 can alleviate liver apoptosis and oxidative stress induced by aflatoxin B1. Food Chem. Toxicol. 2021, 151, 112124. [Google Scholar] [CrossRef] [PubMed]
- Lu, W.; Mei, J.; Yang, J.; Wu, Z.; Liu, J.; Miao, P.; Chen, Y.; Wen, Z.; Zhao, Z.; Kong, H.; et al. ApoE deficiency promotes non-alcoholic fatty liver disease in mice via impeding AMPK/mTOR mediated autophagy. Life Sci. 2020, 252, 117601. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, D.; Li, Z.; Zhang, T.; Peng, B.; Zhang, Y.; Sun, H.; Wang, S. 2-Amino-3-Methylimidazo[4,5-f]quinoline Triggering Liver Damage by Inhibiting Autophagy and Inducing Endoplasmic Reticulum Stress in Zebrafish (Danio rerio). Toxins 2021, 13, 826. https://doi.org/10.3390/toxins13110826
Li D, Li Z, Zhang T, Peng B, Zhang Y, Sun H, Wang S. 2-Amino-3-Methylimidazo[4,5-f]quinoline Triggering Liver Damage by Inhibiting Autophagy and Inducing Endoplasmic Reticulum Stress in Zebrafish (Danio rerio). Toxins. 2021; 13(11):826. https://doi.org/10.3390/toxins13110826
Chicago/Turabian StyleLi, Dan, Zhi Li, Tianchang Zhang, Bo Peng, Yan Zhang, Hongwen Sun, and Shuo Wang. 2021. "2-Amino-3-Methylimidazo[4,5-f]quinoline Triggering Liver Damage by Inhibiting Autophagy and Inducing Endoplasmic Reticulum Stress in Zebrafish (Danio rerio)" Toxins 13, no. 11: 826. https://doi.org/10.3390/toxins13110826