Identification of Shiga-Toxin-Producing Shigella Infections in Travel and Non-Travel Related Cases in Alberta, Canada

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
2.1. stx1-Producing S. flexneri Identified in Clinical Isolates
2.2. Sporadic Cases of stx1 Positive S. flexneri in Alberta, Canada
2.3. Location of stx1 Gene in S. flexneri Strains
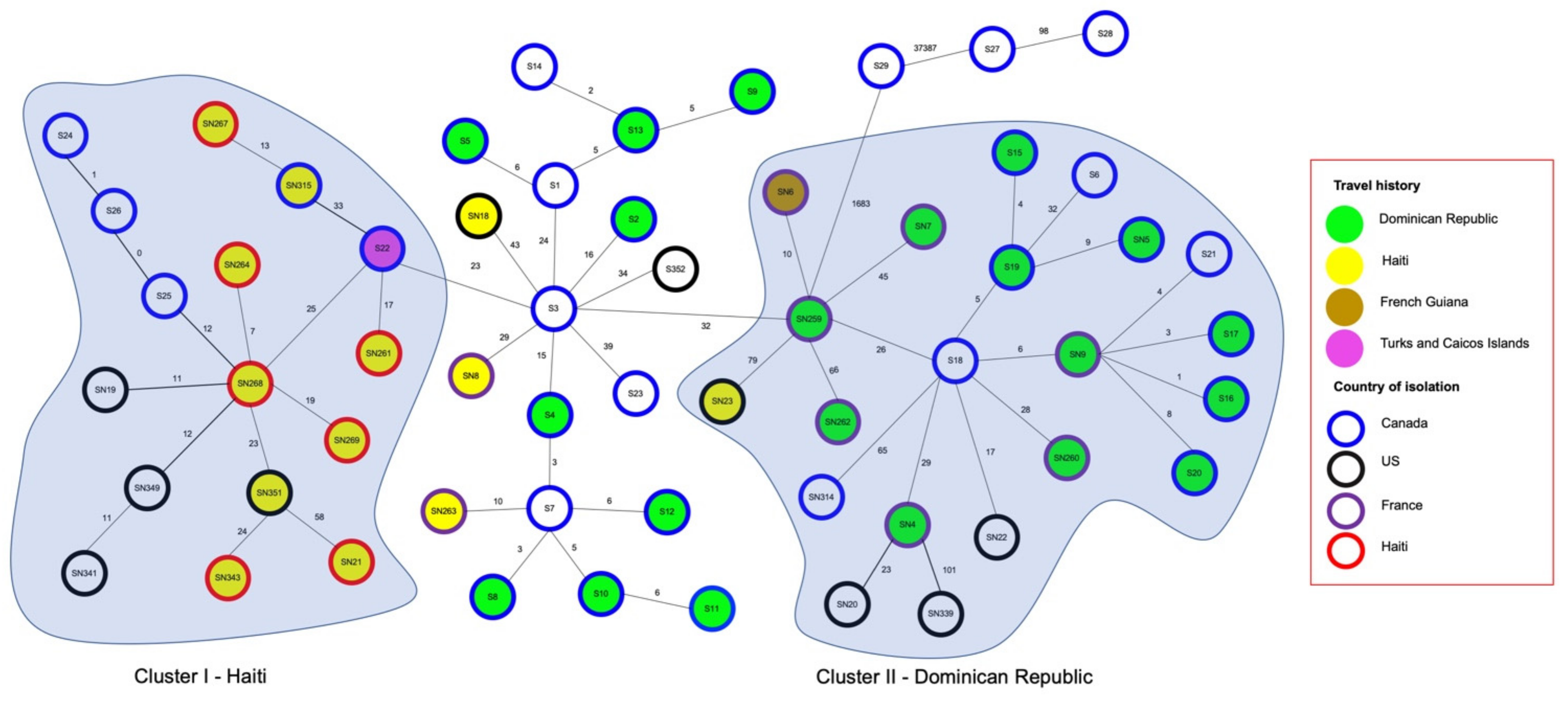
2.4. Relatedness of Travel and Non-Travel stx1 S. flexneri Strains
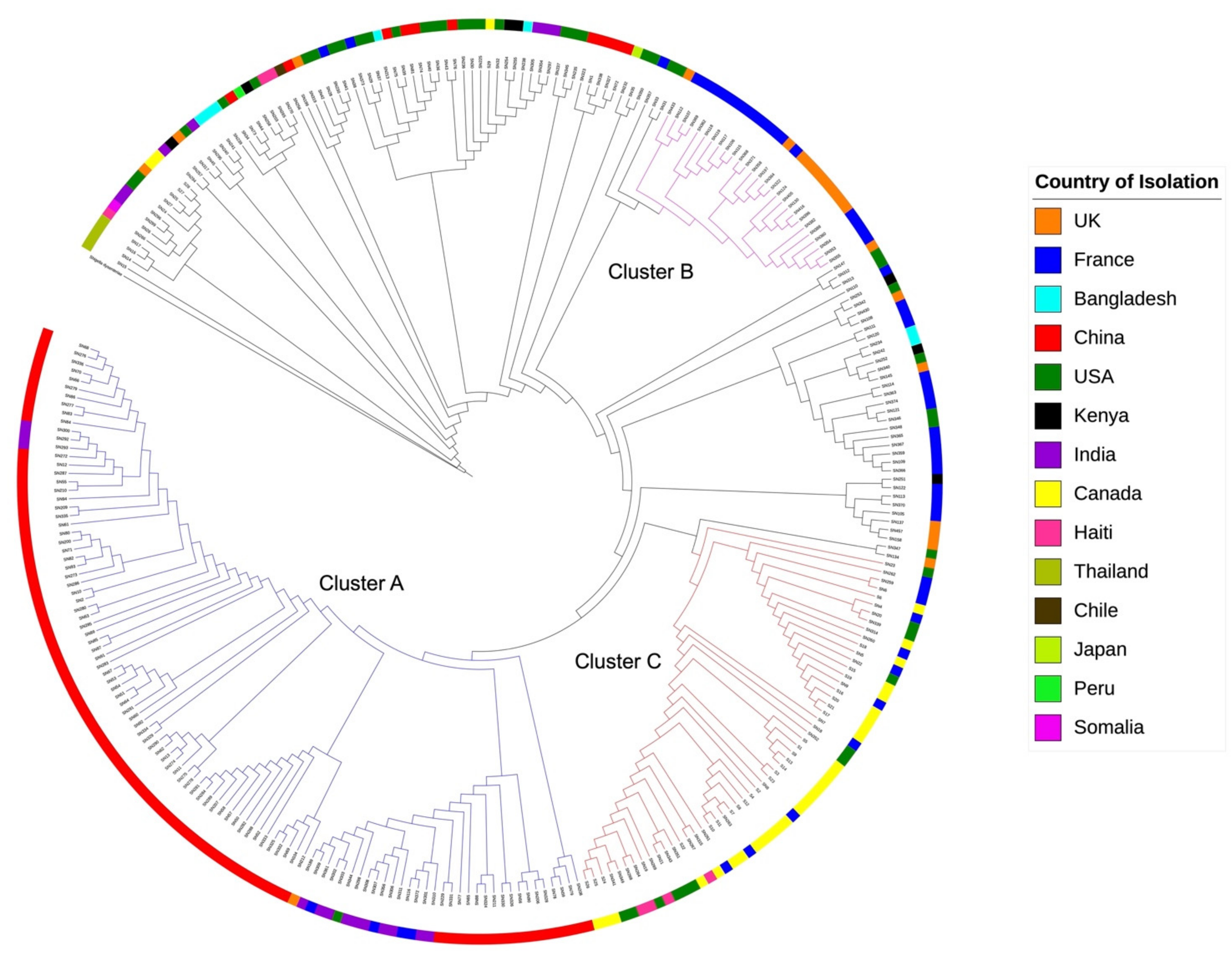
2.5. Relatedness of stx1 Positive S. flexneri Strains from Different Countries
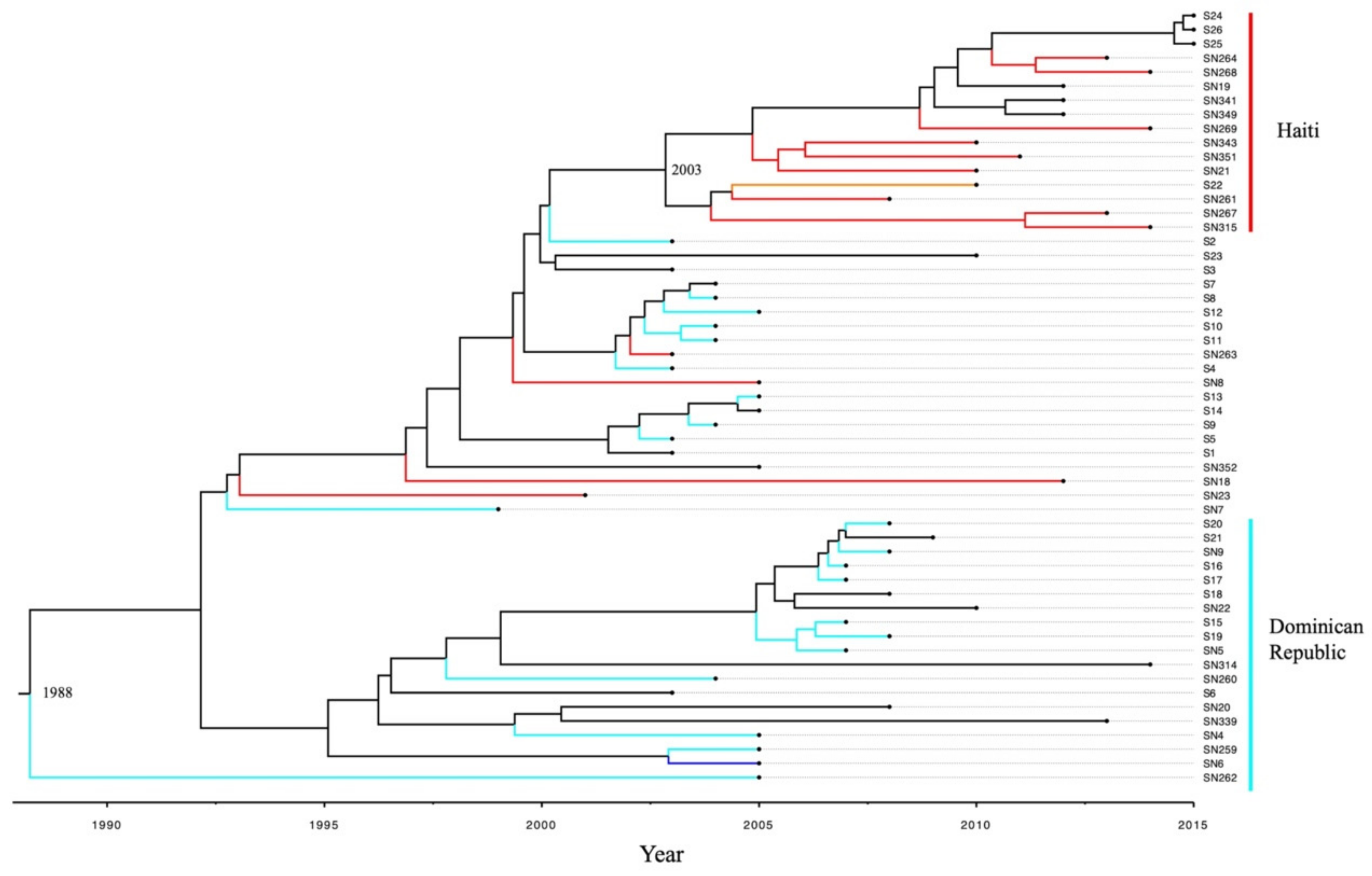
2.6. Bayesian Phylogenomic Analysis of stx1 Positive S. flexneri from Different Countries
2.7. Genome Evolution of stx1 Positive S. flexneri Strains
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Bacterial Strains
5.2. Identification of Shiga Toxin-Producing S. flexneri
5.3. Whole Genome Sequencing and Pairwise Whole Genome Similarity Analysis
5.4. Core Genome SNP Analysis and Minimum Spanning Tree
5.5. Bayesian Phylogenetic Analysis of stx Positive S. flexneri Strains from Different Countries
5.6. Phylogenomic Analysis of S. flexneri
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Zhu, Z.; Wang, W.; Cao, M.; Zhu, Q.; Ma, T.; Zhang, Y.; Liu, G.; Zhou, X.; Li, B.; Shi, Y.; et al. Virulence factors and molecular characteristics of Shigella flexneri isolated from calves with diarrhea. BMC Microbiol. 2021, 21, 214. [Google Scholar] [CrossRef]
- Shi, R.; Yang, X.; Chen, L.; Chang, H.T.; Liu, H.Y.; Zhao, J.; Wang, X.W.; Wang, C.Q. Pathogenicity of Shigella in chickens. PLoS ONE 2014, 9, e100264. [Google Scholar] [CrossRef] [Green Version]
- Cao, M.; Wang, W.; Zhang, L.; Liu, G.; Zhou, X.; Li, B.; Shi, Y.; Zhu, Z.; Zhang, J. Epidemic and molecular characterization of fluoroquinolone-resistant Shigella dysenteriae1 isolates from calves with diarrhea. BMC Microbiol. 2021, 21, 6. [Google Scholar] [CrossRef] [PubMed]
- Hu, G.Z.; Chen, H.Y.; Si, H.B.; Deng, L.X.; Wei, Z.Y.; Yuan, L.; Kuang, X.H. Phenotypic and molecular characterization of TEM-116 extended-spectrum beta-lactamase produced by a Shigella flexneri clinical isolate from chickens. FEMS Microbiol. Lett. 2008, 279, 162–166. [Google Scholar] [CrossRef] [PubMed]
- Kotloff, K.L.; Riddle, M.S.; Platts-Mills, J.A.; Pavlinac, P.; Zaidi, A.K.M. Shigellosis. Lancet 2018, 391, 801–812. [Google Scholar] [CrossRef]
- Taneja, N.; Mewara, A. Shigellosis: Epidemiology in India. Indian J. Med. Res. 2016, 143, 565–576. [Google Scholar] [CrossRef]
- Bowen, A.; Hurd, J.; Hoover, C.; Khachadourian, Y.; Traphagen, E.; Harvey, E.; Libby, T.; Ehlers, S.; Ongpin, M.; Norton, J.C.; et al. Importation and Domestic Transmission of Shigella sonnei Resistant to Ciprofloxacin—United States, May 2014–February 2015. MMWR Morb. Mortal. Wkly. Rep. 2015, 64, 318–320. [Google Scholar]
- Huang, J.Y.; Henao, O.L.; Griffin, P.M.; Vugia, D.J.; Cronquist, A.B.; Hurd, S.; Tobin-D’Angelo, M.; Ryan, P.; Smith, K.; Lathrop, S.; et al. Infection with Pathogens Transmitted Commonly Through Food and the Effect of Increasing Use of Culture-Independent Diagnostic Tests on Surveillance—Foodborne Diseases Active Surveillance Network, 10 U.S. Sites, 2012–2015. MMWR Morb. Mortal. Wkly. Rep. 2016, 65, 368–371. [Google Scholar] [CrossRef]
- Mattock, E.; Blocker, A.J. How Do the Virulence Factors of Shigella Work Together to Cause Disease? Front. Cell Infect. Microbiol. 2017, 7, 64. [Google Scholar] [CrossRef]
- Bryan, A.; Youngster, I.; McAdam, A.J. Shiga Toxin Producing Escherichia coli. Clin. Lab. Med. 2015, 35, 247–272. [Google Scholar] [CrossRef] [PubMed]
- Melton-Celsa, A.R. Shiga Toxin (Stx) Classification, Structure, and Function. Microbiol. Spectr. 2014, 2, 2–4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scheutz, F.; Teel, L.D.; Beutin, L.; Pierard, D.; Buvens, G.; Karch, H.; Mellmann, A.; Caprioli, A.; Tozzoli, R.; Morabito, S.; et al. Multicenter Evaluation of a Sequence-based Protocol for Subtyping Shiga toxins and Standardizing Stx Nomenclature. J. Clin. Microbiol. 2012, 50, 2951–2963. [Google Scholar] [CrossRef] [Green Version]
- Beutin, L.; Strauch, E.; Fischer, I. Isolation of Shigella sonnei Lysogenic for a Bacteriophage Encoding Gene for Production of Shiga toxin. Lancet 1999, 353, 1498. [Google Scholar] [CrossRef]
- Lamba, K.; Nelson, J.A.; Kimura, A.C.; Poe, A.; Collins, J.; Kao, A.S.; Cruz, L.; Inami, G.; Vaishampayan, J.; Garza, A.; et al. Shiga Toxin 1-Producing Shigella sonnei Infections, California, United States, 2014–2015. Emerg. Infect. Dis. 2016, 22, 679–686. [Google Scholar] [CrossRef] [Green Version]
- Adams, C.; Vose, A.; Edmond, M.B.; Lyckholm, L. Shigella sonnei and Hemolytic Uremic Syndrome: A Case Report and Literature Review. IDCases 2017, 8, 6–8. [Google Scholar] [CrossRef]
- Carter, C.C.; Fierer, J.; Chiu, W.W.; Looney, D.J.; Strain, M.; Mehta, S.R. A Novel Shiga Toxin 1a-Converting Bacteriophage of Shigella sonnei With Close Relationship to Shiga Toxin 2-Converting Pages of Escherichia coli. Open Forum Infect. Dis. 2016, 3, ofw079. [Google Scholar] [CrossRef]
- Gray, M.D.; Lampel, K.A.; Strockbine, N.A.; Fernandez, R.E.; Melton-Celsa, A.R.; Maurelli, A.T. Clinical isolates of Shiga toxin 1a-producing Shigella flexneri with An Epidemiological Link to Recent Travel to Hispaniola. Emerg. Infect. Dis. 2014, 20, 1669–1677. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.K.; Strockbine, N.; Omondi, M.; Hise, K.; Fair, M.A.; Mintz, E. Emergence of Shiga toxin 1 genes within Shigella dysenteriae Type 4 Isolates from Travelers Returning from the Island of Hispanola. Am. J. Trop. Med. Hyg. 2007, 76, 1163–1165. [Google Scholar] [CrossRef]
- Bekal, S.; Pilon, P.A.; Cloutier, N.; Doualla-Bell, F.; Longtin, J. Identification of Shigella flexneri isolates carrying the Shiga toxin 1-producing gene in Quebec, Canada, linked to travel to Haiti. Can. J. Microbiol. 2015, 61, 995–996. [Google Scholar] [CrossRef] [Green Version]
- Nogrady, N.; Kiraly, M.; Borbas, K.; Toth, A.; Paszti, J.; Toth, I. Antimicrobial Resistance and Genetic Characteristics of Integron-Carrier Shigellae Isolated in Hungary (1998–2008). J. Med. Microbiol. 2013, 62, 1545–1551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gray, M.D.; Lacher, D.W.; Leonard, S.R.; Abbott, J.; Zhao, S.; Lampel, K.A.; Prothery, E.; Gouali, M.; Weill, F.X.; Maurelli, A.T. Prevalence of Shiga toxin-producing Shigella Species Isolated from French Travellers Returning from the Caribbean: An Emerging Pathogen with International Implications. Clin. Microbiol. Infect. 2015, 21, 765.e9–765.e14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nyholm, O.; Lienemann, T.; Halkilahti, J.; Mero, S.; Rimhanen-Finne, R.; Lehtinen, V.; Salmenlinna, S.; Siitonen, A. Characterization of Shigella sonnei Isolate Carrying Shiga Toxin 2-Producing Gene. Emerg. Infect. Dis. 2015, 21, 891–892. [Google Scholar] [CrossRef]
- Gray, M.D.; Leonard, S.R.; Lacher, D.W.; Lampel, K.A.; Alam, M.T.; Morris, J.G., Jr.; Ali, A.; LaBreck, P.T.; Maurelli, A.T. Stx-Producing Shigella Species From Patients in Haiti: An Emerging Pathogen With the Potential for Global Spread. Open Forum Infect. Dis. 2015, 2, ofv134. [Google Scholar] [CrossRef]
- Freedman, S.B.; Lee, B.E.; Louie, M.; Pang, X.L.; Ali, S.; Chuck, A.; Chui, L.; Currie, G.R.; Dickinson, J.; Drews, S.J.; et al. Alberta Provincial Pediatric EnTeric Infection TEam (APPETITE): Epidemiology, emerging organisms, and economics. BMC Pediatr. 2015, 15, 89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rasko, D.A.; Webster, D.R.; Sahl, J.W.; Bashir, A.; Boisen, N.; Scheutz, F.; Paxinos, E.E.; Sebra, R.; Chin, C.S.; Iliopoulos, D.; et al. Origins of the E. coli strain causing an outbreak of hemolytic-uremic syndrome in Germany. N. Engl. J. Med. 2011, 365, 709–717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kruger, A.; Lucchesi, P.M. Shiga Toxins and stx Phages: Highly Diverse Entities. Microbiology 2015, 161, 451–462. [Google Scholar] [CrossRef] [Green Version]
- Fogolari, M.; Mavian, C.; Angeletti, S.; Salemi, M.; Lampel, K.A.; Maurelli, A.T. Distribution and Characterization of Shiga Toxin Converting Temperate Phages Carried by Shigella flexneri in Hispaniola. Infect. Genet. Evol. 2018, 65, 321–328. [Google Scholar] [CrossRef]
- The World Factbook–Dominican Republic. Available online: https://www.cia.gov/library/publications/the-world-factbook/geos/dr.html (accessed on 1 December 2020).
- The World Factbook–Haiti. Available online: https://www.cia.gov/library/publications/the-world-factbook/geos/ha.html (accessed on 1 December 2020).
- Liu, W.; Zhang, J.; Wu, C.; Cai, S.; Huang, W.; Chen, J.; Xi, X.; Liang, Z.; Hou, Q.; Zhou, B.; et al. Unique Features of Ethnic Mongolian Gut Microbiome Revealed by Metagenomic Analysis. Sci. Rep. 2016, 6, 34826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Labrie, S.J.; Samson, J.E.; Moineau, S. Bacteriophage Resistance Mechanisms. Nat. Rev. Microbiol. 2010, 8, 317–327. [Google Scholar] [CrossRef]
- Davies, T.J.; Pedersen, A.B. Phylogeny and Geography Predict Pathogen Community Similarity in Wild Primates and Humans. Proc. Biol. Sci. 2008, 275, 1695–1701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franz, E.; Rotariu, O.; Lopes, B.S.; MacRae, M.; Bono, J.L.; Laing, C.; Gannon, V.; Soderlund, R.; van Hoek, A.; Friesema, I.; et al. Phylogeographic Analysis Reveals Multiple International Transmission Events Have Driven the Global Emergence of Escherichia coli O157:H7. Clin. Infect. Dis. 2018, 69, 428–437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lemey, P.; Rambaut, A.; Drummond, A.J.; Suchard, M.A. Bayesian Phylogeography Finds Its Roots. PLoS Comput. Biol. 2009, 5, e1000520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bugarel, M.; Beutin, L.; Martin, A.; Gill, A.; Fach, P. Micro-array for the Identification of Shiga toxin-producing Escherichia coli (STEC) Seropathotypes Associated with Hemorrhagic Colitis and Hemolytic Uremic Syndrome in Humans. Int. J. Food Microbiol. 2010, 142, 318–329. [Google Scholar] [CrossRef] [PubMed]
- Untergasser, A.; Cutcutache, I.; Koressaar, T.; Ye, J.; Faircloth, B.C.; Remm, M.; Rozen, S.G. Primer3—New Capabilities and Interfaces. Nucleic Acids Res. 2012, 40, e115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A Flexible Trimmer for Illumina Sequence Data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A New Genome Assembly Algorithm and Its Applications to Single-Cell Sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertels, F.; Silander, O.K.; Pachkov, M.; Rainey, P.B.; van Nimwegen, E. Automated Reconstruction of Whole-Genome Phylogenies from Short-Sequence Reads. Mol. Biol. Evol. 2014, 31, 1077–1088. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Nascimento, M.; Sousa, A.; Ramirez, M.; Francisco, A.P.; Carrico, J.A.; Vaz, C. PHYLOViZ 2.0: Providing Scalable Data Integration and Visualization for Multiple Phylogenetic Inference Methods. Bioinformatics 2017, 33, 128–129. [Google Scholar] [CrossRef]
- Croucher, N.J.; Page, A.J.; Connor, T.R.; Delaney, A.J.; Keane, J.A.; Bentley, S.D.; Parkhill, J.; Harris, S.R. Rapid Phylogenetic Analysis of Large Samples of Recombinant Bacterial Whole Genome Sequences Using Gubbins. Nucleic Acids Res. 2015, 43, e15. [Google Scholar] [CrossRef] [Green Version]
- Bouckaert, R.; Vaughan, T.G.; Barido-Sottani, J.; Duchene, S.; Fourment, M.; Gavryushkina, A.; Heled, J.; Jones, G.; Kuhnert, D.; De Maio, N.; et al. BEAST 2.5: An Advanced Software Platform for Bayesian Evolutionary Analysis. PLoS Comput. Biol. 2019, 15, e1006650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL): An Online Tool for Phylogenetic Tree Display and Annotation. Bioinformatics 2007, 23, 127–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Letunic, I.; Bork, P. Interactive Tree Of Life v2: Online Annotation and Display of Phylogenetic Trees Made Easy. Nucleic Acids Res. 2011, 39, W475–W478. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain Name in This Study | Strains Names in NCBI Database | Country Where Shigellosis Was Diagnosed | Travel History | Year of Diagnosis | Strain Name in This Study | Strains Names in NCBI Database | Country Where Shigellosis Was Diagnosed | Travel History | Year of Diagnosis |
|---|---|---|---|---|---|---|---|---|---|
| S1 | S1 | Canada | NO a | 2003 | SN20 | BS982 | USA | NA b | 2008 |
| S2 | S2 | Canada | Dominican Republic | 2003 | SN21 | BS937 | USA | Haiti | 2010 |
| S3 | S3 | Canada | NO a | 2003 | SN22 | BS942 | USA | NA b | 2010 |
| S4 | S4 | Canada | Dominican Republic | 2003 | SN23 | BS974 | USA | Haiti | 2001 |
| S5 | S5 | Canada | Dominican Republic | 2003 | SN259 | BS1023 | France | Dominican Republic | 2005 |
| S6 | S6 | Canada | NO a | 2003 | SN260 | BS1022 | France | Dominican Republic | 2004 |
| S7 | S7 | Canada | NO a | 2004 | SN261 | BS1025 | France | Haiti | 2008 |
| S8 | S8 | Canada | Dominican Republic | 2004 | SN262 | BS1044 | France | Dominican Republic | 2005 |
| S9 | S9 | Canada | Dominican Republic | 2004 | SN263 | BS1021 | France | Haiti | 2003 |
| S10 | S10 | Canada | Dominican Republic | 2004 | SN264 | BS1057 | Haiti | Haiti | 2013 |
| S11 | S11 | Canada | Dominican Republic | 2004 | SN267 | BS1039 | Haiti | Haiti | 2013 |
| S12 | S12 | Canada | Dominican Republic | 2005 | SN268 | BS1059 | Haiti | Haiti | 2014 |
| S13 | S13 | Canada | Dominican Republic | 2005 | SN269 | BS1060 | Haiti | Haiti | 2014 |
| S14 | S14 | Canada | NO a | 2005 | SN314 | SH200 | Canada | NA b | 2014 |
| S15 | S15 | Canada | Dominican Republic | 2007 | SN315 | SH199 | Canada | Haiti | 2014 |
| S16 | S16 | Canada | Dominican Republic | 2007 | SN339 | BS989 | USA | NA b | 2013 |
| S17 | S17 | Canada | Dominican Republic | 2007 | SN341 | BS972 | USA | NA b | 2012 |
| S18 | S18 | Canada | NO a | 2008 | SN343 | BS968 | USA | Haiti | 2010 |
| S19 | S19 | Canada | Dominican Republic | 2008 | SN349 | BS973 | USA | NA b | 2012 |
| S20 | S20 | Canada | Dominican Republic | 2008 | SN351 | BS971 | USA | Haiti | 2011 |
| S21 | S21 | Canada | NO a | 2009 | SN352 | BS951 | USA | NA b | 2005 |
| S22 | S22 | Canada | Turks and Caicos Islands | 2010 | SN4 | BS1042 | France | Dominican Republic | 2005 |
| S23 | S23 | Canada | NO a | 2010 | SN5 | BS1045 | France | Dominican Republic | 2007 |
| S24 | S24 | Canada | NO a | 2015 | SN6 | BS1024 | France | French Guiana | 2005 |
| S25 | S25 | Canada | NO a | 2015 | SN7 | BS1041 | France | Dominican Republic | 1999 |
| S26 | S26 | Canada | NO a | 2015 | SN8 | BS1043 | France | Haiti | 2005 |
| SN9 | BS1046 | France | Dominican Republic | 2008 | |||||
| SN18 | BS988 | USA | Haiti | 2012 | |||||
| SN19 | BS938 | USA | NA b | 2012 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhi, S.; Parsons, B.D.; Szelewicki, J.; Yuen, Y.T.K.; Fach, P.; Delannoy, S.; Li, V.; Ferrato, C.; Freedman, S.B.; Lee, B.E.; et al. Identification of Shiga-Toxin-Producing Shigella Infections in Travel and Non-Travel Related Cases in Alberta, Canada. Toxins 2021, 13, 755. https://doi.org/10.3390/toxins13110755
Zhi S, Parsons BD, Szelewicki J, Yuen YTK, Fach P, Delannoy S, Li V, Ferrato C, Freedman SB, Lee BE, et al. Identification of Shiga-Toxin-Producing Shigella Infections in Travel and Non-Travel Related Cases in Alberta, Canada. Toxins. 2021; 13(11):755. https://doi.org/10.3390/toxins13110755
Chicago/Turabian StyleZhi, Shuai, Brendon D. Parsons, Jonas Szelewicki, Yue T. K. Yuen, Patrick Fach, Sabine Delannoy, Vincent Li, Christina Ferrato, Stephen B. Freedman, Bonita E. Lee, and et al. 2021. "Identification of Shiga-Toxin-Producing Shigella Infections in Travel and Non-Travel Related Cases in Alberta, Canada" Toxins 13, no. 11: 755. https://doi.org/10.3390/toxins13110755