β-Hemolysis May Not Be a Reliable Indicator of Leukotoxicity of Mannheimia haemolytica Isolates

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
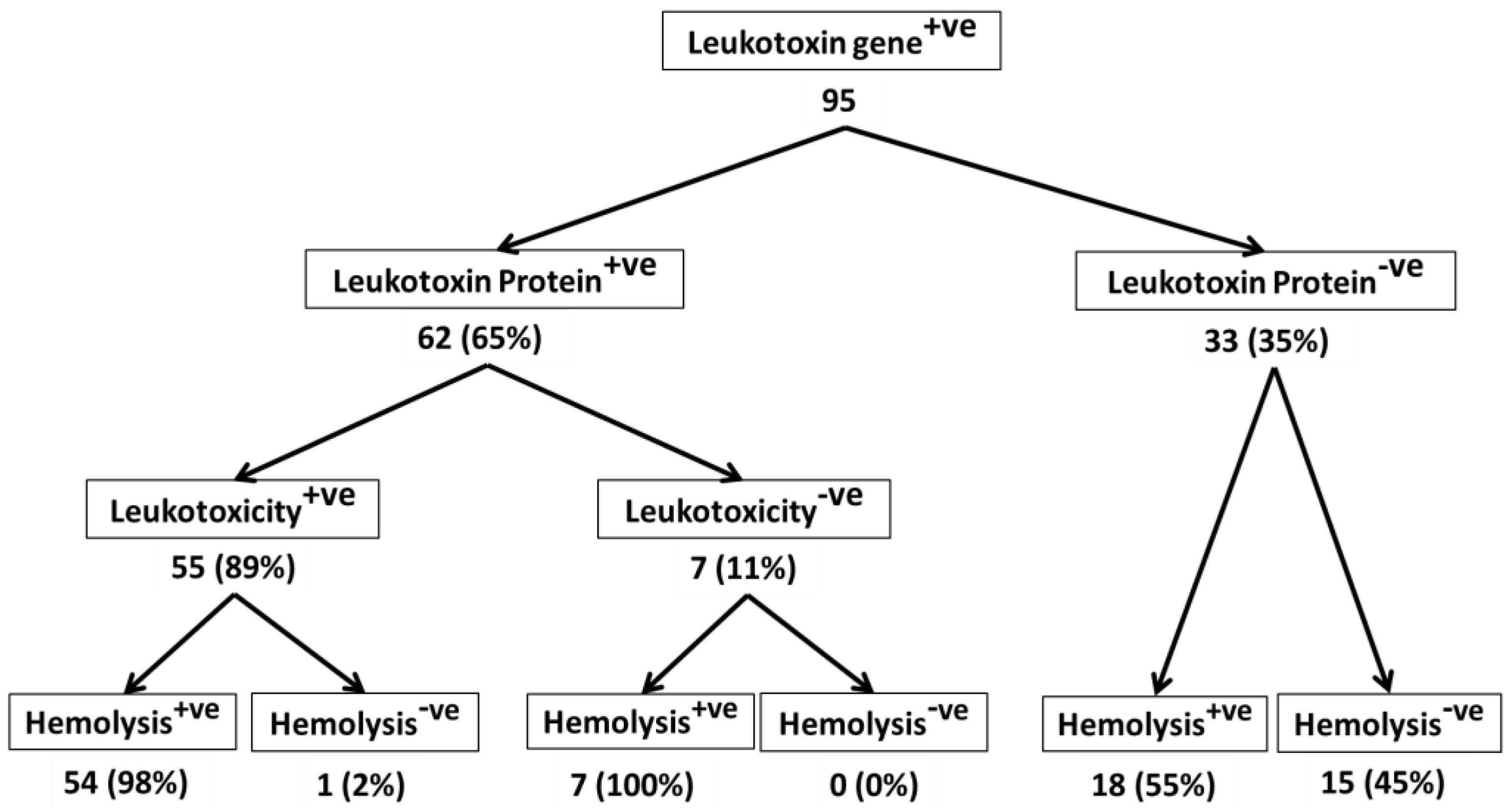
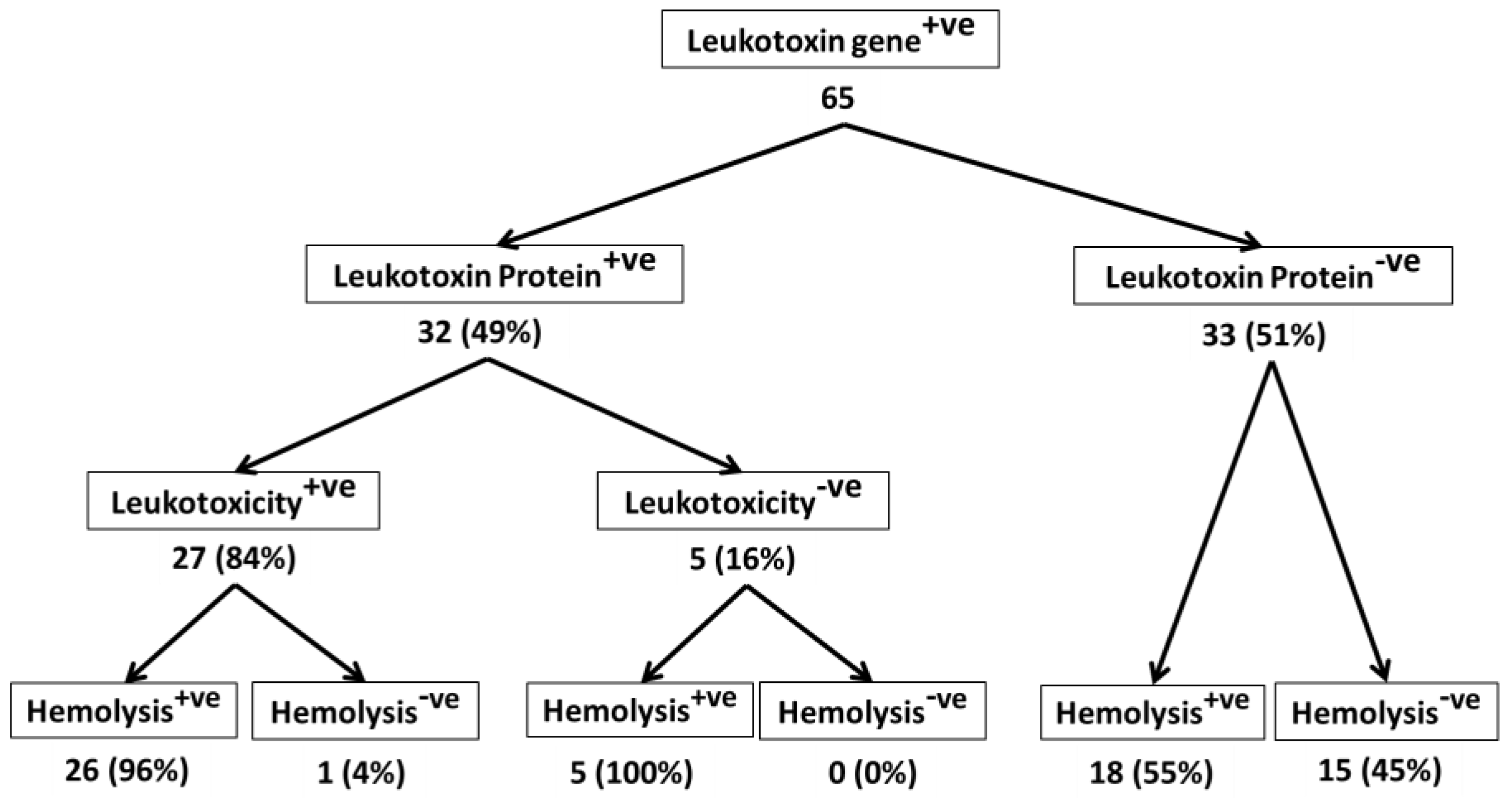
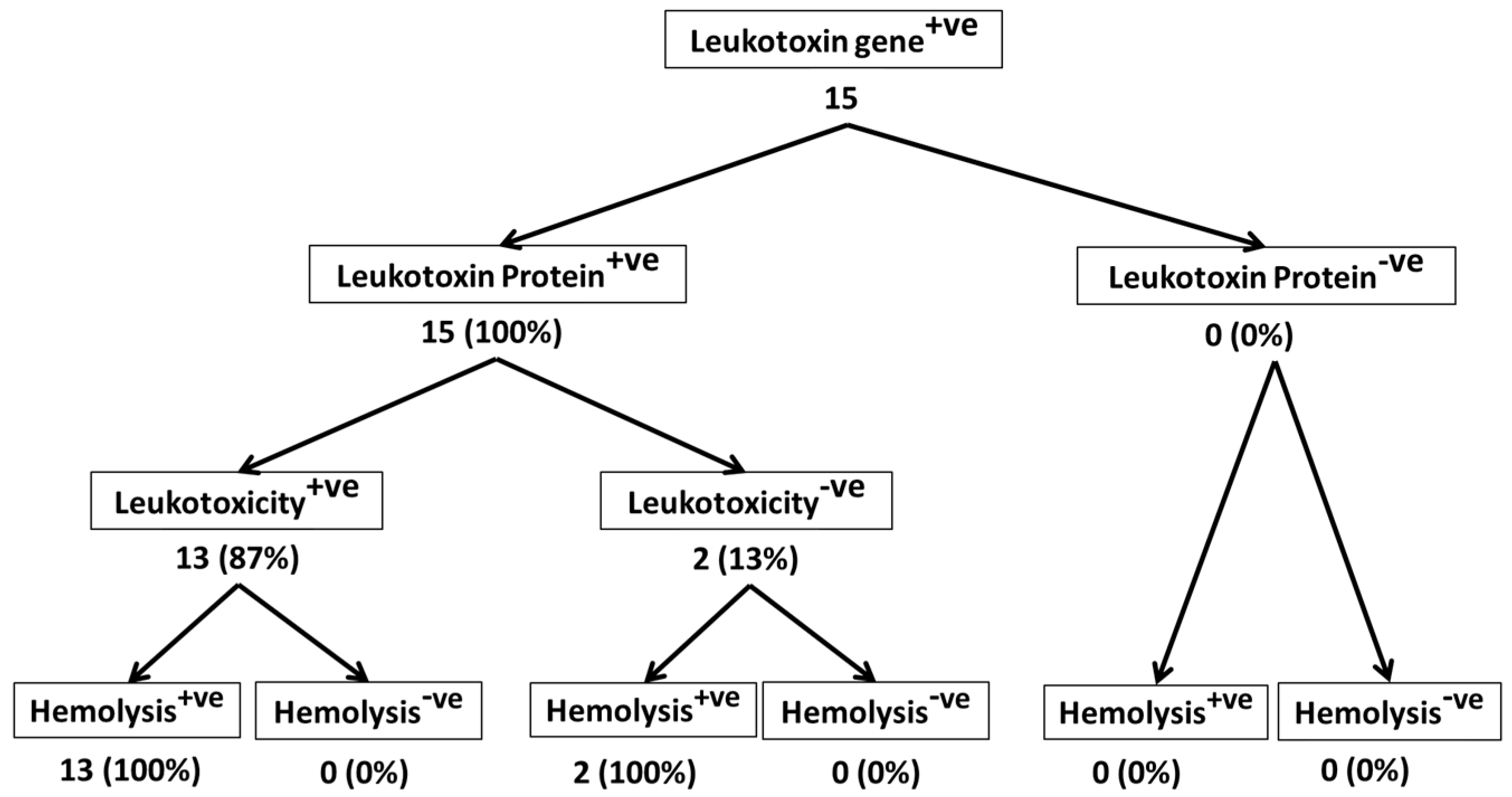
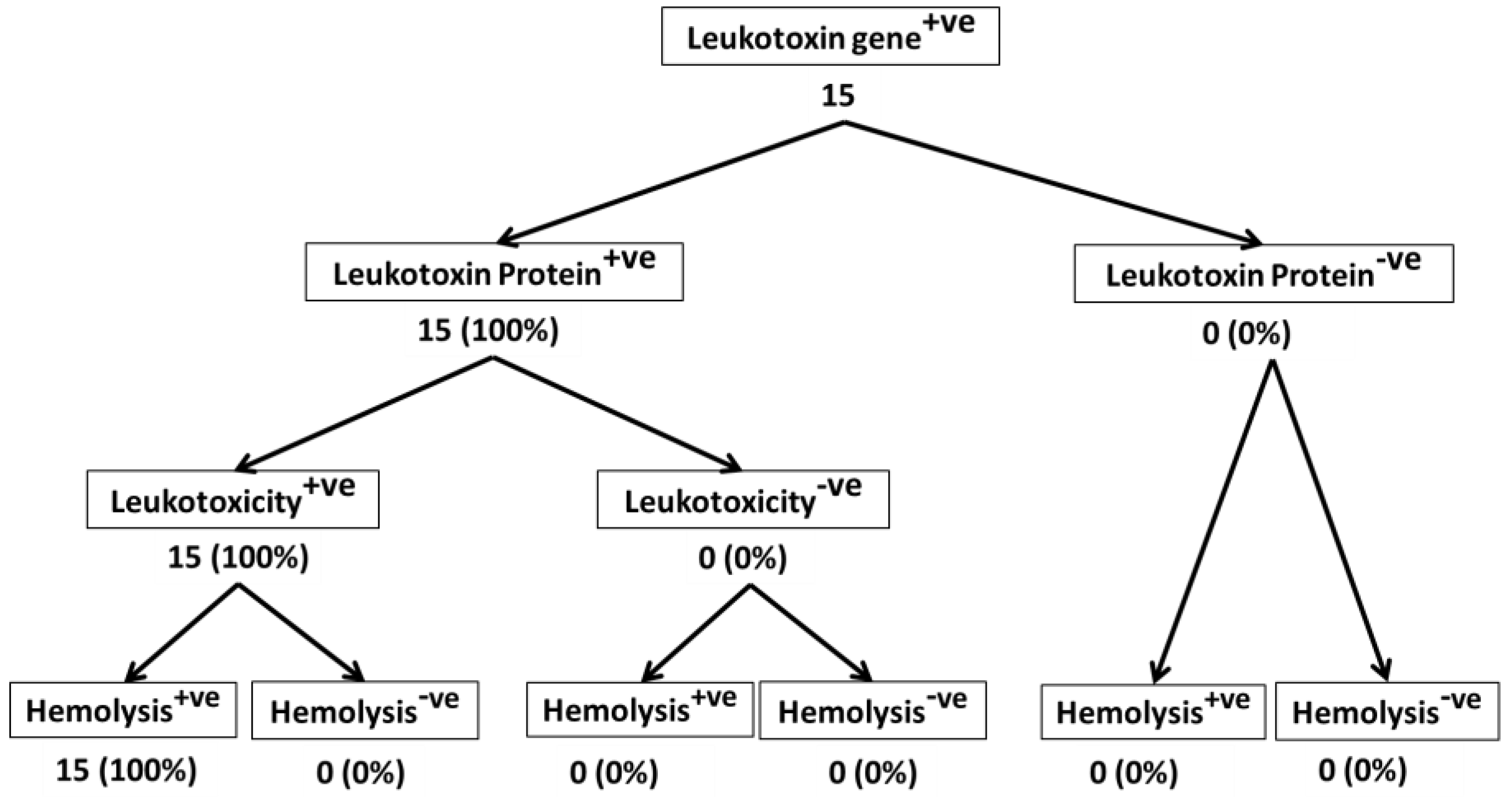
2. Results and Discussion
2.1. All M. haemolytica Isolates that Possess the lktA Gene May Not Produce Lkt Protein
2.2. Lkt-Negative M. haemolytica Isolates Can Induce Hemolysis
2.3. Not All Lkt-Positive M. haemolytica Isolates May Cause Leukotoxicity
2.4. Majority of Leukotoxic M. haemolytica Isolates Are Hemolytic
3. Conclusions
4. Materials and Methods
4.1. Experimental Design
4.2. M. haemolytica and Lkt-Specific PCR Assays
4.3. Leukotoxin Production
4.4. ELISA for Detection of Lkt Protein
4.5. Assay for Detection of Leukotoxic Activity
4.6. Assay for Detection of β-Hemolysis
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Ackermann, M.R.; Brogden, K.A. Response of the ruminant respiratory tract to Mannheimia (Pasteurella) haemolytica. Microb. Infect. 2000, 2, 1079–1088. [Google Scholar] [CrossRef]
- Miller, M.W. Pasteurellosis. In Infectious Diseases of Wild Mammals; Iowa State University Press: Iowa City, IA, USA, 2008; pp. 330–339. [Google Scholar]
- Mosier, D.A. Bacterial pneumonia. Vet. Clin. N. Am. Food Anim. Pract. 1997, 13, 483–493. [Google Scholar] [CrossRef]
- Confer, A.W.; Panciera, R.J.; Clinkenbeard, K.D.; Mosier, D.A. Molecular aspects of virulence of Pasteurella haemolytica. Can. J. Vet. Res. 1990, 54, S48–S52. [Google Scholar] [PubMed]
- Strathdee, C.A.; Lo, R.Y. Cloning, nucleotide sequence, and characterization of genes encoding the secretion function of the Pasteurella haemolytica leukotoxin determinant. J. Bacteriol. 1989, 171, 916–928. [Google Scholar] [CrossRef] [PubMed]
- Devenish, J.; Rosendal, S.; Johnson, R.; Hubler, S. Immunoserological comparison of 104-kilodalton proteins associated with hemolysis and cytolysis in Actinobacillus pleuropneumoniae, Actinobacillus suis, Pasteurella haemolytica, and Escherichia coli. Infect. Immun. 1989, 57, 3210–3213. [Google Scholar] [PubMed]
- Kolodrubetz, D.; Dailey, T.; Ebersole, J.; Kraig, E. Cloning and expression of the leukotoxin gene from Actinobacillus actinomycetemcomitans. Infect. Immun. 1989, 57, 1465–1469. [Google Scholar] [PubMed]
- Kaehler, K.L.; Markham, R.J.; Muscoplat, C.C.; Johnson, D.W. Evidence of species specificity in the cytocidal effects of Pasteurella haemolytica. Infect. Immun. 1980, 30, 615–616. [Google Scholar] [PubMed]
- Chang, Y.F.; Renshaw, H.W.; Martens, R.J.; Livingston, C.W., Jr. Pasteurella haemolytica leukotoxin: Chemiluminescent responses of peripheral blood leukocytes from several different mammalian species to leukotoxin- and opsonin-treated living and killed Pasteurella haemolytica and Staphylococcus aureus. Am. J. Vet. Res. 1986, 47, 67–74. [Google Scholar] [PubMed]
- Shewen, P.E.; Wilkie, B.N. Cytotoxin of Pasteurella haemolytica acting on bovine leukocytes. Infect. Immun. 1982, 35, 91–94. [Google Scholar] [PubMed]
- Dassanayake, R.P.; Shanthalingam, S.; Herndon, C.N.; Lawrence, P.K.; Frances Cassirer, F.E.; Potter, K.A.; Foreyt, W.J.; Clinkenbeard, K.D.; Srikumaran, S. Mannheimia haemolytica serotype A1 exhibits differential pathogenicity in two related species, Ovis canadensis and Ovis aries. Vet. Microbiol. 2009, 133, 366–371. [Google Scholar] [CrossRef] [PubMed]
- Petras, S.F.; Chidambaram, M.; Illyes, E.F.; Froshauer, S.; Weinstock, G.M.; Reese, C.P. Antigenic and Virulence properties of Pasteurella haemolytica leukotoxin mutants. Infect. Immun. 1995, 63, 1033–1039. [Google Scholar] [PubMed]
- Highlander, S.K.; Fedorova, N.D.; Dusek, D.M.; Panciera, R.; Alvarez, L.E.; Rinehart, C. Inactivation of Pasteurella (Mannheimia) haemolytica leukotoxin causes partial attenuation of virulence in a calf challenge model. Infect. Immun. 2000, 68, 3916–3922. [Google Scholar] [CrossRef] [PubMed]
- Slocombe, R.J.; Marark, J.; Ingersoll, R.; Derksen, F.J.; Robinson, N.E. Importance of neutrophils in the pathogenesis of acute pneumonic pasteurellosis in calves. Am. J. Vet. Res. 1985, 46, 2253–2258. [Google Scholar] [PubMed]
- Highlander, S.K. Molecular genetic analysis of virulence in Mannheimia (Pasteurella) haemolytica. Front. Biosci. 2001, 1, D1128–D1150. [Google Scholar]
- Jayaseelan, S.; Sreevatsan, S.; Maheswaran, S.K. Role of Mannheimia haemolytica leukotoxin in the pathogenesis of bovine pneumonic pasteurellosis. Anim. Health Res. Rev. 2002, 3, 69–82. [Google Scholar] [CrossRef]
- Chidambaram, M.; Sharma, B.; Petras, S.F.; Reese, C.P.; Froshauer, S.; Weinstock, G.M. Isolation of Pasteurella haemolytica leukotoxin mutants. Infect. Immun. 1995, 63, 1027–1032. [Google Scholar] [PubMed]
- Tatum, F.M.; Briggs, R.E.; Sreevatsan, S.S.; Zehr, E.S.; Ling Hsuan, S.; Whiteley, L.O.; Ames, T.R.; Maheswaran, S.K. Construction of an isogenic leukotoxin deletion mutant of Pasteurella haemolytica serotype 1: Characterization and virulence. Microb. Pathog. 1998, 24, 37–46. [Google Scholar] [CrossRef] [PubMed]
- Murphy, G.L.; Whitworth, L.C.; Clinkenbeard, K.D.; Clinkenbeard, P.A. Hemolytic activity of the Pasteurella haemolytica leukotoxin. Infect. Immun. 1995, 63, 3209–3212. [Google Scholar] [PubMed]
- Highlander, S.K.; Chidambaram, M.; Engler, M.J.; Weinstock, G.M. DNA sequence of the Pasteurella haemolytica leukotoxin gene cluster. DNA 1989, 8, 15–28. [Google Scholar] [CrossRef] [PubMed]
- Highlander, S.K.; Engler, M.J.; Weinstock, G.M. Secretion and expression of the Pasteurella haemolytica Leukotoxin. J. Bacteriol. 1990, 172, 2343–2350. [Google Scholar] [CrossRef] [PubMed]
- Thumbikat, P.; Briggs, R.E.; Kannan, M.S.; Maheswaran, S.K. Biological effects of two genetically defined leukotoxin mutants of Mannheimia haemolytica. Microb. Pathog. 2003, 34, 217–226. [Google Scholar] [CrossRef]
- Shanthalingam, S.; Goldy, A.; Bavananthasivam, J.; Subramaniam, R.; Batra, S.A.; Kugadas, A.; Raghavan, B.; Dassanayake, R.P.; Jennings-Gaines, J.E.; Killion, H.J.; et al. PCR assay detects Mannheimia haemolytica in culture-negative pneumonic lung tissues of bighorn sheep (Ovis canadensis) from outbreaks in the western USA, 2009–2010. J. Wildl. Dis. 2014, 50, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Subramaniam, R.; Shanthalingam, S.; Bavananthasivam, J.; Kugadas, A.; Potter, K.A.; Foreyt, W.J.; Hodgins, D.C.; Shewen, P.; Barrington, G.M.; Knowles, D.P.; et al. A multivalent Mannheimia/Bibersteinia vaccine protects bighorn sheep against Mannheimia haemolytica challenge. Clin. Vaccine Immunol. 2011, 18, 1689–1694. [Google Scholar] [CrossRef] [PubMed]
- Gentry, M.J.; Srikumaran, S. Neutralizing monoclonal antibodies to Pasteurella haemolytica leukotoxin affinity-purify the toxin from crude culture supernatants. Microb. Pathog. 1991, 10, 411–417. [Google Scholar] [CrossRef]
- Smibert, R.M.; Krieg, N.R. Phenotypic Characterization. In Methods for General and Molecular Bacteriology; Gerhardt, P., Murray, R.G.E., Wood, W.A., Krieg, N.R., Eds.; American Society for Microbiology: Washington, DC, USA, 1994; pp. 607–654. [Google Scholar]
- Miller, M.W.; Hause, B.M.; Killion, H.J.; Fox, K.A.; Edwards, W.H.; Wolfe, L.L. Phylogenetic and epidemiologic relationships among Pasteurellaceae from Colorado bighorn sheep herds. J. Wildl. Dis. 2013, 49, 653–660. [Google Scholar] [CrossRef] [PubMed]
- Fedorova, N.D.; Highlander, S.K. Generation of targeted nonpolar gene insertions and operon fusions in Pasteurella haemolytica and creation of a strain that produces and secretes inactive leukotoxin. Infect. Immun. 1997, 65, 2593–2598. [Google Scholar] [PubMed]




© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bavananthasivam, J.; Shanthalingam, S.; Kugadas, A.; Raghavan, B.; Batra, S.; Srikumaran, S. β-Hemolysis May Not Be a Reliable Indicator of Leukotoxicity of Mannheimia haemolytica Isolates. Toxins 2018, 10, 173. https://doi.org/10.3390/toxins10050173
Bavananthasivam J, Shanthalingam S, Kugadas A, Raghavan B, Batra S, Srikumaran S. β-Hemolysis May Not Be a Reliable Indicator of Leukotoxicity of Mannheimia haemolytica Isolates. Toxins. 2018; 10(5):173. https://doi.org/10.3390/toxins10050173
Chicago/Turabian StyleBavananthasivam, Jegarubee, Sudarvili Shanthalingam, Abirami Kugadas, Bindu Raghavan, Sai Batra, and Subramaniam Srikumaran. 2018. "β-Hemolysis May Not Be a Reliable Indicator of Leukotoxicity of Mannheimia haemolytica Isolates" Toxins 10, no. 5: 173. https://doi.org/10.3390/toxins10050173