
Whole Cigarette Smoke Condensates Induce Accumulation of Amyloid Beta Precursor Protein with Oxidative Stress in Murine Astrocytes


Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
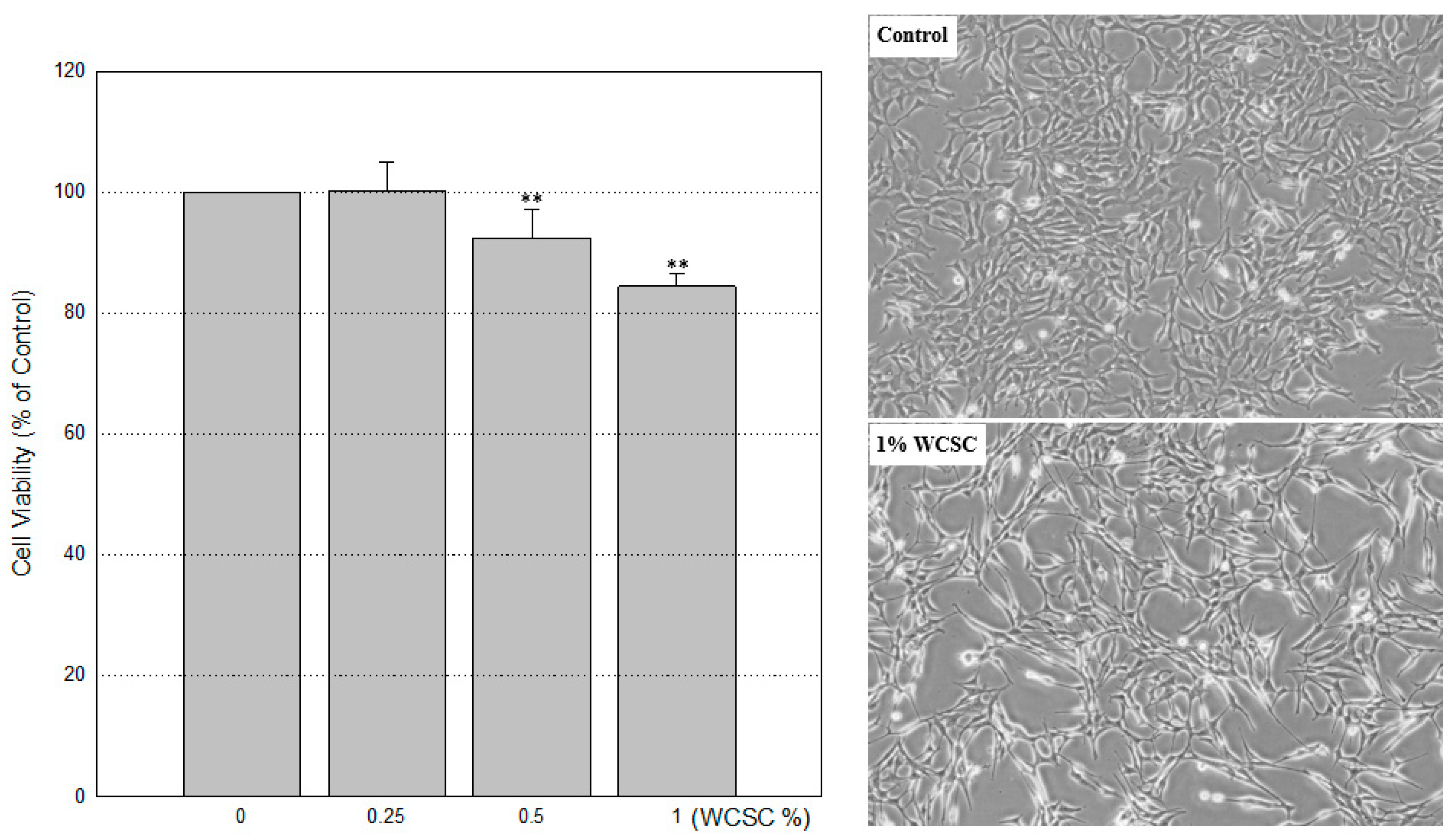
2.2. Cell Viability Assays
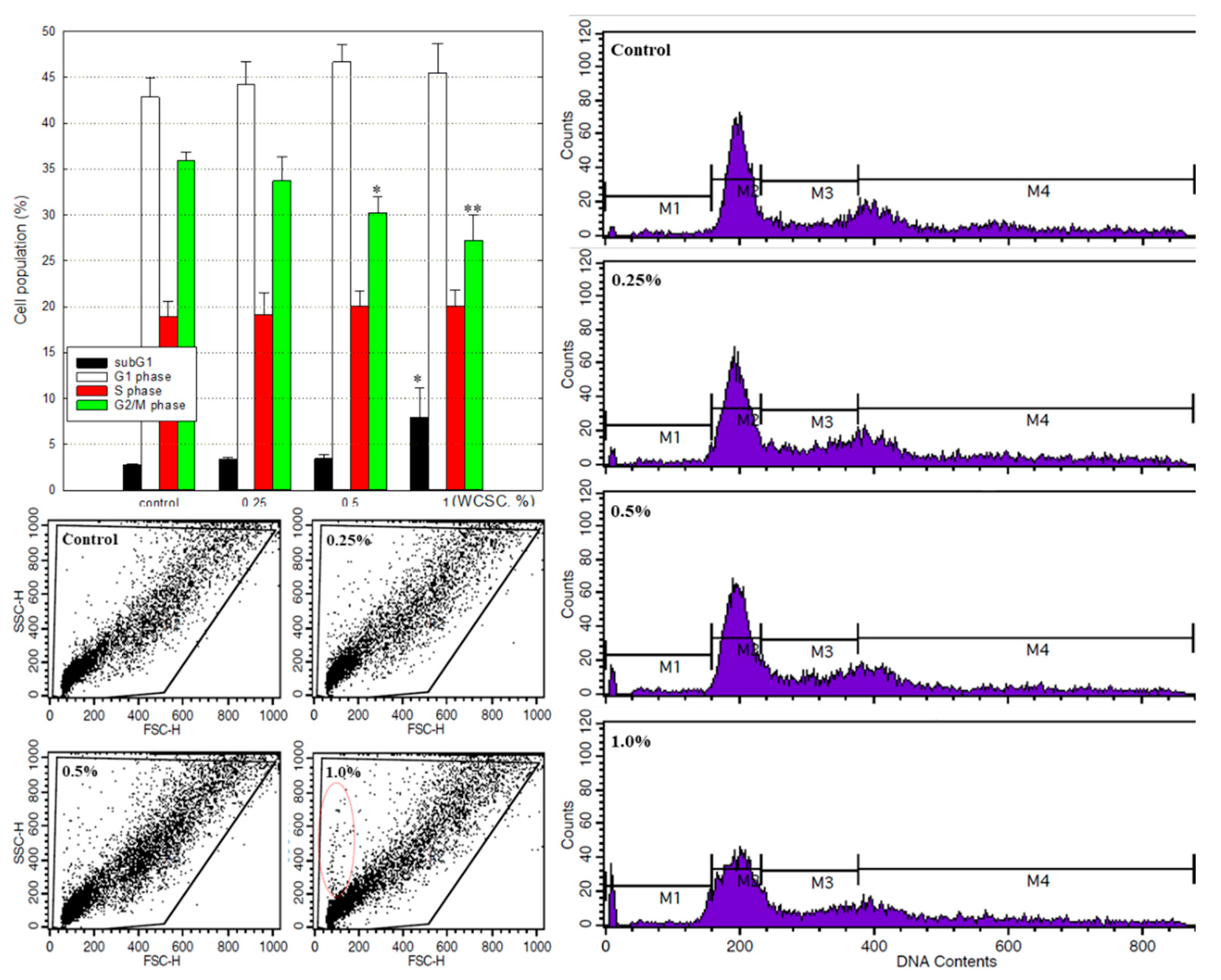
2.3. Cell Cycle Analysis
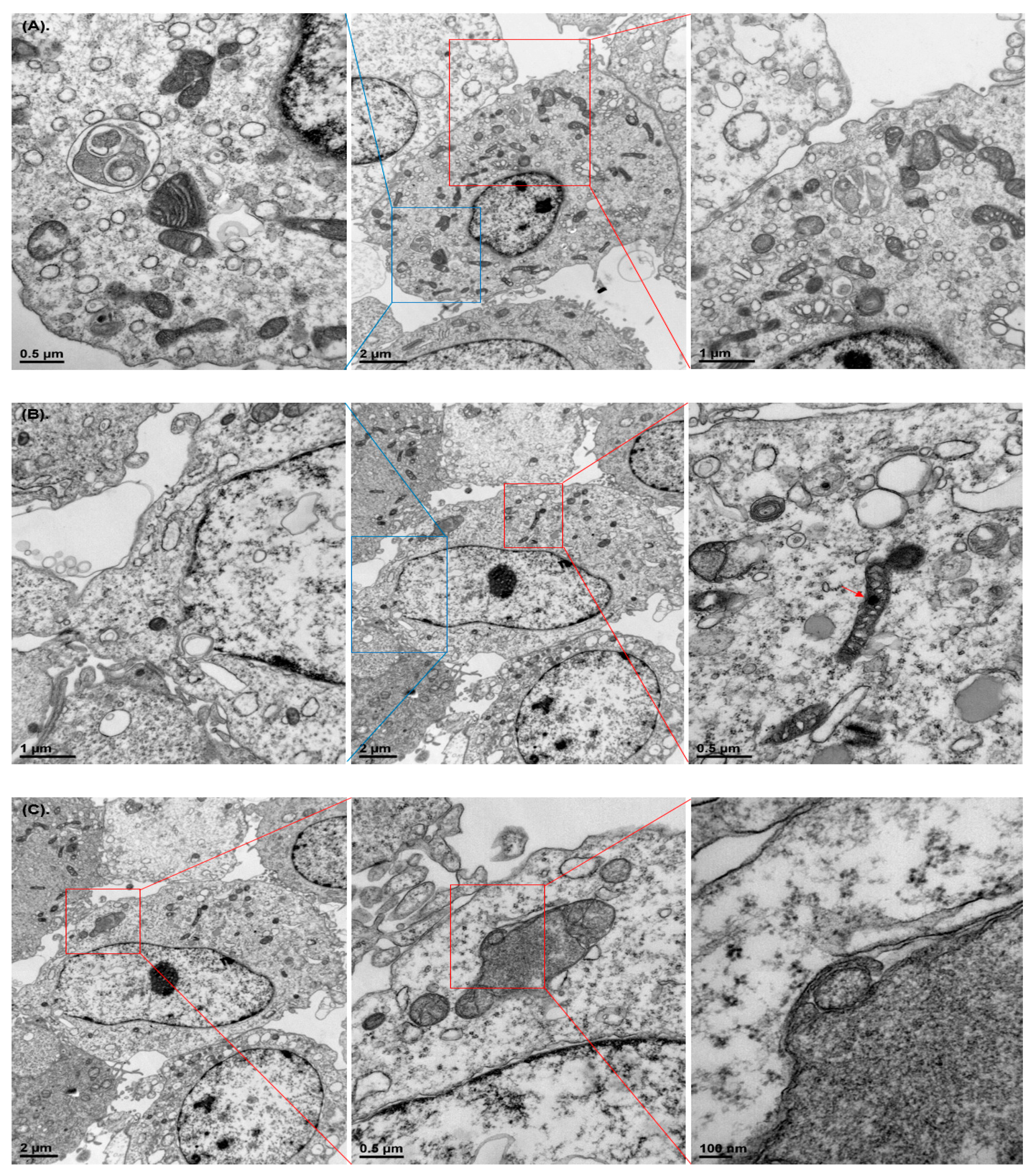
2.4. Transmission Electron Microscopy (TEM)
2.5. Measurement of Intracellular Reactive Oxygen Species
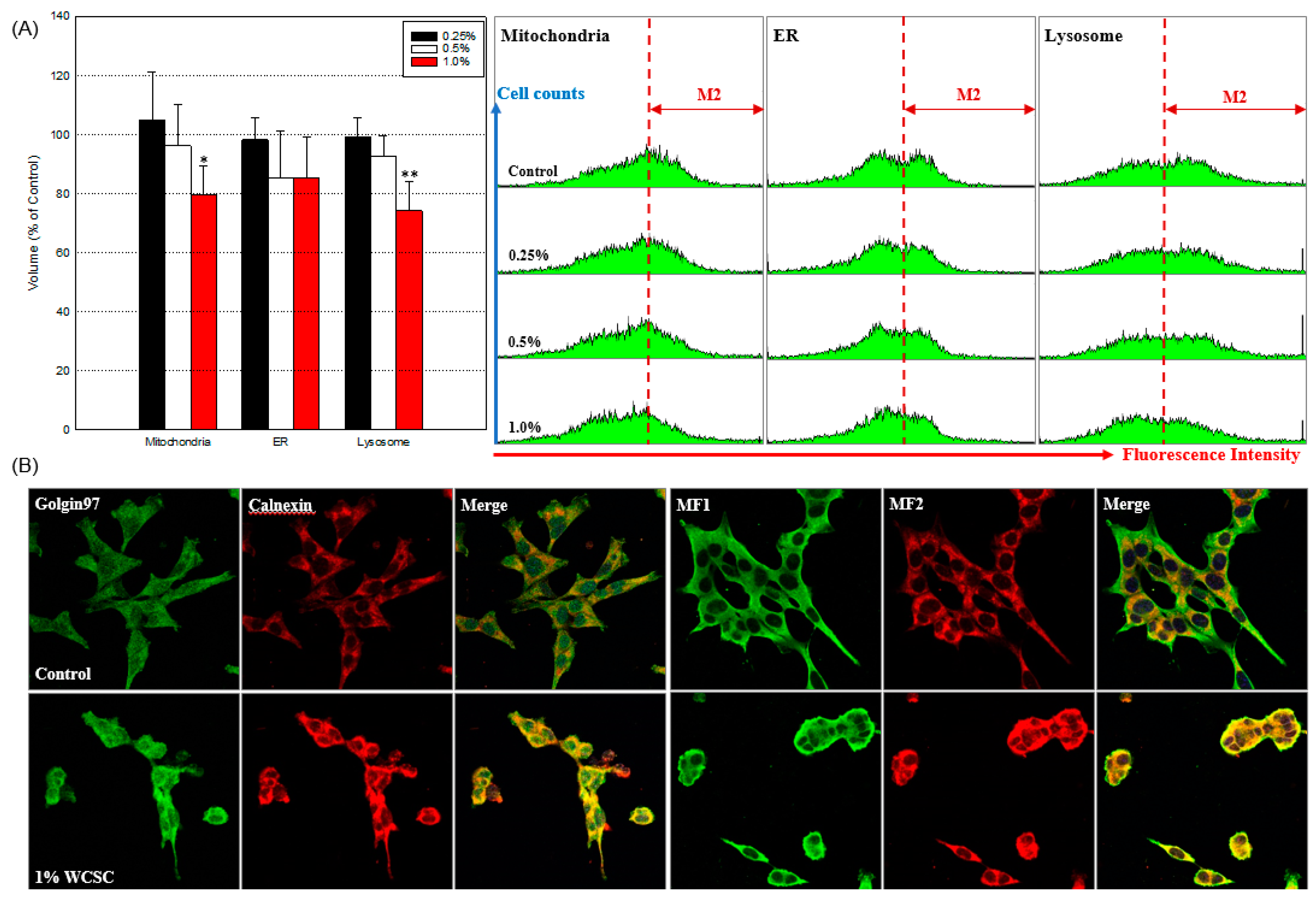
2.6. Effects on Organelle Structure and Function
2.7. Gene Expression
2.8. Protein Expression
2.9. ELISA
2.10. FACS Analysis
2.11. Statistical Analyses
3. Results
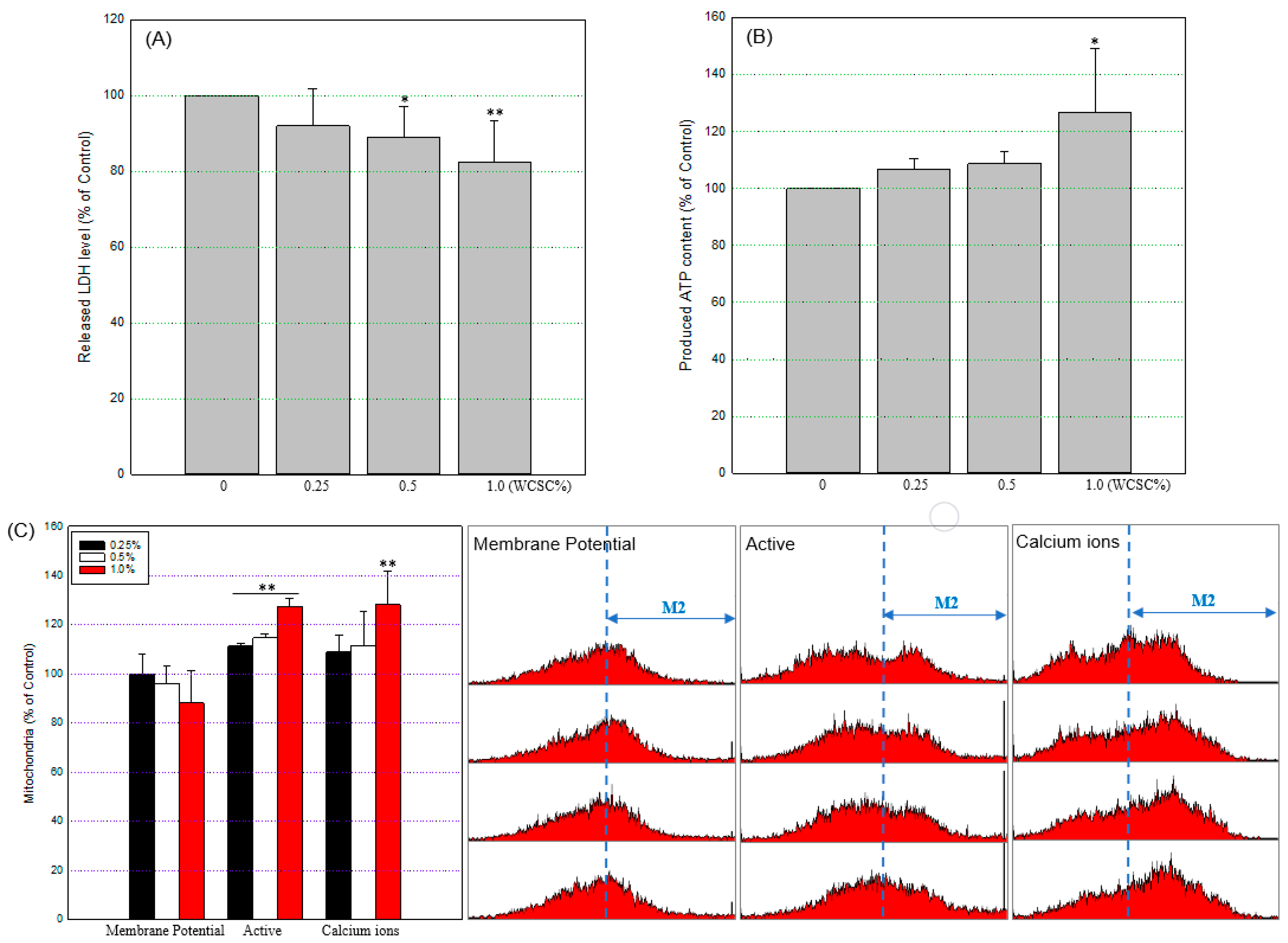
3.1. Reduced Cell Proliferation
3.2. Damaged Organelle Structures
3.3. Mitochondrial Energy Metabolism
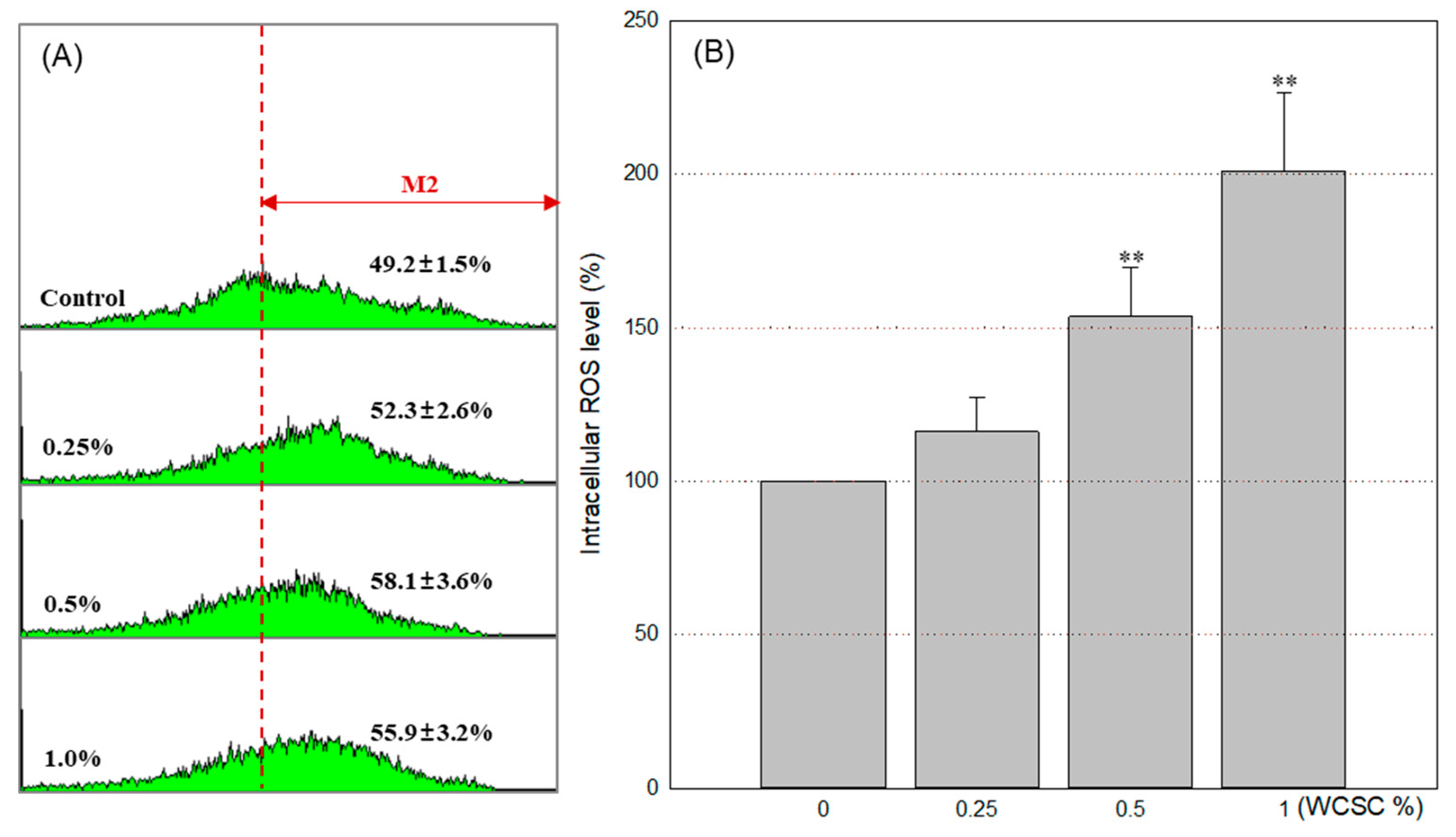
3.4. Increased Intracellular ROS
3.5. Alteration in Gene Profiles
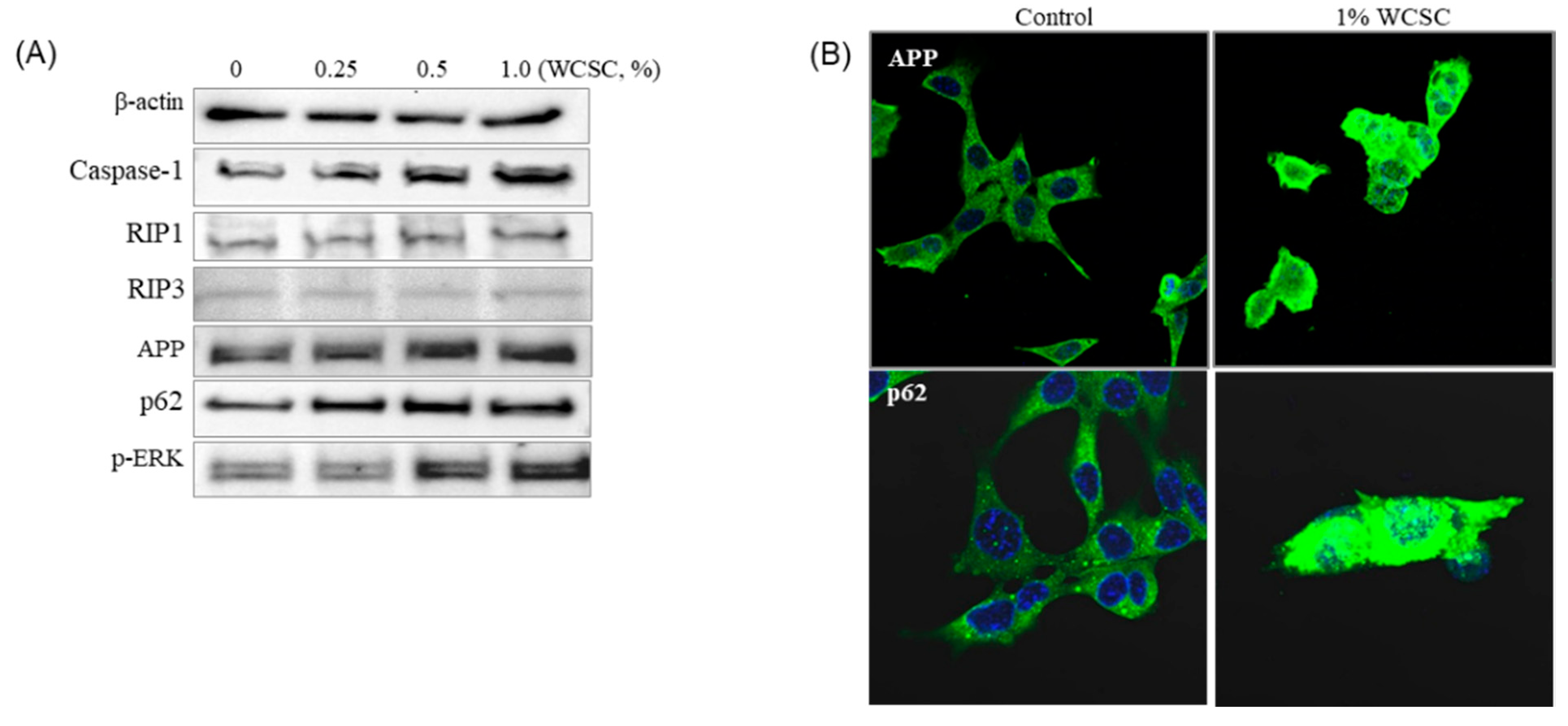
3.6. Accumulation of APP Protein
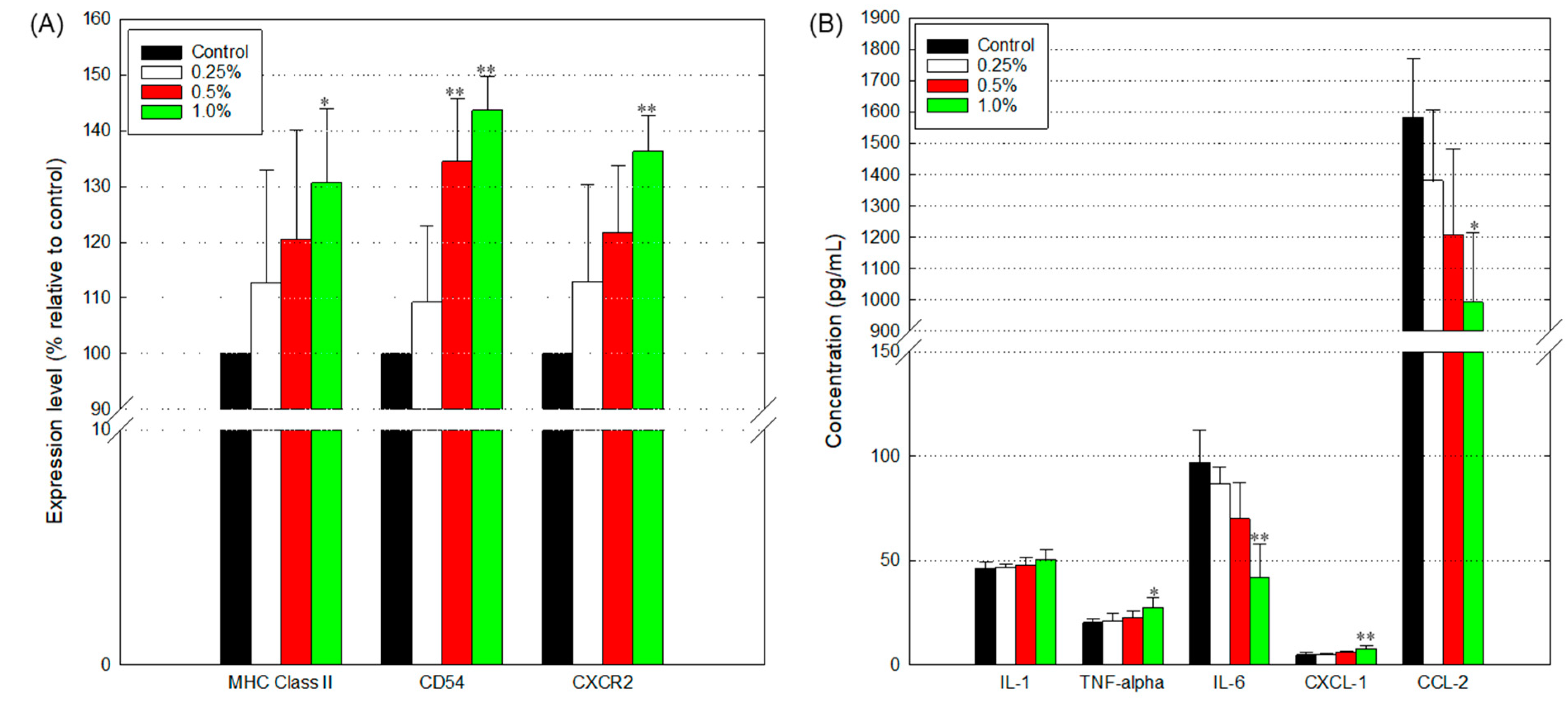
3.7. Disturbed Immune Response
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Desai, A.K.; Grossberg, G.T.; Chibnall, J.T. Healthy brain aging: A road map. Clin. Geriatr. Med. 2010, 26, 1–16. [Google Scholar] [CrossRef]
- World Health Organization. Dementia. 2019. Available online: https://www.who.int/news-room/fact-sheets/detail/dementia (accessed on 11 February 2021).
- Gibson, G.; Gunasekera, N.; Lee, M.; Lelyveld, V.; El-Agnaf, O.M.; Wright, A.; Austen, B. Oligomerization and neurotoxicity of the amyloid Adan peptide implicated in familial Danish dementia. J. Neurochem. 2004, 88, 281–290. [Google Scholar] [CrossRef]
- DaRocha-Souto, B.; Scotton, T.C.; Coma, M.; Serrano-Pozo, A.; Hashimoto, T.; Serenó, L.; Rodríguez, M.; Sánchez, B.; Hyman, B.T.; Gómez-Isla, T. Brain oligomeric β-amyloid but not total amyloid plaque burden correlates with neuronal loss and astrocyte inflammatory response in amyloid precursor protein/Tau transgenic mice. J. Neuropathol. Exp. Neurol. 2011, 70, 360–376. [Google Scholar] [CrossRef] [Green Version]
- Hamley, I.W. The amyloid beta peptide: A chemist’s perspective. Role in Alzheimer’s and fibrillization. Chem. Rev. 2012, 112, 5147–5192. [Google Scholar] [CrossRef]
- Durazzo, T.C.; Mattsson, N.; Weiner, M.W. Alzheimer’s Disease Neuroimaging Initiative. Smoking and increased Alzheimer’s disease risk: A review of potential mechanisms. Alzheimers Dement. 2014, 10 (Suppl. S3), S122–S145. [Google Scholar] [CrossRef] [Green Version]
- Giunta, B.; Deng, J.; Jin, J.; Sadic, E.; Rum, S.; Zhou, H.; Sanberg, P.; Tan, J. Evaluation of how cigarette smoke is a direct risk factor for alzheimer’s disease. Technol. Innov. 2012, 14, 39–48. [Google Scholar] [CrossRef]
- Moreno-Gonzalez, I.; Estrada, L.D.; Sanchez-Mejias, E.; Soto, C. Smoking exacerbates amyloid pathology in a mouse model of Alzheimer’s disease. Nat. Commun. 2013, 4, 1495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hurley, M.A. Light smoking at base-line predicts a higher mortality risk to women than to men; evidence from a cohort with long follow-up. BMC Public Health 2014, 14, 95. [Google Scholar] [CrossRef] [Green Version]
- Liddelow, S.A.; Barres, B.A. Reactive Astrocytes: Production, Function, and Therapeutic Potential. Immunity 2017, 46, 957–967. [Google Scholar] [CrossRef] [Green Version]
- Herculano-Houzel, S. The glia/neuron ratio: How it varies uniformly across brain structures and species and what that means for brain physiology and evolution. Glia 2014, 62, 1377–1391. [Google Scholar] [CrossRef]
- Cohen, J.; Torres, C. Astrocyte senescence: Evidence and significance. Aging Cell. 2019, 18, e12937. [Google Scholar] [CrossRef] [Green Version]
- Fernandez, C.G.; Hamby, M.E.; McReynolds, M.L.; Ray, W.J. The role of APOE4 in disrupting the homeostatic functions of astrocytes and microglia in aging and Alzheimer’s Disease. Front. Aging Neurosci. 2019, 11, 14. [Google Scholar] [CrossRef] [Green Version]
- Verkhratsky, A.; Olabarria, M.; Noristani, H.N.; Yeh, C.Y.; Rodriguez, J.J. Astrocytes in Alzheimer’s disease. Neurotherapeutics 2010, 7, 399–412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aschner, M. Astrocytes as mediators of immune and inflammatory responses in the CNS. Neurotoxicology 1998, 19, 269–281. [Google Scholar]
- Dong, Y.; Benveniste, E.N. Immune function of astrocytes. Glia 2001, 36, 180–190. [Google Scholar] [CrossRef] [PubMed]
- Phatnani, H.; Maniatis, T. Astrocytes in neurodegenerative disease. Cold Spring Harb. Perspect. Biol. 2015, 7, a020628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sica, R.E.; Caccuri, R.; Quarracino, C.; Capani, F. Are astrocytes executive cells within the central nervous system? Arq. Neuropsiquiatr. 2016, 74, 671–678. [Google Scholar] [CrossRef]
- Park, E.J.; Park, Y.J.; Lee, S.J.; Lee, K.; Yoon, C. Whole cigarette smoke condensates induce ferroptosis in human bronchial epithelial cells. Toxicol. Lett. 2019, 303, 55–66. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Cheng, Y.; Mao, C.; Liu, S.; Xiao, D.; Huang, J.; Tao, Y. Emerging mechanisms and targeted therapy of ferroptosis in cancer. Mol. Ther. 2021. [Google Scholar] [CrossRef] [PubMed]
- Jovanovic, K.; Chetty, C.J.; Khumalo, T.; Da Costa Dias, B.; Ferreira, E.; Malindisa, S.T.; Caveney, R.; Letsolo, B.T.; Weiss, S.F. Novel patented therapeutic approaches targeting the 37/67 kDa laminin receptor for treatment of cancer and Alzheimer’s disease. Expert Opin. Ther. Pat. 2015, 25, 567–582. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.L.; Yu, J.T.; Wang, H.F.; Meng, X.F.; Tan, C.C.; Wang, C.; Tan, L. Association between cancer and Alzheimer’s disease: Systematic review and meta-analysis. J. Alzheimers Dis. 2014, 42, 565–573. [Google Scholar] [CrossRef]
- Ou, S.M.; Lee, Y.J.; Hu, Y.W.; Liu, C.J.; Chen, T.J.; Fuh, J.L.; Wang, S.J. Does Alzheimer’s disease protect against cancers? A nationwide population-based study. Neuroepidemiology 2013, 40, 42–49. [Google Scholar] [CrossRef] [PubMed]
- Van der Willik, K.D.; Schagen, S.B.; Ikram, M.A. Cancer and dementia: Two sides of the same coin? Eur. J. Clin. Investig. 2018, 48, e13109. [Google Scholar] [CrossRef]
- Wang, H.; Tan, M.S.; Lu, R.C.; Yu, J.T.; Tan, L. Heat shock proteins at the crossroads between cancer and Alzheimer’s disease. Biomed. Res. Int. 2014, 2014, 239164. [Google Scholar] [CrossRef]
- Huppmann, S.; Römer, S.; Altmann, R.; Obladen, M.; Berns, M. 17β-Estradiol attenuates hyperoxia-induced apoptosis in mouse C8-D1A cell line. J. Neurosci. Res. 2008, 86, 3420–3426. [Google Scholar] [CrossRef] [PubMed]
- Ishihara, N.; Eura, Y.; Mihara, K. Mitofusin 1 and 2 play distinct roles in mitochondrial fusion reactions via GTPase activity. J. Cell Sci. 2004, 117 Pt 26, 6535–6546. [Google Scholar] [CrossRef] [Green Version]
- Wu, F.; Zhao, Y.; Jiao, T.; Shi, D.; Zhu, X.; Zhang, M.; Shi, M.; Zhou, H. CXCR2 is essential for cerebral endothelial activation and leukocyte recruitment during neuroinflammation. J. Neuroinflamm. 2015, 12, 98. [Google Scholar] [CrossRef] [Green Version]
- Linnerbauer, M.; Wheeler, M.A.; Quintana, F.J. Astrocyte crosstalk in CNS inflammation. Neuron 2020, 108, 608–622. [Google Scholar] [CrossRef] [PubMed]
- Palpagama, T.H.; Waldvogel, H.J.; Faull, R.L.M.; Kwakowsky, A. The role of microglia and astrocytes in Huntington’s disease. Front. Mol. Neurosci. 2019, 12, 258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sofroniew, M.V. Astrocyte barriers to neurotoxic inflammation. Nat. Rev. Neurosci. 2015, 16, 249–263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tabata, H. Diverse subtypes of astrocytes and their development during corticogenesis. Front. Neurosci. 2015, 9, 114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matejuk, A.; Ransohoff, R.M. Crosstalk between astrocytes and microglia: An overview. Front. Immunol. 2020, 11, 1416. [Google Scholar] [CrossRef]
- Linnerberg, C.; Toft, C.L.F.; Kjaer-Sorensen, K.; Laursen, L.S. L1cam-mediated developmental processes of the nervous system are differentially regulated by proteolytic processing. Sci. Rep. 2019, 9, 3716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohajeri, M.H.; Bartsch, U.; van der Putten, H.; Sansig, G.; Mucke, L.; Schachner, M. Neurite outgrowth on non-permissive substrates in vitro is enhanced by ectopic expression of the neural adhesion molecule L1 by mouse astrocytes. Eur. J. Neurosci. 1996, 8, 1085–1097. [Google Scholar] [CrossRef] [PubMed]
- Wolfer, D.P.; Mohajeri, H.M.; Lipp, H.P.; Schachner, M. Increased flexibility and selectivity in spatial learning of transgenic mice ectopically expressing the neural cell adhesion molecule L1 in astrocytes. Eur. J. Neurosci. 1998, 10, 708–717. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Qin, C.; Huang, J.; Tang, X.; Liu, C.; Huang, K.; Xu, J.; Guo, G.; Tong, A.; Zhou, L. The role of astrocytes in oxidative stress of central nervous system: A mixed blessing. Cell Prolif. 2020, 53, e12781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.; Dai, C.; Fan, Y.; Guo, B.; Ren, K.; Sun, T.; Wang, W. From autophagy to mitophagy: The roles of p62 in neurodegenerative diseases. J. Bioenerg. Biomembr. 2017, 49, 413–422. [Google Scholar] [CrossRef] [PubMed]
- Genc, S.; Kurnaz, I.A.; Ozilgen, M. Astrocyte- neuron lactate shuttle may boost more ATP supply to the neuron under hypoxic conditions—In silico study supported by in vitro expression data. BMC Syst. Biol. 2011, 5, 162. [Google Scholar] [CrossRef] [Green Version]
- Lemire, J.; Mailloux, R.J.; Appanna, V.D. Mitochondrial lactate dehydrogenase is involved in oxidative-energy metabolism in human astrocytoma cells (CCF-STTG1). J. Neurosci. Res. 2008, 92, 464–475. [Google Scholar] [CrossRef]
- Magistretti, P.J.; Allaman, I. Lactate in the brain: From metabolic end-product to signalling molecule. Nat. Rev. Neurosci. 2018, 19, 235–249. [Google Scholar] [CrossRef] [PubMed]
- Yellen, G. Fueling thought: Management of glycolysis and oxidative phosphorylation in neuronal metabolism. J. Cell Biol. 2018, 217, 2235–2246. [Google Scholar] [CrossRef] [PubMed]
- Youle, R.J.; van der Bliek, A.M. Mitochondrial fission, fusion, and stress. Science 2012, 337, 1062–1065. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.J.; Ye, L.; Huang, W.F.; Guo, L.J.; Xu, Z.G.; Wu, H.L.; Yang, C.; Liu, H.F. p62 links the autophagy pathway and the ubiqutin-proteasome system upon ubiquitinated protein degradation. Cell Mol. Biol. Lett. 2016, 21, 29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, M.; Chen, Z.; Wang, C.; Song, L.; Zou, Y.; Zhang, L.; Hui, R.; Wang, J. Cardiac-specific overexpression of miR-222 induces heart failure and inhibits autophagy in mice. Cell Physiol. Biochem. 2016, 39, 1503–1511. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Yang, J.; Li, F.; Lian, G.; Ouyang, M. Mir-29a inhibited intestinal epithelial cells autophagy partly by decreasing ATG9A in ulcerative colitis. Anticancer Drugs 2018, 29, 652–659. [Google Scholar] [CrossRef]
- Xu, J.; Su, Y.; Xu, A.; Fan, F.; Mu, S.; Chen, L.; Chu, Z.; Zhang, B.; Huang, H.; Zhang, J.; et al. miR-221/222-mediated inhibition of autophagy promotes dexamethasone resistance in multiple myeloma. Mol. Ther. 2019, 27, 559–570. [Google Scholar] [CrossRef] [Green Version]
- Chen, M.J.; Ramesha, S.; Weinstock, L.D.; Gao, T.; Ping, L.; Xiao, H.; Dammer, E.B.; Duong, D.D.; Levey, A.I.; Lah, J.J.; et al. Extracellular signal-regulated kinase regulates microglial immune responses in Alzheimer’s disease. J. Neurosci. Res. 2020, 99, 1704–1721. [Google Scholar] [CrossRef] [PubMed]
- Michael, B.D.; Bricio-Moreno, L.; Sorensen, E.W.; Miyabe, Y.; Lian, J.; Solomon, T.; Kurt-Jones, E.A.; Luster, A.D. Astrocyte- and neuron-derived CXCL1 drives neutrophil transmigration and blood-brain barrier permeability in viral encephalitis. Cell Rep. 2020, 32, 108150. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Primer | Sequence |
|---|---|---|
| β-Actin | Forward | TTCTTTGCAGCTCCTTCGTT |
| Reverse | CGCAGCTCATTGTAGAAGGT | |
| Catalase | Forward | TTCGTCCCGAGTCTCTCCAT |
| Reverse | GAGTGTCCGGGTAGGCAAAA | |
| GADD45a | Forward | TGGAGGAAGTGCTCAGCAAG |
| Reverse | GTCATCTCTGAGCCCTCGTG | |
| HO-1 | Forward | AACAAGCAGAACCCAGTCTA |
| Reverse | CCTTCTGTGCAATCTTCTTC | |
| NQO-1 | Forward | GGTAGCGGCTCCATGTACTC |
| Reverse | TGCCCTGAGGCTCCTAATCT | |
| PMP2 | Forward | AAGCCAAGAGCATCGTGACA |
| Reverse | CCCGTTAGAGCGACTCATCC | |
| SCF | Forward | GCAGACACTGGGTCTCGATT |
| Reverse | GTGGTAGGGACCTTGGGTTG |
| (A) Gene List Upregulated More Than 1.6 Folds | |||||
| Gene_Symbol | mRNA Accession | Folds | Gene_Symbol | mRNA Accession | Folds |
| Hmox1 | NM_010442 | 5.85 | Npc1 | NM_008720 | 1.78 |
| Slc7a11 | NM_011990 | 3.90 | Pgd | NM_001081274 | 1.77 |
| Atp13a4 | NM_001164612 | 3.14 | Disp2 | NM_170593 | 1.77 |
| Cd68 | NM_001291058 | 2.87 | Sh2d6 | XM_006506833 | 1.77 |
| Nqo1 | NM_008706 | 2.84 | Cyb5r1 | NM_028057 | 1.76 |
| Abcc4 | NM_001033336 | 2.75 | Gm23302 | ENSMUST00000082710 | 1.75 |
| Blvrb | NM_001290525 | 2.74 | Il34 | NM_001135100 | 1.75 |
| Mir29a | NR_029744 | 2.55 | Vmn1r62 | NM_030741 | 1.74 |
| Gclm | NM_008129 | 2.44 | Ampd3 | NM_001276301 | 1.73 |
| Gbe1 | NM_028803 | 2.40 | Adh7 | NM_009626 | 1.73 |
| Sox9 | NM_011448 | 2.35 | Sqstm1 | NM_001290769 | 1.73 |
| Gm25126 | ENSMUST00000083343 | 2.30 | Cth | NM_145953 | 1.72 |
| Gm25799 | ENSMUST00000082643 | 2.27 | Cat | NM_009804 | 1.72 |
| Slc40a1 | NM_016917 | 2.25 | Gm3170 | ENSMUST00000168753 | 1.71 |
| Gm10701 | ENSMUST00000098926 | 2.23 | Dusp5 | NM_001085390 | 1.71 |
| Gm25121 | ENSMUST00000179846 | 2.20 | Ptchd1 | NM_001093750 | 1.70 |
| Aldh3a1 | NM_001112725 | 2.12 | Lyst | NM_010748 | 1.69 |
| Fbxo2 | NM_176848 | 2.12 | Bcl6 | NM_009744 | 1.68 |
| Tnfrsf22 | NM_001311145 | 2.12 | Sec11c | NM_025468 | 1.68 |
| Il11 | NM_001290423 | 2.12 | BC048507 | NM_001001185 | 1.67 |
| Ypel5 | NM_027166 | 2.11 | Gtpbp2 | NM_001145979 | 1.67 |
| Sp140 | NM_001013817 | 2.03 | Acot2 | NM_134188 | 1.67 |
| Zfand2a | NM_001159908 | 2.02 | Anxa7 | NM_001110794 | 1.67 |
| 5330438D12Rik | XR_872054 | 2.00 | Prdx1 | NM_011034 | 1.67 |
| Abcc1 | NM_008576 | 1.99 | 2410006H16Rik | NR_030738 | 1.66 |
| Slc48a1 | NM_026353 | 1.98 | Tbc1d30 | NM_029057 | 1.65 |
| Mir708 | NR_030489 | 1.98 | Gm19410 | XM_006509234 | 1.65 |
| Layn | NM_001033534 | 1.97 | Uchl1 | NM_011670 | 1.65 |
| Btc | NM_007568 | 1.96 | 4931413I07Rik | ENSMUST00000173637 | 1.64 |
| Srxn1 | NM_029688 | 1.95 | Gabarapl1 | NM_020590 | 1.64 |
| Sgk1 | NM_001161845 | 1.93 | Zfp2 | NM_001044697 | 1.64 |
| Hgsnat | NM_029884 | 1.93 | Plk3 | NM_001313916 | 1.64 |
| Ptprn | NM_008985 | 1.89 | Taldo1 | NM_011528 | 1.63 |
| Tenm4 | NM_001310760 | 1.89 | Clcn2 | NM_009900 | 1.63 |
| Gm24695 | ENSMUST00000083877 | 1.87 | Tnfaip2 | NM_009396 | 1.63 |
| Creg1 | NM_011804 | 1.85 | Zfp945 | NM_001110254 | 1.63 |
| Mllt11 | NM_019914 | 1.85 | Zfp708 | NM_001012325 | 1.62 |
| Mir222 | NR_029807 | 1.85 | Kdm7a | NM_001033430 | 1.62 |
| Prr13 | NM_001170911 | 1.84 | Dusp4 | NM_176933 | 1.62 |
| LOC102638255 | XR_374951 | 1.83 | Lonp1 | NM_028782 | 1.62 |
| Gm25026 | ENSMUST00000175017 | 1.82 | Hid1 | NM_175454 | 1.62 |
| Gm24431 | ENSMUST00000104073 | 1.81 | Cdkn1a | NM_001111099 | 1.61 |
| Chil3 | NM_009892 | 1.81 | Ptk2b | NM_001162365 | 1.61 |
| KnowTID_00001299 | 1.81 | Rin2 | NM_028724 | 1.61 | |
| Tmem158 | NM_001002267 | 1.80 | Ngf | NM_001112698 | 1.61 |
| Esd | NM_001285423 | 1.80 | Cobll1 | NM_027225 | 1.61 |
| Smox | NM_001177833 | 1.79 | AA467197 | NM_001004174 | 1.60 |
| Fosl1 | NM_010235 | 1.78 | Clcf1 | NM_001310038 | 1.60 |
| (B) Gene List Downregulated More Than 1.9 Folds | |||||
| Gene_Symbol | mRNA Accession | Folds | Gene_Symbol | mRNA Accession | Folds |
| Pmp2 | NM_001030305 | -7.90 | Vmn1r103 | NM_001166737 | -2.20 |
| Gm25664 | ENSMUST00000157697 | -4.74 | Plce1 | NM_019588 | -2.20 |
| Chl1 | NM_007697 | -4.02 | Mcam | NM_023061 | -2.19 |
| Plekhg1 | NM_001033253 | -3.85 | Ogn | NM_008760 | -2.19 |
| Sntb1 | NM_016667 | -3.67 | Cyp51 | NM_020010 | -2.18 |
| Fndc1 | NM_001081416 | -3.62 | Prss23 | NM_029614 | -2.18 |
| Slitrk6 | NM_175499 | -3.50 | Hist1h2ab | NM_175660 | -2.18 |
| Gm22935 | ENSMUST00000103960 | -3.41 | Gas7 | NM_001109657 | -2.17 |
| Hmgcs1 | NM_001291439 | -3.19 | Lrrc8b | NM_001033550 | -2.17 |
| Cd200 | NM_010818 | -3.17 | Dhrs3 | NM_001172424 | -2.14 |
| Itga2 | NM_008396 | -3.16 | Hist4h4 | NM_175652 | -2.13 |
| Ly6a | NM_001271416 | -3.06 | Gpc4 | NM_008150 | -2.13 |
| Slc35f1 | NM_178675 | -3.04 | Epb41l4b | NM_019427 | -2.11 |
| Gm22908 | ENSMUST00000177620 | -2.98 | Sorbs1 | NM_001034962 | -2.09 |
| Gm26140 | ENSMUST00000103807 | -2.98 | H2afx | NM_010436 | -2.09 |
| Gm23256 | ENSMUST00000102235 | -2.98 | Dhcr7 | NM_007856 | -2.09 |
| Ctgf | NM_010217 | -2.97 | Lgi4 | NM_144556 | -2.08 |
| Has2 | NM_008216 | -2.87 | Pls1 | NM_001033210 | -2.08 |
| Adam19 | NM_001291890 | -2.83 | Lss | NM_146006 | -2.07 |
| Rgs8 | NM_026380 | -2.82 | Cmtm5 | NM_026066 | -2.07 |
| Vgll3 | NM_028572 | -2.78 | Prss23os | ENSMUST00000032858 | -2.06 |
| Deptor | NM_001037937 | -2.75 | Arhgef9 | NM_001033329 | -2.06 |
| Postn | NM_001198765 | -2.75 | Olfr97 | NM_146512 | -2.05 |
| Rassf4 | NM_178045 | -2.73 | Acta2 | NM_007392 | -2.05 |
| Plxnb3 | NM_019587 | -2.72 | Rcan2 | NM_001286653 | -2.04 |
| Ldlr | NM_001252658 | -2.69 | L1cam | NM_008478 | -2.03 |
| Dhcr24 | NM_053272 | -2.66 | Hist1h2ag | NM_178186 | -2.03 |
| Igf1 | NM_001111274 | -2.62 | Nrxn1 | NM_020252 | -2.02 |
| Prelp | NM_054077 | -2.62 | Prex2 | NM_001033636 | -2.02 |
| Gabrb3 | NM_001038701 | -2.62 | Ptprz1 | NM_001081306 | -2.02 |
| Deptor | NM_001037937 | -2.61 | Gm10719 | ENSMUST00000099047 | -2.01 |
| Insig1 | NM_153526 | -2.58 | Hist1h2bf | NM_178195 | -2.01 |
| Gm10801 | ENSMUST00000099684 | -2.53 | Scn7a | NM_009135 | -2.00 |
| Idi1 | NM_145360 | -2.51 | Serinc5 | NM_172588 | -2.00 |
| Itga1 | NM_001033228 | -2.51 | Mef2c | NM_001170537 | -2.00 |
| Fdps | NM_001253751 | -2.50 | Lrrc8c | NM_133897 | -1.99 |
| Fabp5 | BC002008 | -2.49 | KnowTID_00007113 | -1.98 | |
| Serpina3n | NM_009252 | -2.45 | Myl9 | NM_172118 | -1.97 |
| Pappa | NM_021362 | -2.41 | Gabra2 | NM_008066 | -1.97 |
| Fabp5 | NM_001272097 | -2.41 | Hist2h2bb | NM_175666 | -1.97 |
| Rgs16 | NM_011267 | -2.40 | Gm11168 | ENSMUST00000178348 | -1.97 |
| Hist1h4n | OTTMUST00000001028 | -2.37 | Npnt | NM_001029836 | -1.96 |
| Ahr | NM_001314027 | -2.36 | Alg6 | NM_001081264 | -1.96 |
| Hist1h1b | NM_020034 | -2.36 | Pcsk6 | NM_001291184 | -1.96 |
| Iqgap2 | NM_027711 | -2.35 | Hist1h2bn | NM_178201 | -1.96 |
| Sparcl1 | NM_010097 | -2.34 | Fdft1 | NM_010191 | -1.95 |
| Nsdhl | NM_010941 | -2.34 | Tgfb2 | NM_009367 | -1.95 |
| Adamts1 | NM_009621 | -2.34 | Arhgef26 | NM_001081295 | -1.94 |
| Cp | NM_001276248 | -2.34 | Prickle1 | NM_001033217 | -1.94 |
| Hist1h1a | NM_030609 | -2.33 | Gm10715 | ENSMUST00000177969 | -1.94 |
| H1fx | NM_198622 | -2.31 | Msmo1 | NM_025436 | -1.94 |
| Kctd12 | NM_177715 | -2.29 | Thbs1 | NM_001313914 | -1.93 |
| Nid2 | NM_008695 | -2.29 | Stard13 | NM_001163493 | -1.93 |
| Gm10717 | ENSMUST00000099042 | -2.29 | Kcna1 | NM_010595 | -1.93 |
| Ephb2 | NM_001290753 | -2.29 | Afap1l2 | NM_001177796 | -1.92 |
| Cdh19 | NM_001081386 | -2.27 | Gm12688 | XR_401838 | -1.92 |
| Tubb3 | NM_023279 | -2.24 | Hist1h2af | NM_175661 | -1.92 |
| Rbms3 | NM_001172121 | -2.23 | Foxd3 | NM_010425 | -1.91 |
| Gm26519 | XR_389454 | -2.22 | Nppb | NM_001287348 | -1.90 |
| Olfr344 | NM_146628 | -2.21 | Tagln2 | NM_178598 | -1.90 |
| Gm24277 | ENSMUST00000158661 | -2.21 | Gm10717 | ENSMUST00000075573 | -1.90 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, E.-J.; Jin, S.-W.; Lim, H.-J.; Kim, H.-Y.; Kang, M.-S.; Yang, S. Whole Cigarette Smoke Condensates Induce Accumulation of Amyloid Beta Precursor Protein with Oxidative Stress in Murine Astrocytes. Toxics 2021, 9, 150. https://doi.org/10.3390/toxics9070150
Park E-J, Jin S-W, Lim H-J, Kim H-Y, Kang M-S, Yang S. Whole Cigarette Smoke Condensates Induce Accumulation of Amyloid Beta Precursor Protein with Oxidative Stress in Murine Astrocytes. Toxics. 2021; 9(7):150. https://doi.org/10.3390/toxics9070150
Chicago/Turabian StylePark, Eun-Jung, Seung-Woo Jin, Hyun-Ji Lim, Hyeon-Young Kim, Min-Sung Kang, and Siyoung Yang. 2021. "Whole Cigarette Smoke Condensates Induce Accumulation of Amyloid Beta Precursor Protein with Oxidative Stress in Murine Astrocytes" Toxics 9, no. 7: 150. https://doi.org/10.3390/toxics9070150