
Nonylphenol Polyethoxylates Enhance Adipose Deposition in Developmentally Exposed Zebrafish

 , ,
, ,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Cell Care
2.3. Adipogenic Differentiation and Outcome Measurements in hMSCs
2.4. Zebrafish Housing and Care
2.5. Zebrafish Exposures
2.6. Zebrafish Metabolic Health Determinations
2.7. Statistical Analysis
3. Results
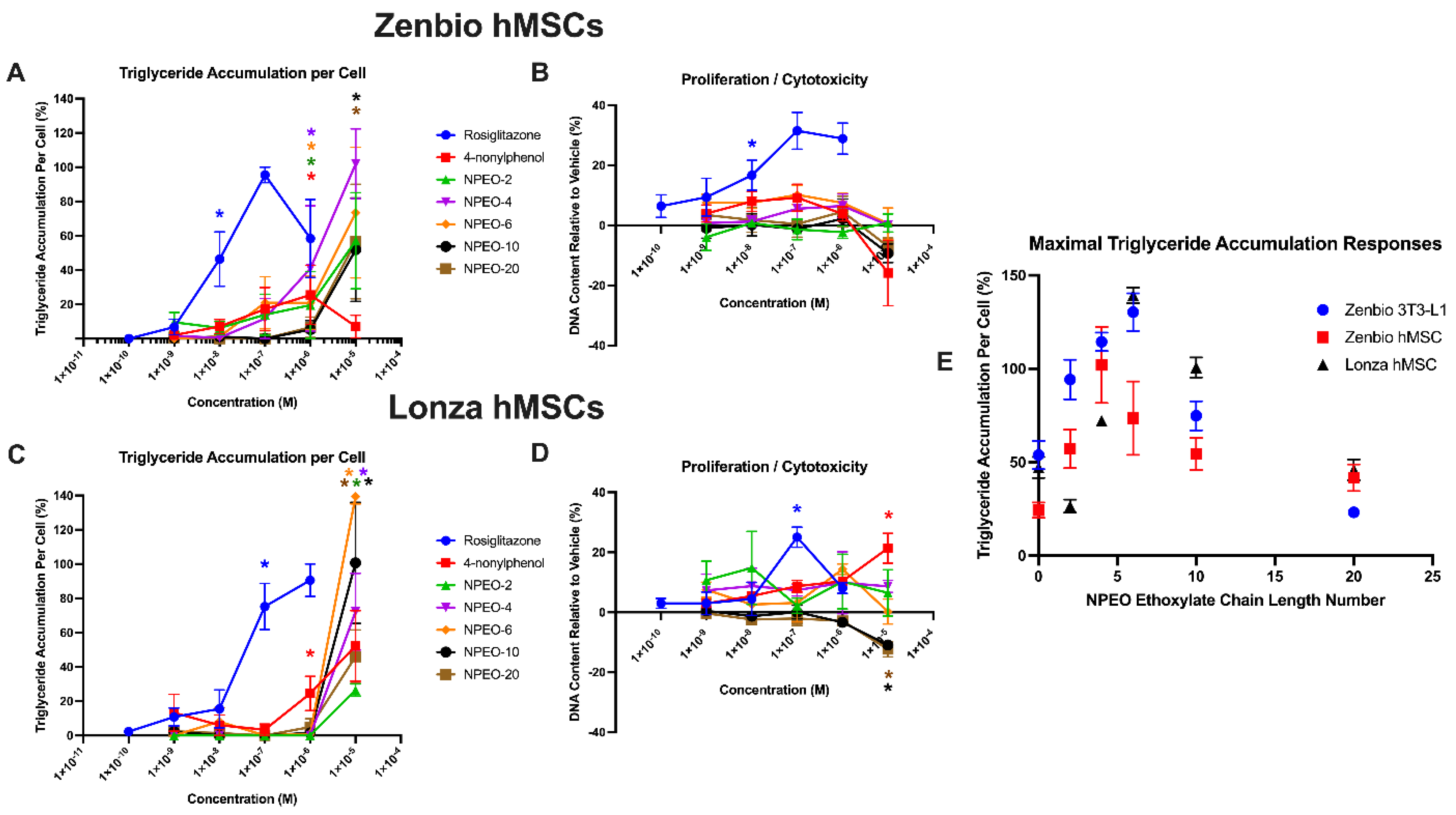
3.1. Adipogenic Activity of NPEOs in MSC Models
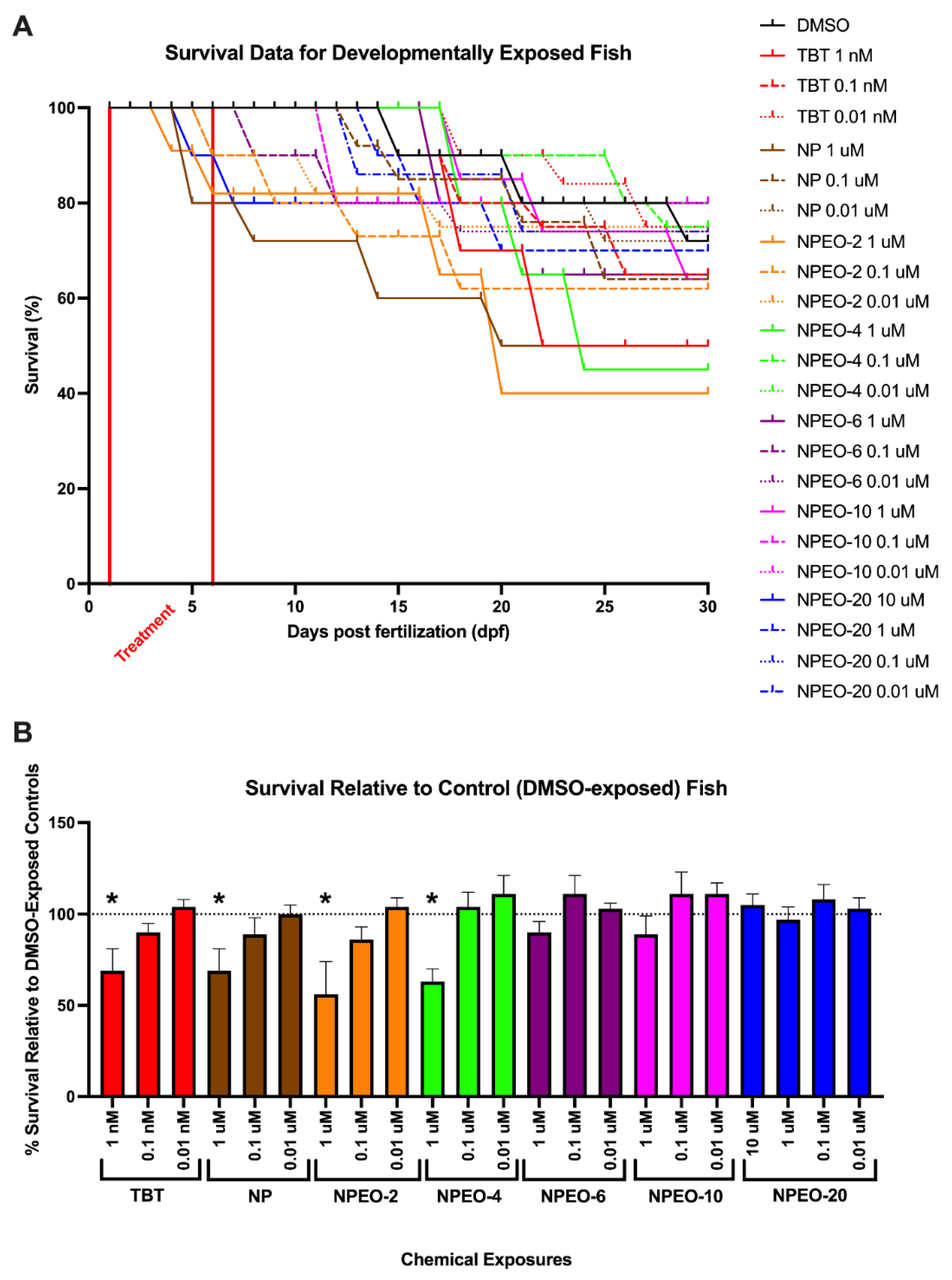
3.2. Lethality of NPEOs on Zebrafish
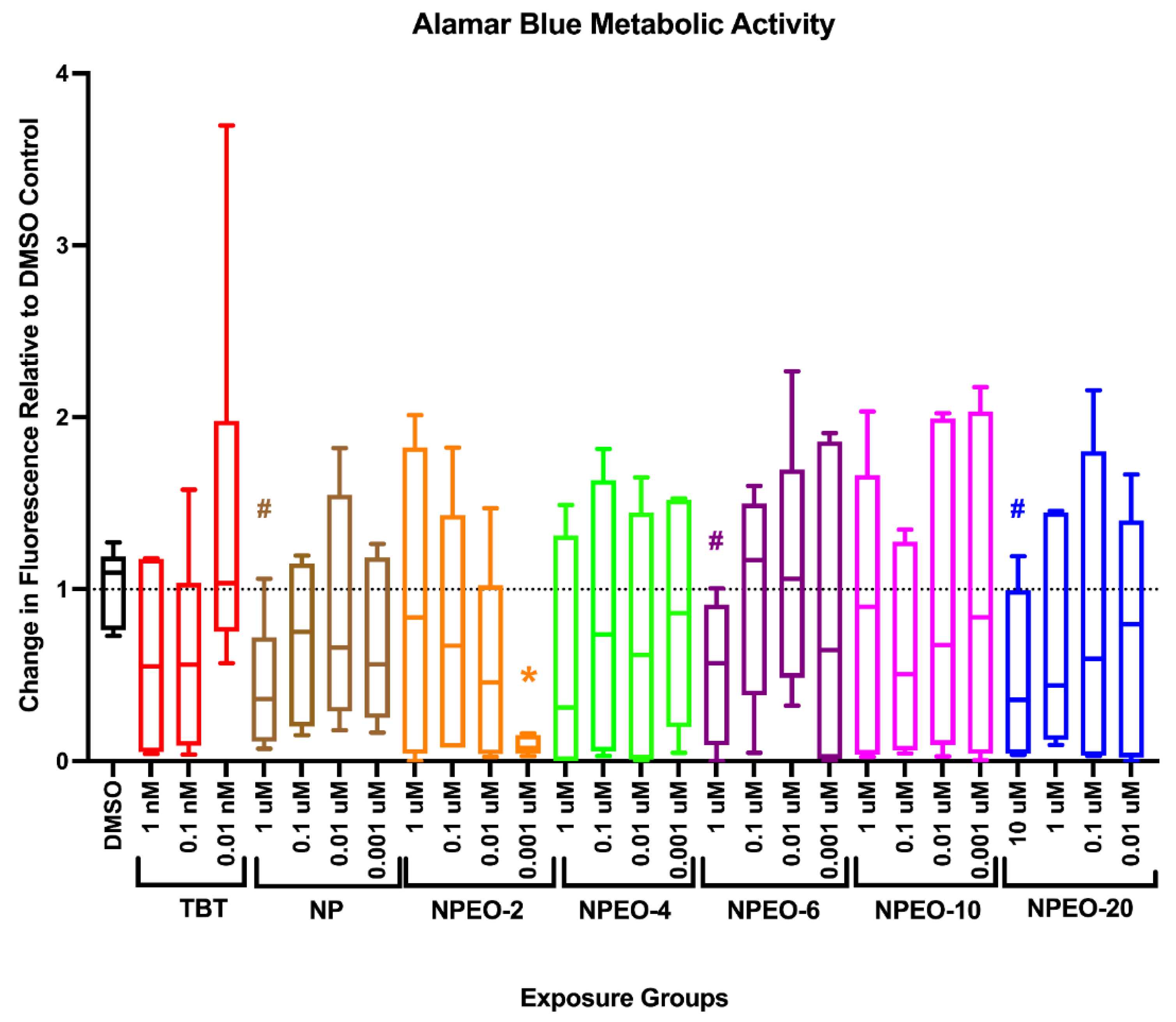
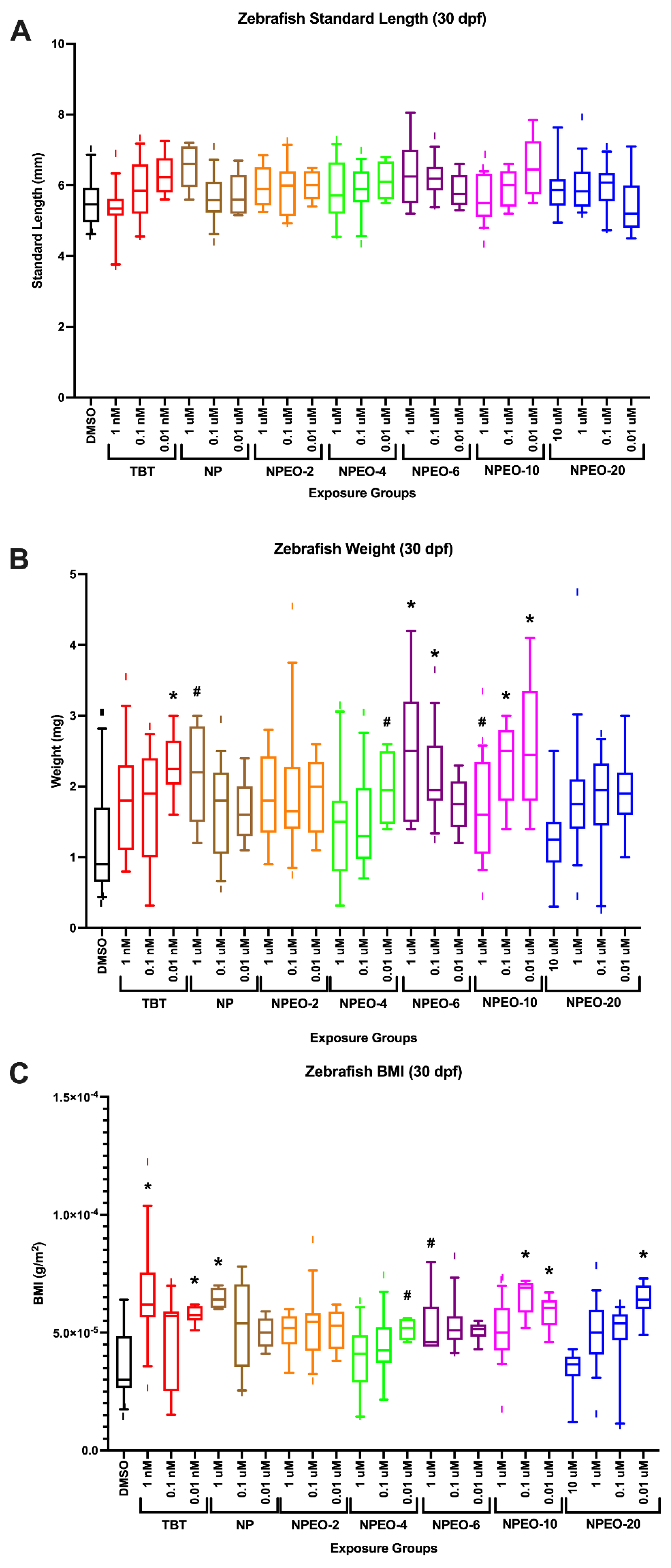
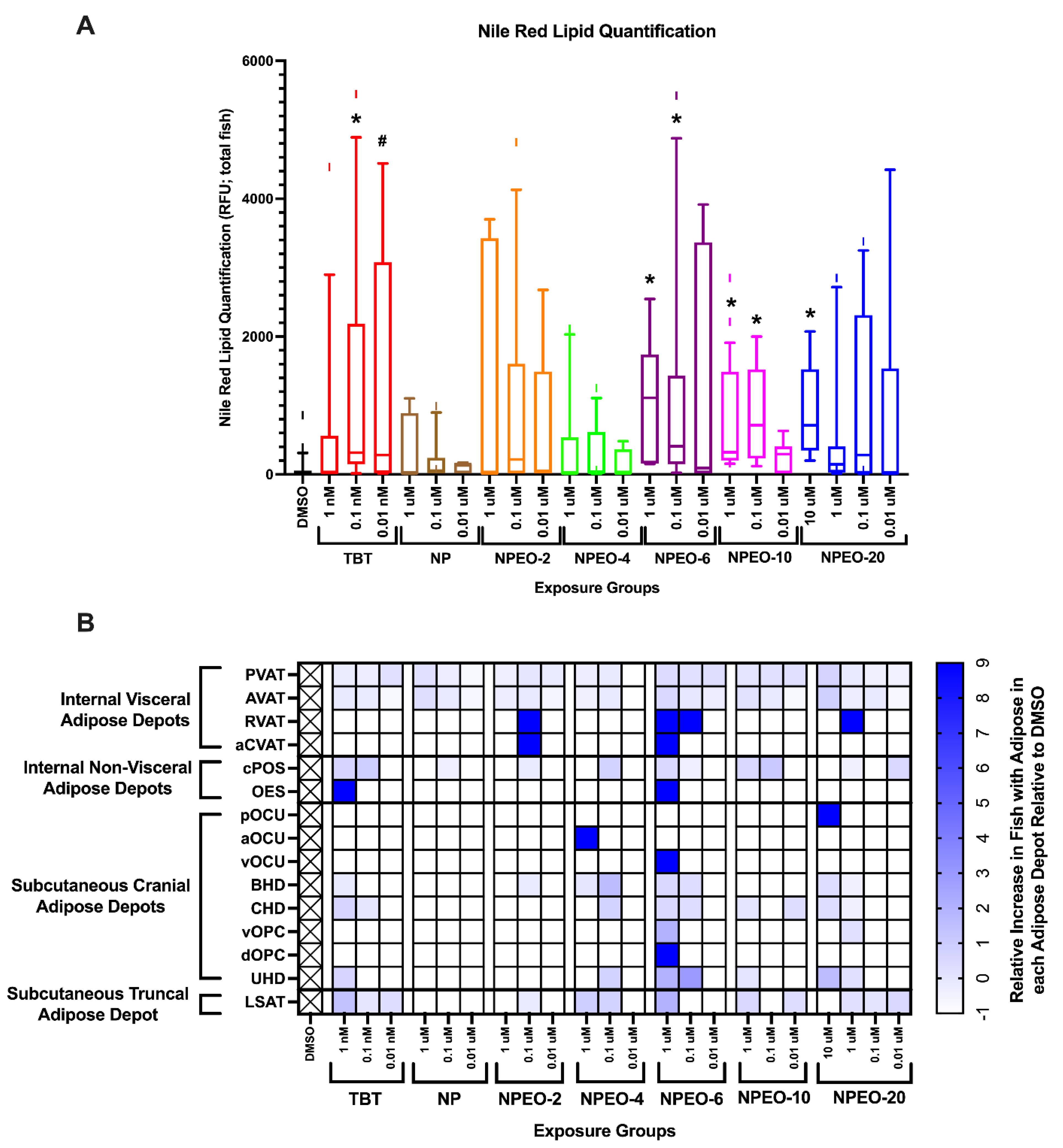
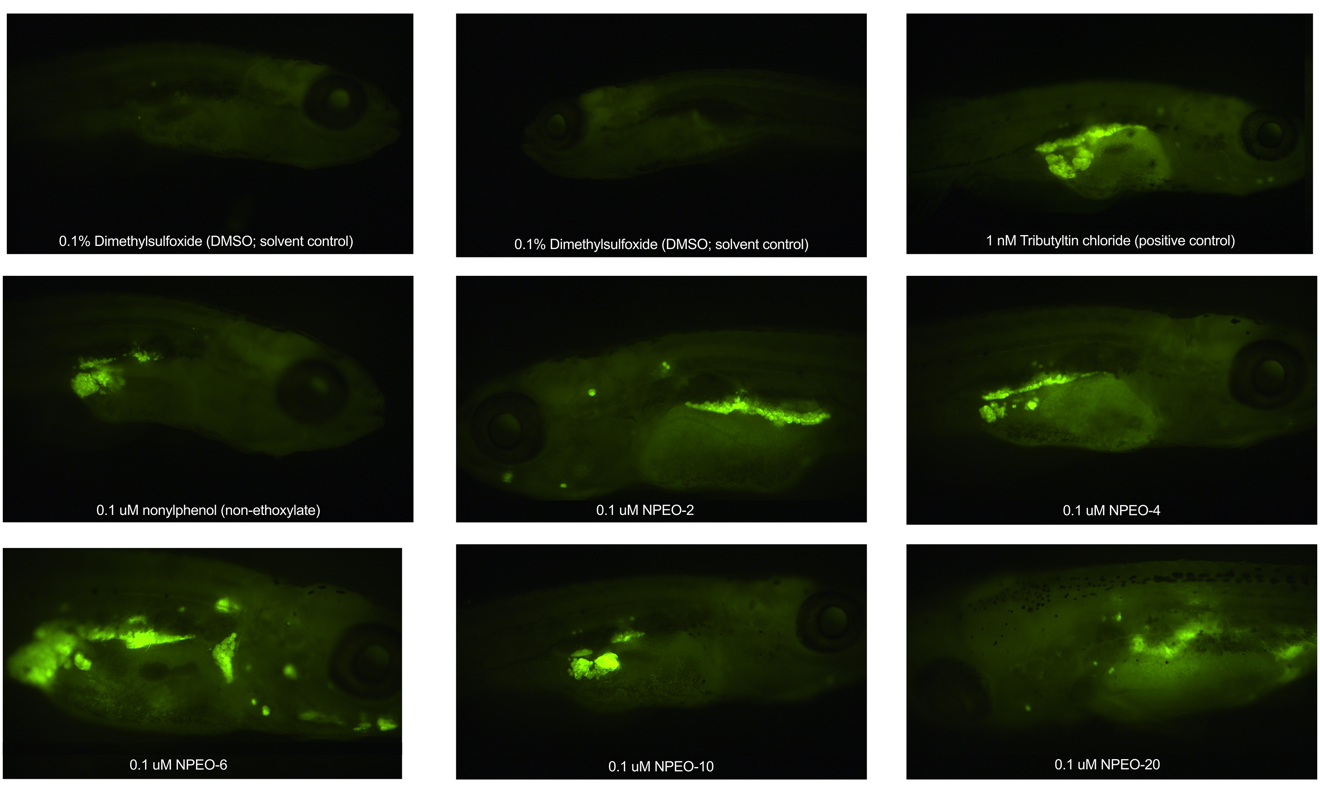
3.3. Metabolic Health, Growth Trajectory, Weights, and Adipose Deposition
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Marcomini, A.; Filipuzzi, F.; Giger, W. Aromatic surfactants in laundry detergents and hard-surface cleaners: Linear alkylbenzenesulphonates and alkylphenol polyethoxylates. Chemosphere 1988, 17, 853–863. [Google Scholar] [CrossRef]
- Marcomini, A.; Stelluto, S.; Pavoni, B. Determination of Linear Alkylbenzenesulphonates and Alkylphenol Polyethoxylates in Commercial Products and Marine Waters by Reversed- and Normal-Phase Hplc. Int. J. Environ. Anal. Chem. 1989, 35, 207–218. [Google Scholar] [CrossRef]
- Maguire, R.J. Review of the Persistence of Nonylphenol and Nonylphenol Ethoxylates in Aquatic Environments. Water Qual. Res. J. 1999, 34, 37–78. [Google Scholar] [CrossRef]
- Ying, G.-G.; Williams, B.; Kookana, R. Environmental fate of alkylphenols and alkylphenol ethoxylates—A review. Environ. Int. 2002, 28, 215–226. [Google Scholar] [CrossRef]
- Loos, R.; Hanke, G.; Umlauf, G.; Eisenreich, S.J. LC–MS–MS analysis and occurrence of octyl- and nonylphenol, their ethoxylates and their carboxylates in Belgian and Italian textile industry, waste water treatment plant effluents and surface waters. Chemosphere 2007, 66, 690–699. [Google Scholar] [CrossRef]
- Reznik, G.O.; Vishwanath, P.; Pynn, M.A.; Sitnik, J.M.; Todd, J.J.; Wu, J.; Jiang, Y.; Keenan, B.G.; Castle, A.B.; Haskell, R.F.; et al. Use of sustainable chemistry to produce an acyl amino acid surfactant. Appl. Microbiol. Biotechnol. 2010, 86, 1387–1397. [Google Scholar] [CrossRef] [PubMed]
- Ceresana. Market Study: Surfactants (2nd Edition). Available online: http://www.ceresana.com/en/market-studies/chemicals/surfactants (accessed on 1 December 2016).
- Giger, W.; Ahel, M.; Koch, M.; Laubscher, H.U.; Schaffner, C.; Schneider, J. Behaviour of alkylphenol polyethoxylate surfactants and of nitrilotriacetate in sewage treatment. Water Sci. Technol. 1987, 19, 449–460. [Google Scholar] [CrossRef]
- Giger, W.; Brunner, P.H.; Schaffner, C. 4-Nonylphenol in Sewage Sludge: Accumulation of Toxic Metabolites from Nonionic Surfactants. Science 1984, 225, 623–625. [Google Scholar] [CrossRef]
- Ahel, M.; Giger, W.; Koch, M. Behaviour of alkylphenol polyethoxylate surfactants in the aquatic environment—I. Occurrence and transformation in sewage treatment. Water Res. 1994, 28, 1131–1142. [Google Scholar] [CrossRef]
- Bandiera, S. Reproductive and Endocrine Effects of p-Nonylphenol and Methoxychlor: A Review. Immunol. Endocr. Metab. Agents Med. Chem. 2006, 6, 15–26. [Google Scholar] [CrossRef]
- Uguz, C.; Varisli, O.; Agca, C.; Agca, Y. Effects of nonylphenol on motility and subcellular elements of epididymal rat sperm. Reprod. Toxicol. 2009, 28, 542–549. [Google Scholar] [CrossRef] [PubMed]
- White, R.; Jobling, S.; Hoare, S.A.; Sumpter, J.P.; Parker, M.G. Environmentally persistent alkylphenolic compounds are estrogenic. Endocrinology 1994, 135, 175–182. [Google Scholar] [CrossRef] [PubMed]
- Talmage, S.S. Environmental and Human Safety of Major Surfactants: Alcohol Ethoxylates and Alkylphenol Ethoxylates; The Soap and Detergent Association: New York, NY, USA, 1994; p. 374. [Google Scholar]
- Marcomini, A.; Capri, S.; Giger, W. Determination of linear alkylbenzenesulphonates, alkylphenol polyethoxylates and nonylphenol in waste water by high-performance liquid chromatography after enrichment on octadecylsilica. J. Chromatogr. A 1987, 403, 243–252. [Google Scholar] [CrossRef]
- Vega-Morales, T.; Sosa-Ferrera, Z.; Santana-Rodríguez, J. Determination of alkylphenol polyethoxylates, bisphenol-A, 17α-ethynylestradiol and 17β-estradiol and its metabolites in sewage samples by SPE and LC/MS/MS. J. Hazard. Mater. 2010, 183, 701–711. [Google Scholar] [CrossRef] [PubMed]
- Clark, L.B.; Rosen, R.T.; Hartman, T.G.; Louis, J.B.; Suffet, I.H.; Lippincott, R.L.; Rosen, J.D. Determination of Alkylphenol Ethoxylates and Their Acetic Acid Derivatives in Drinking Water by Particle Beam Liquid Chromatography/Mass Spectrometry. Int. J. Environ. Anal. Chem. 1992, 47, 167–180. [Google Scholar] [CrossRef]
- Ferguson, P.L.; Bopp, R.F.; Chillrud, S.N.; Aller, R.C.; Brownawell, B.J. Biogeochemistry of Nonylphenol Ethoxylates in Urban Estuarine Sediments. Environ. Sci. Technol. 2003, 37, 3499–3506. [Google Scholar] [CrossRef]
- Thurman, E.M.; Ferrer, I.; Blotevogel, J.; Borch, T. Analysis of Hydraulic Fracturing Flowback and Produced Waters Using Accurate Mass: Identification of Ethoxylated Surfactants. Anal. Chem. 2014, 86, 9653–9661. [Google Scholar] [CrossRef]
- Getzinger, G.J.; O’Connor, M.P.; Hoelzer, K.; Drollette, B.D.; Karatum, O.; Deshusses, M.A.; Ferguson, P.L.; Elsner, M.; Plata, D.L. Natural Gas Residual Fluids: Sources, Endpoints, and Organic Chemical Composition after Centralized Waste Treatment in Pennsylvania. Environ. Sci. Technol. 2015, 49, 8347–8355. [Google Scholar] [CrossRef]
- Lester, Y.; Ferrer, I.; Thurman, E.M.; Sitterley, K.A.; Korak, J.; Aiken, G.; Linden, K.G. Characterization of hydraulic fracturing flowback water in Colorado: Implications for water treatment. Sci. Total Environ. 2015, 512-513, 637–644. [Google Scholar] [CrossRef]
- Ferguson, P.L.; Vogler, B.; Stapleton, H.M. Non-targeted analysis to assess human exposure to semi-volatile organic contaminants in the indoor environment. In Proceedings of the 63rd ASMS Conference on Mass Spectrometry and Allied Topics, St. Louis, MO, USA, 31 May–4 June 2015. [Google Scholar]
- Sabik, H.; Gagné, F.; Blaise, C.; Marcogliese, D.; Jeannot, R. Occurrence of alkylphenol polyethoxylates in the St. Lawrence River and their bioconcentration by mussels (Elliptio complanata). Chemosphere 2003, 51, 349–356. [Google Scholar] [CrossRef]
- Dodder, N.G.; Maruya, K.A.; Ferguson, P.L.; Grace, R.; Klosterhaus, S.; La Guardia, M.; Lauenstein, G.G.; Ramirez, J. Occurrence of contaminants of emerging concern in mussels (Mytilus spp.) along the California coast and the influence of land use, storm water discharge, and treated wastewater effluent. Mar. Pollut. Bull. 2014, 81, 340–346. [Google Scholar] [CrossRef] [PubMed]
- Schmitz-Afonso, I.; Loyo-Rosales, J.E.; Avilés, M.D.L.P.; Rattner, B.A.; Rice, C.P. Determination of alkylphenol and alkylphenolethoxylates in biota by liquid chromatography with detection by tandem mass spectrometry and fluorescence spectroscopy. J. Chromatogr. A 2003, 1010, 25–35. [Google Scholar] [CrossRef]
- Cailleaud, K.; Forget-Leray, J.; Souissi, S.; Lardy, S.; Augagneur, S.; Budzinski, H. Seasonal variation of hydrophobic organic contaminant concentrations in the water-column of the Seine Estuary and their transfer to a planktonic species Eurytemora affinis (Calanoïd, copepod). Part 2: Alkylphenol-polyethoxylates. Chemosphere 2007, 70, 281–287. [Google Scholar] [CrossRef]
- Bennie, D.T. Review of the Environmental Occurrence of Alkylphenols and Alkylphenol Ethoxylates. Water Qual. Res. J. 1999, 34, 79–122. [Google Scholar] [CrossRef]
- Balch, G.; Metcalfe, C. Developmental effects in Japanese medaka (Oryzias latipes) exposed to nonylphenol ethoxylates and their degradation products. Chemosphere 2006, 62, 1214–1223. [Google Scholar] [CrossRef]
- Oliveira-Filho, E.C.; Grisolia, C.K.; Paumgartten, F.J.R. Trans-generation study of the effects of nonylphenol ethoxylate on the reproduction of the snail Biomphalaria tenagophila. Ecotoxicol. Environ. Saf. 2009, 72, 458–465. [Google Scholar] [CrossRef]
- Nichols, K.M.; Snyder, E.M.; Snyder, S.A.; Pierens, S.L.; Miles-Richardson, S.R.; Giesy, J.P. Effects of nonylphenol ethoxylate exposure on reproductive output and bioindicators of environmental estrogen exposure in fathead minnows, Pimephales promelas. Environ. Toxicol. Chem. 2001, 20, 510–522. [Google Scholar] [CrossRef]
- Zoller, U. Estuarine and coastal zone marine pollution by the nonionic alkylphenol ethoxylates endocrine disrupters: Is there a potential ecotoxicological problem? Environ. Int. 2006, 32, 269–272. [Google Scholar] [CrossRef]
- Servos, M.R. Review of the Aquatic Toxicity, Estrogenic Responses and Bioaccumulation of Alkylphenols and Alkylphenol Polyethoxylates. Water Qual. Res. J. 1999, 34, 123–178. [Google Scholar] [CrossRef]
- Ashfield, L.A.; Pottinger, T.G.; Sumpter, J.P. Exposure of female juvenile rainbow trout to alkylphenolic compounds results in modifications to growth and ovosomatic index. Environ. Toxicol. Chem. 1998, 17, 679–686. [Google Scholar] [CrossRef]
- Temkin, A.M.; Bowers, R.R.; Magaletta, M.E.; Holshouser, S.; Maggi, A.; Ciana, P.; Guillette, L.J.; Bowden, J.A.; Kucklick, J.R.; Baatz, J.E.; et al. Effects of Crude Oil/Dispersant Mixture and Dispersant Components on PPARγ Activity In Vitro and In Vivo: Identification of Dioctyl Sodium Sulfosuccinate (DOSS.; CAS #577-11-7) as a Probable Obesogen. Environ. Health Perspect. 2016, 124, 112–119. [Google Scholar] [CrossRef] [PubMed]
- Bowers, R.R.; Temkin, A.M.; Guillette, L.J.; Baatz, J.E.; Spyropoulos, D.D. The commonly used nonionic surfactant Span 80 has RXRα transactivation activity, which likely increases the obesogenic potential of oil dispersants and food emulsifiers. Gen. Comp. Endocrinol. 2016, 238, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Kassotis, C.D.; Kollitz, E.M.; Ferguson, P.L.; Stapleton, H.M. Nonionic ethoxylated surfactants induce adipogenesis in 3T3-L1 cells. Toxicol. Sci. 2018, 162, 124–136. [Google Scholar] [CrossRef] [PubMed]
- Kassotis, C.D.; Hoffman, K.; Völker, J.; Pu, Y.; Veiga-Lopez, A.; Kim, S.M.; Schlezinger, J.J.; Bovolin, P.; Cottone, E.; Saraceni, A.; et al. Reproducibility of adipogenic responses to metabolism disrupting chemicals in the 3T3-L1 pre-adipocyte model system: An interlaboratory study. Toxicology 2021, 461, 152900. [Google Scholar] [CrossRef]
- Zang, L.; Maddison, L.A.; Chen, W. Zebrafish as a Model for Obesity and Diabetes. Front. Cell Dev. Biol. 2018, 6, 91. [Google Scholar] [CrossRef]
- Tingaud-Sequeira, A.; Ouadah, N.; Babin, P.J. Zebrafish obesogenic test: A tool for screening molecules that target adiposity. J. Lipid Res. 2011, 52, 1765–1772. [Google Scholar] [CrossRef]
- Minchin, J.; Rawls, J. In vivo imaging and quantification of regional adiposity in zebrafish. Methods Cell Biol. 2016, 138, 3–27. [Google Scholar] [CrossRef]
- Flynn, E.; Trent, C.M.; Rawls, J.F. Ontogeny and nutritional control of adipogenesis in zebrafish (Danio rerio). J. Lipid Res. 2009, 50, 1641–1652. [Google Scholar] [CrossRef]
- Minchin, J.E.N.; Rawls, J.F. A classification system for zebrafish adipose tissues. Dis. Model. Mech. 2017, 10, 797–809. [Google Scholar] [CrossRef]
- Oka, T.; Nishimura, Y.; Zang, L.; Hirano, M.; Shimada, Y.; Wang, Z.; Umemoto, N.; Kuroyanagi, J.; Nishimura, N.; Tanaka, T. Diet-induced obesity in zebrafish shares common pathophysiological pathways with mammalian obesity. BMC Physiol. 2010, 10, 21. [Google Scholar] [CrossRef]
- Broeder, M.J.D.; Moester, M.J.B.; Kamstra, J.H.; Cenijn, P.H.; Davidoiu, V.; Kamminga, L.M.; Ariese, F.; De Boer, J.F.; Legler, J. Altered Adipogenesis in Zebrafish Larvae Following High Fat Diet and Chemical Exposure Is Visualised by Stimulated Raman Scattering Microscopy. Int. J. Mol. Sci. 2017, 18, 894. [Google Scholar] [CrossRef] [PubMed]
- Landgraf, K.; Schuster, S.; Meusel, A.; Garten, A.; Riemer, T.; Schleinitz, D.; Kiess, W.; Körner, A. Short-term overfeeding of zebrafish with normal or high-fat diet as a model for the development of metabolically healthy versus unhealthy obesity. BMC Physiol. 2017, 17, 4. [Google Scholar] [CrossRef] [PubMed]
- Riu, A.; Mccollum, C.W.; Pinto, C.L.; Grimaldi, M.; Hillenweck, A.; Perdu, E.; Zalko, D.; Bernard, L.; Laudet, V.; Balaguer, P.; et al. Halogenated Bisphenol-A Analogs Act as Obesogens in Zebrafish Larvae (Danio rerio). Toxicol. Sci. 2014, 139, 48–58. [Google Scholar] [CrossRef] [PubMed]
- Lyssimachou, A.; Santos, J.G.; André, A.; Soares, J.; Lima, D.; Guimaraes, L.; Almeida, M.; Teixeira, C.; Castro, L.F.C.; Santos, M. The Mammalian “Obesogen” Tributyltin Targets Hepatic Triglyceride Accumulation and the Transcriptional Regulation of Lipid Metabolism in the Liver and Brain of Zebrafish. PLoS ONE 2015, 10, e0143911. [Google Scholar] [CrossRef]
- Buerger, A.N.; Schmidt, J.; Chase, A.; Paixao, C.; Patel, T.N.; Brumback, B.A.; Kane, A.S.; Martyniuk, C.J.; Bisesi, J.H. Examining the responses of the zebrafish (Danio rerio) gastrointestinal system to the suspected obesogen diethylhexyl phthalate. Environ. Pollut. 2018, 245, 1086–1094. [Google Scholar] [CrossRef]
- Ouadah-Boussouf, N.; Babin, P.J. Pharmacological evaluation of the mechanisms involved in increased adiposity in zebrafish triggered by the environmental contaminant tributyltin. Toxicol. Appl. Pharmacol. 2016, 294, 32–42. [Google Scholar] [CrossRef]
- Wang, W.; Zhang, X.; Wang, Z.; Qin, J.; Wang, W.; Tian, H.; Ru, S. Bisphenol S induces obesogenic effects through deregulating lipid metabolism in zebrafish (Danio rerio) larvae. Chemosphere 2018, 199, 286–296. [Google Scholar] [CrossRef]
- Green, A.J.; Hoyo, C.; Mattingly, C.J.; Luo, Y.; Tzeng, J.-Y.; Murphy, S.K.; Buchwalter, D.B.; Planchart, A. Cadmium exposure increases the risk of juvenile obesity: A human and zebrafish comparative study. Int. J. Obes. 2018, 42, 1285–1295. [Google Scholar] [CrossRef]
- Heindel, J.J.; Vom Saal, F.S.; Blumberg, B.; Bovolin, P.; Calamandrei, G.; Ceresini, G.; Cohn, B.A.; Fabbri, E.; Gioiosa, L.; Kassotis, C.; et al. Parma consensus statement on metabolic disruptors. Environ. Health 2015, 14, 54. [Google Scholar] [CrossRef]
- Janesick, A.; Blumberg, B. Obesogens: An emerging threat to public health. Am. J. Obstet. Gynecol. 2016, 214, 559–565. [Google Scholar] [CrossRef]
- Hales, C.M.; Carroll, M.D.; Fryar, C.D.; Ogden, C.L. Prevalence of Obesity among Adults and Youth: United States, 2015–2016; National Center for Health Statistics: Hyattsville, MD, USA, 2017.
- Skinner, A.C.; Ravanbakht, S.N.; Skelton, J.A.; Perrin, E.M.; Armstrong, S.C. Prevalence of Obesity and Severe Obesity in US Children, 1999–2016. Pediatrics 2018, 141, e20173459. [Google Scholar] [CrossRef] [PubMed]
- Hales, C.M.; Carroll, M.D.; Fryar, C.D.; Ogden, C.L. Prevalence of Obesity and Severe Obesity among Adults: United States, 2017–2018; CDC National Center for Health Statistics: Hyattsville, MD, USA, 2020; pp. 1–8.
- Biener, A.; Cawley, J.; Meyerhoefer, C. The Impact of Obesity on Medical Care Costs and Labor Market Outcomes in the US. Clin. Chem. 2018, 64, 108–117. [Google Scholar] [CrossRef] [PubMed]
- Legler, J.; Fletcher, T.; Govarts, E.; Porta, M.; Blumberg, B.; Heindel, J.J.; Trasande, L. Obesity, Diabetes, and Associated Costs of Exposure to Endocrine-Disrupting Chemicals in the European Union. J. Clin. Endocrinol. Metab. 2015, 100, 1278–1288. [Google Scholar] [CrossRef] [PubMed]
- Kassotis, C.; Masse, L.; Kim, S.; Schlezinger, J.; Webster, T.F.; Stapleton, H.M. Characterization of Adipogenic Chemicals in Three Different Cell Culture Systems: Implications for Reproducibility Based on Cell Source and Handling. Sci. Rep. 2017, 7, srep42104. [Google Scholar] [CrossRef] [PubMed]
- Kassotis, C.D.; Hoffman, K.; Stapleton, H.M. Characterization of Adipogenic Activity of Semi-volatile Indoor Contaminants and House Dust. Environ. Sci. Technol. 2017, 51, 8735–8745. [Google Scholar] [CrossRef] [PubMed]
- Renquist, B.J.; Zhang, C.; Williams, S.Y.; Cone, R.D. Development of an Assay for High-Throughput Energy Expenditure Monitoring in the Zebrafish. Zebrafish 2013, 10, 343–352. [Google Scholar] [CrossRef]
- Reid, R.M.; D’Aquila, A.L.; Biga, P.R. The validation of a sensitive, non-toxic in vivo metabolic assay applicable across zebrafish life stages. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2018, 208, 29–37. [Google Scholar] [CrossRef]
- Imrie, D.; Sadler, K.C. White adipose tissue development in zebrafish is regulated by both developmental time and fish size. Dev. Dyn. 2010, 239, 3013–3023. [Google Scholar] [CrossRef]
- Green, H.; Kehinde, O. An established preadipose cell line and its differentiation in culture II. Factors affecting the adipose conversion. Cell 1975, 5, 19–27. [Google Scholar] [CrossRef]
- Green, H.; Meuth, M. An established pre-adipose cell line and its differentiation in culture. Cell 1974, 3, 127–133. [Google Scholar] [CrossRef]
- Zenbio, I. Subcutaneous Human Adipocytes Manual. 2015. Available online: https://www.zen-bio.com/pdf/ZBM0001.01SQAdipocyteCare.pdf (accessed on 4 November 2021).
- American Type Culture Collection (ATCC). Chemically-Induced Differentiation of ATCC CL-173 (3T3-L1) Using Single-Component Commercially-Available Reagents. Available online: http://www.atcc.org/~/media/6124AF1E4C2A47CF904435117909AC25.ashx (accessed on 17 May 2016).
- Millipore-Sigma. Human Preadipocytes (HPAd) Culture Protocol. Available online: https://www.sigmaaldrich.com/technical-documents/protocols/biology/human-preadipocytes.html (accessed on 4 November 2020).
- Jobling, S.; Sumpter, J. Detergent components in sewage effluent are weakly oestrogenic to fish: An in vitro study using rainbow trout (Oncorhynchus mykiss) hepatocytes. Aquat. Toxicol. 1993, 27, 361–372. [Google Scholar] [CrossRef]
- Newbold, R.R.; Padilla-Banks, E.; Snyder, R.J.; Jefferson, W.N. Developmental exposure to estrogenic compounds and obesity. Birth Defects Res. Part A Clin. Mol. Teratol. 2005, 73, 478–480. [Google Scholar] [CrossRef] [PubMed]
- Newbold, R.R.; Padilla-Banks, E.; Snyder, R.J.; Jefferson, W.N. Perinatal exposure to environmental estrogens and the development of obesity. Mol. Nutr. Food Res. 2007, 51, 912–917. [Google Scholar] [CrossRef] [PubMed]
- Newbold, R.R.; Padilla-Banks, E.; Jefferson, W.N. Environmental estrogens and obesity. Mol. Cell. Endocrinol. 2009, 304, 84–89. [Google Scholar] [CrossRef] [PubMed]
- Mentor, A.; Brunström, B.; Mattsson, A.; Jönsson, M. Developmental exposure to a human relevant mixture of endocrine disruptors alters metabolism and adipogenesis in zebrafish (Danio rerio). Chemosphere 2019, 238, 124584. [Google Scholar] [CrossRef]
- Sant, K.E.; Annunziato, K.; Conlin, S.; Teicher, G.; Chen, P.; Venezia, O.; Downes, G.B.; Park, Y.; Timme-Laragy, A.R. Developmental exposures to perfluorooctanesulfonic acid (PFOS) impact embryonic nutrition, pancreatic morphology, and adiposity in the zebrafish, Danio rerio. Environ. Pollut. 2021, 275, 116644. [Google Scholar] [CrossRef] [PubMed]
- Shuster, A.; Patlas, M.; Pinthus, J.H.; Mourtzakis, M. The clinical importance of visceral adiposity: A critical review of methods for visceral adipose tissue analysis. Br. J. Radiol. 2012, 85, 1–10. [Google Scholar] [CrossRef]
- Overdahl, K.E.; Sutton, R.; Sun, J.; DeStefano, N.J.; Getzinger, G.J.; Ferguson, P.L. Assessment of emerging polar organic pollutants linked to contaminant pathways within an urban estuary using non-targeted analysis. Environ. Sci. Process. Impacts 2021, 23, 429–445. [Google Scholar] [CrossRef]
- Design for the Environment. Alternatives Assessment for Nonylphenol Ethoxylates; United States Environmental Protection Agency: Washington, DC, USA, 2012. Available online: https://www.epa.gov/sites/default/files/2014-06/documents/npe_final.pdf (accessed on 16 November 2021).







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chemical | Acronym | CAS # | Manufacturer | Catalog # | Lot # | Avg MW | Density | Molecular Formula |
|---|---|---|---|---|---|---|---|---|
| Alkylphenols/ethoxylates | ||||||||
| 4-nonylphenol | NPEO(0) | 84852-15-3 | Acros Organics | 416,240,010 | A0216749 | 220.4 | 0.94 | C15H24O |
| nonylphenol ethoxylate (1–2) | NPEO(1–2) | N/A | Chem Service | S-346 | 270–35A | 294 | 1.01 | C15H24O(C2H5O)1–2 |
| nonylphenol ethoxylate (4) | NPEO(4) | N/A | Chem Service | S-347 | 348–75A | 396 | 1.02 | C15H24O(C2H5O)4 |
| nonylphenol ethoxylate (6) | NPEO(6) | N/A | Chem Service | S-348 | 195–130C | 484 | 1.04 | C15H24O(C2H5O)6 |
| nonylphenol ethoxylate (9–10) | NPEO(9–10) | N/A | Chem Service | S-350 | 267–60C | 602.8 | 1.06 | C15H24O(C2H5O)9–10 |
| nonylphenol ethoxylate (20) | NPEO(20) | N/A | Chem Service | S-354 | 127–80C | 1101 | 1.13 | C15H24O(C2H5O)20 |
| Control chemicals | ||||||||
| Tributyltin chloride | TBT | 1461-22-9 | Sigma | 442,869 | - | 325.51 | N/A | [CH3(CH2)3]3SnCl |
| Dimethylsulfoxide | DMSO | 67-68-5 | Sigma | 34,869–100 mL | - | 78.13 | 1.10 | (CH3)2SO |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kassotis, C.D.; LeFauve, M.K.; Chiang, Y.-T.T.; Knuth, M.M.; Schkoda, S.; Kullman, S.W. Nonylphenol Polyethoxylates Enhance Adipose Deposition in Developmentally Exposed Zebrafish. Toxics 2022, 10, 99. https://doi.org/10.3390/toxics10020099
Kassotis CD, LeFauve MK, Chiang Y-TT, Knuth MM, Schkoda S, Kullman SW. Nonylphenol Polyethoxylates Enhance Adipose Deposition in Developmentally Exposed Zebrafish. Toxics. 2022; 10(2):99. https://doi.org/10.3390/toxics10020099
Chicago/Turabian StyleKassotis, Christopher D., Matthew K. LeFauve, Yu-Ting Tiffany Chiang, Megan M. Knuth, Stacy Schkoda, and Seth W. Kullman. 2022. "Nonylphenol Polyethoxylates Enhance Adipose Deposition in Developmentally Exposed Zebrafish" Toxics 10, no. 2: 99. https://doi.org/10.3390/toxics10020099