
Wet Grasslands as a Green Infrastructure for Ecological Sustainability: Wader Conservation in Southern Sweden as a Case Study


Abstract
:1. Introduction
2. Materials and Methods
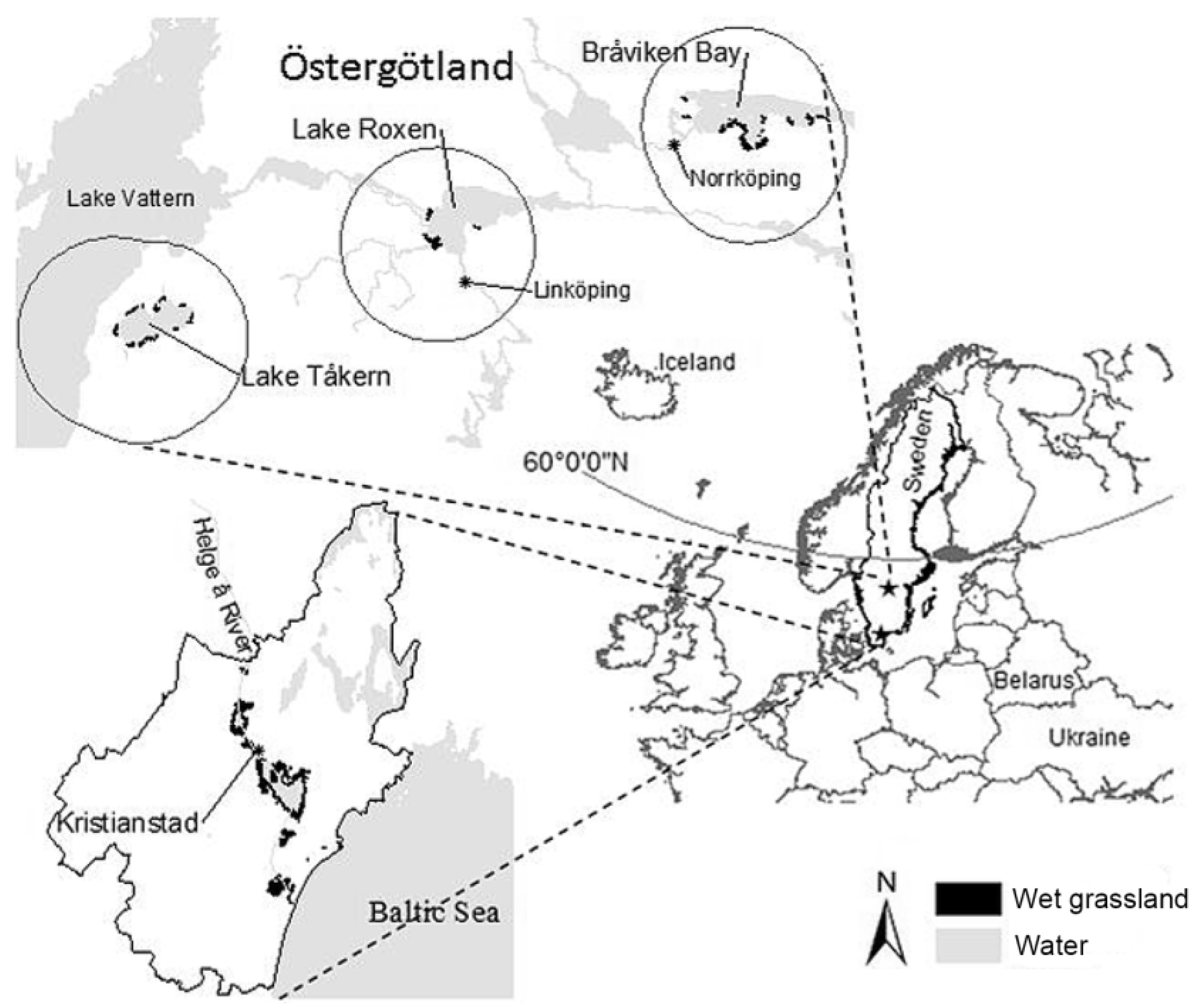
2.1. Study Areas
2.2. Field Observations
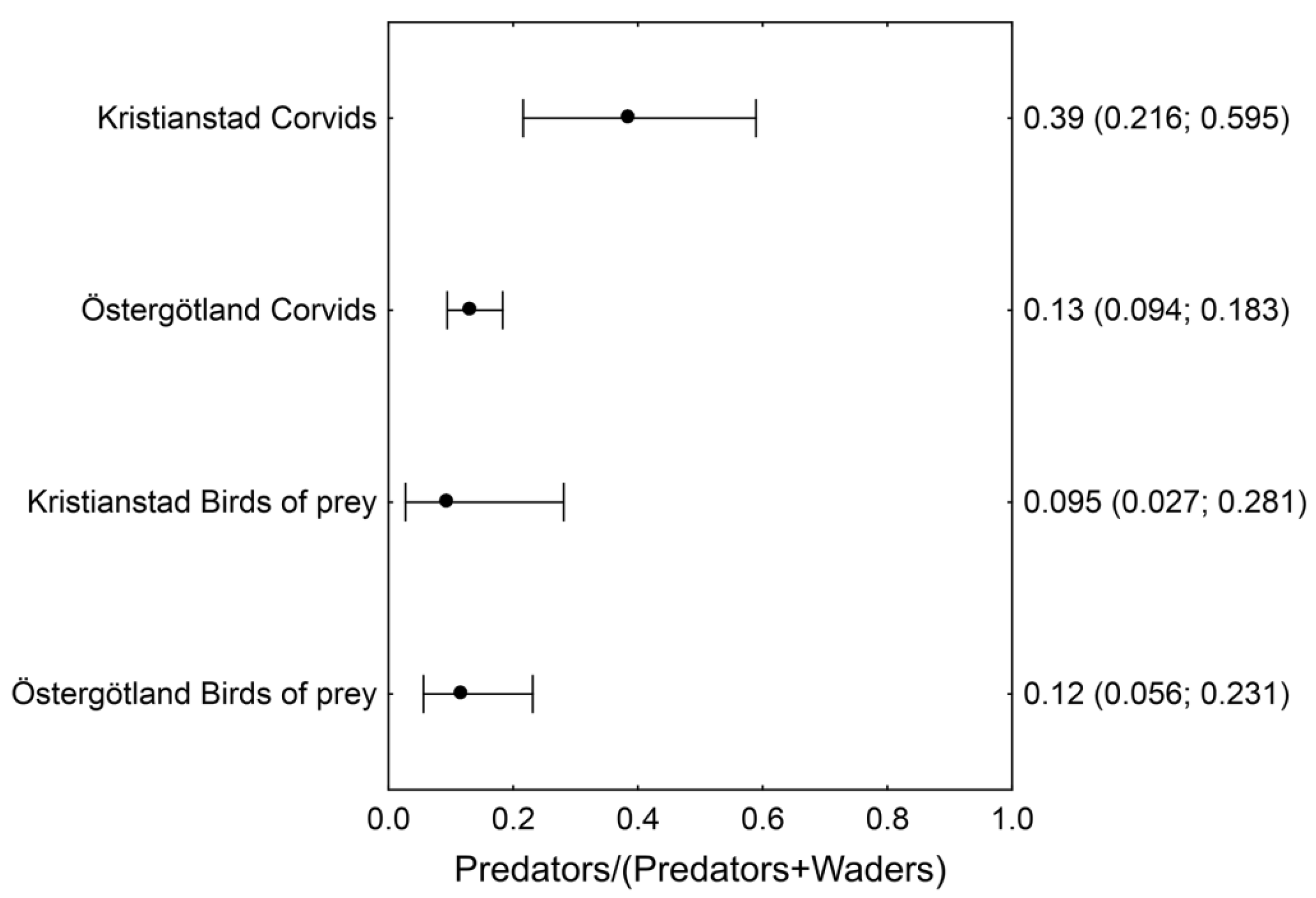
2.2.1. Relative Abundance of Avian Predators and Waders
2.2.2. Landscape Avian Predator Counts
2.2.3. Artificial Nest Predation
3. Results
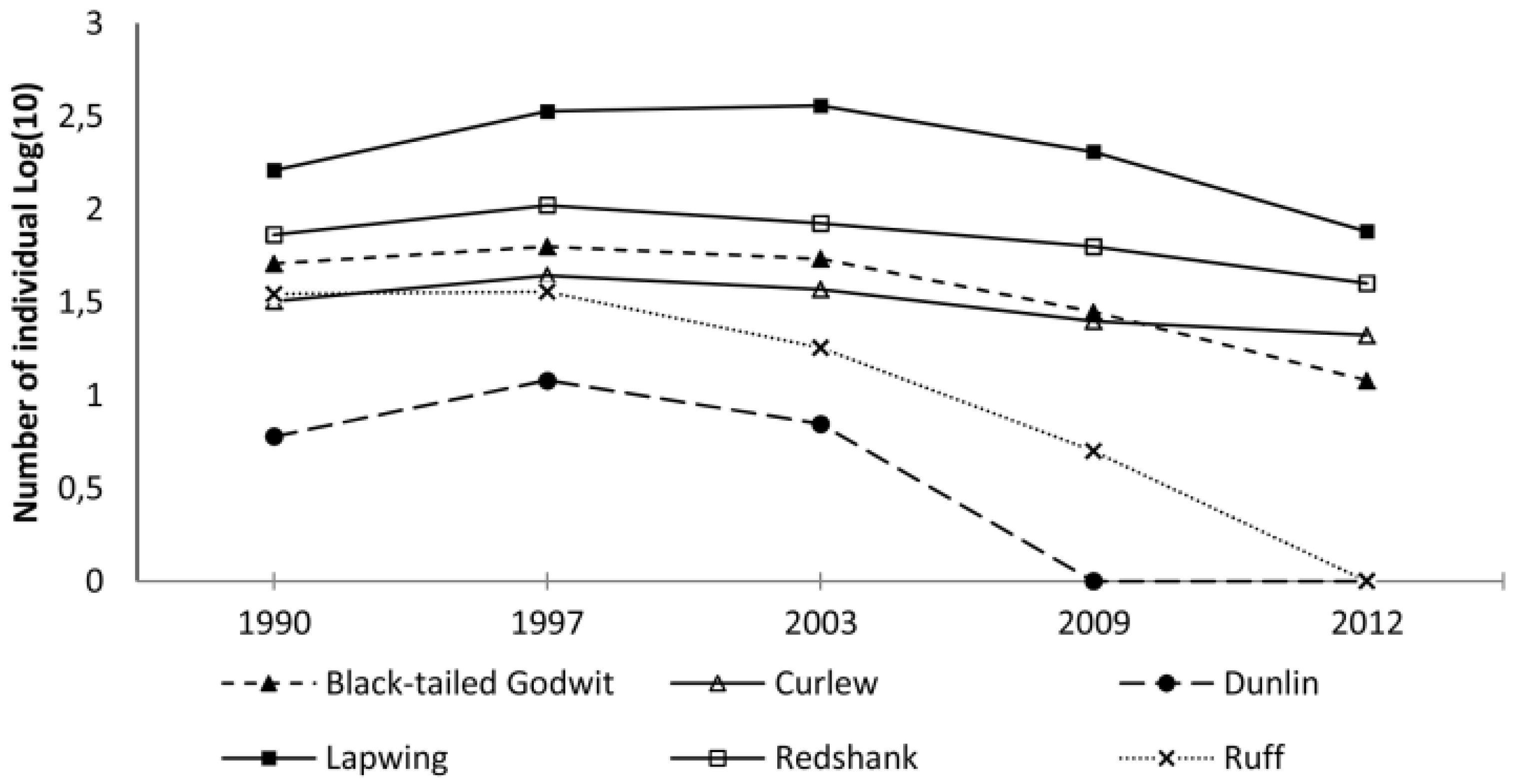
3.1. Relative Abundance of Avian Predators and Waders at the Beginning of the Breeding Season
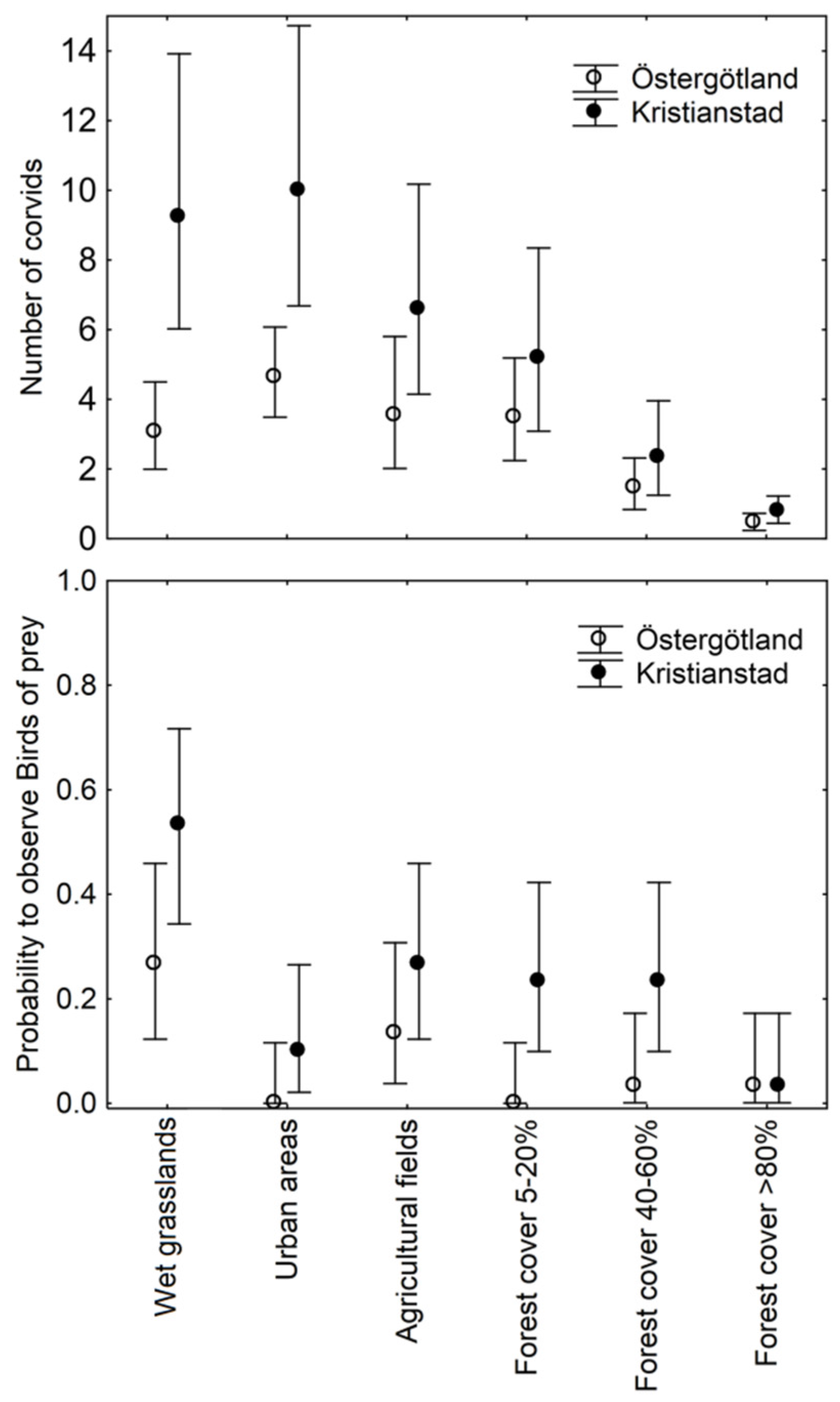
3.2. Landscape Avian Predator Counts during the Latter Part of the Breeding Season
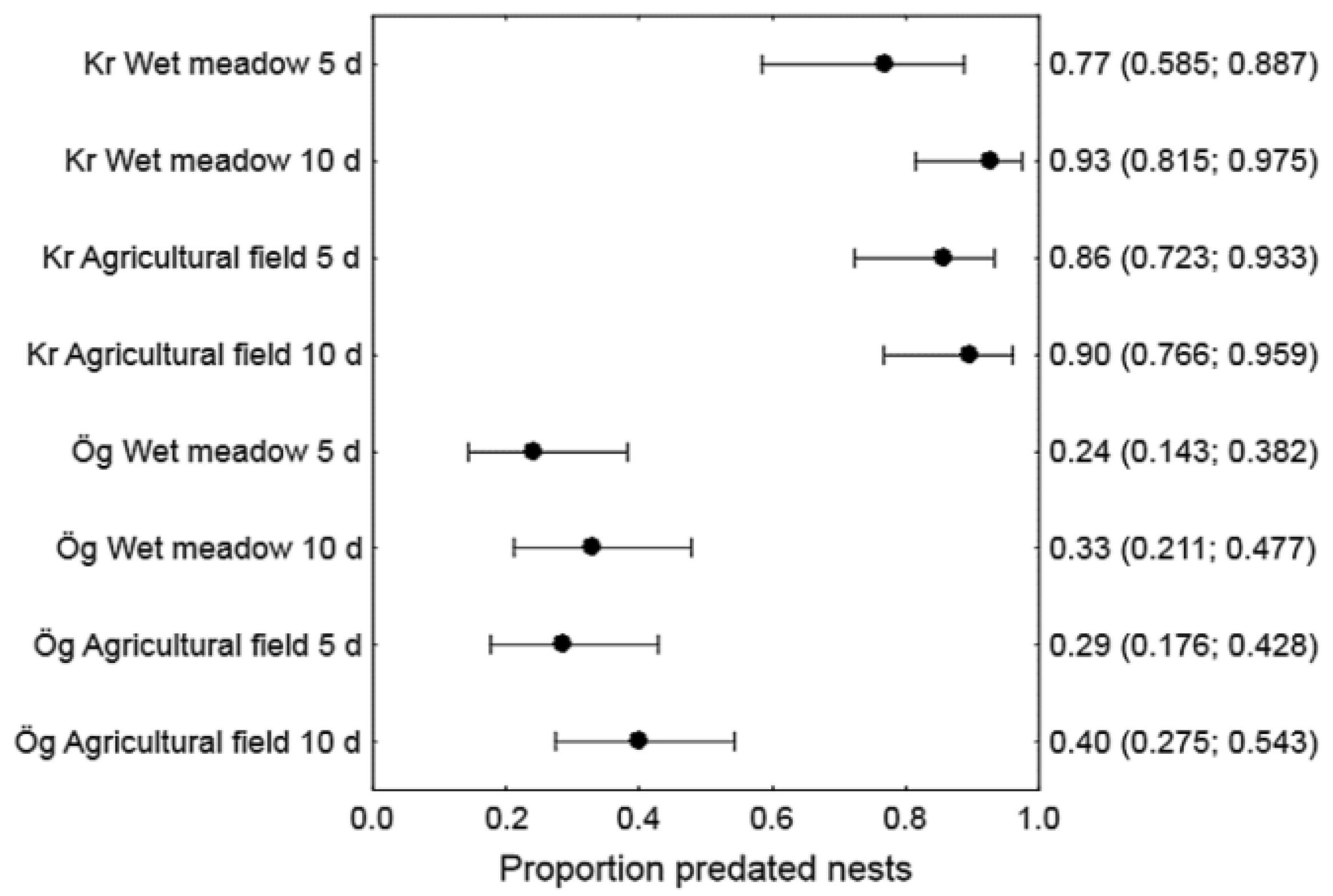
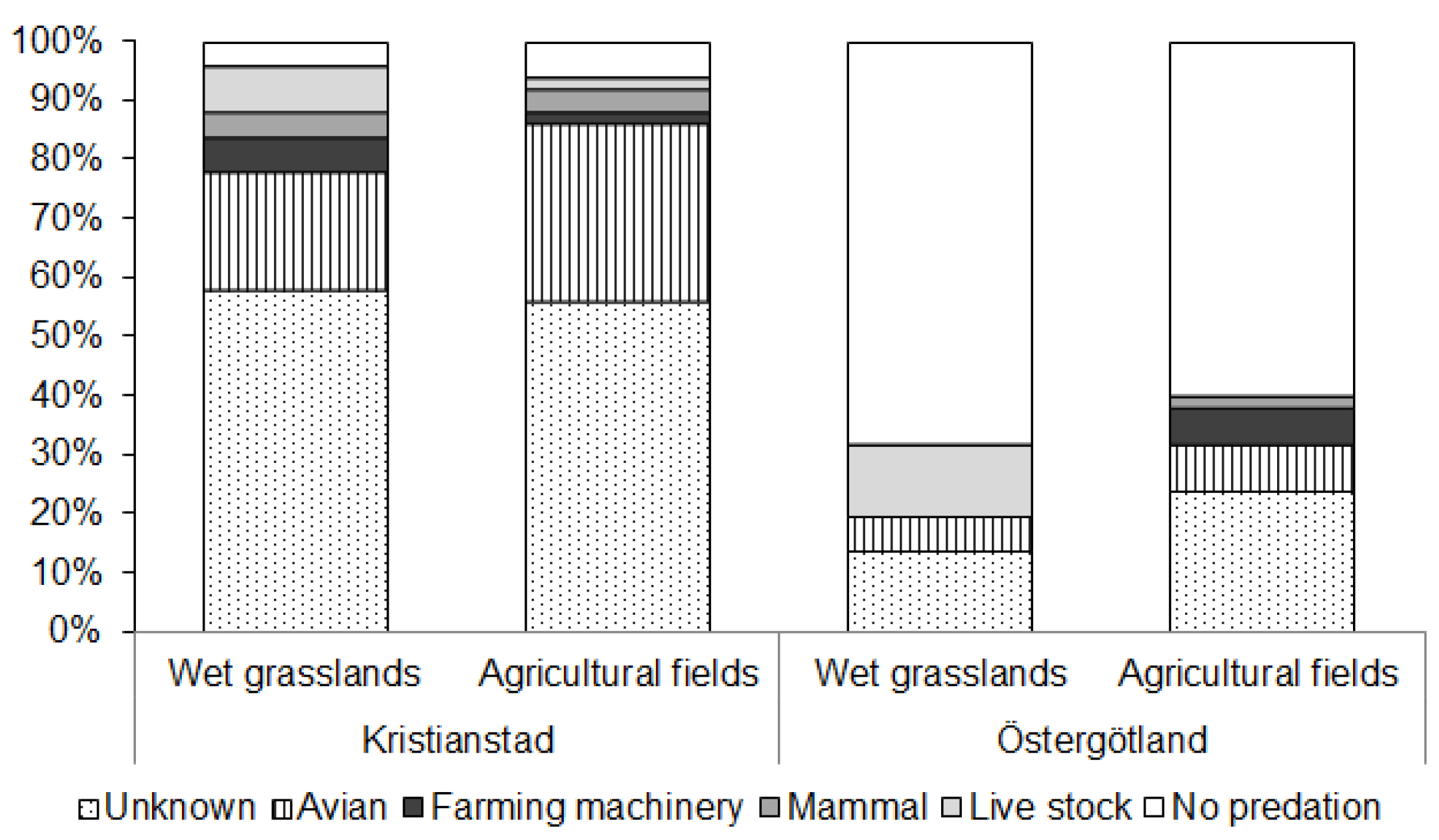
3.3. Artificial Nest Predation
4. Discussion
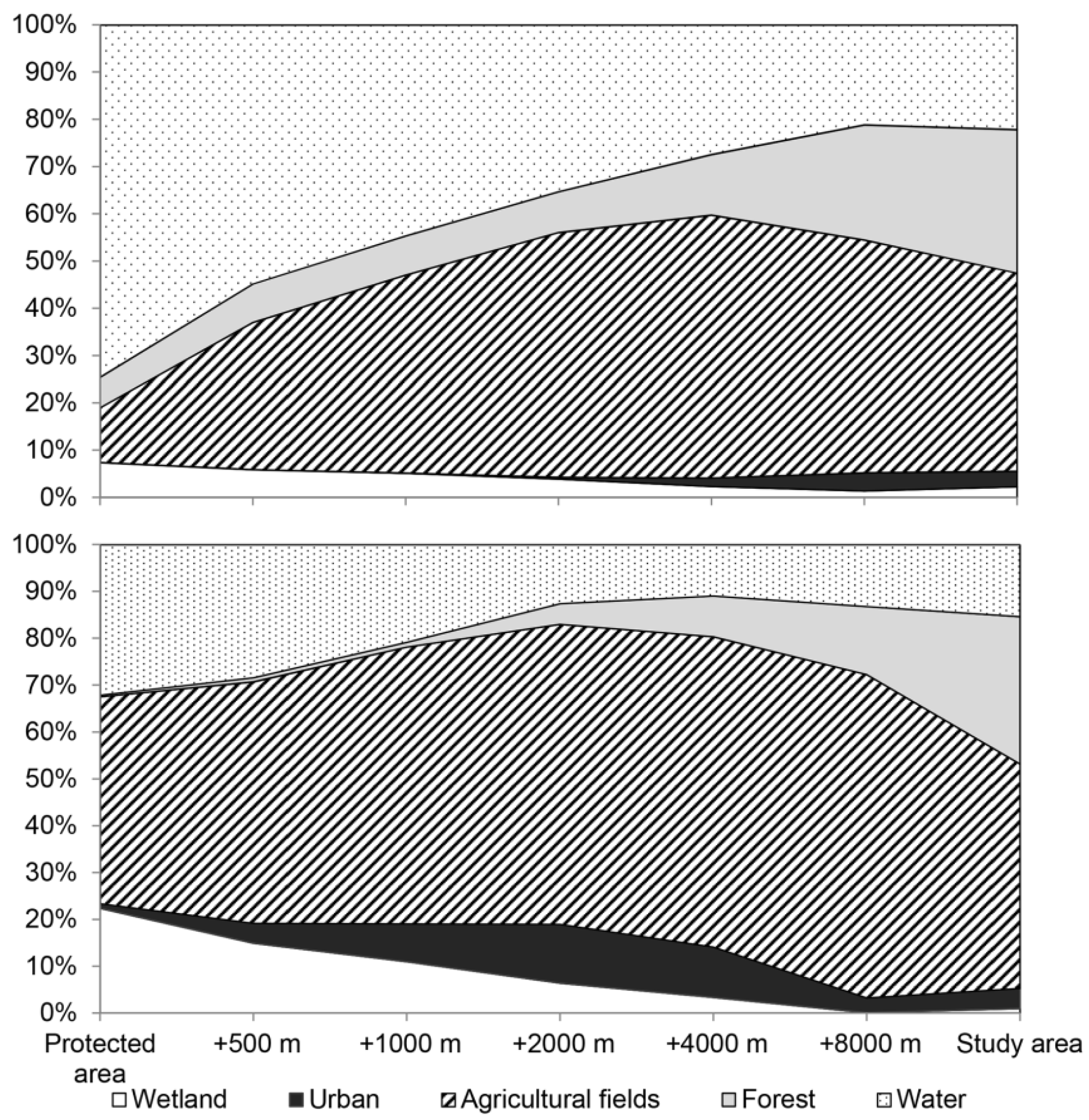
4.1. Management of Trophic Interactions and Land Covers
4.2. Towards Governance of Wet Grasslands as a Green Infrastructure
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| KVBR | Kristianstad Vattenrike Biosphere Reserve |
References
- Norgaard, R.B. Ecosystem services: From eye-opening metaphor to complexity blinder. Ecol. Econ. 2010, 69, 1219–1227. [Google Scholar] [CrossRef]
- European Commission. Green Infrastructure (GI)—Enhancing Europe’s Natural Capital. Available online: http://eur-lex.europa.eu/legal-content/EN/TXT/?uri=celex%3A52013DC0249 (accessed on 28 March 2016).
- Axelsson, R.; Angelstam, P.; Elbakidze, M.; Stryamets, N.; Johansson, K.-E. Sustainable development and sustainability: Landscape approach as a practical interpretation of principles and implementation concepts. J. Landsc. Ecol. 2011, 4, 5–30. [Google Scholar] [CrossRef]
- Elbakidze, M.; Hahn, T.; Mauerhofer, V.; Angelstam, P.; Axelsson, R. Legal framework for biosphere reserves as learning sites for sustainable development: A Comparative analysis of Ukraine and Sweden. Ambio 2013, 42, 174–187. [Google Scholar] [CrossRef] [PubMed]
- Millennium Ecosystem Assessment. Ecosystems and Human Well-Being: Synthesis; Island Press: Washington, DC, USA, 2005. [Google Scholar]
- Hahn, T.; Olsson, P.; Folke, C.; Johansson, K. Trust-building, knowledge generation and organizational innovations: The role of a bridging organization for adaptive comanagement of a wetland landscape around Kristianstad, Sweden. Hum. Ecol. 2006, 34, 573–592. [Google Scholar] [CrossRef]
- Ekberg, B.; Nilsson, L. Skånes Fåglar Idag och i Gången tid; Signum: Lund, Sweden, 1994. (In Swedish) [Google Scholar]
- Magnusson, S.-E.; Magntorn, K.; Wallsten, E.; Cronert, H.; Thelaus, M. Kristianstads Vattenrike Biosphere Reserve Nomination Form; Springer: Berlin, Germany, 2004. [Google Scholar]
- Lindström, M.; Johansson, M.; Herrmann, J.; Johnsson, O. Attitudes towards the conservation of biological diversity—A case study in Kristianstad Municipality, Sweden. J. Environ. Plan. Manag. 2006, 49, 495–513. [Google Scholar] [CrossRef]
- Schultz, L.; Folke, C.; Olsson, P. Enhancing ecosystem management through social-ecological inventories: Lessons from Kristianstads Vattenrike, Sweden. Environ. Conserv. 2007, 34, 140–152. [Google Scholar] [CrossRef]
- Olsson, P.; Folke, C.; Galaz, V.; Hahn, T.; Schultz, L. Enhancing the fit through adaptive co-management: Creating and maintaining bridging functions for matching scales in the Kristianstads Vattenrike Biosphere Reserve Sweden. Available online: http://www.ecologyandsociety.org/vol12/iss1/art28/ (accessed on 28 March 2016).
- Olsson, P.; Schultz, L.; Folke, C.; Hahn, T. Social networks for ecosystem management: A case study of Kristianstads Vattenrike, Sweden. In The Centre for Interdisciplinary Environmental Research (CTM); Stockholm University: Stockholm, Sweden, 2007. [Google Scholar]
- Hahn, T. Self-Organized Governance Networks for Ecosystem Management: Who Is Accountable? Available online: http://www.diva-portal.org/smash/record.jsf?pid=diva2%3A468735&dswid=-3101 (accessed on 28 March 2016).
- Tuvendal, M. Ecosystem Services—A Tool in Sustainable Landscape Management. Ph.D. Thesis, Stockholm University, Stockholm, Sweden, 2012. [Google Scholar]
- Walker, B.; Salt, D. Resilience Thinking: Sustaining Ecosystems and People in a Changing World; Island Press: Washington, DC, USA, 2006. [Google Scholar]
- Rauschmayer, F.; Berghöfer, A.; Omann, I.; Zikos, D. Examining processes or/and outcomes? Evaluation concepts in European governance of natural resources. Environ. Policy Gov. 2009, 19, 159–173. [Google Scholar] [CrossRef]
- Lundquist, L. Implementation Steering. An Actor-Structure Approach; Studentlitteratur: Lund, Sweden, 1987. [Google Scholar]
- Tuvendal, M.; Elmberg, J. A handshake between markets and hierarchies: Geese as an example of successful collaborative management of ecosystem services. Sustainability 2015, 7, 15937–15954. [Google Scholar] [CrossRef]
- Ottosson, U.; Ottvall, R.; Elmberg, J.; Green, M.; Gustafsson, R.; Haas, F.; Holmqvist, N.; Lindström, Å.; Nilsson, L.; Svensson, M.; et al. Fåglarna i Sverige—Antal och Förekomst; SOF: Halmstad, Swedish, 2012. [Google Scholar]
- Lindström, Å.; Green, M. Monitoring Population Changes of Birds in Sweden; Annual Report for 2012; Department of Biology, Lund University: Lund, Sweden, 2013; p. 80. [Google Scholar]
- Nilsson, S. Skandinavisk Fauna. Foglarna; Första Bandet. Available online: http://www.bokborsen.se/S-Nilsson/Skandinavisk-Fauna-Foglarna-Andra-Bandet/5515892 (accessed on 28 March 2016). (In Swedish)
- Ihse, M. Swedish agricultural landscapes—Patterns and changes during the last 50 years, studied by aerial photos. Landsc. Urban Plan. 1995, 31, 21–37. [Google Scholar] [CrossRef]
- Wretenberg, J.; Lindström, A.; Svensson, S.; Pärt, T. Linking agricultural policies to population trends of Swedish farmland birds in different agricultural regions. J. Appl. Ecol. 2007, 44, 933–941. [Google Scholar] [CrossRef]
- Ottvall, R.; Edenius, L.; Elmberg, J.; Engström, H.; Green, M.; Holmqvist, N.; Lindström, Å.; Pärt, T.; Tjernberg, M. Population trends for Swedish breeding birds. Ornis Svec. 2009, 19, 117–192. [Google Scholar]
- Cronert, H. Strandängsfåglar längs nedre Helgeån i Kristianstads Vattenrike 2009—Med jämförelser bakåt i tiden. Anser 2010, 2, 79–94. (In Swedish) [Google Scholar]
- Cronert, H. Strandängsfåglar i Vattenriket. Häckfågelkarteringarna och simultanräkningar. Spoven 2014, 1, 1–19. (In Swedish) [Google Scholar]
- Cronert, H.; Lindblad, T. Strandängsinventering—Längs nedre Helgeån i Kristianstads Vattenrike våren 2003. Anser 2004, 43, 65–78. [Google Scholar]
- Ottvall, R.; Cronert, H.; Flyckt, G.; Lindblad, T.; Olofsson, P.; Strandberg, R.; Waldemarsson, N. Rödspoven i Kristianstads Vattenrike 2008. Anser 2009, 48, 1–10. [Google Scholar]
- Bildstein, K.L.; Bancroft, G.T.; Dugan, P.J.; Gordon, D.H.; Erwin, R.M.; Nol, E.; Payne, L.X.; Senner, S.E. Approaches to the conservation of coastal wetlands in the western hemisphere. Wilson Bull. 1991, 103, 218–254. [Google Scholar]
- Brinson, M.M.; Malvárez, A. Temperate freshwater wetlands: Types, status, and threats. Environ. Conserv. 2002, 29, 115–133. [Google Scholar] [CrossRef]
- Antrop, M. The transformation of the Mediterranean landscapes: An experience of 25 years of observations. Landsc. Urban Plan. 1993, 24, 3–13. [Google Scholar] [CrossRef]
- Thorup, O. Ynglefuglene på Tipperne 1928–1992. Dansk Ornitol. Foren. Tidskr. 1998, 92, 1–192. (In Swedish) [Google Scholar]
- Beintema, A.J. Man-made polders in the Netherlands: A traditional habitat for shorebirds. Colonial Waterbirds 1986, 9, 196–202. [Google Scholar] [CrossRef]
- Gill, J.A.; Langston, R.H.W.; Alves, J.A.; Atkinson, P.W.; Bocher, P.; Cidraes Vieira, N.; Crockford, N.J.; Gélinaud, G.; Groen, N.; Gunnarsson, T.G.; et al. Contrasting trends in two Black-tailed Godwit populations: A review of causes and recommendations. Wader Study Group Bull. 2007, 114, 43–50. [Google Scholar]
- Schekkerman, H.; Teunissen, W.; Oosterveld, E. The effect of “mosaic management” on the demography of Black-tailed Godwit Limosa limosa on farmland. J. Appl. Ecol. 2008, 45, 1067–1075. [Google Scholar]
- Newton, I. The recent declines of farmland bird populations in Britain: An appraisal of causal factors and conservation actions. Ibis 2004, 146, 579–600. [Google Scholar] [CrossRef]
- Alvarez-Cobelas, M.; Cirujano, S.; Sánchez-Carrillo, S. Hydrological and botanical man-made changes in the Spanish wetland of Las Tablas de Daimiel. Biol. Conserv. 2001, 97, 89–98. [Google Scholar] [CrossRef]
- Illyés, E.; Botta-Dukát, Z.; Molnár, Z. Patch and landscape factors affecting the naturalness-based quality of three model grassland habitats in Hungary. Acta Bot. Hung. 2008, 50, 179–197. [Google Scholar] [CrossRef]
- Durant, D.; Tichit, M.; Kernéïs, E.; Fritz, H. Management of agricultural wet grasslands for breeding waders: Integrating ecological and livestock system perspectives—A review. Biodivers. Conserv. 2008, 17, 2275–2295. [Google Scholar] [CrossRef]
- Wretenberg, J.; Lindström, Å.; Svensson, S.; Thierfelder, T.; Pärt, T. Population trends of farmland birds in Sweden and England: Similar trends but different patterns of agricultural intensification. J. Appl. Ecol. 2006, 43, 1110–1120. [Google Scholar] [CrossRef]
- Catry, T.; Alves, J.A.; Andrade, J.; Costa, H.; Dias, M.P.; Fernandes, P.; Leal, A.; Lourenco, P.M.; Martins, R.C.; Moniz, F.; et al. Long-term declines of wader populations at the Tagus estuary, Portugal: A response to global or local factors? Bird Conserv. Int. 2011, 21, 438–453. [Google Scholar] [CrossRef]
- Roodbergen, M.; Werf, B.; Hötker, H. Revealing the contributions of reproduction and survival to the Europe-wide decline in meadow birds: Review and meta-analysis. J. Ornithol. 2011, 153, 53–74. [Google Scholar] [CrossRef]
- Donald, P.F.; Sanderson, F.J.; Burfield, I.J.; van Bommel, F.P.J. Further evidence of continent-wide impacts of agricultural intensification on European farmland birds, 1990–2000. Agric. Ecosyst. Environ. 2006, 116, 189–196. [Google Scholar] [CrossRef]
- Rönkä, A. Distribution, status and population trends in the Temminck’s Stint Calidris temminckii in the Finnish Bothnian Bay. Ornis Fenn. 1996, 73, 1–11. [Google Scholar]
- Raudonikis, L. Black-tailed Godwit: Bird of 1998 in Lithuania. Acta Zool. Litu. 1999, 9, 215–218. [Google Scholar] [CrossRef]
- Schekkerman, H.; Teunissen, W.; Oosterveld, E. Mortality of Black-tailed Godwit Limosa limosa and Northern Lapwing Vanellus vanellus chicks in wet grasslands: Influence of predation and agriculture. J. Ornithol. 2009, 150, 133–145. [Google Scholar] [CrossRef]
- Fahrig, L. Effects of habitat fragmentation on biodiversity. Annu. Rev. Ecol. Evol. Syst. 2003, 34, 487–515. [Google Scholar] [CrossRef]
- Thorup, O. Breeding Waders in Europe 2000; International Wader Study: Norfolk, UK, 2005; Volume 14. [Google Scholar]
- Schekkerman, H. Precocial Problems: Shorebird Chick Performance in Relation to Weather, Farming, and Predation; Alterra, Wageningen University and Research Centre: Wageningen, the Netherlands, 2008. [Google Scholar]
- Fuller, R.J. Avian responses to transitional habitats in temperate cultural landscapes: Woodland edges and young-growth. In Birds and Habitat. Relationships in Changing Landscapes; Cambridge University Press: Cambridge, UK, 2012. [Google Scholar]
- International Wader Study Group. Waders are declining worldwide. Wader Study Group Bull. 2003, 101–102, 8–12. [Google Scholar]
- Wilson, A.M.; Ausden, M.; Milsom, T.P. Changes in breeding wader populations on lowland wet grasslands in England and Wales: Causes and potential solutions. Ibis 2004, 146, 32–40. [Google Scholar] [CrossRef]
- Eglington, S.M.; Gill, J.A.; Bolton, M.; Smart, M.A.; Sutherland, W.J.; Watkinson, A.R. Restoration of wet features for breeding waders on lowland grassland. J. Appl. Ecol. 2008, 45, 305–314. [Google Scholar] [CrossRef]
- Ottvall, R.; Smith, H.G. Effects of an agri-environment scheme on wader populations of coastal meadows of southern Sweden. Agric. Ecosyst. Environ. 2006, 113, 264–271. [Google Scholar] [CrossRef]
- Smart, J.; Gill, J.A.; Sutherland, W.J.; Watkinson, A.R. Grassland-breeding waders: Identifying key habitat requirements for management. J. Appl. Ecol. 2006, 43, 454–463. [Google Scholar] [CrossRef]
- Isaksson, D.; Wallander, J.; Larsson, M. Managing predation on ground-nesting birds: The effectiveness of nest exclosures. Biol. Conserv. 2007, 136, 136–142. [Google Scholar] [CrossRef]
- Wallander, J.; Isaksson, D.; Lenberg, T. Wader nest distribution and predation in relation to man-made structures on coastal pastures. Biol. Conserv. 2006, 132, 343–350. [Google Scholar] [CrossRef]
- Teunissen, W.; Schekkerman, H.; Willems, F.; Majoor, F. Identifying predators of eggs and chicks of Lapwing Vanellus vanellus and Black-tailed Godwit Limosa limosa in the Netherlands and the importance of predation on wader reproductive output. Ibis 2008, 150, 74–85. [Google Scholar] [CrossRef]
- Macdonald, M.A.; Bolton, M. Predation on wader nests in Europe. Ibis 2008, 150, 54–73. [Google Scholar] [CrossRef]
- Koivula, K.; Rönkä, A. Habitat deterioration and efficiency of antipredator strategy in a meadow-breeding wader, Temminck’s stint (Calidris temminckii). Oecologia 1998, 116, 348–355. [Google Scholar] [CrossRef]
- Martin, T.E. Processes organizing open-nesting bird assemblages: Competition or nest predation? Evol. Ecol. 1988, 2, 37–50. [Google Scholar] [CrossRef]
- Pehlak, H.; Lõhmus, A. An artificial nest experiment indicates equal nesting success of waders in coastal meadows and mires. Ornis Fenn. 2008, 85, 66–71. [Google Scholar]
- Elmberg, J.; Gunnarsson, G. Manipulated density of adult mallards affects nest survival differently in different landscapes. Can. J. Zool. 2007, 85, 589–595. [Google Scholar] [CrossRef]
- Baillie, S.R.; Sutherland, W.J.; Freeman, S.N.; Gregory, R.D.; Paradis, E. Consequences of large-scale processes for the conservation of bird populations. J. Appl. Ecol. 2000, 37, 88–102. [Google Scholar] [CrossRef]
- Wiens, J.A. Spatial scaling in ecology. Funct. Ecol. 1989, 3, 385–397. [Google Scholar] [CrossRef]
- Angelstam, P.; Mikusiński, G.; Fridman, J. Natural forest remnants and transport infrastructure: Does history matter for biodiversity conservation planning? Ecol. Bull. 2004, 51, 149–162. [Google Scholar]
- Mikusiński, G.; Angelstam, P. Occurrence of mammals and birds with different ecological characteristics in relation to forest cover in Europe: Do macroecological data make sense? Ecol. Bull. 2004, 51, 265–275. [Google Scholar]
- Flyvbjerg, B. Case study. In The Sage Handbook of Qualative Research, 4th ed.; Denzin, N.K., Lincoln, Y.S., Eds.; Sage: Thousand Oaks, CA, USA, 2011; pp. 301–316. [Google Scholar]
- Angelstam, P.; Elbakidze, M.; Axelsson, R.; Dixelius, M.; Tornblom, J. Knowledge production and learning for sustainable landscapes: Seven steps using social-ecological systems as laboratories. Ambio 2013, 42, 116–128. [Google Scholar] [CrossRef] [PubMed]
- Bergner, A. Fåglar på Strandängar i Östergötland—Inventeringar 1996–2013; Länsstyrelsen: Östergötland, Sweden, 2013. (In Swedish) [Google Scholar]
- Oveson, P. Markhävdkartering 2008. Hävdtillståndet på Betesmarker och Slåtterängar Inom Nedre Helgeåns Våtmarksområde i Kristianstads Vattenrike; Länsstyrelsen i Skåne län: Biosfärkontoret, Sweden; Kristianstads, Sweden; Vattenrike, Sweden, 2009. (In Swedish) [Google Scholar]
- Lasiewski, R.C.; Dawson, W.R. A re-examination of the relation between standard metabolic rate and body weight in birds. Condor 1967, 69, 13–23. [Google Scholar] [CrossRef]
- Environmental Systems Research Institute. ArcGIS Desktop: Release 10.1; Environmental Systems Research Institute: Redlands, CA, USA, 2012. [Google Scholar]
- Angelstam, P. Predation on ground-nesting birds’ nests in relation to predator densities and habitat edge. Oikos 1986, 47, 365–373. [Google Scholar] [CrossRef]
- Andrén, H. Corvid density and nest predation in relation to forest fragmentation: A landscape perspective. Ecology 1992, 73, 794–804. [Google Scholar] [CrossRef]
- Andrén, H.; Angelstam, P. Elevated predation rates as an edge effect in habitat islands: Experimental evidence. Ecology 1988, 69, 544–547. [Google Scholar] [CrossRef]
- Brazaitis, G.; Kurlavičius, P. Green tree retention and bird communities on clearcuts in Lithuania. Balt. For. 2003, 9, 63–70. [Google Scholar]
- Cramp, S. Handbook of the Birds of Europe, the Middle East, and North Africa: The Birds of the Western Palearctic; Hawks to Bustards; Oxford University Press: Oxford, UK, 1980; Volume 2. [Google Scholar]
- Bomholt, P. Bestanden af Rød Glente Milvus milvus i et censusområde i det sydøstlige Jylland, 1980–1995. Dansk Ornithol. Foren. Tidsskr. 1997, 91, 53–58. (in Danish). [Google Scholar]
- Kjellén, N.; Roos, G. Population trends in Swedish raptors demonstrated by migration counts at Falsterbo, Sweden 1942–97. Bird Study 2000, 47, 195–211. [Google Scholar] [CrossRef]
- Valkama, J.; Korpimäki, E.; Arroyo, B.; Beja, P.; Bretagnolle, V.; Bro, E.; Kenward, R.; Mañosa, S.; Redpath, S.M.; Thirgood, S.; et al. Birds of prey as limiting factors of gamebird populations in Europe: A review. Biol. Rev. 2005, 80, 171–203. [Google Scholar] [CrossRef] [PubMed]
- Berg, Å. Predation on artificial, solitary and aggregated wader nests on farmland. Oecologia 1996, 107, 343–346. [Google Scholar] [CrossRef]
- Cramp, S.; Perrins, C.M.; Brooks, D.J. Handbook of the Birds of Europe, the Middle East, and North Africa: The Birds of the Western Palearctic; Crows to Finches; Oxford University Press: Oxford, UK, 1980; Volume 8. [Google Scholar]
- Svensson, L.; Grant, P.J.; Mullarney, K.; Zetterström, D.; Christie, D. Collins Bird Guide; HarperCollins: London, UK, 2009. [Google Scholar]
- Seymour, A.S.; Harris, S.; Ralston, C.; White, P.C.L. Factors influencing the nesting success of Lapwings Vanellus vanellus and behaviour of Red Fox Vulpes vulpes in Lapwing nesting sites. Bird Study 2003, 50, 39–46. [Google Scholar] [CrossRef]
- Bolton, M.; Tyler, G.; Smith, K.E.N.; Bamford, R.O.Y. The impact of predator control on lapwing Vanellus vanellus breeding success on wet grassland nature reserves. J. Appl. Ecol. 2007, 44, 534–544. [Google Scholar] [CrossRef]
- Stien, J.; Yoccoz, N.G.; Ims, R.A. Nest predation in declining populations of common eiders Somateria mollissima: An experimental evaluation of the role of hooded crows Corvus cornix. Wildl. Biol. 2010, 16, 123–134. [Google Scholar] [CrossRef]
- Baines, D.; Moss, R.; Dugan, D. Capercaillie breeding success in relation to forest habitat and predator abundance. J. Appl. Ecol. 2004, 41, 59–71. [Google Scholar] [CrossRef]
- Norrdahl, K.; Suhonen, J.; Hemminki, O.; Korpimäki, E. Predator presence may benefit: Kestrels protect curlew nests against nest predators. Oecologia 1995, 101, 105–109. [Google Scholar] [CrossRef]
- Green, R.E.; Hawell, J.; Johnson, T.H. Identification of predators of wader eggs from egg remains. Bird Study 1987, 34, 87–91. [Google Scholar] [CrossRef]
- Ottvall, R. Breeding success and adult survival of Redshank Tringa totanus on coastal meadows in SE Sweden. Ardea 2005, 93, 225–236. [Google Scholar]
- Olsen, H. Patterns of Predation on Ground Nesting Meadow Birds. Ph.D. Thesis, Royal Veterinarian and Agricultural University, Copenhagen, Denmark, 2002. [Google Scholar]
- Jackson, D.B. Experimental removal of introduced hedgehogs improves wader nest success in the Western Isles, Scotland. J. Appl. Ecol. 2001, 38, 802–812. [Google Scholar] [CrossRef]
- Smith, R.K.; Pullin, A.S.; Stewart, G.B.; Sutherland, W.J. Is nest predator exclusion an effective strategy for enhancing bird populations? Biol. Conserv. 2011, 144, 1–10. [Google Scholar] [CrossRef]
- Ivan, J.S.; Murphy, R.K. What preys on Piping Plover eggs and chicks? Wildl. Soc. Bull. 2005, 33, 113–119. [Google Scholar] [CrossRef]
- Dyrcz, A.; Witkowski, J.; Okulewicz, J. Nesting of “timid” waders in the vicinity of “bold” ones as an antipredator adaption. Ibis 1981, 123, 542–545. [Google Scholar] [CrossRef]
- Pearce-Higgins, J.W.; Grant, M.C. Relationships between bird abundance and the composition and structure of moorland vegetation. Bird Study 2006, 53, 112–125. [Google Scholar] [CrossRef]
- Wilson, J.D.; Anderson, R.; Bailey, S.; Chetcuti, J.; Cowie, N.R.; Hancock, M.H.; Quine, C.P.; Russell, N.; Stephen, L.; Thompson, D.B.A. Modelling edge effects of mature forest plantations on peatland waders informs landscape-scale conservation. J. Appl. Ecol. 2014, 51, 204–213. [Google Scholar] [CrossRef]
- Ottvall, R.; Larsson, K.; Smith, H.G. Nesting success in Redshank Tringa totanus breeding on coastal meadows and the importance of habitat features used as perches by avian predators. Bird Study 2005, 52, 289–296. [Google Scholar] [CrossRef]
- Ottvall, R. Nest survival among waders breeding on coastal meadows: The relative importance of predation and trampling damages by livestock. Ornis Svec. 2005, 15, 89–96. [Google Scholar]
- Beintema, A.J.; Muskens, G.J.D.M. Nesting success of birds breeding in Dutch agricultural grasslands. J. Appl. Ecol. 1987, 24, 743–758. [Google Scholar] [CrossRef]
- Loman, J.; Göransson, G. Egg shell dumps and Crow Corvus cornix predation on simulated birds’ nests. Oikos 1978, 30, 461–466. [Google Scholar] [CrossRef]
- Bell, B.; Merton, D. Critically endangered bird populations and their management. In Conserving Bird Biodiversity: General Principles and Their Application; Norris, K., Pain, D.J., Eds.; Cambridge University Press: Cambridge, UK, 2002. [Google Scholar]
- Melman, T.C.P.; Schotman, A.G.M.; Hunink, S.; de Snoo, G.R. Evaluation of meadow bird management, especially Black-tailed Godwit (Limosa limosa L.), in the Netherlands. J. Nat. Conserv. 2008, 16, 88–95. [Google Scholar] [CrossRef]
- Oosterveld, E.B.; Nijland, F.; Musters, C.J.M.; Snoo, G.R. Effectiveness of spatial mosaic management for grassland breeding shorebirds. J. Ornithol. 2011, 152, 161–170. [Google Scholar] [CrossRef]
- Schroeder, J.; Heckroth, M.; Clemens, T. Against the trend: Increasing numbers of breeding Northern Lapwings Vanellus vanellus and Black-tailed Godwits Limosa limosa on a German Wadden Sea island. Bird Study 2008, 55, 100–107. [Google Scholar] [CrossRef]
- Hersteinsson, P.; Angerbjörn, A.; Frafjord, K.; Kaikusalo, A. The arctic fox in fennoscandia and Iceland: Management problems. Biol. Conserv. 1989, 49, 67–81. [Google Scholar] [CrossRef]
- Gjerde, B.; Dahlgren, S. Corvid birds (Corvidae) act as definitive hosts for Sarcocystis ovalis in moose (Alces alces). Parasitol. Res. 2010, 107, 1445–1453. [Google Scholar] [CrossRef] [PubMed]
- Wilson, A.; Vickery, J.; Pendlebury, C. Agri-environment schemes as a tool for reversing declining populations of grassland waders: Mixed benefits from environmentally sensitive areas in England. Biol. Conserv. 2007, 136, 128–135. [Google Scholar] [CrossRef]
- Martin, T.E. Avian life history evolution in relation to nest sites, nest predation, and food. Ecol. Monogr. 1995, 65, 101–127. [Google Scholar] [CrossRef]
- Van Horne, B. Density as a misleading indicator of habitat quality. J. Wildl. Manag. 1983, 47, 893–901. [Google Scholar] [CrossRef]
- Brazaitis, G.; Roberge, J.-M.; Angelstam, P.; Marozas, V.; Pėtelis, K. Age-related effects of clear-cut–old forest edges on bird communities in Lithuania. Scand. J. For. Res. 2005, 20, 59–67. [Google Scholar] [CrossRef]
- Angelstam, P.; Grodzynskyi, M.; Andersson, K.; Axelsson, R.; Elbakidze, M.; Khoroshev, A.; Kruhlov, I.; Naumov, V. Measurement, collaborative learning and research for sustainable use of ecosystem services: Landscape concepts and Europe as laboratory. Ambio 2013, 42, 129–145. [Google Scholar] [CrossRef] [PubMed]
- Bergman, K.-O.; Jansson, N.; Claesson, K.; Palmer, M.W.; Milberg, P. How much and at what scale? Multiscale analyses as decision support for conservation of saproxylic oak beetles. For. Ecol. Manag. 2012, 265, 133–141. [Google Scholar] [CrossRef]
- Musa, N.; Andersson, K.; Burman, J.; Andersson, F.; Hedenström, E.; Jansson, N.; Paltto, H.; Westerberg, L.; Winde, I.; Larsson, M.C. Using sex pheromone and a multi-scale approach to predict the distribution of a rare saproxylic beetle. PLoS ONE 2013, 8, e66149. [Google Scholar] [CrossRef] [PubMed]
- Folke, C.; Hahn, T.; Olsson, P.; Norberg, J. Adaptive governance of social-ecological systems. Annu. Rev. Environ. Resour. 2005, 30, 441–473. [Google Scholar] [CrossRef]
- Paltto, H.; Nordén, B.; Götmark, F.; Franc, N. At which spatial and temporal scales does landscape context affect local density of Red Data Book and Indicator species? Biol. Conserv. 2006, 133, 442–454. [Google Scholar] [CrossRef]
- Wallin, M.; Wallin, K.; Truvé, J. Fågelfaunan på Ölands Sjömarker—Inventeringar 1988–2008; Länsstyrelsen: Kalmar, Sweden, 2009. [Google Scholar]
- Verkuil, Y.I.; Karlionova, N.; Rakhimberdiev, E.N.; Jukema, J.; Wijmenga, J.J.; Hooijmeijer, J.C.E.W.; Pinchuk, P.; Wymenga, E.; Baker, A.J.; Piersma, T. Losing a staging area: Eastward redistribution of Afro-Eurasian ruffs is associated with deteriorating fuelling conditions along the western flyway. Biol. Conserv. 2012, 149, 51–59. [Google Scholar] [CrossRef]
- Thorup, O. Status of populations and management of Dunlin Calidris alpina, Ruff Philomachus pugnax and Black-tailed Godwit Limosa limosa in Denmark. Dansk Ornitol. Foren. Tidsskr. 2004, 98, 7–20. [Google Scholar]
- Maclean, I.M.D.; Austin, G.E.; Rehfisch, M.M.; Blew, J.A.N.; Crowe, O.; Delany, S.; Devos, K.; Deceuninck, B.; Gunther, K.; Laursen, K.; et al. Climate change causes rapid changes in the distribution and site abundance of birds in winter. Glob. Chang. Biol. 2008, 14, 2489–2500. [Google Scholar] [CrossRef]
- Greenwood, J.J.D. Citizens, science and bird conservation. J. Ornithol. 2007, 148, 77–124. [Google Scholar] [CrossRef]
- Smart, M.; Canters, K.J. Ramsar participation and wise use. Landsc. Urban Plan. 1991, 20, 269–274. [Google Scholar] [CrossRef]
- The United Nations Educational, Scientific and Cultural Organization. Biosphere Reserves: The Seville Strategy & the Statutory Framework of the World Network; UNESCO: Paris, France, 1996. [Google Scholar]
- Morris, K. Wetland Connectivity: Understanding the Dispersal of Organisms that Occur in Victoria's Wetlands; Arthur Rylah Institute for Environmental Research, Department of Sustainability and Environment: Heidelberg, Germany, 2012. [Google Scholar]
- Hout, P.J.V.D. Struggle for Safety: Adaptive Responses of Wintering Waders to Their Avian Predators; University of Groningen: Groningen, the Netherlands, 2010. [Google Scholar]
- Svahn, K. Fåglar i Skåne. Available online: http://www.falsterbofagelstation.se/sales/varuspec.php?artikel=168&artkat=2&lan=sve (accessed on 28 March 2016). (In Swedish)
- Batisse, M. The biosphere reserve: A tool for environmental conservation and management. Environ. Conserv. 1982, 9, 101–111. [Google Scholar] [CrossRef]
- Murtas, D.; Davis, P. The Role of The Ecomuseo Dei Terrazzamenti E Della Vite, (Cortemilia, Italy) in Community Development. Mus. Soc. 2009, 7, 150–186. [Google Scholar]
- Davis, P. Ecomuseums: A Sense of Place; Bloomsbury Publishing: New York, NY, USA, 2011. [Google Scholar]
- Riviére, G.H. Images of the ecomuseum. Museum 1985, 37, 182–244. [Google Scholar]
- De la Rocha Mille, R. Museums without Walls: The Museology of Georges Henri Riviere. Ph.D. Thesis, City University London, London, UK, 2011. [Google Scholar]
- Tuvendal, M.; Elmqvist, T. Ecosystem services linking social and ecological systems: River brownification and the response of downstream stakeholders. Ecol. Soc. 2011, 16, 21. [Google Scholar] [CrossRef]
- Daniels, S.E.; Walker, G.B. Working through Environmental Conflict: The Collaborative Learning Approach; Praeger: London, UK, 2001. [Google Scholar]
- Sandström, U.G.; Angelstam, P.; Khakee, A. Urban comprehensive planning—Identifying barriers for the maintenance of functional habitat networks. Landsc. Urban Plan. 2006, 75, 43–57. [Google Scholar] [CrossRef]
- Elbakidze, M.; Angelstam, P.; Sandström, C.; Stryamets, N.; Crow, S.; Axelsson, R.; Stryamets, G.; Yamelynets, T. Biosphere Reserves for conservation and development in Ukraine? Legal recognition and establishment of the Roztochya initiative. Environ. Conserv. 2013, 40, 157–166. [Google Scholar] [CrossRef]
- Grahn, P.; Stigsdotter, U.A. Landscape planning and stress. Urban For. Urban Green. 2003, 2, 1–18. [Google Scholar] [CrossRef]
- Grahn, P.; Stigsdotter, U.K. The relation between perceived sensory dimensions of urban green space and stress restoration. Landsc. Urban Plan. 2010, 94, 264–275. [Google Scholar] [CrossRef]
- Stoltz, J.; Björk, J.; Grahn, P.; Mattisson, K.; Skärbäck, E. Klassificering av Utemmiljöer i Kristianstad för Hälsa och Välbefinnande. Available online: pub.epsilon.slu.se/9538/1/stoltz_et_al_130315.pdf (accessed on 28 March 2016). (In Swedish)








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study Areas | KVBR (SE) | Östergötland (SE) |
|---|---|---|
| Body mass (g/ha) | −27.4 (2.45) | 0.39 (0.918) |
| Metabolic weight (g/ha) | −4.81 (0.488) | −0.060 (0.2529) |
| Nests (No./ha) | −0.061 (0.00723) | 0.00063 (0.00368) |
| Wader Observations | Relative Abundance Field Observations (Early Season) | Landscape Field Observations (Late Season) | ||
|---|---|---|---|---|
| Kr | Ög | Kr | Ög | |
| Oystercatcher (Haematopus ostralegus) | 6 | 2 | NA | NA |
| Lapwing (Vanellus vanellus) | 215 | 131 | NA | NA |
| Wood Sandpiper (Tringa glareola) | 1 | 3 | NA | NA |
| Common Sandpiper (Actitis hypoleucos) | 2 | 0 | NA | NA |
| Redshank (Tringa totanus) | 5 | 40 | NA | NA |
| Greenshank (Tringa nebularia) | 5 | 4 | NA | NA |
| Black-tailed Godwit (Limosa limosa) | 2 | 9 | NA | NA |
| Curlew (Numenius arquata) | 27 | 3 | NA | NA |
| Snipe (Gallinago gallinago) | 38 | 12 | NA | NA |
| Ruff (Phylomachus pugnax) | 0 | 9 | NA | NA |
| Ringed Plover (Charadrius hiaticula) | 0 | 3 | NA | NA |
| Total | 301 | 216 | NA | NA |
| Corvid Observations | ||||
| Magpie (Pica pica) | 16 | 4 | 76 | 122 |
| Jay (Garrulus glandarius) | 0 | 0 | 8 | 9 |
| Jackdaw (Corvus monedula) | 101 | 8 | 572 | 908 |
| Rook (Corvus frugilegus) | 46 | 0 | 0 | 435 |
| Hooded Crow (Corvus cornix) | 338 | 18 | 149 | 377 |
| Raven (Corvus corax) | 24 | 1 | 16 | 23 |
| Total | 525 | 31 | 821 | 1874 |
| Birds of Prey Observations | ||||
| White-tailed Eagle (Haliaetus albicilla) | 1 | 3 | 0 | 1 |
| Osprey (Pandion haliaetus) | 6 | 0 | 1 | 0 |
| Red Kite (Milvus milvus) | 13 | 0 | 33 | 0 |
| Marsh Harrier (Circus aeruginosus) | 12 | 11 | 14 | 10 |
| Buzzard (Buteo buteo) | 19 | 0 | 20 | 5 |
| Kestrel (Falco tinnunculus) | 0 | 0 | 0 | 1 |
| Hobby (Falco subbuteo) | 0 | 0 | 2 | 1 |
| Peregrine Falcon (Falco peregrinus) | 1 | 0 | 0 | 0 |
| Total | 52 | 14 | 70 | 18 |
| Species | Trends [24] | Status [78,83,84] | Waders in Diet [78,83,84] | ||
|---|---|---|---|---|---|
| Corvids | 1996–2006 | 1977–2006 | Kr | Ög | |
| Magpie (Pica pica) | ↔ | ↔ | R | R | E, Y |
| Jay (Garrulus glandarius) | ↑ | ↓ | R | R | E, Y |
| Jackdaw (Corvus monedula) | ↑ | ↑ | R | R | E, Y |
| Rook (Corvus frugilegus) | ↑+ | ↔ | R | R | A, E, Y |
| Hooded Crow (Corvus cornix) | ↓ | ↓ | R | R | A, E, Y |
| Raven (Corvus corax) | ↑+ | ↔ | R | R | A, E, Y |
| Birds of Prey | |||||
| White-tailed Eagle (Haliaetus albicilla) | ↑+ | ↑+ | R | B | A, E, Y |
| Osprey (Pandion haliaetus) | ↔ | ↑+ | B | B | Fish |
| Golden Eagle (Aquila chrysaetos) | ↑ | ↑ | R | R | A, Y |
| Red Kite (Milvus milvus) | ↑+ | ↑+ | R | M | A, E, Y |
| Marsh Harrier (Circus aeruginosus) | ↔ | ↑+ | B | B | A, E, Y |
| Hen Harrier (Circus cyaneus) | ↔ | ↓ | M | B | A, E, Y |
| Montagu’s Harrier (Circus pygargus) | ↓ | ↓ | B | B | A, E, Y |
| Rough-legged Buzzard (Buteo lagopus) | ↔ | ↓ | M | M | A, Y |
| Buzzard (Buteo buteo) | ↑ | ↔ | B | B | A, Y |
| Honey Buzzard (Pernis apivorus) | ↔ | ↓− | B | B | Insects |
| Sparrowhawk (Accipiter nisus) | ↑ | ↑ | R | R | A, Y |
| Goshawk (Accipiter gentilis) | ↔ | ↔ | R | R | A, Y |
| Kestrel (Falco tinnunculus) | ↑+ | ↑+ | B | B | A, Y |
| Hobby (Falco subbuteo) | ↑ | ↔ | B | B | A, Y |
| Peregrine Falcon (Falco peregrinus) | ↑+ | ↑+ | R | R | A, Y |
| Merlin (Falco columbarius) | ↔ | ↔ | B | M | A, Y |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Manton, M.; Angelstam, P.; Milberg, P.; Elbakidze, M. Wet Grasslands as a Green Infrastructure for Ecological Sustainability: Wader Conservation in Southern Sweden as a Case Study. Sustainability 2016, 8, 340. https://doi.org/10.3390/su8040340
Manton M, Angelstam P, Milberg P, Elbakidze M. Wet Grasslands as a Green Infrastructure for Ecological Sustainability: Wader Conservation in Southern Sweden as a Case Study. Sustainability. 2016; 8(4):340. https://doi.org/10.3390/su8040340
Chicago/Turabian StyleManton, Michael, Per Angelstam, Per Milberg, and Marine Elbakidze. 2016. "Wet Grasslands as a Green Infrastructure for Ecological Sustainability: Wader Conservation in Southern Sweden as a Case Study" Sustainability 8, no. 4: 340. https://doi.org/10.3390/su8040340