
Abandoned Mine Lands Reclamation by Plant Remediation Technologies
by
 , , ,
, , ,
Jesús D. Peco
1,2,* ,
,
Pablo Higueras
2,
Juan A. Campos
1,
José M. Esbrí
2,
Marta M. Moreno
1,
Fabienne Battaglia-Brunet
3 and
Luisa M. Sandalio
4,* 1
Escuela Técnica Superior de Ingenieros Agrónomos, Universidad de Castilla-La Mancha (UCLM), 13071 Ciudad Real, Spain
2
Instituto de Geología Aplicada, Universidad de Castilla-La Mancha (UCLM), 13400 Ciudad Real, Spain
3
Bureau de Recherches Géologiques et Minières (BRGM), 3 Avenue Claude Guillemin, 45060 Orléans, France
4
Departamento de Bioquímica, Biología Celular y Molecular de Plantas, Estación Experimental del Zaidín, (CSIC), 18008 Granada, Spain
*
Authors to whom correspondence should be addressed.
Sustainability 2021, 13(12), 6555; https://doi.org/10.3390/su13126555
Submission received: 13 April 2021
/
Revised: 4 June 2021
/
Accepted: 6 June 2021
/
Published: 8 June 2021
(This article belongs to the Special Issue Chemical Pollution, Prevention, and Environmental Sustainability)
Abstract
:Abandoned mine lands (AMLs), which are considered some of the most dangerous anthropogenic activities in the world, are a source of hazards relating to potentially toxic elements (PTEs). Traditional reclamation techniques, which are expensive, time-consuming and not well accepted by the general public, cannot be used on a large scale. However, plant-based techniques have gained acceptance as an environmentally friendly alternative over the last 20 years. Plants can be used in AMLs for PTE phytoextraction, phytostabilization, and phytovolatilization. We reviewed these phytoremediation techniques, paying particular attention to the selection of appropriate plants in each case. In order to assess the suitability of plants for phytoremediation purposes, the accumulation capacity and tolerance mechanisms of PTEs was described. We also compiled a collection of interesting actual examples of AML phytoremediation. On-site studies have shown positive results in terms of soil quality improvement, reduced PTE bioavailability, and increased biodiversity. However, phytoremediation strategies need to better characterize potential plant candidates in order to improve PTE extraction and to reduce the negative impact on AMLs.
1. Introduction
In recent decades, many countries have realized that abandoned metal mining operations greatly contribute to environmental degradation [1]. Metal mining radically transforms the natural environment, causing modifications to the land, as well as increased accumulation of potentially toxic elements (PTEs) in the ecosystem, which seriously undermines soil health and the viability of all living organisms [2]. The principal problem is caused by mining activity carried out in the past using inefficient technology in the absence of environmental protection regulations, leading to the accumulation of mining waste which is now a source of PTEs [3]. Regarding to their role in biological systems, PTEs can be divided into essential and non-essential categories. Despite being required by living organisms for physiological and biochemical functions, at high concentrations, essential PTEs, such as copper (Cu), nickel (Ni), iron (Fe), and zinc (Zn), can have a toxic environmental impact. On the other hand, living organisms do not require non-essential PTEs such as lead (Pb), mercury (Hg), cadmium (Cd), arsenic (As), chromium (Cr) and antimony (Sb) [4]. Large concentrations of PTEs present in abandoned mine waste can migrate to nearby lands and cause severe agricultural soil and water pollution and undermine food safety and human health [5]. The 2030 Agenda for Sustainable Development, which includes 17 Sustainable Development Goals (SDGs), also refers to this problem. SDG target 15.3, in particular, aims, by 2030, to combat desertification, restore degraded land and soil, including land affected by desertification, drought and floods, and to strive to achieve a land degradation-neutral world [6]. The achievement of these objectives in the near future would also necessitate the identification, implementation, and optimization of AML reclamation strategies.
Over the last ten years, the following remediation strategies have been developed to reclaim abandoned mine lands (AMLs): (I) surface stabilization to prevent eolian and water erosion; (II) prevention of toxic pollutant dispersion to the surrounding environment; and (III) restoration of the degraded natural landscape [7]. AMLs can be treated in situ using the following physico-chemical techniques: soil isolation and containment, solidification and stabilization, vitrification, soil flushing, and electrokinetic reclamation [8]. However, these methods, which are expensive, time-consuming and unacceptable to the general public, cannot be applied on a large scale [9]. On the other hand, many organic and inorganic materials, which can be used as in-situ alternatives to expensive and disruptive physico-chemical techniques, can potentially be used as soil amendments [10]. These organic amendments include wood ash, manure, sawdust, compost, sewage sludge, biosolids, and wood chips. Commonly used inorganic amendments include phosphate-derived products, liming materials, alkaline sludge, and sugar foam [11]. However, despite their productive results, these techniques do not eliminate PTEs from the soil.
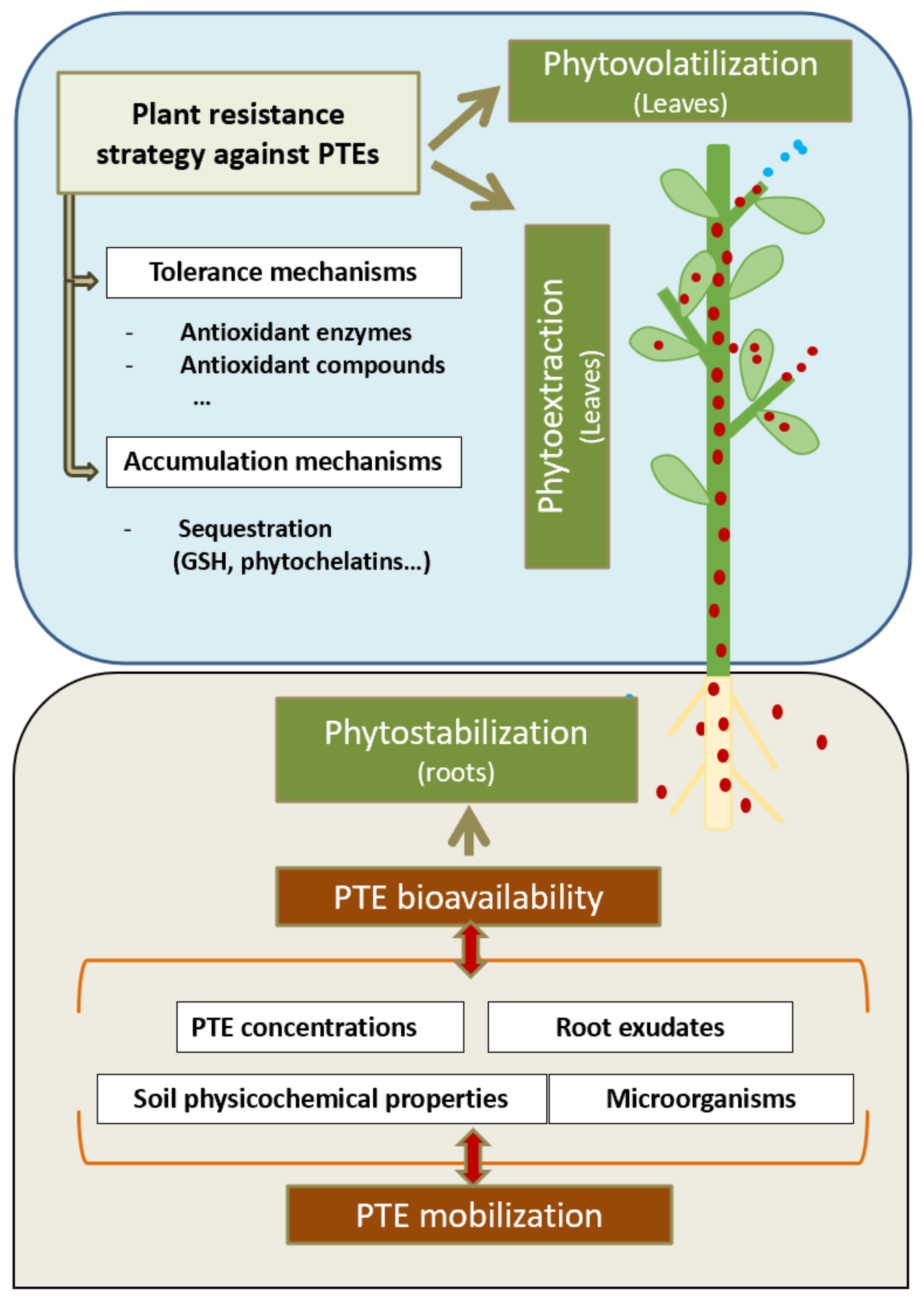
Plant-based techniques can be used to restore degraded soil in a cost-effective and environmentally friendly manner compared with physico-chemical processes [12]. While physico-chemical techniques irreversibly alter soil properties, phytoremediation generally improves the physical, chemical, and biological quality of mining lands [13]. In addition, according to Wan et al. [14], the total cost of a two-year project devoted to the phytoremediation of soil contaminated by As, Cd, and Pb was estimated to be US$ 75,375.2/hm2 or US$ 37.7/m3, which is significantly lower than that for most other techniques studied. Phytoremediation is defined as the use of living plant species to clean up soil, water and air that have been polluted with hazardous contaminants [15]. The following phytoremediation strategies are principally used to restore AMLs: in situ phytostabilization as well as phytoextraction and phytovolatilization clean-up techniques (Figure 1) [16]. These strategies involve selecting suitable plant species that can be used to decontaminate polluted soils. A knowledge of the mechanisms involved in natural plant tolerance to PTEs is therefore crucial in order to design new phytoremediation strategies [17,18,19]. The accumulation of and tolerance to PTEs involve many different genes, whose selection and transfer to candidate plants constitute a potential genetic engineering strategy to obtain new organisms with characteristics more suited to environmental remediation [20,21]. It is important to note that, apart from the need to remove human-made structures and PTEs in AML reclamation projects, it is also necessary to restore the biodiversity [3]. Thus, simultaneously or following phytoremediation, revegetation with native flora suited to the specific ecosystem is required in order to return AMLs to their pre-mining state. However, for natural colonization by native plant species to occur in most AMLs, a prior reclamation process is essential [6].
In recent years, although considerable progress has been made in the application of phytoremediation techniques, significant additional research needs to be carried out. In this review, we discussed the current state of research into AML phytoremediation techniques, analyzed some actual examples, and determined what areas require further research in the future. Particular attention should be paid to the search for appropriate wild plants for use in phytoremediation practices.
2. Occurrence of AMLs and Their Ecological Impact Worldwide
Throughout history, mining activity has produced an enormous number of impacted sites, many of which have undergone remediation, restoration, and reclamation, with differing levels of success, aimed at reducing its environmental impact. However, too often, ancient mining sites, such as those in Italy, Spain, and Latin America, were worked intensely and abandoned without any restorative measures being carried out. More recently, during the first half of the 20th century, very large French and British mining operations such as the Sao Domingos mine in Portugal [22], the Tharsis mine in Spain [23], and the Salafossa mine in Italy [24] left a legacy of abandonment. However, a comprehensive list of mining activities around the world, with different degrees of disruption, is too long for inclusion in this review. Most of these sites contain wastes accumulated in tailing dumps, which are major sources of pollution for the surrounding environment [25] though aerial dispersion, percolation, water leakage, and acid mine drainage [26].
AML abandonment has had a diverse range of consequences for the local and regional environment, some of which have had catastrophic effects, particularly in the form of waste accumulation, direct mine residue dumps, and processed waste dams, which have passed through various mineral processing stages, including froth flotation, calcination, and fusion. Each stage produces modifications in the characteristics of the original accumulated materials. Some of these changes further mobilize chemical pollutants and hence maximize the release of chemicals into the human food chain. An important related issue is the degradation of soil health [27], which includes a reduction in certain soil enzyme activities, mainly dehydrogenase, as well as changes in bacterial communities [28].
3. Plant Remediation Technologies for AML Restoration
Over the last thirty years, the use of phytoremediation for AML regeneration has been gaining in importance. Plant-based remediation, which is eco-friendly, innovative, cost-effective, practical, accepted by the general public, and carried out in-situ, can increasingly be used for PTE-contaminated soils [29]. The use of plants on AMLs enables PTEs to be extracted and immobilized without damaging the soil structure while regenerating soil fertility and quality [16]. The potential negative effects of metal-tolerant and hyperaccumulator plants are offset by their efficient uptake and accumulation of toxic elements. This, together with their adaptation to most environmental conditions, makes these plants a versatile remediation tool [30]. However, the use of plants for AML remediation has certain limitations; with their low organic content, plant nutrient levels and pH, as well as high toxic element levels, mine tailings usually lack the appropriate edaphic characteristics to establish vegetation [31]. Consequently, phytoremediation, which involves the selection of suitable plants and their establishment in AMLs, could be slower or even a long-term process as compared with physico-chemical processes [32]. In addition, implementation initially requires AML evaluation, mineralogical and physico-chemical analysis, monitoring of pH extremes, salinity and erodibility, as well as plant nutrient and microbial community analyses (Figure 1) [28,33,34]. Another important factor to be taken into account is that phytoremediation, which most often involves AML revegetation, can be considered a reconstruction and reestablishment of the original ecosystem [35].
3.1. Clean-Up Techniques
Despite being thought of as the most effective and practical plant-based PTE removal technique [30,36], phytoextraction is the most difficult to carry out given the need to find good plant candidates [16]. This technique involves selecting plants that can grow in contaminated soils and of taking up large amounts of PTEs from the soil for final accumulation in the aerial part of the plant [30]. At the end of this process, plants are harvested and incinerated after drying, which generates concentrations of ash pollutants, which need to be properly disposed of, processed to recover the metals, or simply used for landfill, with their incineration potentially producing an alternative source of energy [37]. While the removal of PTE contamination from a two-acre site to a depth of 18 inches normally produces 5000 tons of contaminated soil, the ash produced by phytoremediation techniques would only amount to between 25 and 30 tons [38]. On the other hand, PTE-contaminated biomass combustion can be hazardous due to metal dispersion through flying ashes. However, this can be counteracted using scrubbers, filters, and electrostatic precipitators which remove most PTEs and flying ash from incinerator air emissions to comply with regulatory thresholds [39]. In addition to cleaning up toxic metal-contaminated areas, at the current world prices of some PTEs, phytoextraction could be an economically viable phytomining alternative [40]. Phytomining is a phytoextraction-based technology that uses hyperaccumulator plant capacity to extract valuable elements from metal-contaminated soils; this process produces bio-ore from the harvested biomass, which facilitates metal recovery through smelting [41]. Thus, mine tailings could be used as secondary sources of valuable metals through the incineration of the harvested biomass to obtain bio-ore in much higher concentrations. Li et al. [42] reported that the 20,000 μg g−1 Ni contained in Alyssum murale biomass became 32 wt.% in ash. Recently, the establishment of the first tropical ‘metal farm’, using the ‘metal crop’ species Phyllanthus rufuschaneyi, was reported in Malaysia [43]. Metal extraction from ash could, in turn, be carried out using different processes. For example, an efficient method to recover valuable elements from the ash of hyperaccumulating Alyssum murale plants by leaching the ashes using a sulphuric acid solution has been reported [44]. Other similar processes that could be used to recover metals from incineration ashes from solid municipal waste, including magnetic separation for ferrous compounds, Eddy current separation for non-ferrous metals, and X-ray transmission sorting for light/heavy metal separation [45], could also be applied to plant ashes. The benefits and limitations of phytoextraction were elegantly reviewed by Corzo et al. [40].
The most critical step in the implementation of phytoextraction is the selection of hyperaccumulator plants capable of accumulating at least 10 g kg−1 (1%) of Mn and Zn, 1 g kg−1 (0.1%) of As, Co, Cr, Cu, Ni, Pb, Sb, Se, and Tl, and 0.1 g kg−1 (0.01%) of Cd in the aerial parts of the plant without affecting its survival [16]. Research carried out on plant populations collected from soils severely contaminated with PTEs has identified many plant species with high PTE tolerance and accumulation capacities, although this is not the only requirement. According to Alkorta et al. [36], the appropriate plants for phytoextraction should have the following features: high tolerance to PTE exposure, PTE accumulation in the aerial parts of the plant, rapid growth rates, high biomass production, a proper root system, easy crop management, genetic stability, and non-edibility for animals. Table 1 provides a summary of potentially good hyperaccumulator candidates for use in phytoextraction strategies. However, these plant populations cannot easily compensate for all the contaminated AMLs, where hyperaccumulator plants growing on this land are highly sensitive to changes in prevailing soil and climate conditions. In addition, as the tolerance and accumulation mechanisms of these plants are usually PTE-specific, the presence of other contaminants could negatively affect plant growth [7,16]. Some studies have found that a plant’s phytoextraction capacity is linearly correlated with the amount of PTEs in the soil, meaning that, after successive crops, phytoextraction ceases to be effective, with a certain amount of contaminants remaining in the soil [46].
As phytoextraction can be a very slow process as compared to other physico-chemical techniques [16], plant communities and high biomass generation capacity need to be established for one or more seasons, depending on whether herbaceous, shrub, or arboreal plants are used. The need for long periods of time to remediate polluted soils sometimes renders phytoextraction nonviable and impractical [16]. In order to accelerate the extraction process, the use of soil amendments, such as compost and shredded tree bark, with their higher seed germination rate and plant growth potential due to their enhanced physico-chemical soil properties and organic matter supply [31], as well as the use of chelators, which increase the plant metal absorption, have been proposed [47]. For example, the use of ethylenediamine disuccinic acid (EDDS) and vermicompost in Helianthus annuus L. exposed to Cd and Pb has notably been found to increase the extraction of the metals mentioned above [48]. Similarly, marble waste, when added to the soil, increased leaf Pb accumulation 2.5-fold and that of Cd by 55% in Atriplex halimus as compared with control plants [49]. On the other hand, despite the reported induced phytoextraction potential of ethylenediaminetetraacetic acid (EDTA) and diethylenetriaminepentaacetic acid (DTPA), these chelators exhibit poor biodegradability, which may trigger groundwater contamination [50,51]. This was demonstrated by Grčman et al. [47] in soil from a former Pb and Zn industrial smelter site. Despite its enhancement of PTE accumulation in the green parts of Brassica rapa L. var. pekinensis, they showed that the addition of EDTA also caused Pb, Zn, and Cd leaching through the soil and increased the toxic effects on test plants and soil microorganisms. Many studies have investigated the possibility of using other less toxic compounds for the environment than EDTA and DTPA. Meers et al. [52] evaluated whether the biodegradable chelator EDDS could be used to enhance phytoextractive processes. EDDS, which initially sparked an intense mobilization of PTEs, followed by a sharp decrease due to its rapid degradation over a 3.8–7.5 day period, was judged as unsuitable for phytoremedial use. Similarly, nitrilotriacetic acid (NTA) and other organic acids were found to require continuous supplementation in large quantities due to their rapid biodegradation also [53]. Although the addition of citric acid, NH4-citrate/citric acid, oxalic acid, EDDS, and NTA to Brassica juncea and Lolium perenne sharply increased PTE phytoextraction, their use was found to be impractical due to the time required to achieve acceptable levels of reduction in soil contaminant concentrations [54]. The goal of many studies to obtain compounds to facilitate absorption by the plant without any adverse effects [53,54] has yet to be achieved. Thus, improvements in phytoextraction techniques are currently based on appropriate plant selection and soil enhancement.
Phytovolatilization clean-up involves plant uptake of contaminants and their release into the atmosphere in a less toxic form [32]. This technique, which enables gaseous volatilized PTEs to be redeposited in surrounding lands, lakes, and oceans, can be precisely analyzed [15]. In particular, the most common PTEs, Hg, As, and Se, can be removed by their conversion into volatile forms [29]. Some interesting examples of plants with a phytovolatilization capability are shown in Table 2 and many more exist in the literature, some of which are described below. Pteris vittata has been shown to efficiently volatilize approximately 90% of total As uptake from contaminated soils in greenhouse experiments [55]. A bio-phytoremediation technique was tested for As remediation potential by adding Stenotrophomonas maltophilia and Agrobacterium sp. to the roots of Arundo donax. By the end of the experiment, ≈25% of As remained in the sand, ≈0.15% was accumulated in the plant, while the remaining 75% was volatilized by transpiration [56]. Another experiment with Zea mays exposed to Hg showed that plants inoculated with Serratia marcescens and Pseudomonas sp. increased mercury volatilization by 47.16% and 62.42%, respectively [57]. Although phytovolatilization could be used for effective AML remediation, prior evaluations of possible environmental risks linked to certain volatilized PTEs are required.
3.2. On-Site Stabilization Techniques
Phytostabilization, a remediation technique currently attracting increasing interest, involves the use of plant roots to limit PTE mobility and bioavailability in the soil [6]. Plants used for AML phytostabilization need to: (I) decrease the amount of water in the soil matrix, which prevents metals from leaching into aquifer systems; (II) prevent soil erosion and reduce PTE dispersion to other areas; (III) act as a barrier to reduce direct contact between animals and contaminated soils; (IV) sequester large amounts of PTEs through plant roots, thus reducing their bioavailability and leaching; and (V) to promote PTE immobilization in the rhizosphere through complexation/precipitation associated with the production of certain root exudates [16,58]. As with the other techniques described above involving PTE-tolerant plant selection, in the case of phytostabilization, the plant species selected also need a high root biomass production capacity, as well as root systems with PTE accumulation and uptake capabilities [59]. Most phytostabilization research has focused on the initial stage of selecting appropriate candidate plant species, some of which are listed in Table 3. These species accumulate large pollutant concentrations in the roots rather than in the aerial part of the plant, thus avoiding the drawbacks associated with phytoextraction, such as the final harvesting and the possible ingestion of the green parts and fruits by animals [7,36]. However, as these species have other drawbacks such as their inability to reduce the total amount of PTEs in the soil, plants with a long lifespan are preferable to ensure long-term phytostabilization [60].
In order to effectively establish appropriate plant communities to implement phytostabilization programs, the use of soil amendments has been proposed to improve nutritional status and soil structure [59]. The treatment of Pb/Zn mine tailings with Fe-rich compounds, furnace slag and red mud, combined with Miscanthus sinensis and Pteridium aquilinum, led to a sharp decrease in Pb, Cd, and Zn bioavailability following phytostabilization. Treatment with various amendments and the use of M. sinensis decreased the mobility of Cd, Pb, and Zn to 72–92%, 83–90%, and 79–88% of control, respectively, while, with P. aquilinum, the decrease in Cd, Pb, and Zn mobility was 79–96%, 77–91%, and 77–96% of control, respectively [61]. Another study found that a combination of compost, cyclonic ash, and steel shots had a beneficial effect on the plant growth and colonization of an industrial soil highly contaminated with Zn (1620 mg kg−1), Cd (60 mg kg−1), Cu (319 mg kg−1), and Pb (756 mg kg−1), suggesting that these amendments could also be used to enhance phytostabilization [62]. Similarly, the addition of manure was shown to be effective for PTE stabilization by enhancing the phytostabilization capacity of Brassica juncea [63]. In summary, soil amendments can be combined with phytostabilization to overcome physico-chemical limitations and to establish green cover.
It is important to note that many microorganisms in the rhizosphere play an important role in soil structure maintenance and plant nutrient recycling as a barrier against pathogens, as well as in the detoxification of some PTEs [64]. These microorganisms can modify soil metal speciation and PTE bioavailability and enhance phytostabilization in some cases [53]. However, the ecological effect of foreign bacteria on indigenous microbial communities and their possible impact on human health need to be taken into account. It has been reported that the addition of Pseudomonas fluorescens and Microbacterium sp. to Brassica napus stimulates plant growth and Pb accumulation [65] and that the addition of Sanguibacter sp. and Pseudomonas sp. increases Nicotiana tabacum growth, as well as Cd, Fe, and Zn uptake [66]. It is important to point out that changes in the plant rhizosphere can also enhance tolerance to certain PTEs. Thus, Quiñones et al. [67] found that Lupinus albus plants inoculated with a Bradyrhizobium canariense bacterium became more Hg-tolerant as compared with control plants. In summary, increases or modifications in the rhizosphere enhance phytoextraction, phytostabilization, and adaptation to AMLs and also increase PTE uptake and immobilization by the plant.
4. Selection of Suitable Plants for AML Restoration
Plant selection is critical for the development of an effective phytoremediation strategy. It is vital to carry out a preliminary study of the AMLs to be restored and to set clear objectives [17]. The major plant selection criteria for phytoremediation include a good level of tolerance to the PTEs present in the AMLs, a sufficiently high potential level of PTE accumulation, translocation, and uptake, a high growth rate, and biomass yield and extreme weather tolerance. Soil physico-chemical properties, the plant’s terrestrial, aquatic, or semi-aquatic habitat, as well as the characteristics and depth of roots, also need to be considered [18]. However, exotic plants, even those with good phytoremediation, rapid growth, and excellent colonization capacity, which could lead to unwanted invasions of whole ecosystems, should not be used [68]. In general, the marketing of plants, which have highly specific PTE accumulation and tolerance capacities, as well as specific climate and soil requirements, may not be economically viable [16]. Plants also have certain maintenance requirements such as the addition of fertilizers, periodic replanting and harvesting, as well as the implementation of feasibility studies [29]. In summary, for the purposes of AML remediation, it is necessary to select plants with sufficient growth capacity in the degraded areas and with specific PTE detoxification characteristics [19,69,70]. Researchers have also tried to identify natural metallophyte species that could be useful for AML reclamation. In recent years, molecular genetic techniques, which can be used to improve plant phytoremediation capacity, have been playing an increasingly important role.
4.1. Wild Plant Species
Great strides have been made in the search for and characterization of PTE hyperaccumulating and hypertolerant plants to PTEs. All these data have been collected in databases such as PHYTOREM in Canada and the metal-accumulating plants database METALS funded by the Environmental Consultancy of the University of Sheffield (ECUS) in the UK [71]. However, their use by phytoremediation researchers has been limited. Unfortunately, PTE accumulation and tolerance are influenced by many variables such as AML physico-chemical characteristics and climatic conditions, in addition to plant physiology. Thus, given the extreme difficulty of using non-native plants for remediation of specific AML areas, the first step in any phytoremediation strategy remains the selection and characterization of AML native plants.
Hyperaccumulator and hyper-tolerant plants often appear to be restricted to metalliferous soils, where their tolerance and accumulation mechanisms operate against PTEs [69,72]. The collection and spread of hyperaccumulators may therefore present challenges due to the difficulty of harvesting, separating, and cleaning natural seeds, as well as to the germination and cultivation requirements of the places where hyperaccumulators naturally grow [73]. The characteristics of hyperaccumulator plant populations often depend on whether they come from metalliferous or nonmetalliferous soils [74]. In research carried out by Poscic et al. [72], thallium tolerance and accumulation were compared in one non-metallicolous and three metallicolous populations of Tl hyperaccumulator Biscutella laevigata. Their results show that Tl hyperaccumulation and hyper-tolerance are population-specific traits of B. laevigata. In research by Ke et al. [75], a Rumex japonicus plant population from a copper mine and another from an uncontaminated site were exposed to Cu. Their results showed that metallicolous plant populations had higher tolerance to Cu and lower nutrient availability than populations from the uncontaminated site. This indicated that the tolerance and accumulation of certain PTEs are population-specific.
Once the plants to be used have been selected, a preliminary study of their PTE accumulation capacity and tolerance mechanisms is recommended (Figure 1) [17,18] in order to assess the suitability of these plants for AML phytoremediation.
4.1.1. Metal Uptake and Accumulation Mechanisms
Several mechanisms enable accumulator plants to absorb PTEs into the roots which are then translocated via various systems to the plant’s aerial parts for eventual storage [76]. PTE accumulation goes through four main stages: (1) PTE phytoavailability and uptake from the soil through the roots; (2) accumulation in the roots; (3) translocation from roots to shoots through xylem bundles; and (4) accumulation in the aerial parts of the plant through various mechanisms [33]. PTE phytoavailability depends on soil characteristics such as pH [63,77,78], salinity [79], humidity [80], soil organic matter content [63,77,79], speciation [81], inter-metal competition [78], and cation exchange capacity [77]. These characteristics enable plants to accumulate PTEs in different forms such as free ions or complexes containing organic and inorganic matter [82]. The synergism and antagonism among PTEs and other elements can also have a major impact on their absorption, translocation, and accumulation. In research carried out by Dong [83], competition between Cd and Mn absorption and translocation was observed in Lycopersicon esculentum exposed to Cd. Similarly, the addition of Ca reduced Cd absorption by soybean and wheat roots, while the addition of K only diminished Cd absorption by wheat roots [84]. On the other hand, plant roots produce chelating agents and compounds which promote changes in soil pH, as well as redox reactions. Thus, plants are able to solubilize and absorb PTEs present in the substrate [85]. Triticum aestivum and Sorghum bicolor, which produce phytosiderophores under Fe deficiency stress, facilitate the uptake of other PTEs [86]. Similarly, the use of root exudates from Belamcanda chinensis in Echinochloa crus-galli exposed to Pb, Cd, and Cu increases plant metal uptake levels [87]. An opposite pattern is observed with respect to Medicago polymorpha, whose root organic acid production reduces PTE phytoavailability [88]. The ability of this plant to produce certain compounds that facilitate PTE uptake may be of particular interest with regard to enhancing phytoremediation efficiency.
Once phytoavailable, PTEs can enter roots via apoplastic and symplastic pathways. In the apoplastic pathway, PTEs enter the cells through intracellular spaces, while in the symplastic pathway they are absorbed by the roots through protein transporters located in the membrane [30]. These transporters specialize in the translocation of nutrients within the root, although some PTEs, whose chemical characteristics are very similar to those of certain nutrients, can use the same entry channels [17]. Since PTE uptake depends on the number of membrane proteins present, PTE absorption is saturable according to Michaelis-Menten kinetics [89]. Each species contains different types and numbers of transporters in each cell, thus making the absorption of a certain element characteristic of the species [76]. Numerous studies of these transporters have been carried out, with the most important Cd transporter families being ZIP, YSL, and NRAMP [90,91,92], while those for Cu uptake are ZIP, COPT, Cu-ATPases, ATX, CCS, and YSL [93,94,95,96]. Thus, PTE absorption by plants, which is influenced by many factors, is a highly complex process. After entering the root, PTEs are translocated to the aerial parts of the plant through xylem bundles, which are involved in the transport of water and dissolved salts [97]. This energy-consuming process only permits the entry of metals through the symplastic pathway, as the Casparian strip in the endodermis closes off access during transport to the xylem [30]. Many natural chelates are synthesized in order to transport the metal to the xylem, which is then transported to the aerial part of the plant. Glutathione, phytochelatins (PCs), organic acids, histidine, and nicotinamide have been reported to facilitate this process [98,99,100,101,102].
After their translocation from the roots to the aerial part of the plant, a large number of chelator compounds play an important role in the accumulation of PTEs, which bind to thiol-rich peptides and phytochelatins (PCs) to facilitate their accumulation in specific locations in the roots and aerial parts of plants [19,69,103,104,105]. PCs are enzymatically synthesized from glutathione or its homologs through a reaction catalyzed by the PC synthase (PCS), a PTE-activated enzyme [105,106]. These peptides are made up of three amino acids with a typical (y-GluCys) n-Gly (n = 2–11) sequence. In some plants, the C-terminal glycine is replaced by serine, glutamine, glutamate, and alanine with (γ-Glu-Cys)n-β-Ala, (γ-Glu-Cys)n-Ser and (γ-Glu-Cys)n-Glu sequences [107,108,109,110,111], whose mechanisms of action have been elucidated in Nicotiana tabacum in response to Cd. After crossing the plasma membrane, Cd forms complexes with PCs in the cytosol, which are then actively transported into the vacuole [112]. This mechanism facilitates the sequestering of Cd, which is kept away from cellular machinery to prevent any toxic effects on the plant [15]. PCs are induced in response to many PTE ions such as Ag+, H2AsO3−, H2AsO4−, Cd2+, Cu2+, Ca3+, Hg2+, Ln3+, Ni2+, Pb2+, SeO42−, and Zn2+ [107].
Hyperaccumulating plants are usually characterized by limited compartmentalization of PTEs in root vacuoles, which are mainly translocated to the aerial parts of the plant. This was demonstrated by Yang et al. [113], who compared two populations of Sedum alfredii. Their results showed that the non-accumulating plant population retained 2.7 times more Zn in root vacuoles than the hyperaccumulating population. The bioaccumulation factor (BF) and translocation factor (TF) are used to evaluate the phytoextraction potential of a given species. The BF indicates a plant’s PTE accumulation efficiency in leaves as compared to PTE soil concentrations, while the TF indicates a plant’s efficiency in transporting PTEs from roots to leaves [114]. Thus, the BF, which was found to be less than one in metal excluder species, was greater than one in metal hyperaccumulator species [64]. The TF, which indicates where most of the metal is accumulated in roots and shoots, could be an accurate indicator of whether a species is a good phytoextraction or phytostabilization candidate. A TF of less than one indicates that the metal is mainly accumulated in roots, while a TF of greater than one indicates that most of the metal is accumulated in leaves [19,69,70].
4.1.2. Tolerance Mechanisms
Plant growth and development are directly dependent on optimal environmental and nutritional factors, any changes in which produce stress [115]. PTE-related stress can lead to multiple toxic effects of differing intensities depending on plant species, concentration, and phytoavailability, as well as the duration of exposure [16]. PTE toxicity triggers morphological and physiological imbalances in plants [19,48,69,70,116,117]. This is mainly due to the inhibition of most cellular processes at different metabolic levels caused by the appearance of reactive oxygen species (ROS), which produces a state of oxidative stress in the plant [19,118]. From a physiological perspective, it is important to highlight the decrease in photosynthetic pigment content and in leaf water potential and stomatal closure due to oxidative stress and reduction in photosynthesis [119]. At the biochemical level, the similarity of some PTEs to certain essential elements can result in the replacement of elements in active enzyme sites with PTEs. This results in a reduction or inhibition in enzyme activity which undermines the correct functioning of the entire system [120]. Many PTEs also affect the uptake of essential nutrients which compete for the same transporters, thus causing deficiencies in nutritional status [121].
PTE-related stress is directly correlated with an increase in the accumulation of ROS, which are a group of free radicals produced by sequential reductions in molecular oxygen (O2). The most common ROS include singlet oxygen (1O2), hydrogen peroxide (H2O2), as well as superoxide (O2−) and hydroxyl (⋅OH) radicals [122], which play important roles in the plant’s normal metabolism by maintaining a balance between their production and elimination by antioxidant systems. However, exposure to PTEs can cause ROS to exceed the threshold levels required for plant metabolism and trigger damage to proteins, lipids, and nucleic acids, which are the main cellular macromolecules [123]. Tolerance to PTEs in plants is dependent on their ability to activate certain tolerance systems that enable ROS levels to be maintained within normal limits [124]. Plants have complex enzymatic and non-enzymatic antioxidant defenses, which are their main protection against PTEs and enable them to maintain ROS levels compatible with those of regularly functioning cells [124]. The principal enzymatic antioxidative defenses are catalase (CAT), superoxide dismutase (SOD), peroxidase (POD), NADP-dependent dehydrogenases, glutathione S-transferase (GST), and the Foyer-Halliwell-Asada cycle enzymes ascorbate peroxidase (APX), monodehydroascorbate reductase (MDHAR), dehydroascorbate reductase (DHAR), and glutathione reductase (GR) [122]. SOD, which removes O2−, is one of the primary antioxidative defenses against PTE toxicity [125]. An increase in SOD activity against a large number of PTEs, such as Pb [69], Cd [126], Cu [127], Hg [128], As [129], Sb [130], Cr [131], and Zn [132], has been reported. To deal with an increase in H2O2, plants activate different enzymes, particularly CAT, POD, and the Foyer-Halliwell-Asada cycle enzymes. Many studies have shown that, by using these enzymes to increase the rate of H2O2 removal, certain plants can better tolerate PTEs such as Cd [19], Cu [70], As [129], Sb [133], Cr [131], and Hg [134]. The correct functioning of the Foyer-Halliwell-Asada cycle is promoted by an increase in the NADP-dehydrogenase activities NADP-G6PDH, NADP-IDH, and NADP-MS, which supply the reduction power required by the AsA-GSH cycle [135]. An induction of certain NADP dehydrogenases against Cd [19], Cu [70], and As has been reported [136]. GST enzymes are a diverse group of enzymes which catalyse the conjugation of electrophilic xenobiotic substrates with GSH, thus removing cytotoxic and genotoxic compounds that could damage the cells [137]. This group of enzymes plays an important role in defenses against As [129,137], Cd [19], Cu [70], Zn [138], and Cr [139]. Non-enzymatic components of the antioxidative defense system include ascorbate (AsA), glutathione (GSH), tocopherol, carotenoids, and phenolic compounds [118]. GSH, which is the most abundant low-molecular weight thiol-bearing molecule in cells, constitutes an essential component of PTE resistance due to its role as a substrate in enzymatic antioxidant systems and as a precursor of the PTE chelating molecules phytochelatins and metallothioneins [140]. More recent studies have reported that nitric oxide, (NO), which can prevent oxidative damage by enhancing antioxidant defenses, could be a key factor in tolerance to PTEs [124,141].
4.2. Genetically Modified Plants
Although plants use their natural metabolic mechanisms to take up PTEs from the soil, these mechanisms are often insufficient for the purposes of phytoremediation, which is sometimes ruled out due to the difficulty of finding plants with all the characteristics required [30]. The use of genetic engineering to modify plant characteristics has emerged as a promising solution to overcome the common drawbacks of conventional plants used in phytoremediation strategies [142]. Scientists have managed to develop genetically modified (GM) plants to overcome the limitations of natural plants, including low biomass production, slow growth, and the lack of tolerance and accumulation mechanisms [143]. However, certain safety and regulatory requirements can limit the commercial use of GM plants [29]. The main problem associated with these types of plants, which show greater survival potential than wild plants, is their possible invasiveness [144]. Consequently, phytoremediation techniques involving GM plants are still at the initial stage of seeking genes that improve the efficiency and development of new transgenic plants which are then tested under laboratory conditions. Several studies, including one carried out by Sanz-Fernández et al. [20], have assembled a “molecular toolbox” of genes for phytoremediation purposes which could be used by other scientists to obtain GM plants. Research carried out on these types of plants shows that genetic engineering could become an important remediation tool. After exposure to PTEs, Beta vulgaris L. plants, which were transformed by gamma-glutamylcysteine synthetase (GGCS)-glutathione synthetase (GS) from Streptococcus thermophilus, accumulated more Cd, Zn and Cu than wild-type plants and showed higher GSH and PC levels [145]. PpMT2, a metallothionein-encoding gene from Physcomitrella patens, was used to improve the tolerance of Arabidopsis thaliana plants to high concentrations of Cu and Cd [146]. Similarly, Arabidopsis thaliana transgenic plants, expressing the natural resistance-associated macrophage protein 6 (Nramp6) gene from Sedum alfredii, exhibited higher Cd accumulation levels as compared with wild-type plants [91]. Although GM plants have yet to be tested on natural AMLs, the ability to genetically engineer plants with ideal characteristics, unlike other remediation techniques, could be of great significance in the future.
5. Real Examples of Using Plants for AML Restoration
Although often misunderstood as simply the removal of human-created structures and waste, AML restoration involves the regeneration of all previously existing biodiversity in an area prior to the commencement of mining operations [3]. While the removal of PTEs accumulated over decades in the environment is insufficient, it is essential to study the whole range of organisms that previously inhabited the location to be returned to its initial state [147]. AML restoration projects initially involve field experiments to regenerate the essential soil fertility conditions through the removal and immobilization of PTEs and through soil restoration. Once this initial stage is completed, the next step will be the restoration of the entire ecosystem.
Given the extent of the progress made, one of the most interesting examples of using plants for AML restoration is the area affected by a mining accident in Aznalcóllar in southwestern Spain in 1998. It was caused by a dam collapse due to an excessive volume of mining waste deposits. The spill affected approximately 4286 ha of alluvial soil, which were flooded by slurry containing high PTE concentrations, including 0.27 mg L−1 of As, 0.85 mg L−1 of Cd, 3.6 mg L−1 of Pb, and 463 mg L−1 of Zn, a few hours after the accident [148]. Following the accident and clean-up operations, a large number of studies were carried out on the defensive behavior of herbaceous plants against PTEs accumulated in nearby areas for at least 20 years [149,150,151,152]. Later studies focused on testing the potential of using different plant species for phytoremediation purposes. For example, Helianthus annuus plants, which were grown in the spill-affected soil, were evaluated for their PTE removal capacity. Unfortunately, the results showed that plant growth in contaminated soils was negatively affected and that amount of PTEs absorbed was very low. Though ruled out for use in phytoextraction, some studies favored this plant species for use in soil conservation [153]. Similarly, two Brassica juncea crops containing different organic soil amendments were analyzed on the site affected by the toxic spillage. However, as in the previous case, total PTE uptake was relatively low, rendering its use in the phytoextraction of pluri-contaminated sites ill-advised [154]. Likewise, two cereal crops, barley and triticale, and two Brassica sp. crops were grown, whose absorption rates were excessively slow, which cast doubt on the viability of using these crops for decontamination [155]. On the other hand, despite their slow PTE uptake rates, Lupinus albus plants showed an increase in acid soil pH, a decrease in soluble As and Cd fractions, as well as high As and Cd accumulation levels in roots. According to the results obtained, it would be possible to extract approximately 0.54 g Cd ha−1 year−1 and 5.26 g As ha−1 year−1 from the contaminated soil by harvesting and removing the lupin shoots. Given these positive results, this plant species was recommended for use in the phytostabilization and revegetation of the affected area [156]. Large samplings were carried out to determine which plants could be used in phytoremediation programs. It is important to highlight the research by Del Rio et al. [149], who carried out periodic field surveys to identify spontaneously growing metal-tolerant species in the area affected by the Aználcollar accident. Their results show that, of the 99 plants studied, Anchusa azurea, Beta vulgaris, Chamaemelum fuscatum, Convolvulus arvensis, Cynodon dactylon, Diplotaxis virgata, Erodium aethiopicum, Lavatera cretica, Malva nicaeensis, Silybum marianum and, particularly, Amaranthus blitoides could be used in the remediation of the contaminated area. A phytoremediation experiment was therefore performed to assess the potential use of native shrub species for soil restoration in Aznalcóllar. The results showed that Retama sphaerocarpa was the most effective plant for use in phytostabilization programs [157]. Later on, an afforestation soil remediation program was carried out involving the planting of Mediterranean riparian woodland species, such as Populus alba, Tamarix africana, Fraxinus angustifolia, and Salix atrocinerea, and the usual drier upland trees such as Quercus ilex subsp. ballota, Olea europaea var. sylvestris, Phillyrea angustifolia, Pistacia lentiscus, Rosmarinus officinalis, and Retama sphaerocarpa [158]. Domínguez et al. [159] monitored survival and growth patterns in these plantations over a 3-year period. Plant survival rates reached nearly 100% in riparian sites, while some species in the upland terrace sites showed the lowest survival rates. They did not find any relationship between plant efficiency and soil pollution, with PTE accumulation rates found to be very low, except for the Salicaceae species, which showed high Cd and Zn accumulation rates (1 and 200 mg kg−1, respectively). Another study carried out in the area found that nurse shrubs, such as Retama sphaerocarpa, were highly effective in reducing Quercus ilex mortality, and thus are a promising tool for the revegetation of polluted areas [160]. Retama spaherocarpa, which recorded the highest survival rates and strong tolerance levels, was found to be the most effective plant species for the restoration of these soils [161]. In addition to the revegetation mentioned above, plant growth phytostabilization had a considerable impact on PTE bioavailability [148,157]. In general, the amount of PTEs accumulated in the aerial parts of trees and shrubs was usually observed to be at the very low normal levels for higher plants [158,159], while different levels of accumulated PTEs were recorded in roots [157,162]. For example, Quercus ilex was capable of accumulating up to 7 g kg−1 of Cd in its fine roots [162] and Myrtus communis, Arbutus unedo, and Retama sphaerocarpa accumulated As in their roots [163]. The area affected by the Aznalcóllar mining accident has become an exceptional location for studying the phytoremediation potential of numerous species in a realistic context and for providing input for future phytoremediation projects.
Given Spain’s long history of a highly dynamic mining industry, large areas of the country have been affected by abandoned mining waste [164]. Some studies have collected the results of different plant-based rehabilitation projects in these locations [10,49,165,166]. For example, a reclamation project involving the plantation of Pinus sp. or Eucalyptus sp. trees and waste material amendments (sewage sludge and paper mill residues) in a Cu mining area located in Touro (Galicia, Spain) [165]. An evaluation of the different reclamation practices showed that soil planted with pines and eucalyptus trees increased Ni, Pb, and Zn retention in non-mobile soil fractions, while the amendments also attenuated Cr- and Cu-related pollution. The results obtained showed that vegetation provided by these tree species, together with the amendments, decreased PTE pollution in the area affected [165]. A field experiment involving the agricultural crop plants Hordeum vulgare, Triticum aestivum, and Lupinus luteus was also performed in an area called Almadén (Ciudad Real, Spain) containing the most unusual large soil concentrations of Hg in the world [167]. The wheat crop was found to extract concentrations of Hg of up to 0.479 µg g−1. However, the most interesting plant subjected to phytoextraction was barley, which accumulated up to 719 mg ha−1 of Hg [168]. Later studies carried out in this area showed that most Hg uptake by plants was through leaves, with root uptake rates being rather slow [169]. The Cartagena-La Union mining district, located on the Mediterranean coast in the southeast of Spain, is also affected by high levels of PTE accumulation (≈1100, 4700 and 5000 mg kg−1 of As, Pb, and Zn, respectively) [49,166]. A remediation experiment combining red mud, compost, and hydrated lime soil amendments with the halophyte plant species Atriplex helium and Zygophyllum fabago was performed. On the whole, soil amendments increased tailing pH and reduced PTE solubility and extractability, while the halophytes tested, which managed to grow only in amended tailings, accumulated low levels of Al, As, Cd, and Zn. This research showed that the combined use amendments and halophyte plant species is an effective AML phytostabilization strategy [166]. A large-scale field experiment, also conducted in this area, involved the reclamation of a tailing’s disposal pond through the spontaneous colonization of native species mediated by tilling, marble waste, and pig slurry. Five years later, Atriplex halimus, showing high metal concentrations, mainly in leaves, had become the predominant species. Accumulation levels were increased differentially using different amendments, with pig slurry increasing Cu concentrations and marble waste increasing Pb and Cd accumulation levels. In summary, Atriplex halimus could be a good candidate for AML phytoremediation [49]. Another study carried out in the Sarzedas mining area (Castelo Branco, Portugal), which was greatly affected by As (11.1–651.1 mg kg−1), Sb (30.5–5986.4 mg kg−1), and W (0.8–684.0 mg kg−1) contamination, analyzed spontaneous vegetation growth. The results showed that the following plants could be used to reduce PTE bioavailability: Pinus pinaster for As and W, Cistus ladanifer and Cistus vulgaris for Sb and W, Digitalis purpurea and Erica umbellata for Sb, and Quercus ilex, Chamaespartium tridentatum, and Eucalyptus globulus for W [170]. Elsewhere in Europe, with its large number of AMLs, a study of a tin mining area in Abbaretz (Loire-Atlantique, Pays de la Loire, France) highlighted the importance of improving soil conditions to establish vegetation correctly. In this research, the effect of biochar amendments combined with iron sulphate supplements on the growth and uptake of three Salicaceae species was evaluated. The results showed that these amendments improved soil health and raised PTE concentrations in the roots of Salix purpurea, which could be a good candidate for PTE phytostabilization [171]. Similarly, a field experiment was conducted in the Campo Pisano mining area (Sardinia, Italy), with 3250 mg kg−1 of Pb and 11,949 mg kg−1 of Zn, to study the feasibility of using Pistacia lentiscus and Scrophularia bicolor combined with soil amendments for phytoremediation purposes. The results showed that the plant species Pistacia lentiscus could be used effectively for PTE phytostabilization due to its capacity to accumulate PTEs in roots, as well as its high resistance and biomass production [172]. Other studies have investigated the establishment of plant cover to improve soil quality. For example, in two AMLs in northwestern Transylvania (Romania), a study was carried out to monitor the effect of different plant covers on soil health after 40 years of vegetation growth. The results revealed that the chemical, physical, and biological properties of the soil, which improved significantly during the period of the study, depended on the plant cover [35].
In the United States, over 550,000 abandoned mining sites have generated 45 billion tons of mining waste, including waste rock and tailing material, containing worrying levels of PTE concentrations [3]. While some researchers have been evaluating emerging phytoremediation technologies, most researchers have focused on the use of revegetation techniques to improve soil health. A revegetation study of three mining sites in Ohio, using different mixes of standard reclamation and native plants, compared the growth of each type of revegetation and its impact on soil microorganisms over a 2-year period. The results obtained highlighted the importance of using native plants which can improve soil health more effectively than non-native plants [173]. An Iron King Mine and Humboldt Smelter Superfund (IKMHSS) field study carried out by Gil-Loaiza, which found over 2000 mg kg−1 of Pb, As, and Zn, also demonstrated the importance of incorporating good native plant cover in AMLs [174]. Six different treatments containing different amounts of compost and a mix of native plants, which were pre-tested in the laboratory, were studied. It is interesting to note that plant growth in tailing soils was found to stabilize pH levels through rhizosphere alkalinisation and to prevent smelter tailing acidification. In summary, both studies showed that the establishment of vegetation cover improved soil quality through increased microbial activity and PTE phytostabilization [173,174].
In China, about 2.88 Mha of land, which is increasing at an alarming rate of 46,700 ha per year, has been degraded by mining operations [175]. The many studies of phytoremediation techniques used in China, outlining the most important advances made in this field, are reviewed below. The remediation of the Dabaoshan polymetallic mining area in the Guangdong province, with 681, 343 and 339 mg kg−1 of Pb, Cu and Zn, respectively, combining lime, organic fertilizer, and biochar soil amendments with Hibiscus cannabinus and Boehmeria nivea plant species, was studied. The results showed that amendments and plant growth improved soil health by reducing PTE bioavailability, solubilization, and migration in mine tailings [176]. The phytoextraction capacity of heavy metal-accumulating plants and chemical chelators has also been evaluated in Guangdong paddy fields that had been continuously irrigated with mining wastewater for 55 years (868.5, 433.6, 58.6, and 2.2 mg kg−1 of Pb, Zn, Cu, and Cd, respectively). The results suggested that Amaranthus hypochondriacus was highly effective for in situ PTE phytoremediation and that the application of ethylenediaminetetraacetic acid (EDTA) accelerated the phytoextraction effect [177]. A field experiment involving plants, microorganisms, and amendments was also carried out on the Anshan Mining Group Corporation’s metal-contaminated mine tailings in the Liaoning province (51.7, 151.5, 43.6, and 1960.0 mg kg−1 of Cu, Zn, Pb, and Mn, respectively). The experiment, which combined Glycine max and Mucor circinelloides fungi, as well as the application of rice husk, biochar, and ceramsite soil amendments in 17 phytoremediation tree plots, was carried out over a period of 105 days. The combination of these techniques was found to improve PTE phytoextraction capacity, reduce PTE soil concentrations, and to increase microbial soil activity [178]. Similar results were reported by Yu et al. [179] using Celosia argentea and ammonium chloride, Bacillus megaterium, and citric acid soil amendments for 10 months in mine tailing-contaminated soil in southern China (3.68 mg kg−1 of Cd). Soil Cd concentrations were decreased after three harvesting events, with overall reductions in Cd of 40.5%, 46,1%, and 105% following applications of ammonium chloride, B. megaterium, and citric acid, respectively. The study showed that the use of C. argentea combined with soil amendments could greatly improve soil enzyme activity and the quality of Cd-contaminated soils. Another experiment in Chenzhou (Hunan province), one of the largest ferrous metal mines in China, assessed the restoration and accumulation potential of selected hybrid sweet sorghum species in contaminated areas (288–1270, 97.7–1250 and 2–5.7 mg kg−1 of Zn, Pb, and Cd, respectively). The results showed that hybrid sweet sorghum species had greater accumulation and tolerance capacity than local species. The study concluded that it would be economically viable to use sweet sorghum, which densifies biofuel production, for PTE remediation in this mining area [180]. Similarly, an evaluation of the Manihot esculenta bioethanol plantation in a severely PTE-contaminated mining area found that Manihot esculenta could produce 3680–4160 L of bioethanol per hectare (L ha−1) on contaminated soils. Manihot esculenta also improved soil fertility by reducing mobile and bioavailable metal fractions. The cultivation of Manihot esculenta could therefore be used for AML remediation and also generate economic benefits [181]. Other AML phytoremediation projects have evaluated the relationship between revegetation and improved chemical, physical, and microbiological soil properties. Research conducted in the AnTaibao, Anjialing, and East opencast coal mining areas analyzed the effects of revegetation on physical, chemical, and microbiological soil properties in nine plots for 1–13 years. The results obtained showed that the plant species significantly enhanced soil health by improving soil field capacity, organic content, aggregation, microbiological composition, and total microorganisms excluding other factors such as soil texture in these areas [175]. Revegetation projects were carried out in the Lechand Pb/Zn mine in Guangdong province, which is severely contaminated by Pb (29.9 mg kg−1), Zn (50.7 mg kg−1) Cu (14.7 mg kg−1), and Cd (0.52 mg kg−1). A combination of domestic waste and complex fertilizer amendments and plants was evaluated to assess their suitability for revegetation purposes. The results showed that Vertiveria zizanioides was the best plant species for tailings revegetation, followed by Paspalum notatum, Cynodon dactylon, and Imperata cylindrica var. major [182]. Similarly, research carried out in the Jiangxi province, which is affected by ionic rare earth element mine tailings, showed that a combination of organic amendments and plants could regenerate bacterial community structures and diversity, improve soil quality, and possibly reduce PTE bioavailability [183]. Similar results were reported by Mi et al. [184] in the Kee Open Pit Mine in Shanxi province, where the use of mine dumps for crop cultivation improved soil health as measured by soil properties and microbial communities. On the other hand, different periods of revegetation were evaluated in metal mining dump areas (up to 306.5, 1863.9, 494.5, 353, and 133.3 mg kg−1 of Cu, Mn, Cr, Ni, and Zn, respectively) in the Sichuan province to determine the long-term impact of revegetation. The results showed that plant species with low vegetation diversity had a strong impact on PTE remediation, but that the effect of soil fertility restoration was less than that of natural vegetation, while spontaneous vegetation succession reduced the rate of PTE remediation. The study suggested that revegetation techniques need to be monitored to ensure that succession does not disrupt soil remediation [185]. Finally, it is important to highlight the study carried out by Wan et al. [14], in which the costs and benefits of a phytoremediation project involving soil contaminated with As, Cd, and Pb are calculated. They concluded that phytoremediation costs for this project were lower than those reported for other remediation projects. Moreover, given the cost of environmental pollution, the benefits of phytoremediation are expected to offset project costs in less than seven years.
The Asia-Pacific region, one the largest producers of coal, accounts for over 67% of total world production. Land degradation due to coal mining operations is reported to affect 1400 additional hectares each year [186]. Several studies have described phytoremediation projects, in which different soil restoration strategies involving herbaceous plants and trees in coal mining areas have been used [186,187,188,189]. The use of Miscanthus x giganteus as an effective plant to restore soil in a lignite mine has been analyzed by Jeżowski et al. [188]. This 3-year field study of lignite mining waste heaps showed positive results in terms of soil regeneration. A phytoremediation study of the removal of toxic iron and sulphur aqueous waste spilled on soils around coal mines was also carried out in the Jaintia hills (Meghalaya, northeastern India). The use of agricultural lime and organic manure soil amendments, together with plants, reduced sulphate concentrations from 22 to 5 mg kg−1 and soluble iron from 476 to 109 mg kg−1. Moreover, this reclamation strategy improved soil fertility by increasing plant nutrients, pH (up to normal levels) and microbial biomass [186]. The phytoextraction capacity of Eucalyptus sp. was evaluated in the Jharia coal mine (Dhanbad, India), the results of which were compared with PTE content measured in soil and trees in 1995. The study concluded that plant-based reclamation reduced Pb, Zn, and Cu concentrations to their levels in 1995, while PTEs mainly accumulated in the stem bark (Pb), leaves (Zn and Mn), and roots (Cu and Co) of trees [189].

{kind=link}
Table 1.
List of hyperaccumulator plants that could be used for phytoextraction. Asterisks indicate approximate values (*).
Table 1.
List of hyperaccumulator plants that could be used for phytoextraction. Asterisks indicate approximate values (*).
| Species | Maximum Data of PTEs Concentration in Leaves (mg kg−1 DW) | References | |
|---|---|---|---|
| Alyssum sp. | Ni | 20,100 | [190] |
| Arabidopsis halleri | Cd | 5722 | [191] |
| Zn | 9491 | ||
| Arabis gemmifera | Cd | 1810 | [192] |
| Zn | 20,300 | ||
| Atriplex halimus | Cd | 850 * | [193] |
| Berkheya coddii | Ni | 31,300 | [194] |
| Bidens pilosa | Cd | 144.1 | [195] |
| Biscutella laevigata | Tl | 66.7 | [196] |
| Brassica pekinensis | Pb | 2670 | [197] |
| Carpobrotus rossi | Cd | 172 | [198] |
| Lantana cámara | Cd | 423.06 | [46] |
| Leersia hexandra | Cr | 5005 | [199] |
| Lonicera japonica | Cd | 57.22 | [200] |
| Phytolacca acinosa | Mn | 19,300 | [201] |
| Polygonum lapathifolium | Mn | 18,841.7 | [202] |
| Pteris vittata | As | 1530 | [203] |
| Schima superba | Mn | 9975.61 | [204,205] |
| Sedum alfredii | Cd | 9000 | [206] |
| Zn | 4807 | [207] | |
| Sesbania drummondi | Pb | 5000 * | [208] |
| Solanum sp. | Cd | 142.7 | [209] |
| Thlaspi sp. | Cd | 164 | [210] |
| Ni | 16,600 | [190] | |
| Zn | 19,070 | [210] | |
Table 2.
List of plants capable of volatilizing PTEs.
| Species | PTEs | References |
|---|---|---|
| Pteris vittata | As | [211] |
| Arundo donax | [56] | |
| Transgenic Arabidopsis thaliana (fungal gene WaarsM) | [212] | |
| Dittrichia viscosa | [213] | |
| Oryza sativa | [214] | |
| Zea mays | Hg | [57] |
| Transgenic Nicotiana tabacum (bacterial gene merA and merB) | [215] | |
| Haliminone portulacoides | [216] | |
| Arabidopsis thaliana | [217] | |
| Brassica juncea | Se | [218] |
| Oryza sativa Brassica oleracea Daucus carota Hordeum vulgare Medicago sativa Lycopersicon esculentum Cucumis sativus Gossypium hirsutum Solanum melongena Zea mays | [219] | |
| Populus tremula x alba | [220] | |
| Stanleya pinnata | [221] | |
| Agrostis tenuis Hordeum vulgare | [222] |
Table 3.
List of suitable plants for phytostabilization purposes.
| Species | PTEs | References |
|---|---|---|
| Acanthus ilicifolius | Cd | [223] |
| Agrostis capillaris | Pb | [224] |
| Arundo donax | Ni, Pb and Hg | [225] |
| Atriplex halimus | Pb and Cd | [49,94] |
| Biscutella auriculata | Pb, Cu and Cd | [19,69,70] |
| Brassica juncea | Cu and Zn | [63] |
| Brassica juncea Dactylis glomerata | Cd, Pb and Zn | [226] |
| Casdaminopsis arenosa | Zn, Cd and Pb | [227] |
| Conocarpus erectus Populus deltoides | As | [228] |
| Dahlia pinnata | As | [229] |
| Imperata condensata | Cu | [230] |
| Iris lactea | Cd | [126] |
| Jatropha curcas | Al, Zn and Cu | [231] |
| Lolium perenne | Cu | [232] |
| Lupinus albus | Cd, As and Pb | [156,233] |
| Miscanthus sacchariflorus | Cd | [234] |
| Miscanthus sinensis x giganteus | Zn, Cd and Pb | [235] |
| Pistacia lentiscus | Pb | [236] |
| Pteridium aquilinum | Cd, Cu, Pb and Zn | [236] |
| Quercus ilex | Cd | [162] |
| Ricinus communis | Pb | [237] |
| Salix purpurea | As | [171] |
| Typha latifolia | Co, Cd, Ni, Mn, Cr and As | [238] |
The results of AML restoration projects described above showed that the establishment of vegetation cover in these areas effectively reduced PTE bioavailability and increased soil quality, which constituted a further step towards a total restoration of the ecosystem.
6. Conclusions
Soil restoration in accordance with ecologically sustainable development norms is essential once mining operations are closed. The goal of land reclamation is to return degraded AMLs to their original state and to ensure that the ecosystem’s normal interactions between biotic and abiotic components take place. Plant-based techniques, which include phytoextraction, phytostabilization, and phytovolatilization, are environmentally-friendly and cost-effective alternatives to traditional remediation procedures which are usually more destructive to soils. However, the principal drawback of this remediation strategy is the need to select plants that can meet the objectives set out in the strategy. Their PTE tolerance and accumulation systems must enable the plants selected to play an effective role in AML reclamation. Implementation of a phytoremediation strategy will therefore include the following steps: (a) soil physical-chemical characterization; (b) description of climatic conditions; (c) identification of plants capable of growing in the area; (d) assessment of phytoremediation capacities; (e) characterization of PTE tolerance and accumulation mechanisms; and, finally, (f) in situ experiments to assess the success of the strategy. In many cases, little is known about the underlying tolerance and accumulation mechanisms. A better understanding of these mechanisms is crucial to decipher and thus to enhance a plant’s capacity to be used for phytoextraction, phytostabilization, and phytovolatilization. This will also facilitate the development of transgenic plants, which may be of great potential significance in the future. In situ phytoremediation studies of AMLs demonstrated that plant-based techniques regenerate soil fertility, reduce PTE bioavailability, and increase biodiversity on-site. New market opportunities in mine land phytoremediation, which uses plants and bio-ores to obtain, in particular, critical precious metals, should be explored. However, research in this field, which is still in its infancy, needs to better identify plants suitable for these purposes in order to enhance their capacity to extract, reduce, and stabilize PTE concentrations in AMLs, as well as to accumulate metals in shoots to facilitate precious metal recovery.
Author Contributions
Conceptualization, J.D.P.; resources and investigation, J.D.P., P.H., J.A.C., J.M.E., L.M.S., M.M.M. and F.B.-B.; writing—original draft preparation, J.D.P. and P.H.; writing—review and editing, P.H., J.A.C., J.M.E., L.M.S., M.M.M. and F.B.-B.; supervision, L.M.S., P.H., and J.A.C.; project administration, P.H. and L.M.S.; funding acquisition, L.M.S., P.H. and M.M.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Spanish Ministry of Science, Innovation and Universities (CGL2015–67644–R and PGC2018–098372–B–100), the Castilla–La Mancha Regional Government (SBPLY/17/180501/000273) and the University of Castilla La–Mancha RDi Internal Research Plan (predoctoral contract 2016/14100).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
We wish to thank Michael O’Shea for proofreading the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Elmayel, I.; Esbrí, J.M.; García-Ordiales, E.; Elouaer, Z.; Garcia-Noguero, E.M.; Bouzid, J.; Campos, J.A.; Higueras, P.L. Biogeochemical assessment of the impact of Zn mining activity in the area of the Jebal Trozza mine, Central Tunisia. Environ. Geochem. Health 2020, 42, 3529–3542. [Google Scholar] [CrossRef] [PubMed]
- Jain, M.K.; Das, A. Impact of Mine Waste Leachates on Aquatic Environment: A Review. Curr. Pollut. Rep. 2017, 3, 31–37. [Google Scholar] [CrossRef]
- Venkateswarlu, K.; Nirola, R.; Kuppusamy, S.; Thavamani, P.; Naidu, R.; Megharaj, M. Abandoned metalliferous mines: Ecological impacts and potential approaches for reclamation. Rev. Environ. Sci. Biotechnol. 2016, 15, 327–354. [Google Scholar] [CrossRef]
- Tchounwou, P.B.; Yedjou, C.G.; Patlolla, A.K.; Sutton, D.J. Heavy metal toxicity and the environment. Mol. Clin. Environ. Toxicol. 2012, 101, 133–164. [Google Scholar]
- Sengupta, M. Environmental Impacts of Mining: Monitoring, Restoration, and Control; Routledge: London, UK, 2018; ISBN 9781351450539. [Google Scholar]
- United Nations. Transforming Our World: The 2030 Agenda for Sustainable Development. Available online: https://sustainabledevelopment.un.org/post2015/transformingourworld (accessed on 27 May 2021).
- Xie, L.; van Zyl, D. Distinguishing reclamation, revegetation and phytoremediation, and the importance of geochemical processes in the reclamation of sulfidic mine tailings: A review. Chemosphere 2020, 252, 126446. [Google Scholar] [CrossRef] [PubMed]
- Dada, E.; Njoku, K.; Osuntoki, A.; Akinola, M. A review of current techniques of in situ Physico-chemical and biological remediation of heavy metals polluted soil. Ethiop. J. Environ. Stud. Manag. 2015, 8, 606. [Google Scholar] [CrossRef] [Green Version]
- Mahar, A.; Wang, P.; Ali, A.; Awasthi, M.K.; Lahori, A.H.; Wang, Q.; Li, R.; Zhang, Z. Challenges and opportunities in the phytoremediation of heavy metals contaminated soils: A review. Ecotoxicol. Environ. Saf. 2016, 126, 111–121. [Google Scholar] [CrossRef]
- Rodríguez, L.; Gómez, R.; Sánchez, V.; Villaseñor, J.; Alonso-Azcárate, J. Performance of waste-based amendments to reduce metal release from mine tailings: One-year leaching behaviour. J. Environ. Manag. 2018, 209, 1–8. [Google Scholar] [CrossRef]
- Rodríguez, L.; Gómez, R.; Sánchez, V.; Alonso-Azcárate, J. Chemical and plant tests to assess the viability of amendments to reduce metal availability in mine soils and tailings. Environ. Sci. Pollut. Res. 2016, 23, 6046–6054. [Google Scholar] [CrossRef]
- Ruiz, E.; Rodríguez, L.; Alonso-Azcárate, J.; Rincón, J. Phytoextraction of metal polluted soils around a Pb-Zn mine by crop plants. Int. J. Phytoremediat. 2009, 11, 360–384. [Google Scholar] [CrossRef]
- Yan, A.; Wang, Y.; Tan, S.N.; Mohd Yusof, M.L.; Ghosh, S.; Chen, Z. Phytoremediation: A Promising Approach for Revegetation of Heavy Metal-Polluted Land. Front. Plant Sci. 2020, 11, 1–15. [Google Scholar] [CrossRef]
- Wan, X.; Lei, M.; Chen, T. Cost-benefit calculation of phytoremediation technology for heavy-metal-contaminated soil. Sci. Total Environ. 2016, 563–564, 796–802. [Google Scholar] [CrossRef]
- Ali, H.; Khan, E.; Sajad, M.A. Phytoremediation of heavy metals—Concepts and applications. Chemosphere 2013, 91, 869–881. [Google Scholar] [CrossRef]
- Suman, J.; Uhlik, O.; Viktorova, J.; Macek, T. Phytoextraction of heavy metals: A promising tool for clean-up of polluted environment? Front. Plant Sci. 2018, 9, 1476. [Google Scholar] [CrossRef] [Green Version]
- Dickinson, N. Phytoremediation. Encycl. Appl. Plant Sci. 2016, 3, 327–331. [Google Scholar]
- Sarma, H. Metal hyperaccumulation in plants: A review focusing on phytoremediation technology. J. Environ. Sci. Technol. 2011, 4, 118–138. [Google Scholar] [CrossRef] [Green Version]
- Peco, J.D.; Campos, J.A.; Romero-Puertas, M.C.; Olmedilla, A.; Higueras, P.; Sandalio, L.M. Characterization of mechanisms involved in tolerance and accumulation of Cd in Biscutella auriculata L. Ecotoxicol. Environ. Saf. 2020, 201, 110784. [Google Scholar] [CrossRef]
- Sanz-Fernández, M.; Rodríguez-Serrano, M.; Sevilla-Perea, A.; Pena, L.; Mingorance, M.D.; Sandalio, L.M.; Romero-Puertas, M.C. Screening Arabidopsis mutants in genes useful for phytoremediation. J. Hazard. Mater. 2017, 335, 143–151. [Google Scholar] [CrossRef]
- Macek, T.; Kotrba, P.; Svatos, A.; Novakova, M.; Demnerova, K.; Mackova, M. Novel roles for genetically modified plants in environmental protection. Trends Biotechnol. 2008, 26, 146–152. [Google Scholar] [CrossRef]
- Ferreira, R.A.; Pereira, M.F.; Magalhães, J.P.; Maurício, A.M.; Caçador, I.; Martins-Dias, S. Assessing local acid mine drainage impacts on natural regeneration-revegetation of São Domingos mine (Portugal) using a mineralogical, biochemical and textural approach. Sci. Total Environ. 2021, 755, 142825. [Google Scholar] [CrossRef]
- Moreno González, R.; Cánovas, C.R.; Olías, M.; Macías, F. Seasonal variability of extremely metal rich acid mine drainages from the Tharsis mines (SW Spain). Environ. Pollut. 2020, 259, 113829. [Google Scholar] [CrossRef]
- Pavoni, E.; Covelli, S.; Adami, G.; Baracchini, E.; Cattelan, R.; Crosera, M.; Higueras, P.; Lenaz, D.; Petranich, E. Mobility and fate of Thallium and other potentially harmful elements in drainage waters from a decommissioned Zn-Pb mine (North-Eastern Italian Alps). J. Geochem. Explor. 2018, 188, 1–10. [Google Scholar] [CrossRef]
- Sánchez-Donoso, R.; Martín-Duque, J.F.; Crespo, E.; Higueras, P.L. Tailing’s geomorphology of the San Quintín mining site (Spain): Landform catalogue, aeolian erosion and environmental implications. Environ. Earth Sci. 2019, 78, 166. [Google Scholar] [CrossRef]
- Akcil, A.; Koldas, S. Acid Mine Drainage (AMD): Causes, treatment and case studies. J. Clean. Prod. 2006, 14, 1139–1145. [Google Scholar] [CrossRef]
- United States Department of Agriculture of Soil Health. NRCS Soils. Available online: https://www.nrcs.usda.gov/wps/portal/nrcs/main/soils/health/ (accessed on 2 March 2021).
- Gallego, S.; Esbrí, J.M.; Campos, J.A.; Peco, J.D.; Martin-Laurent, F.; Higueras, P. Microbial diversity and activity assessment in a 100-year-old lead mine. J. Hazard. Mater. 2020, 420, 124618. [Google Scholar] [CrossRef]
- Muthusaravanan, S.; Sivarajasekar, N.; Vivek, J.S.; Paramasivan, T.; Naushad, M.; Prakashmaran, J.; Gayathri, V.; Al-Duaij, O.K. Phytoremediation of heavy metals: Mechanisms, methods and enhancements. Environ. Chem. Lett. 2018, 16, 1339–1359. [Google Scholar] [CrossRef]
- Ghori, Z.; Iftikhar, H.; Bhatti, M.F.; Nasar-um-Minullah; Sharma, I.; Kazi, A.G.; Ahmad, P. Phytoextraction; Elsevier Inc.: Amsterdam, The Netherlands, 2015; ISBN 9780128031582. [Google Scholar]
- Grobelak, A. Organic soil amendments in the phytoremediation process. In Phytoremediation: Management of Environmental Contaminants; Springer International Publishing: Cham, Switzerland, 2016; Volume 4, pp. 21–39. ISBN 9783319418117. [Google Scholar]
- Jadia, C.D.; Fulekar, M.H. Phytoremediation of heavy metals: Recent techniques. Afr. J. Biotechnol. 2009, 8, 921–928. [Google Scholar]
- Zhao, X.; Sun, Y.; Huang, J.; Wang, H.; Tang, D. Effects of soil heavy metal pollution on microbial activities and community diversity in different land use types in mining areas. Environ. Sci. Pollut. Res. 2020, 27, 20215–20226. [Google Scholar] [CrossRef]
- Sheoran, V.; Sheoran, A.S.; Poonia, P. Soil Reclamation of Abandoned Mine Land by Revegetation: A Review. Int. J. Soil Sediments Water 2010, 3, 13. [Google Scholar]
- Buta, M.; Blaga, G.; Paulette, L.; Păcurar, I.; Roșca, S.; Borsai, O.; Grecu, F.; Sînziana, P.E.; Negrușier, C. Soil Reclamation of Abandoned Mine Lands by Revegetation in Northwestern Part of Transylvania: A 40-Year Retrospective Study. Sustainability 2019, 11, 3393. [Google Scholar] [CrossRef] [Green Version]
- Alkorta, I.; Hernández-Allica, J.; Becerril, J.M.; Amezaga, I.; Albizu, I.; Garbisu, C. Recent findings on the phytoremediation of soils contaminated with environmentally toxic heavy metals and metalloids such as zinc, cadmium, lead, and arsenic. Rev. Environ. Sci. Biotechnol. 2004, 3, 71–90. [Google Scholar] [CrossRef]
- Liu, L.; Li, W.; Song, W.; Guo, M. Remediation techniques for heavy metal-contaminated soils: Principles and applicability. Sci. Total Environ. 2018, 633, 206–219. [Google Scholar] [CrossRef] [PubMed]
- Black, H. Absorbing possibilities: Phytoremediation. Environ. Health Perspect. 1995, 103, 1106–1108. [Google Scholar] [CrossRef] [PubMed]
- Chalot, M.; Blaudez, D.; Rogaume, Y.; Provent, A.S.; Pascual, C. Fate of trace elements during the combustion of phytoremediation wood. Environ. Sci. Technol. 2012, 46, 13361–13369. [Google Scholar] [CrossRef]
- Corzo Remigio, A.; Chaney, R.L.; Baker, A.J.M.; Edraki, M.; Erskine, P.D.; Echevarria, G.; van der Ent, A. Phytoextraction of high value elements and contaminants from mining and mineral wastes: Opportunities and limitations. Plant Soil. 2020, 449, 11–37. [Google Scholar] [CrossRef]
- Novo, L.A.B.; Castro, P.M.L.; Alvarenga, P.; da Silva, E.F. Phytomining of rare and valuable metals. In Phytoremediation: Management of Environmental Contaminants; Springer International Publishing: Cham, Switzerland, 2017; Volume 5, pp. 469–486. ISBN 9783319523811. [Google Scholar]
- Li, Y.M.; Chaney, R.L.; Brewer, E.P.; Angle, J.S.; Nelkin, J. Phytoextraction of nickel and cobalt by hyperaccumulator Alyssum species grown on nickel-contaminated soils. Environ. Sci. Technol. 2003, 37, 1463–1468. [Google Scholar] [CrossRef]
- Nkrumah, P.N.; Echevarria, G.; Erskine, P.D.; Chaney, R.L.; Sumail, S.; van der Ent, A. Growth effects in tropical nickel-agromining ‘metal crops” in response to nutrient dosing. J. Plant Nutr. Soil Sci. 2019, 182, 715–728. [Google Scholar] [CrossRef]
- Barbaroux, R.; Plasari, E.; Mercier, G.; Simonnot, M.O.; Morel, J.L.; Blais, J.F. A new process for nickel ammonium disulfate production from ash of the hyperaccumulating plant Alyssum murale. Sci. Total Environ. 2012, 423, 111–119. [Google Scholar] [CrossRef]
- Gisbertz, K.; Hilgendorf, S.; Friedrich, B.; Heinrichs, S.; Rüßmann, D.; Pretz, T. Maximising metal recovery from incineration ashes. In Proceedings of the European Metallurgical Conference, EMC 2013, Weimar, Germany, 23–26 June 2013; pp. 1127–1132. [Google Scholar]
- Liu, S.; Ali, S.; Yang, R.; Tao, J.; Ren, B. A newly discovered Cd-hyperaccumulator Lantana camara L. J. Hazard. Mater. 2019, 371, 233–242. [Google Scholar] [CrossRef]
- Grčman, H.; Velikonja-Bolta, Š.; Vodnik, D.; Kos, B.; Leštan, D. EDTA enhanced heavy metal phytoextraction: Metal accumulation, leaching and toxicity. Plant Soil 2001, 235, 105–114. [Google Scholar] [CrossRef]
- Moslehi, A.; Feizian, M.; Higueras, P.; Eisvand, H.R. Assessment of EDDS and vermicompost for the phytoextraction of Cd and Pb by sunflower (Helianthus annuus L.). Int. J. Phytoremediat. 2019, 21, 191–199. [Google Scholar] [CrossRef]
- Acosta, J.A.; Abbaspour, A.; Martínez, G.R.; Martínez-Martínez, S.; Zornoza, R.; Gabarrón, M.; Faz, A. Phytoremediation of mine tailings with Atriplex halimus and organic/inorganic amendments: A five-year field case study. Chemosphere 2018, 204, 71–78. [Google Scholar] [CrossRef]
- Nörtemann, B. Biodegradation of Chelating Agents: EDTA, DTPA, PDTA, NTA, and EDDS. Biogeochem. Chelating Agents 2005, 150–170. [Google Scholar] [CrossRef]
- Sun, B.; Zhao, F.J.; Lombi, E.; McGrath, S.P. Leaching of heavy metals from contaminated soils using EDTA. Environ. Pollut. 2001, 113, 111–120. [Google Scholar] [CrossRef]
- Meers, E.; Ruttens, A.; Hopgood, M.J.; Samson, D.; Tack, F.M.G. Comparison of EDTA and EDDS as potential soil amendments for enhanced phytoextraction of heavy metals. Chemosphere 2005, 58, 1011–1022. [Google Scholar] [CrossRef]
- Meers, E.; Hopgood, M.; Lesage, E.; Vervaeke, P.; Tack, F.M.G.; Verloo, M.G. Enhanced phytoextraction: In search of EDTA alternatives. Int. J. Phytoremediat. 2004, 6, 95–109. [Google Scholar] [CrossRef]
- Duquène, L.; Vandenhove, H.; Tack, F.; Meers, E.; Baeten, J.; Wannijn, J. Enhanced phytoextraction of uranium and selected heavy metals by Indian mustard and ryegrass using biodegradable soil amendments. Sci. Total Environ. 2009, 407, 1496–1505. [Google Scholar] [CrossRef]
- Sakakibara, M.; Watanabe, A.; Sano, S.; Inoue, M.; Kaise, T. Phytoextraction and phytovolatili-zation of arsenic from as-contaminated soils by Pteris vittata. In Proceedings of the Association for Environmental Health and Sciences—22nd Annual International Conference on Contaminated Soils, Sediments and Water 2006, Amherst, MA, USA, 16–19 October 2006; Volume 12, pp. 258–263. [Google Scholar]
- Guarino, F.; Miranda, A.; Castiglione, S.; Cicatelli, A. Arsenic phytovolatilization and epigenetic modifications in Arundo donax L. assisted by a PGPR consortium. Chemosphere 2020, 251, 126310. [Google Scholar] [CrossRef]
- Mello, I.S.; Targanski, S.; Pietro-Souza, W.; Frutuoso Stachack, F.F.; Terezo, A.J.; Soares, M.A. Endophytic bacteria stimulate mercury phytoremediation by modulating its bioaccumulation and volatilization. Ecotoxicol. Environ. Saf. 2020, 202, 110818. [Google Scholar] [CrossRef]
- Raskin, I.; Ensley, B.D. Phytoremediation of Toxic Metals-Using Plants to Clean Up the Environment; Wiley Inter-Science Publication: New York, NY, USA, 2001. [Google Scholar]
- Shackira, A.M.; Puthur, J.T. Phytostabilization of Heavy Metals: Understanding of Principles and Practices. In Plant-Metal Interactions; Springer International Publishing: Cham, Switzerland, 2019; pp. 263–282. [Google Scholar]
- Marques, A.P.G.C.; Rangel, A.O.S.S.; Castro, P.M.L. Remediation of heavy metal contaminated soils: Phytoremediation as a potentially promising clean-Up technology. Crit. Rev. Environ. Sci. Technol. 2009, 39, 622–654. [Google Scholar] [CrossRef]
- Lee, S.H.; Park, H.; Koo, N.; Hyun, S.; Hwang, A. Evaluation of the effectiveness of various amendments on trace metals stabilization by chemical and biological methods. J. Hazard. Mater. 2011, 188, 44–51. [Google Scholar] [CrossRef]
- Ruttens, A.; Mench, M.; Colpaert, J.V.; Boisson, J.; Carleer, R.; Vangronsveld, J. Phytostabilization of a metal contaminated sandy soil. I: Influence of compost and/or inorganic metal immobilizing soil amendments on phytotoxicity and plant availability of metals. Environ. Pollut. 2006, 144, 524–532. [Google Scholar] [CrossRef]
- Pérez-Esteban, J.; Escolástico, C.; Moliner, A.; Masaguer, A.; Ruiz-Fernández, J. Phytostabilization of metals in mine soils using Brassica juncea in combination with organic amendments. Plant Soil 2014, 377, 97–109. [Google Scholar] [CrossRef] [Green Version]
- McGrath, S.P.; Zhao, F.J.; Lombi, E. Plant and rhizosphere processes involved in phytoremediation of metal-contaminated soils. Plant Soil 2001, 232, 207–214. [Google Scholar] [CrossRef]
- Sheng, X.F.; Xia, J.J.; Jiang, C.Y.; He, L.Y.; Qian, M. Characterization of heavy metal-resistant endophytic bacteria from rape (Brassica napus) roots and their potential in promoting the growth and lead accumulation of rape. Environ. Pollut. 2008, 156, 1164–1170. [Google Scholar] [CrossRef]
- Mastretta, C.; Taghavi, S.; Van Der Lelie, D.; Mengoni, A.; Galardi, F.; Gonnelli, C.; Barac, T.; Boulet, J.; Weyens, N.; Vangronsveld, J. Endophytic bacteria from seeds of Nicotiana tabacum can reduce cadmium phytotoxicity. Int. J. Phytoremediat. 2009, 11, 251–267. [Google Scholar] [CrossRef]
- Quiñones, M.A.; Ruiz-Díez, B.; Fajardo, S.; López-Berdonces, M.A.; Higueras, P.L.; Fernández-Pascual, M. Lupinus albus plants acquire mercury tolerance when inoculated with an Hg-resistant Bradyrhizobium strain. Plant Physiol. Biochem. 2013, 73, 168–175. [Google Scholar] [CrossRef] [Green Version]
- Rock, S.; Pivetz, B.; Madalinski, K.; Adams, N.; Wilson, T. Introduction to Phytoremediation; U.S. Environmental Protection Agency: Washington, DC, USA, 2019.
- Peco, J.D.; Higueras, P.; Campos, J.A.; Olmedilla, A.; Romero-Puertas, M.C.; Sandalio, L.M. Deciphering lead tolerance mechanisms in a population of the plant species Biscutella auriculata L. from a mining area: Accumulation strategies and antioxidant defenses. Chemosphere 2020, 261, 127721. [Google Scholar] [CrossRef]
- Peco, J.D.; Sandalio, L.M.; Higueras, P.; Olmedilla, A.; Campos, J.A. Characterization of the biochemical basis for copper homeostasis and tolerance in Biscutella auriculata L. Physiol. Plant. 2020. [Google Scholar] [CrossRef]
- Reeves, R.D.; Baker, A.J.M.; Jaffré, T.; Erskine, P.D.; Echevarria, G.; van der Ent, A. A global database for plants that hyperaccumulate metal and metalloid trace elements. New Phytol. 2018, 218, 407–411. [Google Scholar] [CrossRef] [Green Version]
- Pošćić, F.; Marchiol, L.; Schat, H. Hyperaccumulation of thallium is population-specific and uncorrelated with caesium accumulation in the thallium hyperaccumulator, Biscutella laevigata. Plant Soil 2013, 365, 81–91. [Google Scholar] [CrossRef]
- McIntyre, T. Phytoremediation of heavy metals from soils. Adv. Biochem. Eng. Biotechnol. 2003, 78, 97–123. [Google Scholar] [PubMed]
- Pollard, A.J.; Powell, K.D.; Harper, F.A.; Smith, J.A.C. The genetic basis of metal hyperaccumulation in plants. Crit. Rev. Plant Sci. 2002, 21, 539–566. [Google Scholar] [CrossRef]
- Ke, W.; Xiong, Z.T.; Chen, S.; Chen, J. Effects of copper and mineral nutrition on growth, copper accumulation and mineral element uptake in two Rumex japonicus populations from a copper mine and an uncontaminated field sites. Environ. Exp. Bot. 2007, 59, 59–67. [Google Scholar] [CrossRef]
- Kabata-Pendias, A. Trace Elements in Soils and Plants; CRC press: Boca Raton, FL, USA, 2001; ISBN 0849315751. [Google Scholar]
- Wang, X.P.; Shan, X.Q.; Zhang, S.Z.; Wen, B. A model for evaluation of the phytoavailability of trace elements to vegetables under the field conditions. Chemosphere 2004, 55, 811–822. [Google Scholar] [CrossRef]
- Kim, K.-R.; Owens, G.; Naidu, R. Heavy metal distribution, bioaccessibility, and phytoavailability in long-term contaminated soils from Lake Macquarie, Australia. Soil Res. 2009, 47, 166. [Google Scholar] [CrossRef]
- Filipović, L.; Romić, M.; Romić, D.; Filipović, V.; Ondrašek, G. Organic matter and salinity modify cadmium soil (phyto) availability. Ecotoxicol. Environ. Saf. 2018, 147, 824–831. [Google Scholar] [CrossRef]
- Stafford, A.; Jeyakumar, P.; Hedley, M.; Anderson, C. Influence of Soil Moisture Status on Soil Cadmium Phytoavailability and Accumulation in Plantain (Plantago lanceolata). Soil Syst. 2018, 2, 9. [Google Scholar] [CrossRef] [Green Version]
- Ren, J.H.; Ma, L.Q.; Sun, H.J.; Cai, F.; Luo, J. Antimony uptake, translocation and speciation in rice plants exposed to antimonite and antimonate. Sci. Total Environ. 2014, 475, 83–89. [Google Scholar] [CrossRef]
- Kim, R.Y.; Yoon, J.K.; Kim, T.S.; Yang, J.E.; Owens, G.; Kim, K.R. Bioavailability of heavy metals in soils: Definitions and practical implementation—a critical review. Environ. Geochem. Health 2015, 37, 1041–1061. [Google Scholar] [CrossRef]
- Dong, J.; Wu, F.; Zhang, G. Influence of cadmium on antioxidant capacity and four microelement concentrations in tomato seedlings (Lycopersicon esculentum). Chemosphere 2006, 64, 1659–1666. [Google Scholar] [CrossRef]
- Yang, C.M.; Juang, K.W. Alleviation effects of calcium and potassium on cadmium rhizotoxicity and absorption by soybean and wheat roots. J. Plant Nutr. Soil Sci. 2015, 178, 748–754. [Google Scholar] [CrossRef]
- Tangahu, B.V.; Sheikh Abdullah, S.R.; Basri, H.; Idris, M.; Anuar, N.; Mukhlisin, M.; Sheikh Abdullah, S.R.; Idris, M.; Tangahu, B.V.; Basri, H. A Review on Heavy Metals (As, Pb, and Hg) Uptake by Plants through Phytoremediation. Int. J. Chem. Eng. 2011, 2011, 939161. [Google Scholar] [CrossRef]
- Römheld, V.; Awad, F. Significance of root exudates in acquisition of heavy metals from a contaminated calcareous soil by graminaceous species. J. Plant Nutr. 2000, 23, 1857–1866. [Google Scholar] [CrossRef]
- Kim, S.; Lim, H.; Lee, I. Enhanced heavy metal phytoextraction by Echinochloa crus-galli using root exudates. J. Biosci. Bioeng. 2010, 109, 47–50. [Google Scholar] [CrossRef]
- Montiel-Rozas, M.M.; Madejón, E.; Madejón, P. Effect of heavy metals and organic matter on root exudates (low molecular weight organic acids) of herbaceous species: An assessment in sand and soil conditions under different levels of contamination. Environ. Pollut. 2016, 216, 273–281. [Google Scholar] [CrossRef] [Green Version]
- Martin, M.H.; Marschner, H. The Mineral Nutrition of Higher Plants. J. Ecol. 1988, 76, 1250. [Google Scholar] [CrossRef] [Green Version]
- Lombi, E.; Tearall, K.L.; Howarth, J.R.; Zhao, F.J.; Hawkesford, M.J.; McGrath, S.P. Influence of iron status on cadmium and zinc uptake by different ecotypes of the hyperaccumulator Thlaspi caerulescens. Plant Physiol. 2002, 128, 1359–1367. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Han, X.; Fang, J.; Lu, Z.; Qiu, W.; Liu, M.; Sang, J.; Jiang, J.; Zhuo, R. Sedum alfredii SaNramp6 Metal Transporter Contributes to Cadmium Accumulation in Transgenic Arabidopsis thaliana. Sci. Rep. 2017, 7, 13318. [Google Scholar] [CrossRef] [Green Version]
- Feng, S.; Tan, J.; Zhang, Y.; Liang, S.; Xiang, S.; Wang, H.; Chai, T. Isolation and characterization of a novel cadmium-regulated Yellow Stripe-Like transporter (SnYSL3) in Solanum nigrum. Plant Cell Rep. 2017, 36, 281–296. [Google Scholar] [CrossRef]
- Yruela, I. Copper in plants: Acquisition, transport and interactions. Funct. Plant Biol. 2009, 36, 409–430. [Google Scholar] [CrossRef] [Green Version]
- Puig, S. Function and Regulation of the Plant COPT Family of High-Affinity Copper Transport Proteins. Adv. Bot. 2014, 2014, 476917. [Google Scholar] [CrossRef] [Green Version]
- Andrés-Colás, N.; Sancenón, V.; Rodríguez-Navarro, S.; Mayo, S.; Thiele, D.J.; Ecker, J.R.; Puig, S.; Peñarrubia, L. The Arabidopsis heavy metal P-type ATPase HMA5 interacts with metallochaperones and functions in copper detoxification of roots. Plant J. 2006, 45, 225–236. [Google Scholar] [CrossRef]
- Dai, J.; Wang, N.; Xiong, H.; Qiu, W.; Nakanishi, H.; Kobayashi, T.; Nishizawa, N.K.; Zuo, Y. The yellow stripe-like (YSL) gene functions in internal copper transport in peanut. Genes 2018, 9, 635. [Google Scholar] [CrossRef] [Green Version]
- Uraguchi, S.; Mori, S.; Kuramata, M.; Kawasaki, A.; Arao, T.; Ishikawa, S. Root-to-shoot Cd translocation via the xylem is the major process determining shoot and grain cadmium accumulation in rice. J. Exp. Bot. 2009, 60, 2677–2688. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Guo, J.; Zhang, X.; Yu, H.; Huang, F.; Zhang, L.; Zhang, M.; Li, T. Changes of non-protein thiols in root and organic acids in xylem sap involved in cadmium translocation of cadmium-safe rice line (Oryza sativa L.). Plant Soil 2019, 439, 475–486. [Google Scholar] [CrossRef]
- Kerkeb, L.; Krämer, U. The role of free histidine in xylem loading of nickel in Alyssum lesbiacum and Brassica juncea. Plant Physiol. 2003, 131, 716–724. [Google Scholar] [CrossRef] [Green Version]
- Centofanti, T.; Sayers, Z.; Cabello-Conejo, M.I.; Kidd, P.; Nishizawa, N.K.; Kakei, Y.; Davis, A.P.; Sicher, R.C.; Chaney, R.L. Xylem exudate composition and root-to-shoot nickel translocation in Alyssum species. Plant Soil 2013, 373, 59–75. [Google Scholar] [CrossRef]
- Saathoff, A.J.; Ahner, B.; Spanswick, R.M.; Walker, L.P. Detection of Phytochelatin in the Xylem Sap of Brassica napus. Environ. Eng. Sci. 2011, 28, 103–111. [Google Scholar] [CrossRef]
- Wei, Z.G.; Wong, J.W.C.; Zhao, H.Y.; Zhang, H.J.; Li, H.X.; Hu, F. Separation and determination of heavy metals associated with low molecular weight chelators in xylem saps of Indian mustard (Brassica juncea) by size exclusion chromatography and atomic absorption spectrometry. Biol. Trace Elem. Res. 2007, 118, 146–158. [Google Scholar] [CrossRef]
- Yadav, S.K.K. Heavy metals toxicity in plants: An overview on the role of glutathione and phytochelatins in heavy metal stress tolerance of plants. S. Afr. J. Bot. 2010, 76, 167–179. [Google Scholar] [CrossRef] [Green Version]
- Dago, À.; González, I.; Ariño, C.; Martínez-Coronado, A.; Higueras, P.; Díaz-Cruz, J.M.; Esteban, M. Evaluation of mercury stress in plants from the Almadén mining district by analysis of phytochelatins and their Hg complexes. Environ. Sci. Technol. 2014, 48, 6256–6263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dago, A.; Ariño, C.; Díaz-Cruz, J.M.; Esteban, M. Analysis of phytochelatins and Hg-phytochelatin complexes in Hordeum vulgare plants stressed with Hg and Cd: HPLC study with amperometric detection. Int. J. Environ. Anal. Chem. 2014, 94, 668–678. [Google Scholar] [CrossRef]
- Heiss, S.; Wachter, A.; Bogs, J.; Cobbett, C.; Rausch, T. Phytochelatin synthase (PCS) protein is induced in Brassica juncea leaves after prolonged Cd exposure. J. Exp. Bot. 2003, 54, 1833–1839. [Google Scholar] [CrossRef]
- Liu, W.; Zhang, X.; Liang, L.; Chen, C.; Wei, S.; Zhou, Q. Phytochelatin and oxidative stress under heavy metal stress tolerance in plants. In Reactive Oxygen Species and Oxidative Damage in Plants under Stress; Springer International Publishing: Cham, Switzerland, 2015; pp. 191–217. ISBN 9783319204215. [Google Scholar]
- Grill, E.; Zenk, M.H.; Winnacker, E.-L. Induction of heavy metal-sequestering phytochelatin by cadmium in cell cultures of Rauvolfia serpentina. Naturwissenschaften 1985, 72, 432–433. [Google Scholar] [CrossRef]
- Klapheck, S.; Chrost, B.; Starke, J.; Zimmermann, H. γ-Glutamylcysteinylserine—A New Homologue of Glutathione in Plants of the Family Poaceae. Bot. Acta 1992, 105, 174–179. [Google Scholar] [CrossRef]
- Meuwly, P.; Thibault, P.; Rauser, W.E. γ-Glutamylcysteinylglutamic acid-a new homologue of glutathione in maize seedlings exposed to cadmium. FEBS Lett. 1993, 336, 472–476. [Google Scholar] [CrossRef] [Green Version]
- Kubota, H.; Sato, K.; Yamada, T.; Maitani, T. Phytochelatin homologs induced in hairy roots of horseradish. Phytochemistry 2000, 53, 239–245. [Google Scholar] [CrossRef]
- Sylwia, W.; Anna, R.; Ewa, B.; Stephan, C.; Danuta Maria, A. The role of subcellular distribution of cadmium and phytochelatins in the generation of distinct phenotypes of AtPCS1- and CePCS3-expressing tobacco. J. Plant Physiol. 2010, 167, 981–988. [Google Scholar] [CrossRef]
- Yang, X.E.; Li, T.Q.; Long, X.X.; Xiong, Y.H.; He, Z.L.; Stoffella, P.J. Dynamics of zinc uptake and accumulation in the hyperaccumulating and non-hyperaccumulating ecotypes of Sedum alfredii Hance. Plant Soil 2006, 284, 109–119. [Google Scholar] [CrossRef]
- Melo, E.E.C.; Costa, E.T.S.; Guilherme, L.R.G.; Faquin, V.; Nascimento, C.W.A. Accumulation of arsenic and nutrients by castor bean plants grown on an As-enriched nutrient solution. J. Hazard. Mater. 2009, 168, 479–483. [Google Scholar] [CrossRef]
- Pandey, S.; Fartyal, D.; Agarwal, A.; Shukla, T.; James, D.; Kaul, T.; Negi, Y.K.; Arora, S.; Reddy, M.K. Abiotic Stress Tolerance in Plants: Myriad Roles of Ascorbate Peroxidase. Front. Plant Sci. 2017, 8, 581. [Google Scholar] [CrossRef] [Green Version]
- Fuentes, A. Phytotoxicity and heavy metals speciation of stabilised sewage sludges. J. Hazard. Mater. 2004, 108, 161–169. [Google Scholar] [CrossRef]
- Sandalio, L.M.; Dalurzo, H.C.; Gómez, M.; Romero-Puertas, M.C.; del Río, L.A. Cadmium-induced changes in the growth and oxidative metabolism of pea plants. J. Exp. Bot. 2001, 52, 2115–2126. [Google Scholar] [CrossRef]
- Sharma, P.; Jha, A.B.; Dubey, R.S.; Pessarakli, M. Reactive Oxygen Species, Oxidative Damage, and Antioxidative Defense Mechanism in Plants under Stressful Conditions. J. Bot. 2012, 2012, 1–26. [Google Scholar] [CrossRef] [Green Version]
- Kalaivanan, D.; Ganeshamurthy, A.N. Mechanisms of Heavy Metal Toxicity in Plants. In Abiotic Stress Physiology of Horticultural Crops; Springer: New Delhi, India, 2016; pp. 85–102. [Google Scholar]
- Nagajyoti, P.C.; Lee, K.D.; Sreekanth, T.V.M. Heavy metals, occurrence and toxicity for plants: A review. Environ. Chem. Lett. 2010, 8, 199–216. [Google Scholar] [CrossRef]
- Manzoor, J.; Sharma, M.; Wani, K.A. Heavy metals in vegetables and their impact on the nutrient quality of vegetables: A review. J. Plant Nutr. 2018, 41, 1744–1763. [Google Scholar] [CrossRef]
- Sandalio, L.M.; Rodríguez-Serrano, M.; Gupta, D.K.; Archilla, A.; Romero-Puertas, M.C.; Del Río, L.A. Reactive Oxygen species and Nitric Oxide in plants under cadmium stress: From toxicity to signaling. In Environmental Adaptations and Stress Tolerance of Plants in the Era of Climate Change; Springer: New York, NY, USA, 2012; pp. 199–215. ISBN 9781461408154. [Google Scholar]
- Molina-Moya, E.; Terrón-Camero, L.C.; Pescador-Azofra, L.; Sandalio, L.M.; Romero-Puertas, M.C. Reactive Oxygen Species and Nitric Oxide Production, Regulation and Function During Defense Response. In Reactive Oxygen, Nitrogen and Sulfur Species in Plants; Wiley: Hoboken, NJ, USA, 2019; pp. 573–590. [Google Scholar]
- Romero-Puertas, M.C.; Terrón-Camero, L.C.; Peláez-Vico, M.Á.; Olmedilla, A.; Sandalio, L.M. Reactive oxygen and nitrogen species as key indicators of plant responses to Cd stress. Environ. Exp. Bot. 2019, 161, 107–119. [Google Scholar] [CrossRef]
- Gupta, A.S.; Webb, R.P.; Holaday, A.S.; Allen, R.D. Overexpression of superoxide dismutase protects plants from oxidative stress. Induction of ascorbate peroxidase in superoxide dismutase-overexpressing plants. Plant Physiol. 1993, 103, 1067–1073. [Google Scholar] [CrossRef] [Green Version]
- Guo, Q.; Meng, L.; Zhang, Y.-N.N.; Mao, P.-C.C.; Tian, X.-X.X.; Li, S.-S.S.; Zhang, L. Antioxidative systems, metal ion homeostasis and cadmium distribution in Iris lactea exposed to cadmium stress. Ecotoxicol. Environ. Saf. 2017, 139, 50–55. [Google Scholar] [CrossRef]
- Gong, Q.; Wang, L.; Dai, T.; Zhou, J.; Kang, Q.; Chen, H.; Li, K.; Li, Z. Effects of copper on the growth, antioxidant enzymes and photosynthesis of spinach seedlings. Ecotoxicol. Environ. Saf. 2019, 171, 771–780. [Google Scholar] [CrossRef]
- Xun, Y.; Feng, L.; Li, Y.; Dong, H. Mercury accumulation plant Cyrtomium macrophyllum and its potential for phytoremediation of mercury polluted sites. Chemosphere 2017, 189, 161–170. [Google Scholar] [CrossRef]
- Singh, S.; Sounderajan, S.; Kumar, K.; Fulzele, D.P. Investigation of arsenic accumulation and biochemical response of in vitro developed Vetiveria zizanoides plants. Ecotoxicol. Environ. Saf. 2017, 145, 50–56. [Google Scholar] [CrossRef]
- Ortega, A.; Garrido, I.; Casimiro, I.; Espinosa, F. Effects of antimony on redox activities and antioxidant defence systems in sunflower (Helianthus annuus L.) plants. PLoS ONE 2017, 12, e0183991. [Google Scholar] [CrossRef] [Green Version]
- Zaheer, I.E.; Ali, S.; Saleem, M.H.; Imran, M.; Alnusairi, G.S.H.; Alharbi, B.M.; Riaz, M.; Abbas, Z.; Rizwan, M.; Soliman, M.H. Role of iron-lysine on morpho-physiological traits and combating chromium toxicity in rapeseed (Brassica napus L.) plants irrigated with different levels of tannery wastewater. Plant Physiol. Biochem. 2020, 155, 70–84. [Google Scholar] [CrossRef]
- Singh, P.; Shukla, A.K.; Behera, S.K.; Tiwari, P.K. Zinc Application Enhances Superoxide Dismutase and Carbonic Anhydrase Activities in Zinc-Efficient and Zinc-Inefficient Wheat Genotypes. J. Soil Sci. Plant Nutr. 2019, 19, 477–487. [Google Scholar] [CrossRef]
- Zhu, Y.M.; Wu, Q.H.; Lv, H.Q.; Chen, W.X.; Wang, L.Z.; Shi, S.J.; Yang, J.G.; Zhao, P.P.; Li, Y.P.; Christopher, R.; et al. Toxicity of different forms of antimony to rice plants: Effects on reactive oxidative species production, antioxidative systems, and uptake of essential elements. Environ. Pollut. 2020, 263, 114544. [Google Scholar] [CrossRef]
- Pirzadah, T.B.; Malik, B.; Tahir, I.; Irfan, Q.M.; Rehman, R.U. Characterization of mercury-induced stress biomarkers in Fagopyrum tataricum plants. Int. J. Phytoremediat. 2018, 20, 225–236. [Google Scholar] [CrossRef]
- Corpas, F.J.; Barroso, J.B.; Sandalio, L.M.; Distefano, S.; Palma, J.M.; Lupiáñez, J.A.; Del Río, L.A. A dehydrogenase-mediated recycling system of NADPH in plant peroxisomes. Biochem. J. 1998, 330 Pt 2, 777–784. [Google Scholar] [CrossRef]
- Rodríguez-Ruiz, M.; Aparicio-Chacón, M.V.; Palma, J.M.; Corpas, F.J. Arsenate disrupts ion balance, sulfur and nitric oxide metabolisms in roots and leaves of pea (Pisum sativum L.) plants. Environ. Exp. Bot. 2019, 161, 143–156. [Google Scholar] [CrossRef]
- Kumar, S.; Trivedi, P.K. Glutathione S-transferases: Role in combating abiotic stresses including arsenic detoxification in plants. Front. Plant Sci. 2018, 9, 751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Namdjoyan, S.; Kermanian, H.; Abolhasani Soorki, A.; Modarres Tabatabaei, S.; Elyasi, N. Interactive effects of Salicylic acid and nitric oxide in alleviating zinc toxicity of Safflower (Carthamus tinctorius L.). Ecotoxicology 2017, 26, 752–761. [Google Scholar] [CrossRef] [PubMed]
- Handa, N.; Kohli, S.K.; Sharma, A.; Thukral, A.K.; Bhardwaj, R.; Abd_Allah, E.F.; Alqarawi, A.A.; Ahmad, P. Selenium modulates dynamics of antioxidative defence expression, photosynthetic attributes and secondary metabolites to mitigate chromium toxicity in Brassica juncea L. plants. Environ. Exp. Bot. 2019, 161, 180–192. [Google Scholar] [CrossRef]
- Noctor, G.; Mhamdi, A.; Chaouch, S.; Han, Y.; Neukermans, J.; Marquez-Garcia, B.; Queval, G.; Foyer, C.H. Glutathione in plants: An integrated overview. Plant. Cell Environ. 2012, 35, 454–484. [Google Scholar] [CrossRef]
- Terrón-Camero, L.C.; Peláez-Vico, M.Á.; Del-Val, C.; Sandalio, L.M.; Romero-Puertas, M.C.; Brouquisse, R. Role of nitric oxide in plant responses to heavy metal stress: Exogenous application versus endogenous production. J. Exp. Bot. 2019, 70, 4477–4488. [Google Scholar] [CrossRef]
- Maestri, E.; Marmiroli, N. Transgenic plants for phytoremediation. Int. J. Phytoremediat. 2011, 13, 264–279. [Google Scholar] [CrossRef]
- Cherian, S.; Oliveira, M.M. Transgenic plants in phytoremediation: Recent advances and new possibilities. Environ. Sci. Technol. 2005, 39, 9377–9390. [Google Scholar] [CrossRef]
- Gunarathne, V.; Mayakaduwa, S.; Ashiq, A.; Weerakoon, S.R.; Biswas, J.K.; Vithanage, M. Transgenic Plants: Benefits, Applications, and Potential Risks in Phytoremediation. Benefits, Applications, and Potential Risks in Phytoremediation. In Transgenic Plant Technology for Remediation of Toxic Metals and Metalloids; Elsevier: Amsterdam, The Netherlands, 2018; pp. 89–102. ISBN 9780128143902. [Google Scholar]
- Liu, D.; An, Z.; Mao, Z.; Ma, L.; Lu, Z. Enhanced Heavy Metal Tolerance and Accumulation by Transgenic Sugar Beets Expressing Streptococcus thermophilus StGCS-GS in the Presence of Cd, Zn and Cu Alone or in Combination. PLoS ONE 2015, 10, e0128824. [Google Scholar] [CrossRef]
- Liu, Y.; Kang, T.; Cheng, J.S.; Yi, Y.J.; Han, J.J.; Cheng, H.L.; Li, Q.; Tang, N.; Liang, M.X. Heterologous expression of the metallothionein PpMT2 gene from Physcomitrella patens confers enhanced tolerance to heavy metal stress on transgenic Arabidopsis plants. Plant Growth Regul. 2020, 90, 63–72. [Google Scholar] [CrossRef]
- Evans, D.M.; Zipper, C.E.; Burger, J.A.; Strahm, B.D.; Villamagna, A.M. Reforestation practice for enhancement of ecosystem services on a compacted surface mine: Path toward ecosystem recovery. Ecol. Eng. 2013, 51, 16–23. [Google Scholar] [CrossRef]
- Madejón, P.; Domínguez, M.T.; Madejón, E.; Cabrera, F.; Marañón, T.; Murillo, J.M. Soil-plant relationships and contamination by trace elements: A review of twenty years of experimentation and monitoring after the Aznalcóllar (SW Spain) mine accident. Sci. Total Environ. 2018, 625, 50–63. [Google Scholar] [CrossRef] [Green Version]
- Del Río, M.; Font, R.; Almela, C.; Vélez, D.; Montoro, R.; De Haro Bailón, A. Heavy metals and arsenic uptake by wild vegetation in the Guadiamar river area after the toxic spill of the Aznalcóllar mine. In Proceedings of the Journal of Biotechnology. J. Biotechnol. 2002, 98, 125–137. [Google Scholar] [CrossRef]
- Madejón, P.; Murillo, J.M.; Marañón, T.; Cabrera, F. Bioaccumulation of trace elements in a wild grass three years after the Aznalcóllar mine spill (South Spain). Environ. Monit. Assess. 2006, 114, 169–189. [Google Scholar] [CrossRef]
- Madejón, P.; Murillo, J.M.; Marañón, T.; Lepp, N.W. Factors affecting accumulation of thallium and other trace elements in two wild Brassicaceae spontaneously growing on soils contaminated by tailings dam waste. Chemosphere 2007, 67, 20–28. [Google Scholar] [CrossRef]
- Murillo, J.M.; Marañón, T.; Cabrera, F.; López, R. Accumulation of heavy metals in sunflower and sorghum plants affected by the Guadiamar spill. Sci. Total Environ. 1999, 242, 281–292. [Google Scholar] [CrossRef]
- Madejón, P.; Murillo, J.M.; Marañón, T.; Cabrera, F.; Soriano, M.A. Trace element and nutrient accumulation in sunflower plants two years after the Aznalcóllar mine spill. Sci. Total Environ. 2003, 307, 239–257. [Google Scholar] [CrossRef]
- Clemente, R.; Walker, D.J.; Bernal, M.P. Uptake of heavy metals and As by Brassica juncea grown in a contaminated soil in Aznalcóllar (Spain): The effect of soil amendments. Environ. Pollut. 2005, 138, 46–58. [Google Scholar] [CrossRef]
- Soriano, M.A.; Fereres, E. Use of crops for in situ phytoremediation of polluted soils following a toxic flood from a mine spill. Plant Soil 2003, 256, 253–264. [Google Scholar] [CrossRef]
- Vázquez, S.; Agha, R.; Granado, A.; Sarro, M.J.; Esteban, E.; Peñalosa, J.M.; Carpena, R.O. Use of white lupin plant for phytostabilization of Cd and as polluted acid soil. Water Air Soil Pollut. 2006, 177, 349–365. [Google Scholar] [CrossRef]
- Moreno-Jiménez, E.; Vázquez, S.; Carpena-Ruiz, R.O.; Esteban, E.; Peñalosa, J.M. Using Mediterranean shrubs for the phytoremediation of a soil impacted by pyritic wastes in Southern Spain: A field experiment. J. Environ. Manag. 2011, 92, 1584–1590. [Google Scholar] [CrossRef] [Green Version]
- Domínguez, M.T.; Marañón, T.; Murillo, J.M.; Schulin, R.; Robinson, B.H. Trace element accumulation in woody plants of the Guadiamar Valley, SW Spain: A large-scale phytomanagement case study. Environ. Pollut. 2008, 152, 50–59. [Google Scholar] [CrossRef] [Green Version]
- Domínguez, M.T.; Madejón, P.; Marañón, T.; Murillo, J.M. Avorestation of a trace-element polluted area in SW Spain: Woody plant performance and trace element accumulation. Eur. J. For. Res. 2010, 129, 47–59. [Google Scholar] [CrossRef]
- Domínguez, M.T.; Pérez-Ramos, I.M.; Murillo, J.M.; Marañón, T. Facilitating the afforestation of Mediterranean polluted soils by nurse shrubs. J. Environ. Manag. 2015, 161, 276–286. [Google Scholar] [CrossRef] [Green Version]
- De la Fuente, C.; Pardo, T.; Alburquerque, J.A.; Martínez-Alcalá, I.; Bernal, M.P.; Clemente, R. Assessment of native shrubs for stabilisation of a trace elements-polluted soil as the final phase of a restoration process. Agric. Ecosyst. Environ. 2014, 196, 103–111. [Google Scholar] [CrossRef]
- Domínguez, M.T.; Madrid, F.; Marañón, T.; Murillo, J.M. Cadmium availability in soil and retention in oak roots: Potential for phytostabilization. Chemosphere 2009, 76, 480–486. [Google Scholar] [CrossRef]
- Moreno-Jiménez, E.; Peñalosa, J.M.; Carpena-Ruiz, R.O.; Esteban, E. Comparison of arsenic resistance in Mediterranean woody shrubs used in restoration activities. Chemosphere 2008, 71, 466–473. [Google Scholar] [CrossRef]
- Martín-Crespo, T.; Gómez-Ortiz, D.; Martín-Velázquez, S.; Esbrí, J.M.; de Ignacio-San José, C.; Sánchez-García, M.J.; Montoya-Montes, I.; Martín-González, F. Abandoned mine tailings in cultural itineraries: Don Quixote Route (Spain). Eng. Geol. 2015, 197, 82–93. [Google Scholar] [CrossRef]
- Asensio, V.; Vega, F.A.; Singh, B.R.; Covelo, E.F. Effects of tree vegetation and waste amendments on the fractionation of Cr, Cu, Ni, Pb and Zn in polluted mine soils. Sci. Total Environ. 2013, 443, 446–453. [Google Scholar] [CrossRef] [PubMed]
- Pardo, T.; Bernal, M.P.; Clemente, R. Phytostabilisation of severely contaminated mine tailings using halophytes and field addition of organic and inorganic amendments. Chemosphere 2017, 178, 556–564. [Google Scholar] [CrossRef]
- Hernández, A.; Jébrak, M.; Higueras, P.; Oyarzun, R.; Morata, D.; Munhá, J. The Almaden mercury mining district, Spain. Miner. Depos. 1999, 34, 539–548. [Google Scholar] [CrossRef]
- Rodriguez, L.; Lopez-Bellido, F.J.; Carnicer, A.; Recreo, F.; Tallos, A.; Monteagudo, J.M. Mercury recovery from soils by phytoremediation. In Environmental Chemistry: Green Chemistry and Pollutants in Ecosystems; Springer: Berlin/Heidelberg, Germany, 2005; pp. 197–204. ISBN 3540228608. [Google Scholar]
- Naharro, R.; Esbrí, J.M.; Amorós, J.Á.; García-Navarro, F.J.; Higueras, P. Assessment of mercury uptake routes at the soil-plant-atmosphere interface. Geochem. Explor. Environ. Anal. 2018, 19, 146–154. [Google Scholar] [CrossRef]
- Pratas, J.; Prasad, M.N.V.; Freitas, H.; Conde, L. Plants growing in abandoned mines of Portugal are useful for biogeochemical exploration of arsenic, antimony, tungsten and mine reclamation. J. Geochem. Explor. 2005, 85, 99–107. [Google Scholar] [CrossRef] [Green Version]
- Simiele, M.; Lebrun, M.; Miard, F.; Trupiano, D.; Poupart, P.; Forestier, O.; Scippa, G.S.; Bourgerie, S.; Morabito, D. Assisted phytoremediation of a former mine soil using biochar and iron sulphate: Effects on As soil immobilization and accumulation in three Salicaceae species. Sci. Total Environ. 2020, 710, 136203. [Google Scholar] [CrossRef] [PubMed]
- Bacchetta, G.; Cao, A.; Cappai, G.; Carucci, A.; Casti, M.; Fercia, M.L.; Lonis, R.; Mola, F. A field experiment on the use of Pistacia lentiscus L. and Scrophularia canina L. subsp. bicolor (Sibth. et Sm.) Greuter for the phytoremediation of abandoned mining areas. Plant Biosyst. 2012, 146, 1054–1063. [Google Scholar] [CrossRef]
- Swab, R.M.; Lorenz, N.; Byrd, S.; Dick, R. Native vegetation in reclamation: Improving habitat and ecosystem function through using prairie species in mine land reclamation. Ecol. Eng. 2017, 108, 525–536. [Google Scholar] [CrossRef]
- Gil-Loaiza, J.; White, S.A.; Root, R.A.; Solís-Dominguez, F.A.; Hammond, C.M.; Chorover, J.; Maier, R.M. Phytostabilization of mine tailings using compost-assisted direct planting: Translating greenhouse results to the field. Sci. Total Environ. 2016, 565, 451–461. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Z.; Shahrour, I.; Bai, Z.; Fan, W.; Feng, L.; Li, H. Soils development in opencast coal mine spoils reclaimed for 1–13 years in the West-Northern Loess Plateau of China. Eur. J. Soil Biol. 2013, 55, 40–46. [Google Scholar] [CrossRef]
- Zhang, P.; Yang, F.; Lan, M.; Liu, W.; Yang, W.; Tang, Y.; Qiu, R. Phytostabilization with tolerant plants and soil amendments of the tailings of the Dabaoshan polymetallic mine in Guangdong Province. Huanjing Kexue Xuebao/Acta Sci. Circumstantiae 2019, 39, 545–552. [Google Scholar]
- Tai, Y.P.; Yang, Y.F.; Li, Z.A.; Yang, Y.; Wang, J.X.; Zhuang, P.; Zou, B. Phytoextraction of 55-year-old wastewater-irrigated soil in a Zn-Pb mine district: Effect of plant species and chelators. Environ. Technol. 2018, 39, 2138–2150. [Google Scholar] [CrossRef]
- Li, X.; Wang, X.; Chen, Y.; Yang, X.; Cui, Z. Optimization of combined phytoremediation for heavy metal contaminated mine tailings by a field-scale orthogonal experiment. Ecotoxicol. Environ. Saf. 2019, 168, 1–8. [Google Scholar] [CrossRef]
- Yu, G.; Jiang, P.; Fu, X.; Liu, J.; Sunahara, G.I.; Chen, Z.; Xiao, H.; Lin, F.; Wang, X. Phytoextraction of cadmium-contaminated soil by Celosia argentea Linn.: A long-term field study. Environ. Pollut. 2020, 266, 115408. [Google Scholar] [CrossRef]
- Yuan, X.; Xiong, T.; Yao, S.; Liu, C.; Yin, Y.; Li, H.; Li, N. A real filed phytoremediation of multi-metals contaminated soils by selected hybrid sweet sorghum with high biomass and high accumulation ability. Chemosphere 2019, 237, 124536. [Google Scholar] [CrossRef]
- Shen, S.; Chen, J.; Chang, J.; Xia, B. Using bioenergy crop cassava (Manihot esculenta) for reclamation of heavily metal-contaminated land. Int. J. Phytoremediat. 2020, 22, 1313–1320. [Google Scholar] [CrossRef]
- Shu, W.S.; Xia, H.P.; Zhang, Z.Q.; Lan, C.Y.; Wong, M.H. Use of vetiver and three other grasses for revegetation of Pb/Zn mine tailings: Field experiment. Int. J. Phytoremediat. 2002, 4, 47–57. [Google Scholar] [CrossRef]
- Liu, Y.; Zhong, X.; Huot, H.; Liu, W.; Liu, C.; Guo, M.; Li, Y.; Fei, Y.; Chao, Y.; Wang, S.; et al. Reclamation with organic amendments and plants remodels the diversity and structure of bacterial community in ion-adsorption rare earth element mine tailings. J. Soils Sediments 2020, 20, 3669–3680. [Google Scholar] [CrossRef]
- Mi, J.; Hou, H.; Raval, S.; Yang, Y.; Zhang, S.; Hua, Y.; Wang, C.; Chen, F. Effect of crop cultivation on the soil carbon stock in mine dumps of the Loess Plateau, China. Sci. Total Environ. 2020, 741, 139809. [Google Scholar] [CrossRef]
- Chen, F.; Yang, Y.; Mi, J.; Liu, R.; Hou, H.; Zhang, S. Effects of vegetation pattern and spontaneous succession on remediation of potential toxic metal-polluted soil in mine dumps. Sustainability 2019, 11, 397. [Google Scholar] [CrossRef] [Green Version]
- Singh, A.K. Toxic aqueous discharge of iron and sulphur from spoiled coal mined lands and its control by phytostabilization process. Curr. Sci. 2018, 115, 529–534. [Google Scholar] [CrossRef]
- Kamble, S.S.; Dod, R.D. Management of coal mine overburden and fly ash using bamboo cultivation. Int. J. Innov. Technol. Explor. Eng. 2019, 8, 4223–4226. [Google Scholar]
- Jeżowski, S.; Mos, M.; Buckby, S.; Cerazy-Waliszewska, J.; Owczarzak, W.; Mocek, A.; Kaczmarek, Z.; McCalmont, J.P. Establishment, growth, and yield potential of the perennial grass Miscanthus × Giganteus on degraded coal mine soils. Front. Plant Sci. 2017, 8, 726. [Google Scholar] [CrossRef] [Green Version]
- Bandyopadhyay, S.; Rana, V.; Maiti, S.K. Chronological Variation of Metals in Reclaimed Coal Mine Soil and Tissues of Eucalyptus Hybrid Tree After 25 Years of Reclamation, Jharia Coal Field (India). Bull. Environ. Contam. Toxicol. 2018, 101, 604–610. [Google Scholar] [CrossRef]
- Pavlova, D.; Echevarria, G.; Mullaj, A.; Reeves, R.D.; Morel, J.-L.; Sulçe, S. Nickel hyperaccumulation by the species of Alyssum and Thlaspi (Brassicaceae) from the ultramafic soils of the Balkans. Bot. Serbica 2010, 34, 3–10. [Google Scholar]
- Küpper, H.; Lombi, E.; Zhao, F.J.; McGrath, S.P. Cellular compartmentation of cadmium and zinc in relation to other elements in the hyperaccumulator Arabidopsis halleri. Planta 2000, 212, 75–84. [Google Scholar] [CrossRef] [Green Version]
- Kubota, H.; Takenaka, C. Arabis gemmifera is a Hyperaccumulator of Cd and Zn. Int. J. Phytoremediat. 2003, 5, 197–201. [Google Scholar] [CrossRef]
- Mesnoua, M.; Mateos-Naranjo, E.; Barcia-Piedras, J.M.; Pérez-Romero, J.A.; Lotmani, B.; Redondo-Gómez, S. Physiological and biochemical mechanisms preventing Cd-toxicity in the hyperaccumulator Atriplex halimus L. Plant Physiol. Biochem. 2016, 106, 30–38. [Google Scholar] [CrossRef]
- Ugustyniak, M.; Arnawska, M.; Eimold, W. Uptake of cadmium, lead, nickel and zinc from soil and water solutions by the nickel hyperaccumulator. Acta Biol. Cracoviensia 2004, 46, 75–85. [Google Scholar]
- Sun, Y.; Zhou, Q.; Wang, L.; Liu, W. Cadmium tolerance and accumulation characteristics of Bidens pilosa L. as a potential Cd-hyperaccumulator. J. Hazard. Mater. 2009, 161, 808–814. [Google Scholar] [CrossRef]
- Wierzbicka, M.; Pielichowska, M.; Abratowska, A.; WiŁkomirski, B.; Wysocka, I.; Panufnik-Mędrzycka, D.; Bulska, E.; Panufnik-Mȩdrzycka, D.; Bulska, E. Thallium Hyperaccumulation in Polish Populations of Biscutella laevigata (Brassicaceae). Acta Biol. Cracoviensia Ser. Bot. 2016, 58, 7–19. [Google Scholar] [CrossRef] [Green Version]
- Xiong, Z.T. Lead uptake and effects on seed germination and plant growth in a Pb hyperaccumulator Brassica pekinensis Rupr. Bull. Environ. Contam. Toxicol. 1998, 60, 285–291. [Google Scholar] [CrossRef]
- Liu, W.; Zhang, C.; Hu, P.; Luo, Y.; Wu, L.; Sale, P.; Tang, C. Influence of nitrogen form on the phytoextraction of cadmium by a newly discovered hyperaccumulator Carpobrotus rossii. Environ. Sci. Pollut. Res. 2016, 23, 1246–1253. [Google Scholar] [CrossRef]
- Zhang, X.H.; Liu, J.; Huang, H.T.; Chen, J.; Zhu, Y.N.; Wang, D.Q. Chromium accumulation by the hyperaccumulator plant Leersia hexandra Swartz. Chemosphere 2007, 67, 1138–1143. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; He, X.; Chen, W.; Yuan, F.; Yan, K.; Tao, D. Accumulation and tolerance characteristics of cadmium in a potential hyperaccumulator—Lonicera japonica Thunb. J. Hazard. Mater. 2009, 169, 170–175. [Google Scholar] [CrossRef] [PubMed]
- Xue, S.G.; Chen, Y.X.; Reeves, R.D.; Baker, A.J.; Lin, Q.; Fernando, D.R. Manganese uptake and accumulation by the hyperaccumulator plant Phytolacca acinosa Roxb. (Phytolaccaceae). Environ. Pollut. 2004, 131, 393–399. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Yu, F.; Chen, M.; Zhou, Z.; Chen, C.; Li, M.S.; Zhu, J. A newly found manganese hyperaccumulator—Polygonum lapathifolium Linn. Int. J. Phytoremediat. 2016, 18, 348–353. [Google Scholar] [CrossRef]
- Chen, T.; Wei, C.; Huang, Z.; Huang, Q.; Lu, Q.; Fan, Z. Arsenic hyperaccumulator Pteris vittata L. and its arsenic accumulation. Chin. Sci. Bull. 2002, 47, 902–905. [Google Scholar] [CrossRef]
- Li, M.S.; Yang, S.X. Heavy metal contamination in soils and phytoaccumulation in a manganese mine Wasteland, South China. Air Soil Water Res. 2008, 1, 31–41. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.X.; Deng, H.; Li, M.S. Manganese uptake and accumulation in a woody hyperaccumulator, Schima superba. Plant Soil Environ. 2008, 54, 441–446. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.E.; Long, X.X.; Ye, H.B.; He, Z.L.; Calvert, D.V.; Stoffella, P.J. Cadmium tolerance and hyperaccumulation in a new Zn-hyperaccumulating plant species (Sedum alfredii Hance). Plant Soil 2004, 259, 181–189. [Google Scholar] [CrossRef]
- Yang, X.; Long, X.; Ni, W.; Fu, C. Sedum alfredii H: A new Zn hyperaccumulating plant first found in China. Chin. Sci. Bull. 2002, 47, 1634–1637. [Google Scholar] [CrossRef]
- Sahi, S.V.; Bryant, N.L.; Sharma, N.C.; Singh, S.R. Characterization of a lead hyperaccumulator shrub, Sesbania drummondii. Environ. Sci. Technol. 2002, 36, 4676–4680. [Google Scholar] [CrossRef]
- Gao, Y.; Zhou, P.; Mao, L.; Shi, W.J.; Zhi, Y.E. Phytoextraction of cadmium and physiological changes in Solanum nigrum as a novel cadmium hyperaccumulator. Russ. J. Plant Physiol. 2010, 57, 501–508. [Google Scholar] [CrossRef]
- Baker, A.J.M.; Reeves, R.D.; Hajar, A.S.M. Heavy metal accumulation and tolerance in British populations of the metallophyte Thlaspi caerulescens J. & C. Presl (Brassicaceae). New Phytol. 1994, 127, 61–68. [Google Scholar]
- Sakakibara, M.; Watanabe, A.; Inoue, M.; Sano, S.; Kaise, T. Phytoextraction and phytovolatilization of arsenic from As-contaminated soils by Pteris vittata. In Proceedings of the Annual International Conference on Soils, Sediments, Water and Energy, Amherst, MA, USA, 16–19 October 2010; Volume 12. [Google Scholar]
- Verma, S.; Verma, P.K.; Pande, V.; Tripathi, R.D.; Chakrabarty, D. Transgenic Arabidopsis thaliana expressing fungal arsenic methyltransferase gene (WaarsM) showed enhanced arsenic tolerance via volatilization. Environ. Exp. Bot. 2016, 132, 113–120. [Google Scholar] [CrossRef]
- Guarino, F.; Conte, B.; Improta, G.; Sciarrillo, R.; Castiglione, S.; Cicatelli, A.; Guarino, C. Genetic characterization, micropropagation, and potential use for arsenic phytoremediation of Dittrichia viscosa (L.) Greuter. Ecotoxicol. Environ. Saf. 2018, 148, 675–683. [Google Scholar] [CrossRef]
- Jia, Y.; Huang, H.; Sun, G.X.; Zhao, F.J.; Zhu, Y.G. Pathways and relative contributions to arsenic volatilization from rice plants and paddy soil. Environ. Sci. Technol. 2012, 46, 8090–8096. [Google Scholar] [CrossRef]
- Hussein, H.S.; Ruiz, O.N.; Terry, N.; Daniell, H. Phytoremediation of mercury and organomercurials in chloroplast transgenic plants: Enhanced root uptake, translocation to shoots, and volatilization. Environ. Sci. Technol. 2007, 41, 8439–8446. [Google Scholar] [CrossRef] [Green Version]
- Cabrita, M.T.; Duarte, B.; Cesário, R.; Mendes, R.; Hintelmann, H.; Eckey, K.; Dimock, B.; Caçador, I.; Canário, J. Mercury mobility and effects in the salt-marsh plant Halimione portulacoides: Uptake, transport, and toxicity and tolerance mechanisms. Sci. Total Environ. 2019, 650, 111–120. [Google Scholar] [CrossRef]
- Battke, F.; Ernst, D.; Fleischmann, F.; Halbach, S. Phytoreduction and volatilization of mercury by ascorbate in Arabidopsis thaliana, European beech and Norway spruce. Appl. Geochemistry 2008, 23, 494–502. [Google Scholar] [CrossRef]
- De Souza, M.P.; Chu, D.; Zhao, M.; Zayed, A.M.; Ruzin, S.E.; Schichnes, D.; Terry, N. Rhizosphere bacteria enhance selenium accumulation and volatilization by indian mustard. Plant Physiol. 1999, 119, 565–573. [Google Scholar] [CrossRef] [Green Version]
- Terry, N.; Carlson, C.; Raab, T.K.; Zayed, A.M. Rates of Selenium Volatilization among Crop Species. J. Environ. Qual. 1992, 21, 341–344. [Google Scholar] [CrossRef]
- Pilon-Smits, E.A.H.; De Souza, M.P.; Lytle, C.M.; Shang, C.; Lugo, T.; Terry, N. Selenium volatilization and assimilation by hybrid poplar (Populus tremula x alba). J. Exp. Bot. 1998, 49, 1889–1892. [Google Scholar] [CrossRef] [Green Version]
- Parker, D.R.; Feist, L.J.; Varvel, T.W.; Thomason, D.N.; Zhang, Y. Selenium phytoremediation potential of Stanleya pinnata. Plant Soil 2003, 249, 157–165. [Google Scholar] [CrossRef]
- Zieve, R.; Peterson, P.J. Volatilization of selenium from plants and soils. Sci. Total Environ. 1984, 32, 197–202. [Google Scholar] [CrossRef]
- Shackira, A.M.; Puthur, J.T. Enhanced phytostabilization of cadmium by a halophyte—Acanthus ilicifolius L. Int. J. Phytoremediat. 2017, 19, 319–326. [Google Scholar] [CrossRef]
- Nandillon, R.; Lebrun, M.; Miard, F.; Gaillard, M.; Sabatier, S.; Morabito, D.; Bourgerie, S. Contrasted tolerance of Agrostis capillaris metallicolous and non-metallicolous ecotypes in the context of a mining technosol amended by biochar, compost and iron sulfate. Environ. Geochem. Health 2019, 43, 1457–1475. [Google Scholar] [CrossRef] [PubMed]
- Cristaldi, A.; Oliveri Conti, G.; Cosentino, S.L.; Mauromicale, G.; Copat, C.; Grasso, A.; Zuccarello, P.; Fiore, M.; Restuccia, C.; Ferrante, M. Phytoremediation potential of Arundo donax (Giant Reed) in contaminated soil by heavy metals. Environ. Res. 2020, 185, 109427. [Google Scholar] [CrossRef]
- Visconti, D.; Álvarez-Robles, M.J.; Fiorentino, N.; Fagnano, M.; Clemente, R. Use of Brassica juncea and Dactylis glomerata for the phytostabilization of mine soils amended with compost or biochar. Chemosphere 2020, 260, 127661. [Google Scholar] [CrossRef]
- Nadgórska-Socha, A.; Ptasiński, B.; Kita, A. Heavy metal bioaccumulation and antioxidative responses in Cardaminopsis arenosa and Plantago lanceolata leaves from metalliferous and non-metalliferous sites: A field study. Ecotoxicology 2013, 22, 1422–1434. [Google Scholar] [CrossRef] [Green Version]
- Hussain, S.; Akram, M.; Abbas, G.; Murtaza, B.; Shahid, M.; Shah, N.S.; Bibi, I.; Niazi, N.K. Arsenic tolerance and phytoremediation potential of Conocarpus erectus L. and Populus deltoides L. Int. J. Phytoremediat. 2017, 19, 985–991. [Google Scholar] [CrossRef]
- Raza, A.; Khan, A.H.A.; Nawaz, I.; Qu, Z.; Yousaf, S.; Ali, M.A.; Sayal, A.U.; Iqbal, M. Evaluation of Arsenic-Induced Stress in Dahlia pinnata Cav.: Morphological and Physiological Response. Soil Sediment Contam. 2019, 28, 716–728. [Google Scholar] [CrossRef]
- Meier, S.; Alvear, M.; Borie, F.; Aguilera, P.; Ginocchio, R.; Cornejo, P. Influence of copper on root exudate patterns in some metallophytes and agricultural plants. Ecotoxicol. Environ. Saf. 2012, 75, 8–15. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Wang, S.; Thangavel, P.; Li, Q.; Zheng, H.; Bai, J.; Qiu, R. Phytostabilization potential of Jatropha curcas L. in polymetallic acid mine tailings. Int. J. Phytoremediat. 2011, 13, 788–804. [Google Scholar] [CrossRef] [PubMed]
- Santibáñez, C.; Verdugo, C.; Ginocchio, R. Phytostabilization of copper mine tailings with biosolids: Implications for metal uptake and productivity of Lolium perenne. Sci. Total Environ. 2008, 395, 1–10. [Google Scholar] [CrossRef] [PubMed]
- García, S.; Zornoza, P.; Hernández, L.E.; Esteban, E.; Carpena, R.O. Response of Lupinus albus to Pb-EDTA indicates relatively high tolerance. Toxicol. Environ. Chem. 2017, 99, 1378–1388. [Google Scholar] [CrossRef]
- Zhang, J.; Yang, S.; Huang, Y.; Zhou, S. The Tolerance and Accumulation of Miscanthus Sacchariflorus (maxim.) Benth., an Energy Plant Species, to Cadmium. Int. J. Phytoremediat. 2015, 17, 538–545. [Google Scholar] [CrossRef]
- Pavel, P.B.; Puschenreiter, M.; Wenzel, W.W.; Diacu, E.; Barbu, C.H. Aided phytostabilization using Miscanthus sinensis×giganteus on heavy metal-contaminated soils. Sci. Total Environ. 2014, 479–480, 125–131. [Google Scholar] [CrossRef]
- Lee, S.H.; Ji, W.H.; Lee, W.S.; Koo, N.; Koh, I.H.; Kim, M.S.; Park, J.S. Influence of amendments and aided phytostabilization on metal availability and mobility in Pb/Zn mine tailings. J. Environ. Manag. 2014, 139, 15–21. [Google Scholar] [CrossRef]
- Kiran, B.R.; Prasad, M.N.V. Assisted phytostabilization of Pb-spiked soils amended with charcoal and banana compost and vegetated with Ricinus communis L. (Castor bean). Environ. Geochem. Health 2021, 43, 1507–1521. [Google Scholar] [CrossRef]
- Varun, M.; Souza, R.D.; Pratas, J.; Paul, M.S.; De Ciências, D.; Coimbra, U. De Evaluation of phytostabilization, a green technology to remove heavy metals from industrial sludge using Typha latifolia L. Biotechnol. Bioinf. Bioeng. 2011, 1, 137–145. [Google Scholar]
Figure 1.
Principal mechanisms involved in phytoremediation.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Peco, J.D.; Higueras, P.; Campos, J.A.; Esbrí, J.M.; Moreno, M.M.; Battaglia-Brunet, F.; Sandalio, L.M. Abandoned Mine Lands Reclamation by Plant Remediation Technologies. Sustainability 2021, 13, 6555. https://doi.org/10.3390/su13126555
AMA Style
Peco JD, Higueras P, Campos JA, Esbrí JM, Moreno MM, Battaglia-Brunet F, Sandalio LM. Abandoned Mine Lands Reclamation by Plant Remediation Technologies. Sustainability. 2021; 13(12):6555. https://doi.org/10.3390/su13126555
Chicago/Turabian StylePeco, Jesús D., Pablo Higueras, Juan A. Campos, José M. Esbrí, Marta M. Moreno, Fabienne Battaglia-Brunet, and Luisa M. Sandalio. 2021. "Abandoned Mine Lands Reclamation by Plant Remediation Technologies" Sustainability 13, no. 12: 6555. https://doi.org/10.3390/su13126555
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.