Evaluation of Brassicaceae Seedlings as Trap Plants for Bagrada Hilaris Burmeister in Caper Bush Cultivations




1
Department of Agriculture, Food and Forest Sciences, University of Palermo, Viale delle Scienze, Building 5, 90128 Palermo, Italy
2
Institute of Biosciences and Bioresources (IBBR), National Research Council of Italy (CNR), Corso Calatafimi 414, 90129 Palermo, Italy
*
Author to whom correspondence should be addressed.
Sustainability 2020, 12(16), 6361; https://doi.org/10.3390/su12166361
Submission received: 27 May 2020
/
Revised: 2 August 2020
/
Accepted: 3 August 2020
/
Published: 7 August 2020
(This article belongs to the Special Issue Sustainable Defense Strategies for Pest Management)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:The caper bush, Capparis spinosa (Brassicales: Capparaceae), is intensively grown on Pantelleria Island (Trapani, Sicily, Sicilian channel) where it has been granted protected geographical indication (PGI) by the EU. On this island, Bagrada hilaris, a stink bug native of Asia and Africa, is the major pest of caper crops. Recent studies have shown the attraction of B. hilaris to volatiles of brassicaceous plants at the seedling stage. The objective of this study was to evaluate three cotyledon-stage seedlings of host plants, Brassica oleracea var. botrytis (cauliflower), Eruca sativa (rocket) and Brassica carinata (Abyssinian cabbage), as potential trap plants for B. hilaris. The relative preferences of these species were first evaluated in laboratory and field experiments, carried out during summer when the level of B. hilaris infestation was the highest. Behavioral bioassays in the laboratory conditions showed that adults of B. hilaris preferred to orient toward seedlings of B. oleracea and E. sativa over B. carinata. Field experiments confirmed these results. Then seedlings were tested in trap plant trials, by sowing them in artificial pots formed with aluminum trays and placing them in caper fields infested with B. hilaris. Results showed that E. sativa and B. oleracea diverted hundreds of B. hilaris individuals from the capers to these sources of attraction. Overall, these results suggest that B. oleracea and E. sativa seedlings used as lure inside traps or as trap plants may be a useful tool in the management of B. hilaris populations.
1. Introduction
Capparis spinosa L. (Brassicales: Capparaceae), also known as the caper bush, is a perennial shrub cultivated mainly for the flower buds known as capers, used for human consumption as spices [1,2]. Caper bush is a xerophilous plant able to develop and survive in environments with dry summer climates, typical of the islands of the Mediterranean Basin [3]. In Italy, capers are cultivated in Sicily and its minor islands where the positive use of marginal lands allows it, and has higher profit margins compared to other local crops [4]. In particular, caper cultivation on Pantelleria island has been granted protected geographical indication (Commission Regulation No. 1107/96 - EU) and produces 300–400 tons per year [5]. Caper plants are attacked by a large group of phytophagous insects including Capparimya savastanoi (Martelli) (Diptera: Tephritidae), Asphondylia gennadii (Marchal) (Diptera: Cecydomidae) and several pentatomid bugs (Heteroptera: Pentatomidae) such as Bagrada hilaris (Burmeister), Antheminia lunulata (Goeze), Eurydema ornata L., Eurydema ventralis Kol., Holcostethus punctatus (Lindberg) and Nezara viridula L. [4].
Among these pentatomids, the painted bug B. hilaris is considered the key pest of caper bush plants on Pantelleria Island [5]. After its accidental introduction in 1978 [6], this species progressively extended its area of diffusion on the island and caused increasing economic damages to the growers. The plant is injured by the feeding activity of adults and nymphs, which with their pierce-sucking mouthparts cause necrotic spots that compromise the plant’s photosynthetic activity. The injuries on the flower buds determine deformations that negatively affect their marketability [4,5]. Furthermore, B. hilaris can form large aggregations of individuals on host plants, in some cases causing the death of the plant itself [5]. To keep populations of B. hilaris below the threshold of economic damage, in the past, caper growers have relied on the use of chemical insecticides with four to five applications per year sprayed after each harvest [5]. Finally, the management of this species is also compromised by the lack of effective sampling strategies and appropriate monitoring tools, for example based on semiochemicals that still have not been developed [7]. In this context, new tools for controlling the B. hilaris population are urgently necessary in order to reduce treatments costs, have pesticide-free products and give caper growers the opportunity to switch to organic agriculture.
Among the possible alternative strategies for managing invasive insect pests, in recent years the use of trap plants or trap crops has been receiving attention as having low environmental impact and being useful in organic farming contexts [8,9]. Trap cropping is a technique based usually on the use of host plants that are independently deployed to attract, divert, intercept, and/or retain targeted insects that are successively treated or destroyed [10]. Trap crops have been shown to have high potential for stink bug control in conventional and organic crop production systems [11,12,13]. For examples, the use of sorghum as a trap crop for N. viridula allows for consistently reducing the insecticide treatments on cotton [12]; a complex of triticale, sorghum, and sunflower as potential trap plants can be potentially useful in cotton fields against the stink bugs (Heteroptera: Pentatomidae) Euschistus servus (Say), Chinavia hilaris (Say) and N. viridula [14]. A prerequisite for trap plants is high attractiveness to the pest in comparison with the plant to be protected. Consequently, the choice of the attractant source used (i.e., a preferred host in a specific stage) is of crucial importance [10,15,16].
In the case of B. hilaris, a pest native of Asia and Africa known mainly for its damage to brassicaceous plants [17,18,19,20], a distinct predilection for attacking few species and varieties has been shown, such as Brassica oleracea var botrytis L. [21], Raphanus sativus L. [22], and Eruca sativa L. [23]. Moreover, unlike other phytophagous stink bugs of Brassicaceae, B. hilaris showed a preference for the plants at seedling stage [22]. A recent study by Guarino et al. [24] suggested that B. hilaris attraction toward Brassica seedlings is determined by the emission of a few diterpene hydrocarbons, the main one recently identified and named brassicadiene [25]. These data already suggest the possibility of using these seedlings as attractive sources for B. hilaris. However, an important step to evaluate these seedlings as a potential trap plant is to test them directly in the field, where the background of stimuli is much more complex than in the laboratory.
The objective of this study was to evaluate the attractiveness of host plant seedling from three species adapted to the Mediterranean climate, B. oleracea var. botrytis (cauliflower), E. sativa (rocket) and Brassica carinata Braun (Abyssinian cabbage), as potential trap plants for B. hilaris. The host choice preference for seedlings was first evaluated in laboratory and field experiments, also noting the different response of the insects to the growth stages of the seedlings.
Finally, the seedlings were tested in a trap plant evaluation trial in Pantelleria caper bush cultivations to test the attractiveness of B. oleracea and E. sativa in the presence of caper bushes and their ability to divert B. hilaris bugs and protect the caper bushes from attack.
2. Materials and Methods
2.1. Insects
The colony of B. hilaris was established and restocked regularly at the Department of Agriculture, Food and Forest Sciences, University of Palermo (Italy) with individuals collected from caper bush fields on the island of Pantelleria (Italy). Insects were reared in climatic cell (30 ± 2 °C, 70 ± 10% RH, photoperiod 16 L:8 D) in 47.5 × 47.5 × 47.5 cm cages (BugDorm-44545; MegaView Science Co. Ltd., Taichung, Taiwan). The colony was fed with cauliflower and cabbage plants, depending on seasonal availability. Because B. hilaris lays eggs in the soil, paper dishes (6-cm Ø) with a mixture of sand, silt, and clay (33% for each soil component) were placed in the cages as oviposition sites. Dishes were changed weekly, and those with eggs were kept in separate cages until the emergence of nymphs. The nymphs were then kept in separate cages until the final molt to adults. Newly emerging adults were used individually in laboratory experiments.
2.2. Seedlings
Seeds of B. oleracea var. botrytis (afterward named B. oleracea), E. sativa, and B. carinata were placed on cotton wool (10 g) soaked with distilled water and held in glass containers. Seeds were sown at a density of about 0.5/cm2. The containers were placed in an environmentally controlled growth chamber (25 ± 1 °C, 70 ± 10% RH, photoperiod 16 L: 8 D) equipped with lights with a photosynthetic flux density (PPFD) of 600 mol photons m−2 s−1. Cotyledon-stage seedling clusters, made by 50 plants at 3–9 days from emergence, were prepared; the cotton wool substrate was wrapped in aluminum foil to avoid desiccation and then used for laboratory bioassays. The seedlings used for the experiments carried out in the field were grown in a nursery in Pantelleria, in open condition.
2.3. Host Choice Assesment
2.3.1. Laboratory Experiments
Host choice assessment of E. sativa, B. oleracea, and B carinata was carried out in the laboratory with a dual choice arena and in open vertical olfactometer bioassays. The B. hilaris choice was evaluated in two separate pairwise comparison experiments of the different species using plants at the ages of 3 and 9 days from emergence.
The arena consisted of a Plexiglas cage 50 cm × 30 cm × 30 cm high. The two side walls of the cage were made with net for ventilation. Inside the arena, two clusters of different plant species were placed 20 cm apart in the arena. Each cluster was made by n = 50 cotyledon-stage seedlings. Individuals (n = 6 per replicate) were released into the cage at 2:00 PM and the total number of insects on each seedling cluster was observed and recorded after 20 h, at 10:00 AM the next day (final choice). The position of the treatments inside the arena was alternated after each replication. After each bioassay, the Plexiglas cage was cleaned with water and dried. Observations were conducted in static air, under controlled laboratory temperature and humidity conditions (25 ± 2 °C, and 50 ± 15% RH). A total of 11 replications were carried out for each test.
Olfactometer bioassays were carried out to verify the role of volatile organic compounds (VOCs) in B. hilaris attraction to the tested cotyledon-stage seedlings. The open vertical Y-shaped olfactometer consisted of a brass rod (left and right arms 20 cm long, central arm 25 cm long, 1 cm diameter). The left and right arms were partially covered with two glass tubes (18 cm long, 5 cm diameter) terminating in hose nipples connected by Tygon tubes to a high–purity air source, and air flow was controlled with a flow-meter at a rate of 0.2 L/min. The air flowed through two glass chambers (500 mL each), which held the test stimuli (seedling clusters N = 50). Light was provided with a halogen lamp (Osram, 12 V–35 W, Münich, Germany) hanging 30 cm above the olfactometer. Experiments were carried out under controlled laboratory temperature and humidity conditions (25 ± 2 °C, and 50 ± 15% RH) starting from 10:00 AM to 15:00 PM. For each replicate, a single adult individual was gently placed at the bottom of the central arm of the olfactometer with a paintbrush and allowed 10 min to respond. Both males and females were individually used for the experiment, at a ratio of 1:1. The bugs moved from the bottom upward toward the light source and upon arriving at the Y junction, had the option to choose between the different volatile stimuli. The criterion for a response was that the test bug walked in the test arm or the control arm for at least 5 cm past the Y junction (first choice). Bugs that did not move into one of the two arms during the 10 min trial were scored as non-responders and were not included in the analysis. After 8 replications, the glass parts of the apparatus were washed with water and detergent, then wiped with acetone and the brass rod was cleaned with distilled water and acetone and baked at 200 °C for 60 min.
Insects were used separately for dual choice arena and olfactometer experiments; each individual was tested only once and then placed in rearing cages. All the insects were starved for one day before the start of the experiments.
2.3.2. Field Trap Bioassays
Cotyledon-stage seedlings of B. oleracea and E. sativa were tested as attractant lures in traps in a caper bush field infested by B. hilaris. A cluster of 50 cotyledon-stage seedlings (7 days old) with soaked cotton used as growing substrate were placed inside the traps. For this experiment a horizontal plastic trap (25 × 15 × 15 cm) furnished by GEA S.r.L., (Settimo Milanese Milan—Italy) was used, with a funnel-like shape on both sides to allow insect entrance. The trap used (S1) was specifically conceived and designed in consideration of B. hilaris behavior characterized by oviposition activity and mating mainly happening on the ground [5]. The traps were placed near a randomly chosen caper bush plant and partially buried in order to facilitate insect entrance and to prevent wind damage (S1). Paraffin oil was applied on the board of the inner part of the funnels to prevent the insects’ escape. Four traps per each seedling species and control (empty traps with soaked cotton wool) were used. Traps were placed in a Pantelleria caper bush field of 0.5 hectares (36°46’15.5” N 11°57’43.9” E) homogeneously infested by B. hilaris, and disposed using a Latin square design. The distance between traps was approximately 8 m. The traps were inspected every 3 days for 15 days and the trapped individuals (adults and nymphs) were counted and removed. During inspections, water was provided to the seedlings to avoid their desiccation and also to control trap.
2.4. Trap Plant Evaluation
Host preference bioassays using B. oleracea, E. sativa, and B. carinata were carried out in a caper bush field in Pantelleria (36°46′21.7” N 11°57′38.1” E). The caper bush field chosen for the experiment was 40 m long and 80 m wide and presented a heavy infestation of B. hilaris. The experiment was carried out using aluminum trays (40 × 30 × 10 cm) as artificial pots, containing cotton soaked with water used as growing substrate. Each species (treatment) was sown in four trays (n = 4 for each of the three treatments) using 5 g of seeds for each tray. The seeds were sown in July 2019 in a nursery, irrigated and transferred to the field one day after emergence. Irrigation was carried out every two days. As the rows of caper bush closer to the dry stone walls surroundings the field presented the highest level of infestation, the trays were placed between the cultivating rows and distributed around the field’s perimeter, alternating the treatments at a distance of 20 m from one to each other. The number of total B. hilaris individuals present on each cotyledon-stage seedling tray was counted and removed from the pot using a paint brush every 2 days until the 15th day after the seedlings’ emergence. Trays were rotated clock-wise by one position after each inspection, to avoid possible position bias.
2.5. Statistical Analysis
The data obtained from the laboratory experiments in dual choice arena were analyzed using a paired t-test, while the ones obtained from the open vertical Y-shaped olfactometer bioassay were analyzed using χ2 tests. The data obtained from the field trap bioassays were analyzed using a one-way ANOVA followed by Tukey test. Finally, the data obtained from the trap plant evaluation bioassays were analyzed using two-way ANOVA, considering the factors “treatment”, “time of inspection” and their interaction, while Fisher least significant difference analysis (LSD) was used to distinguish differences among means for individuals for each inspection day. All the statistical analyses were performed using Statistica 10.0 for Windows (Statsoft, Vigonza, Padova, Italy).
3. Results
3.1. Host Choice Assesment
3.1.1. Laboratory Experiments
The response of B. hilaris adults to the different cotyledon-stage seedling clusters at 3 and 9 days in the dual choice arena is reported in Figure 1. Overall, the seedlings that attracted the highest number of B. hilaris adults were B. oleracea and E. sativa, with B. carinata resulting as the least preferred. More specifically, using 3-day old seedlings, B. hilaris adults showed a similar attraction response toward B. oleracea and E. sativa (t = 1.30; df = 10; p = 0.22), while B. hilaris adults preferred B. oleracea over B. carinata (t = 5.01; df = 10; p < 0.001), and E. sativa over B. carinata (t = 3.19; df = 10; p = 0.009). Furthermore, using 9-day old seedlings, B. hilaris adults preferred B. oleracea over E. sativa (t = 3.54; df = 10; p = 0.005), and over B. carinata (t = 6.62; df = 10; p < 0.001), as well as B. hilaris preferred E. sativa over B. carinata (t = 5.58; df = 10; p < 0.001).
The response of B. hilaris adults to the volatiles of the different 3 or 9-day old seedling clusters in the open vertical Y-shaped olfactometer is reported in Figure 2.
In the experiments using 3-day old seedlings, B. hilaris adults oriented toward E. sativa over B. carinata volatiles (χ2 = 6.52, df = 1, p = 0.012, n = 64), but did not show significant responses between volatiles of B. oleracea and E. sativa (χ2 = 0.40, df = 1, p = 0.52, n = 61), or between B. oleracea and B. carinata (χ2 = 2.25, df = 1, p = 0.13, n = 64).
In the experiments with 9-day old seedlings, B. hilaris adults were also more strongly attracted to volatiles from B. oleracea than to volatiles of B. carinata (χ2 = 8.53, df = 1, p = 0.03, n = 120), and to E. sativa volatiles more than to B. carinata volatiles (χ2 = 7.71, df = 1, p = 0.005, n = 109). However, the adult bugs were equally attracted to volatiles from B. oleracea and E. sativa (χ2 = 2.61, df = 1, p = 0.10, n = 98).
3.1.2. Field Trap Bioassays
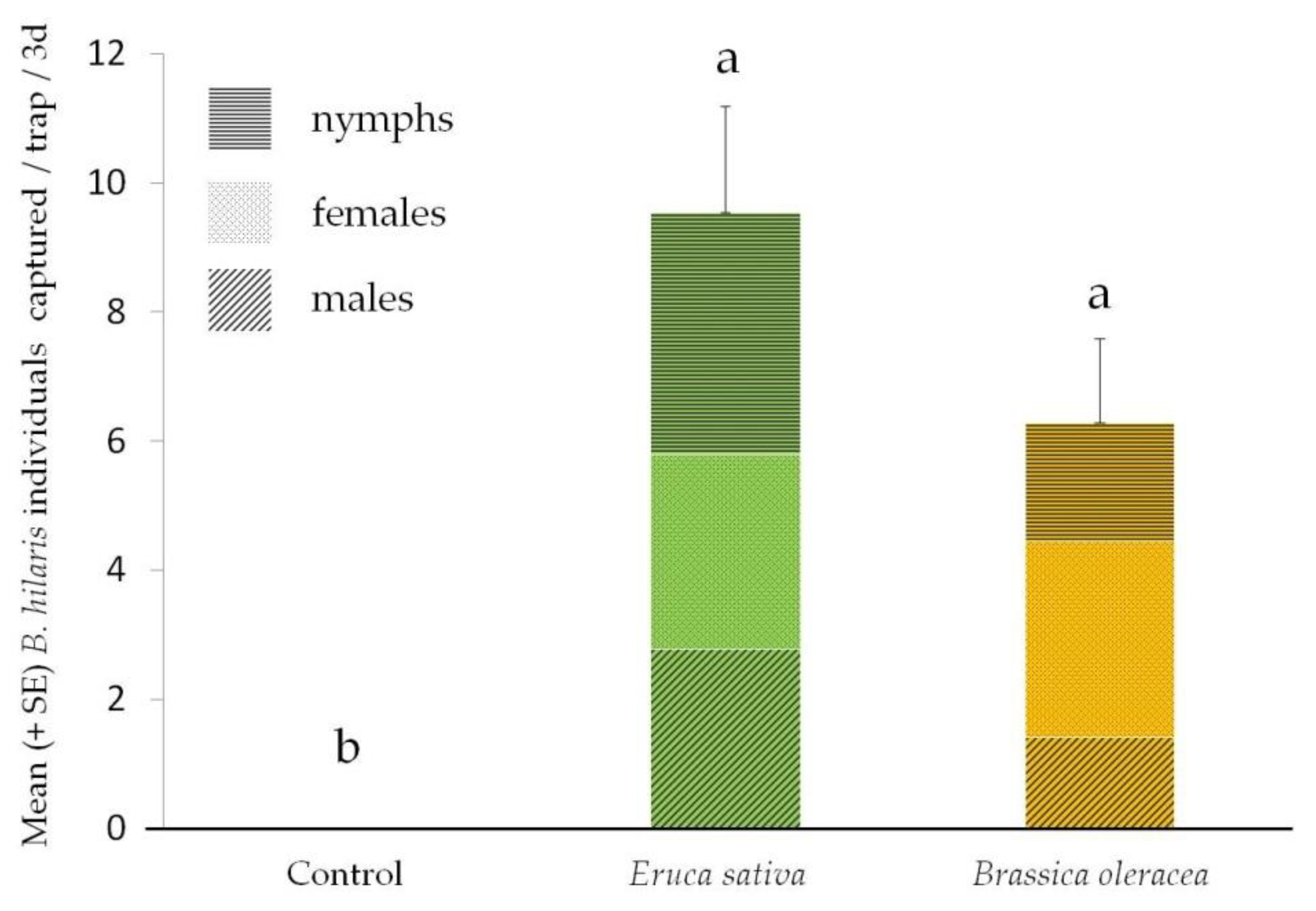
The results of the field trap bioassays using cotyledon-stage seedlings as lures are reported in Figure 3. Overall, the traps baited with B. oleracea and E. sativa captured more individuals than the control (F = 16.03, df = 2, p < 0.0001). More specifically, the traps baited with B. oleracea and E. sativa captured respectively a mean (± SE) of 6.29 ± 1.2 and 9.54 ± 1.65 individuals per trap per 3 days, while no captures were recorded in the control traps. However, the number of captures determined by B. oleracea and E. sativa did not differ statistically (p = 0.14; Tukey test).
3.2. Trap Plant Evaluation
The results of the trap plant evaluation are reported in Figure 4. Overall, the effect of time of inspection and treatment determined differences statistically significant (p < 0.01; ANOVA), while the interaction for the two factors was marginally not significant (p = 0.08; ANOVA). The trays with cotyledon-stage seedlings of E. sativa attracted a higher number of B. hilaris individuals (adults and nymphs) compared to B. oleracea and B. carinata at the 5th and 7th day from emergence (p < 0.05; ANOVA). However, successively the E. sativa attracted a number of individuals similar to the other species. Moreover, at the 9th, 11th, and 13th day from their emergence, the B. oleracea attracted a higher number of individuals compared to B. carinata (p < 0.05; ANOVA). Overall, the attraction of B. oleracea and E. sativa cotyledon-stage seedlings progressively decreased after the 11th day from emergence. This was in part determined by the strong feeding damage by B. hilaris that led to the death of the majority of the cotyledon-stage seedlings.
4. Discussion
The results provide evidence that cotyledon-stage seedlings of B. oleracea and E. sativa are good candidates as attractant sources for B. hilaris and may find application as a lure for traps or in trap cropping techniques.
The attraction of B. hilaris to these seedlings is mediated by the specific volatile organic compounds (VOCs) emitted from the species tested. In fact, the olfactometer bioassays firstly evidenced that the attraction to preferred hosts is mediated by olfactory cues. In particular, testing the seedlings at 3 and 9 days from emergence, B. hilaris adults oriented toward VOCs from E. sativa rather than from those of B. carinata.
Differently from the majority of phytophagous-plant interaction cases [26], the attraction in this case seems to be determined by a key odorant molecule [24,25] rather than by a blend of ubiquitous compounds as observed. It is possible that the attraction of B. hilaris adults toward E. sativa, already observed also by Joseph et al. [23], can be determined by a few key odorant molecules, which however have not yet been investigated. In the case of B. oleracea, Guarino et al. [24] found that the B. hilaris attraction to the cotyledon-stage seedling VOCs is in particular determined by an uncommon volatile diterpene hydrocarbon of novel identification and named brassicadiene [25], not present in the VOCs of B. carinata.
Field bioassays carried out using living seedlings inside the traps as an attractant lure not only confirmed the high attraction of B. hilaris to B. oleracea and E. sativa, but also poses the basis for the use of living germinating seeds as a new type of attractant for monitoring this species. Even if the use of vegetal material as attractant lure for trapping insects has already been exploited for other pests such as the Red Palm weevil [27,28], the use of such seedlings as an attractant is novel to our knowledge. The use of such a lure could be also of interest considering how recent experiments with pheromone-based traps for monitoring B. hilaris captured a low number of individuals [29].
Finally, the data obtained from the trap plant experiments showed that both E. sativa and B. oleracea cotyledon-stage seedlings can attract a consistent number of B. hilaris individuals to a lesser extent than B. carinata, diverting them from the surrounding caper bush plants. These results are encouraging for more in-depth exploration into the possibility of using such seedlings for trap cropping purposes to protect caper bush plants. Furthermore, in accordance with the laboratory bioassays results, B. oleracea and E. sativa elicited a different attraction on B. hilaris individuals over time, probably related to the seedling growth rate and the relative volatiles emitted. More specifically, while E. sativa attracted the highest number of individuals during the first part of the experiment, later on B. oleracea attracted a similar number of individuals. The individuals, adults of both sexes and nymphs at all stages, were pooled together to simplify the observation. It should also be pointed out that the decrease in seedling cluster attraction in the later observations could partially have been determined by the mortality caused by the large aggregations of B. hilaris individuals on the seedlings, particularly evident in the E. sativa clusters. This may require that, before E. sativa trap plants die, they must be destroyed or treated with a pesticide to avoid the B. hilaris individuals from re-infesting the caper bush plants. Otherwise, the B. oleracea seedlings showed greater attraction to B. hilaris at the 9th day after emergence.
Overall, these results suggest that the cotyledon-stage seedlings used could have potential use as trap plants to manage this pest, also encouraged by the strong needs Pantelleria caper growers have for alternative and reliable methods to control Painted bug populations. However, it is also to be stated that, even if these seedlings have showed great potential as attractant sources, more studies are needful to better understand if their application as trap plants can determine a damage reduction of B. hilaris on the caper bush, as this was not specifically assessed in this study.
In our trap plant evaluation, we decided to carry out the experiment in the summer when the B. hilaris population was at the highest level in order to attract the highest number of individuals. However, due to the high temperature, it was necessary to periodically irrigate the trays containing the young seedlings to prevent their desiccation. In order to exploit the trap cropping technique successfully, and in consideration of the well-known water shortage in the island of Pantelleria [30], it could be alternatively recommended that the brassiceous seeds are sown during rainfall periods, for example in early autumn. This tactic was also suggested by Ludwig and Kok [31], as applying the trap plants in fall could limit the size of the population entering winter diapause. However, alternatively or simultaneously the trap plants could be used in spring to catch the newly emerging colonizers before they infest the caper bushes. In consideration of the fact that B. hilaris generally overwinter in the stone walls surrounding the caper bush fields [5], this last technique could be useful if the trap plants are planted around the perimeter of the main crop in order to resemble the so-called “perimeter trap cropping method” [32]. In fact, as suggested by Shelton and Badenes-Perez [9] “the potential success of a trap cropping system depends on the interaction of the characteristics of the trap plant and its deployment with the ecology and behavior of the targeted insect pest”. To our knowledge, very few cases have been reported of commercially successful trap planting applications [9]. On the other hand, it is encouraging that the use of trap plants using Brassica plants to reduce infestation was reported successfully for other pests such as the coleopteran Psylliodes chrysocephala (L.) and Ceutorhynchus pallidactylus (Marsh) [33] and for the lepidopterean Plutella xylostella (L.) [34].
5. Conclusions
These results have ecological value, demonstrating the attractiveness of B. hilaris to host plant cotyledon-stage seedlings other than the caper crop. The results demonstrate the ability of these seedlings to strategically divert the B. hilaris from caper bushes and be attracted to the attractant trap plants E. sativa and B. oleracea. These new findings offer the opportunity to explore these seedlings as candidates for trap planting in different times of the year, to kill the emerging individuals from diapauses or to suppress the overwintering B. hilaris. These strategies can be considered of particular interest for caper growers who are applying organic farming methods.
Author Contributions
Conceptualization, S.G., M.A.A., S.C., E.P.; methodology, S.G., M.A.A, S.C., E.P.; data curation S.G., analysis, S.G., original draft preparation, S.G., M.A.A.; review & editing, S.G., M.A.A., S.C., E.P., project administration, E.P., S.C.; funding acquisition, S.C.; resources, S.G., M.A.A., S.C., E.P.; validation, S.G., M.A.A., S.C., E.P. All authors have read and agreed to the published version of the manuscript.
Funding
Part of this work was made carried out within the Project “High Studies Programme (Master and Ph.D.) for Iraqi officials at Italian Academic Institutions (HiSPro)” supported by the Iraqi Government and the Italian Ministry of Foreign Affairs (MAE).
Acknowledgments
The authors would like to thank Rosario Cappadona of the Cooperative of Caper Producers of Pantelleria and Erik Vallini and Gabriele Lasagni of Bonomo and Giglio Srl for their helpful assistance during the collection of insects in the field.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Sozzi, G.O. Caper bush: Botany and horticulture. Hort. Rev. 2001, 27, 1251–1288. [Google Scholar]
- Rivera, D.; Inocencio, C.; Obón, C.; Alcaraz, F. Review of food and medicinal uses of Capparis L. Subgenus Capparis (capparidaceae) revisión de los usos alimentarios y medicinales de Capparis Subgénero Capparis (capparidaceae). Econ. Bot. 2003, 57, 515. [Google Scholar] [CrossRef]
- Simoglou, K.B.; Dioli, P. First report of caper (Capparis spinosa) serious infestation by Eurydema eckerleini in cyclades islands, Greece (hemiptera: heteroptera, pentatomidae). Fragm. Entomol. 2017, 49, 656–659. [Google Scholar] [CrossRef] [Green Version]
- Infantino, A.; Tomassoli, L.; Peri, E.; Colazza, S. Viruses, fungi and insect pests affecting caper. Eur. J. Plant Sci. Biotechnol. 2007, 1, 1701–1779. [Google Scholar]
- Colazza, S.; Guarino, S.; Peri, E. Bagrada hilaris (burmeister) (heteroptera: pentatomidae) fitofago dannoso al cappero nell’isola di pantelleria. Inf. Fitopatol. 2004, 12, 30–34. [Google Scholar]
- Carapezza, A. Gli eterotteri dell’isola di pantelleria (insecta, heteroptera). Nat. Sicil. 1981, 5, 73–91. [Google Scholar]
- Bundy, C.S.; Perring, T.M.; Reed, D.A.; Palumbo, J.C.; Grasswitz, T.R.; Jones, W.A. Bagrada hilaris. In Invasive Stink Bugs and Related Species (Pentatomoidea): Biology, Higher Systematics, Semiochemistry, and Management; McPherson, J.E., Ed.; CRC Press: Boca Raton, FL, USA, 2018; pp. 205–242. [Google Scholar]
- Khan, Z.R.; Midega, C.A.; Hutter, N.J.; Wilkins, R.M.; Wadhams, L.J. Assessment of the potential of napier grass (Pennisetum purpureum) varieties as trap plants for management of Chilo partellus. Entomol. Exp. Appl. 2006, 119, 15–22. [Google Scholar] [CrossRef]
- Shelton, A.; Badenes-Perez, F. Concepts and applications of trap cropping in pest management. Ann. Rev. Entomol. 2006, 51, 285–308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hokkanen, H.M. Trap cropping in pest management. Ann. Rev. Entomol. 1991, 36, 119–138. [Google Scholar] [CrossRef]
- Todd, J.W.; Schumann, F.W. Combination of insecticide applications with trap crops of early maturing soybean and southern peas for population management of Nezara viridula in soybean (hemiptera: pentatomidae). J. Entomol. Sci. 1988, 23, 192–199. [Google Scholar] [CrossRef]
- Tillman, P.G. Sorghum as a trap crop for Nezara viridula L. (heteroptera: pentatomidae) in cotton in the southern United States. Environ. Entomol. 2006, 35, 771–783. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, A.L.; Dively, G.; Pote, J.M.; Zinati, G.; Mathews, C. Identifying a potential trap crop for a novel insect pest, Halyomorpha halys (hemiptera: pentatomidae), in organic farms. Environ. Entomol. 2016, 45, 472–478. [Google Scholar] [CrossRef] [PubMed]
- Mizell, R.F.; Riddle, T.C.; Blount, A.S. Trap cropping system to suppress stink bugs in the southern coastal plain. Proc. Fla. State Hortic. Soc. 2008, 121, 377–382. [Google Scholar]
- Cook, S.M.; Smart, L.E.; Martin, J.L.; Murray, D.A.; Watts, N.P.; Williams, I.H. Exploitation of host plant preferences in pest management strategies for oilseed rape (Brassica napus). Entomol. Exp. Appl. 2006, 119, 221–229. [Google Scholar] [CrossRef]
- Guarino, S.; Peri, E.; Colazza, S. Plant and Stink Bug Interactions at Different Trophic Levels; Stinkbugs Borges, M., Cokl, A., Eds.; CRC Press: Boca Raton, FL, USA, 2017; pp. 180–199. [Google Scholar]
- Palumbo, J.C.; Natwick, E.T. The bagrada bug (hemiptera: pentatomidae): A new invasive pest of cole crops in Arizona and California. Plant Health Prog. 2010, 11, 50–53. [Google Scholar] [CrossRef] [Green Version]
- Bundy, C.S.; Grasswitz, T.; Sutherland, C. First report of the invasive stink bug Bagrada hilaris (burmeister) (heteroptera: pentatomidae) from New Mexico, with notes on its biology. Southwest. Entomol. 2012, 37, 411–414. [Google Scholar] [CrossRef]
- Reed, D.A.; Palumbo, J.C.; Perring, T.M.; May, C. Bagrada hilaris (hemiptera: pentatomidae), an invasive stink bug attacking cole crops in the southwestern United States. J. Integr. Pest Manag. 2013, 4, C1–C7. [Google Scholar] [CrossRef] [Green Version]
- Torres-Acosta, R.I.; Sánchez-Peña, S.R.; Torres-Castillo, J.A. Feeding by bagrada bug, Bagrada hilaris, on Moringa oleifera (brassicales: moringaceae) in Mexico. Southwest. Entomol. 2017, 42, 919–922. [Google Scholar] [CrossRef]
- Guarino, S.; Peri, E.; Colazza, S.; Luchi, N.; Michelozzi, M.; Loreto, F. Impact of the invasive painted bug Bagrada hilaris on physiological traits of its host Brassica oleracea var botrytis. Arthropod Plant Interact. 2017, 11, 649–658. [Google Scholar] [CrossRef]
- Huang, T.I.; Reed, D.A.; Perring, T.M.; Palumbo, J.C. Host selection behavior of Bagrada hilaris (hemiptera: pentatomidae) on commercial cruciferous host plants. Crop Prot. 2014, 59, 7–13. [Google Scholar] [CrossRef] [Green Version]
- Joseph, S.V.; Grettenberger, I.M.; Godfrey, L.D.; Zavala, N. Susceptibility of germinating cruciferous seeds to Bagrada hilaris (hemiptera: pentatomidae) feeding injury. Arthropod Plant Interact. 2017, 11, 577–590. [Google Scholar] [CrossRef]
- Guarino, S.; Arif, M.A.; Millar, J.G.; Colazza, S.; Peri, E. Volatile unsaturated hydrocarbons emitted by seedlings of brassica species provide host location cues to Bagrada hilaris. PLoS ONE 2018, 13, e0209870. [Google Scholar] [CrossRef] [PubMed]
- Arriola, K.; Guarino, S.; Schlawis, C.; Arif, M.A.; Colazza, S.; Peri, E.; Schulz, S.; Millar, J.G. Identification of brassicadiene, a diterpene hydrocarbon attractive to the invasive stink bug Bagrada hilaris, from volatiles of cauliflower seedlings, Brassica oleracea var. botrytis. Org. Lett 2020, 22, 2972–2975. [Google Scholar] [CrossRef] [PubMed]
- Bruce, T.J.; Pickett, J.A. Perception of plant volatile blends by herbivorous insects–finding the right mix. Phytochemistry 2011, 72, 1605–1611. [Google Scholar] [CrossRef] [PubMed]
- Giblin-Davis, R.M.; Weissling, T.J.; Oehlschlager, A.C.; Gonzalez, L.M. Field response of Rhynchophorus cruentatus (coleoptera: curculionidae) to its aggregation pheromone and fermenting plant volatiles. Fla. Entomol. 1994, 77, 164–177. [Google Scholar] [CrossRef]
- Peri, E.; Rochat, D.; Belušic, G.; Ilic, M.; Soroker, V.; Barkan, S.; Guarino, S.; Lo Bue, P.; Colazza, S. Rhynchophorus ferrugineus: Behavior, ecology, and communication. In Handbook of Major Palm Pests: Biology and Management; Soroker, V., Colazza, S., Eds.; Wiley-Blackwell: Oxford, UK, 2016; pp. 105–130. [Google Scholar]
- Arif, M.A.; Guarino, S.; Colazza, S.; Peri, E. The role of (E)-2-octenyl acetate as a pheromone of Bagrada hilaris (burmeister): Laboratory and field evaluation. Insects 2020, 11, 109. [Google Scholar] [CrossRef] [Green Version]
- Manenti, F.; Masi, M.; Santucci, G.; Manenti, G. Parametric simulation and economic assessment of a heat integrated geothermal desalination plant. Desalination 2013, 317, 193–205. [Google Scholar] [CrossRef]
- Ludwig, S.; Kok, L. Evaluation of trap crops to manage harlequin bugs, Murgantia histrionica (hahn) (hemiptera: pentatomidae) on broccoli. Crop Prot. 1998, 17, 123–128. [Google Scholar] [CrossRef]
- Boucher, T.J.; Ashley, R.; Durgy, R.; Sciabarrasi, M.; Calderwood, W. Managing the pepper maggot (diptera: tephritidae) using perimeter trap cropping. J. Econ. Entomol. 2003, 96, 420–432. [Google Scholar] [CrossRef]
- Barari, H.; Cook, S.M.; Clark, S.J.; Williams, I.H. Effect of a turnip rape (Brassica rapa) trap crop on stem-mining pests and their parasitoids in winter oilseed rape (Brassica napus). BioControl 2005, 50, 69–86. [Google Scholar] [CrossRef]
- Charleston, D.S.; Kfir, R. The possibility of using Indian mustard, Brassica juncea, as a trap crop for the diamondback moth, Plutella xylostella, in south Africa. Crop Prot. 2000, 19, 455–460. [Google Scholar] [CrossRef]
Figure 1.
Host choice responses (final choice) of Bagrada hilaris adults to 3 and 9-day old cotyledon-stage seedlings of Brassica oleracea, Brassica carinata and Eruca sativa. * = p < 0.01 (t-test for dependent samples), ns = not significant.
Figure 1.
Host choice responses (final choice) of Bagrada hilaris adults to 3 and 9-day old cotyledon-stage seedlings of Brassica oleracea, Brassica carinata and Eruca sativa. * = p < 0.01 (t-test for dependent samples), ns = not significant.

Figure 2.
Y-shaped olfactometer bioassays: Responses (% first choice) of Bagrada hilaris adults to 3- and 9-day old cotyledon-stage seedlings of Brassica oleracea, Brassica carinata and Eruca sativa. n = number of replications; * = p < 0.05; ** = p < 0.01; ns = not significant.
Figure 2.
Y-shaped olfactometer bioassays: Responses (% first choice) of Bagrada hilaris adults to 3- and 9-day old cotyledon-stage seedlings of Brassica oleracea, Brassica carinata and Eruca sativa. n = number of replications; * = p < 0.05; ** = p < 0.01; ns = not significant.

Figure 3.
Mean (+ SE) Bagrada hilaris individuals captured in traps lured by Brassica oleracea and Eruca sativa cotyledon-stage seedling or in control traps. The different letters indicate that values differ statistically at p < 0.05 (one-way ANOVA, followed by Tukey test).
Figure 3.
Mean (+ SE) Bagrada hilaris individuals captured in traps lured by Brassica oleracea and Eruca sativa cotyledon-stage seedling or in control traps. The different letters indicate that values differ statistically at p < 0.05 (one-way ANOVA, followed by Tukey test).

Figure 4.
Mean (+ SE) Bagrada hilaris individuals observed in the cotyledon-stage seedlings trays of Eruca sativa, Brassica oleracea and Brassica carinata in Pantelleria caper bush field for each day of inspection. Different letters within the same day of observation indicate that values differ statistically at p < 0.05 (two-way ANOVA, followed by LSD test).
Figure 4.
Mean (+ SE) Bagrada hilaris individuals observed in the cotyledon-stage seedlings trays of Eruca sativa, Brassica oleracea and Brassica carinata in Pantelleria caper bush field for each day of inspection. Different letters within the same day of observation indicate that values differ statistically at p < 0.05 (two-way ANOVA, followed by LSD test).

© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Arif, M.A.; Guarino, S.; Peri, E.; Colazza, S. Evaluation of Brassicaceae Seedlings as Trap Plants for Bagrada Hilaris Burmeister in Caper Bush Cultivations. Sustainability 2020, 12, 6361. https://doi.org/10.3390/su12166361
AMA Style
Arif MA, Guarino S, Peri E, Colazza S. Evaluation of Brassicaceae Seedlings as Trap Plants for Bagrada Hilaris Burmeister in Caper Bush Cultivations. Sustainability. 2020; 12(16):6361. https://doi.org/10.3390/su12166361
Chicago/Turabian StyleArif, Mokhtar Abdulsattar, Salvatore Guarino, Ezio Peri, and Stefano Colazza. 2020. "Evaluation of Brassicaceae Seedlings as Trap Plants for Bagrada Hilaris Burmeister in Caper Bush Cultivations" Sustainability 12, no. 16: 6361. https://doi.org/10.3390/su12166361
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.