Germplasm Development of Underutilized Temperate U.S. Tree Crops


1
Department of Crop Sciences, Institute of Sustainability, Energy, and Environment, Plant Sciences Laboratory, University of Illinois at Urbana-Champaign, 1201 S. Dorner Drive, Urbana, IL 61820, USA
2
Savanna Institute, 1360 Regent Street #124, Madison, WI 53715, USA
3
Department of Plant Biology and Pathology, Foran Hall, 59 Dudley Road, Rutgers, University, New Brunswick, NJ 08901, USA
*
Authors to whom correspondence should be addressed.
Sustainability 2019, 11(6), 1546; https://doi.org/10.3390/su11061546
Submission received: 3 February 2019
/
Revised: 5 March 2019
/
Accepted: 5 March 2019
/
Published: 14 March 2019
(This article belongs to the Special Issue Sustainable Agroforestry Systems)
Abstract
:In the Midwest U.S. dominated corn-soybean landscape, agroforestry systems can be particularly valuable for increasing the provisioning and regulatory capacity of the agricultural landscape. However, these systems have not yet been broadly integrated into the landscape of this region since they are mostly relegated to marginal lands. A growing body of literature suggests a path to increase the adoption of agroforestry in the Midwest U.S. lies in the incorporation of low-input food-producing tree species that provide economic incentives for farmers. Studies of the system-level integration of such approaches have proceeded by using the currently available cultivars and breeding selections of various tree nut and fruit species. While existing varieties and breeding selections provide the opportunity for initial system development and integration, their broad adaptability to the Midwest U.S. and its marginal land-types is unexplored. Thus, a second tier of research includes the genetic improvement and adaptation of tree crop selections to their respective target environments throughout the Midwest U.S. Fortunately, select tree crops of interest are amendable to systematic breeding and have wild relatives that are endemic across the region. In this paper, we discuss the value of these wild relatives for broadening the adaption of cultivated tree crop selections by using the hazelnut as an example species. We present a framework using geospatial tools to define and prioritize target environments for breeding and, in turn, exploiting wild relative germplasm.
1. Introduction
The benefits of agroforestry’s regulatory services are well characterized for the Midwest U.S., where annual cropping systems such as the corn-soybean rotation dominate the landscape [1,2,3]. Under the appropriate conditions, select agroforestry systems have even demonstrated superior production capacity and profitability compared to their respective annual grain counterparts [4,5,6], which brings pragmatism to the strategic diversification within Midwest U.S. Nevertheless, adoption of agroforestry continues to be relatively limited throughout this region. A growing body of research suggests a path to increase the adoption of U.S. agroforestry systems lies in the integration of low-input fruit and nut producing tree species (henceforth, referred to as tree crops) [1,7,8,9,10,11,12,13,14]. Such systems, described by Lovell et al. [7], provide a unique opportunity to integrate new food production capacity into the Corn Belt while simultaneously providing regulatory services [7,14]. A primary constraint in considering these systems more broadly is the limited availability of tree crop germplasm and the extent to which cultivated selections can be improved and adapted to target environments across the Midwest U.S.
Numerous tree crop species are available for integration into Midwest U.S. agroforestry systems at some scale and range [10]. In recent years, cultivated selections of tree crop species have been tested or released for particular regions of the Midwest U.S. (Table 1), which has subsequently led to new regional crop markets (reviewed in part by Reference [11]). These germplasm improvement efforts provide valuable genetic resources that are foundational for tree crop development, but a systematic framework is needed to effectively exploit these genetic resources within environments across the Midwest U.S. Tree crops (such as those in Table 1), which are often underutilized species and have varying degrees of assembled genetic resources. Additionally, the development of these tree crops will, in many cases, be restricted to the marginal lands of row-crop regions, which have not been studied for suitability with respect to productivity of specific tree crops. Fortunately, many tree crops are good targets for schematic breeding, and they have wild relatives that are endemic to the range of the Midwest U.S. To be relevant for broad adoption within the region, a framework for tree crop improvement must include systematic steps to target selection environments within the marginal lands of row-crops and, in doing so, identify the associated adaptive traits required of tree crop wild relatives. Forming and implementing such a framework persists as a critical gap in tree crop development.
In this review, we synthesize literature to support a case for the broad integration of select tree crops into the Midwest agricultural landscape. We first contextualize the roles of tree crops within the landscape and then present focal considerations for species development. Thereafter, we make a case for the value of tree crop wild relatives and conceptually define target environments for broad integration into the landscape especially lands that are marginal to row-crops. Lastly, we present a framework using geospatial tools to systematically target environments of the Midwest U.S. for tree crop adaptation using corresponding wild relatives.
2. Finding a Place for Tree Crops in the Midwest U.S.
The suitability of tree crops to the Midwest U.S. relates to the species’ capacity to yield both provisioning and regulatory services when cultivated within the marginal lands of the maize-soybean rotation. Moreover, an element of this capacity is the species’ propensity for genetic improvement within these respective marginal lands. Henceforth, we refer to the marginal lands of the maize-soybean rotation in the Midwest U.S. just as marginal lands, despite the fact that these areas can still be productive and might not be considered marginal in comparison to lands in other regions.
In the Midwest U.S., maize and soybean yields have increased linearly over the past several decades [29,30]. Recent satellite mapping suggests that maize production gains are disproportionally occurring on well-suited parcels and within-field locations [30], as opposed to low-yielding lands or marginal lands. This trend is in part due to the responsiveness of well suited land-types to the maize system’s prominent scientific improvements (i.e., high-density planting, variable rate technologies, and selection of maize hybrids within high-yielding environments) [30]. In fact, maize yield improvements have been accompanied by a higher sensitivity to drought stress, which may be induced more rapidly in low-yielding, marginal environments [31]. Nevertheless, the relative lack of maize yield improvement within low-yielding and marginal lands alone provides justification to consider tree crops that are better suited to these environments [4,30].
Considering tree crops for cultivation on marginal lands requires a procedural approach to pair discrete locations with corresponding well-suited tree crops, so that their adoption and subsequent improvement can proceed accordingly. Low crop productivity and profitability are often used to define marginal land-types [32,33,34]. Collectively, such marginal lands represent millions of underutilized hectares throughout the Midwest U.S. [35], which possess varying characteristics that cause marginality. Select tree crops have been described as compliments to the complex row-crop landscape [7,10,14] in that they are low-input [36,37,38] and suited to attributes typical of certain marginal land types (e.g., highly erodible, sloped lands) [10,39]. However, in practice, the extent to which particular tree crop selections are well suited to marginal lands remains to be determined. Filling this knowledge gap will guide development initiatives by clarifying the extent to which a given tree crop can be integrated into marginal lands, as well as the geographic distribution of those lands and the secondary considerations those locations might carry.
Geospatial mapping and analysis could offer insight regarding the extent to which marginal lands exhibit characteristics that match the suitability of respective tree crops, with a potential to estimate profitability. To date, bioenergy grasses have been the primary perennial crops modeled as alternatives for the Midwest U.S., and they demonstrate the viability of agricultural diversification with perennial species for the region [4,5]. Spatial mapping and economic analysis have demonstrated improvement of farm-level profitability through targeted subfield cultivation of perennial grasses, switchgrass (Panicum virgatum), and miscanthus (Miscanthus giganteus), in place of low-yielding maize and soybean [4,5]. A recent study by Wolz and DeLucia [6] utilized similar spatial mapping of timber-based black walnut (Juglans nigra) plantations and alley cropping systems to demonstrate increased profitability through targeted system placement. Across four states in the Midwest U.S., this study showed that black walnut alley cropping systems could increase profitability for landowners on 23.4% of cultivated land, assuming a 5% discount rate in future yields [6]. In effect, the studies by Brandes et al. [4,5] and Wolz and DeLucia [6] provide evidence for considering the targeted integration of other low-input perennials, which extends to a growing number of tree crop species.
Select tree crops demonstrate potential as new or emerging commercial crops for the Midwest U.S. Research to advance the commercialization of these tree crops has progressed over the past decade, with emphasis on adapting cultivated selections to particular regions across the Eastern and Midwest U.S. (Table 1). Variety improvement has stimulated new regional tree crop industries [11] and, in some cases, assembled resources requisite to access domestic markets [26], e.g., diverse and characterized breeding material. Systematic exploitation of existing resources offers an opportunity to scale these crops and capitalize on regional and domestic market growth [11,40]. Foremost, this exploitation entails expanding the production range well-suited for cultivated tree crop selections to additional regions throughout the Midwest U.S., which is discussed at depth in subsequent sections.
The benefits of incorporating tree cultivation and conservation into agriculture (i.e., agroforestry) are diverse and well-documented [41,42,43], even for the Midwest U.S. [1,44,45,46,47,48,49]. Strategic integration of trees into temperate croplands adds substantial capacity to mitigate climate change by reducing agricultural greenhouse gas emissions [47,50,51] and sequestering carbon [42,49]. Udawatta and Jose [49] provide an in-depth synthesis of the carbon sequestration capacity of temperate North American agroforestry systems and, conservatively, estimate the sequestration potential of agroforestry systems at 12.4 Mg C ha−1 year−1 to a landscape-level capacity of 65.7 Tg C year−1, excluding silvopastoral systems. Diversifying the tree species that are common to North American agroforestry systems—beyond timber species like walnut (Juglans sp.) and poplar (Populus sp.)—to include tree crops will help realize agroforestry’s sequestration potential [14].
Temperate agroforestry systems also provide services to filter run-off [52], reduce erosion [9,53], and create habitat that fosters biodiversity [1,2,3]. Deep tree roots and a longer growing season enable agroforestry systems to reduce nitrate leaching compared to crop systems and perennial pasture [52,54,55], with increasing reductions of leaching at greater soil depths [54]. Consistent with the concept of integrating agroforestry on marginal lands, various studies have demonstrated that subfield row-crop areas that are the lowest yielding coincide with those of the highest environmental risks [56], including soil erosion [9], water, and soil quality [4,57]. Disproportional reductions in soil erosion and nitrate leaching have been demonstrated by targeting these areas with perennial cropping systems [9].
3. Considerations for Tree Crop Development
Numerous tree crops are endemic or somewhat well-adapted to the U.S. [58,59]. Prioritizing species with sufficient potential to warrant development is essential [60]. Measures to prioritize the development of underutilized species are seldom discussed with respect to the temperate U.S. with the exception of several related studies that have systematically categorized the relative importance of wild crop relatives for collection [59,61,62] and generally detailed the use of wild crop relatives in systematic breeding [63,64].
Opportunities exist to bring new crop species into a production region when a respective trait outperforms the region’s major crop under select conditions. Referred to as a trait-based approach, the strategy is useful to gauge the potential of a particular underutilized species, in which the production or quality of a specific commercial trait is compared to the major crop when cultivated on a particular marginal land-type or under relevant abiotic stresses (e.g., heat and drought stress) [60]. When considering such opportunities, the economic value of the tree crop or its trait of interest must be apparent and exploitable within existing markets [60]. Trait-based comparisons may be most applicable to tree crop species with high value markets (e.g., oil, protein, vitamin, or phytonutrient). One such example is elderberry (Sambucus sp.) and a growing market for healthy plant-based food dyes made from phytonutrients [65]. High phytonutrient content is a trait intrinsic to the elderberry genus [66,67]. The concentration of total phytonutrients in elderberry are naturally higher than the relevant accessions of maize and various vegetable species. Furthermore, elderberry is adapted to a wide range soils in the Midwest U.S. [68], including those that are sloped and moderately fertile. In these environments, elderberry’s field-level phytonutrient production can be compared to that of maize and vegetable selections. If elderberry provides a production advantage within these environments, there is precedent for crop integration.
While intuitive, basic market trends provide valuable support for prioritization of tree crops for improvement. Market data may reveal shifts in global production or indicate whether markets can stably accommodate an increased supply. Drivers of market growth or gaps are also valuable considerations since they shed light on constraints for market access or even competitive opportunities for market newcomers. Box 1 uses the hazelnut to emphasize the value such information offers. Collectively, such material is valuable to improve stakeholder engagement and garner industry development.
Box 1. An economic case to broaden hazelnut cultivation in the U.S.
Commercial cultivation of hazelnut has been an ongoing pursuit in the eastern and north-central regions. The global hazelnut market is forecasted to grow substantially from 2017–2021 from $4.15 to $5.75 billion [40]. Although the U.S. hazelnut market is small and young compared to the European market, the Americas are expected to be the fastest growing market during this time (7.57% annual growth, $0.262 billion) [40,69]. Ferrero SpA (Alba, Italy)—manufacturer of Nutella® and Ferrero Rocher® and purchaser of 25% of the annual global hazelnut supply—has helped drive new growth for hazelnut-based products in the U.S. confectionary markets, which has, in turn, spurred new purchasing of hazelnut by American chocolate companies, like The Hershey Company [40].
Growth in the global hazelnut market has incentivized minor hazelnut producing countries to support development initiatives aimed at expanding cultivation [25,70,71,72,73,74]. This incentive has been furthered by price peaks caused by constraints to the Turkish hazelnut supply, which accounts for 60% to 70% of the global supply. Turkish cultivation relies primarily on superior wild selections or landraces, and most orchards are over a half-century old and gradually declining in productivity [75]. Moreover, new high-yielding varieties are not available for a new orchard establishment because of the absence of variety improvement programs in the region [76]. Additionally, the Turkish hazelnut supply has greatly fluctuated in recent years due to abiotic stresses [77]. In 2013 and 2014, serve winter frost and subsequent drought damaged the Turkish crop to cause 30% and 50% reductions to production, respectively [78]. The resulting shortages led to peak prices, motivating Ferrero SpA and companies alike to lessen their dependency on the Turkish supply. In recent years, Ferrero SpA has heavily invested in hazelnut industry development in several non-traditional nations (e.g., Canada, Chile, and Australia).
These market trends highlight an opportunity for new growing regions to gain a share of the hazelnut market. The U.S. maintains mature and dedicated variety improvement programs, while development initiatives in other countries primarily focus on variety trials of traditional landraces and cultivars. These U.S. breeding programs are a competitive advantage for supporting the growth of U.S. hazelnut production. The endemic American hazelnut species also offers an additional competitive advantage in that it provides adaptability traits requisite for the comprehensive adaptation of the commercial Corylus avellana (European hazelnut) to the Eastern and Midwest U.S.
The genetic resources of a given species and its wild relatives are fundamental parameters that determine the outlook of tree crop development. Tree crops with superior wild selections or suitable breeding selections that can serve as first generation varieties hold an invaluable advantage to help facilitate grower adoption and prime markets. The availability of tree crop wild relatives adapted to the targeted cultivation regions is critical. Sufficient genetic diversity and ability to discover adaptive traits is an essential parameter to species prioritization.
4. Expanding the Cultivated Range of Tree Crops Using Wild Relatives
Wild crop relatives are increasingly discussed as a source of novel traits relevant to the agriculture’s challenges [63,64,79,80,81]. Such germplasm offers the ability to introduce traits from the far side of the domestication bottleneck and expand breeding pools with alleles that contribute to improved yields, adaptation to abiotic stresses, and an expanded cultivation range. While cultivated tree species have undergone less severe domestication bottlenecks than annual crops [82], the systematic introduction of wild germplasm still provides novel allelic diversity to breeding pools. The wild relatives of the tree crops discussed in this case carry a variety of traits of commercial value, including fruit quality, disease resistance, and abiotic stress tolerance (Table 2). For example, the American hazelnut, Corylus americana, is a wild relative of growing value since breeders seek to expand the cultivated range of the European hazelnut to non-traditional climates within the U.S. [83]. The species is endemic to a vast and continuous area of the eastern U.S. and represents a diverse source of adaptability traits to the primary stressors that limit commercial cultivation—eastern filbert blight resistance and climatic stressors (discussed in Box 2). The introgression of such traits in perennial systems is increasingly more feasible and efficient due to genomic-assisted techniques that accelerate allele discovery and progeny selection [80,84,85].
Box 2. Expanding the cultivated range of the European hazelnut using the American hazelnut.

Commercial hazelnut cultivation is an ongoing pursuit in the Eastern and North-central U.S. over the last several decades [25,26]. Development initiatives center on the adaptation of cultivated European hazelnut selections (Corylus avellana) through the introgression of regional climatic adaptations and eastern filbert blight (EFB) resistance [24,83,96].
The endemic hazelnut species (C. americana) is native to a vast extent of the eastern half of North America [97]. This extensive geography suggests that C. americana holds a diverse source of resistance, and, moreover, a broad source of climatic adaptations [98,99]. In nature, the pathogen causing eastern filbert blight, A. anomala, co-exists exclusively with C. americana as its native host, occasionally producing minor, inconsequential cankers [98,99,100], while the infection of C. avellana causes perennial cankering that leads to branch dieback, yield decline, and, in many cases, plant death [101]. Additionally, recent examination of A. anomala diversity demonstrates population differentiation [102] and supports the need for integrating durable multigenic resistance like that of C. americana into breeding programs.
While C. americana is a diverse source of climatic adaptations and EFB resistance, its nut quality and plant architecture are not suited for commercial production and existing markets. Compared to cultivated C. avellana, the kernels of C. americana are much smaller, enclosed in thicker shells and involucres, which complicates their retrieval during harvest and processing, and retain a higher percent of their pellicle following blanching (above). The species’ shrub architecture and aggressive basal sprouts also deviate from standard single-stemmed hazelnut orchards. Fortunately, reciprocal crosses between C. americana and C. avellana are viable [103], and breeders suggest a modified multi-generational backcross to recover commercial traits of the recurrent parent(s) in which the recurrent parents are complimentary and unrelated [24]. Promisingly, breeders have observed the quick recovery of commercial traits in the first and second backcross generations [104]. C. americana germplasm collection and characterization has expanded accordingly to enable broad use of the species in breeding.
A majority of tree crop species are amenable to clonal propagation (i.e., asexual and vegetative), and, as a consequence, many tree crop progenitors and even cultivated varieties are clonal propagates of superior wild selections [82]. The ability for “instant” domestication through clonal propagation means that current tree crop varieties are typically only a few generations removed from their wild relatives, and, collectively, these selections often maintain similar levels of neutral genetic variation compared to their wild populations [82]. Additionally, woody perennial varieties often remain genetically indistinct from their uncultivated wild counterparts [84]. Comparative multivariate analysis of genome-wide DNA markers shows that cultivated individuals of apple [105], grape [106,107], and European hazelnut [108] appear undifferentiated from either their progenitors or wild relatives, even following selection. Such genetic similarities suggest ongoing gene flow between cultivated selections and wild germplasm following domestication [106], which occurs very regularly [109]. Although not requisite, genetic similarities between these two categories of germplasm provide an optimistic outlook for the role of wild relatives in the introgression of adaptive traits from wild germplasm into breeding selections.
While clonal propagation has been advantageous for prompt domestication and cultivation, it has negated the need to comprehensively introduce wild germplasm into breeding programs. Thus, to-date, wild relatives have been used only sparingly and often just for select traits [81,110,111]. Despite their historical underutilization, wild relatives of tree crops hold extensive phenotypic and genetic diversity critical to broadening the cultivated range of existing tree crop selections. This diversity arises as a consequence of tree reproductive biology (e.g., outcrossing, wind-dispersed pollen, long lifespan) that causes extensive gene flow [82]. In turn, tree species often hold high within-population diversity and a weak population structure [82,112,113,114,115].
Even with minimal among-population structure and high gene flow, divergent selection between geographically distinct natural tree populations still gives rise to local adaptations [82,116]. Local adaptation can arise as a response to a variety of geographically specific selection pressures, e.g., climate, photoperiod, soil characteristics, and pathogens [117,118,119]. While different traits respond variably to selection, many local adaptations of natural tree populations evolve through gradual allele frequency changes at small effects loci of polygenic traits [120,121].
The highly polygenetic architecture of adaptive traits provides insight into how local populations could diverge despite high gene flow. Divergent selection pressure occurs on the many small effects loci individually rather than the polygenic trait itself [122]. Therefore, adaptive traits arise from a collection of allele frequency changes at the small effects loci. These allelic changes diverge weakly at individual loci and are not reflected in population structure [121,123]. Additionally, adaptive traits within local tree populations are often genetically diverse. Due to high levels of outcrossing, linkage disequilibrium (the pattern of non-random allele assortment) is quite low in natural tree populations [124,125,126]. Therefore, the recombination rate among small effects loci that comprise adaptive traits is high. Consequently, a diversity of recombinant offspring are produced annually in healthy tree populations with high fecundity, giving rise to genotypes with different allelic combinations for a given adaptive trait [122].
The local adaptations, as well as other traits of interest (e.g., Table 2) from wild germplasm, can be leveraged through their systematic integration into breeding pools [81]. The desirable traits of wild relatives must be integrated into the genetic background of cultivated selections without the loss of traits of commercial importance. Backcrossing is a common approach for trait introgression, where selected progeny that inherited the wild trait of interest are crossed recurrently to advanced breeding selections. Numerous adaptive traits of both monogenic and polygenic architecture have been introgressed into elite varieties. Linkage drag, however, can produce undesirable progeny in backcross generations, if undesirable phenotypes from wild germplasm are linked to the introgressed loci [127]. Increasing progeny size and additional backcross generations can facilitate the development of rare recombinants that break linkage drag.
Additionally, rapid linkage disequilibrium decay (i.e., the decrease of non-random segregation) is common in high diversity tree crops [107,124,128]. This decay is the consequence of high levels of ancestral recombination occurring in natural outcrossing tree populations, which improves genetic mapping resolution and enables genome-wide association studies (GWAS) [129]. GWAS can proceed using existing plant populations of unrelated individuals (i.e., natural populations or germplasm collections) and, thus, circumvents the time needed to develop and grow linkage-mapping populations. Consequently, GWAS creates the potential to quickly identify markers tightly linked with the causal loci and accelerate the identification of rare recombinants in backcrossing generations [130].
When considering the introgression of adaptive traits, recombinant progeny are most effectively identified when selection occurs within the target environment (e.g., marginal lands) [131,132]. However, much of the historic selection has prioritized the most ideal, productive environments [132]. Decentralizing selection to the targeted cultivation environment(s) ultimately makes the selection environment as similar to the cultivated environment as possible [133]. As a result, decentralization improves the response to selection for specific adaptations within the respective environments of the target region (compared to selecting for wide adaption across the region) [134]. While decentralizing selection to improve the selection response is intuitive, the selection environment for public plant breeding programs has historically been restricted to centralized research stations, and on-farm testing is restricted to the trialing of only a few varieties [132]. The decentralization of selection is particularly important in the breeding of tree crops for the Midwest U.S., where breeding goals center on expanding the cultivated range of respective species.
5. Defining Target Environments for Decentralized Selection
While the decentralization of selection is conceptually straightforward, its implementation is complex. Target environment selection is based upon an assortment of characteristics that determine whether a given location is a suitable cultivation environment for the tree crop. These characteristics are highly heterogeneous among and within environments across the landscape, which leads to the recombination of environmental stressors and the creation of unique environments [30]. Despite high heterogeneity, breeders must choose the most opportune series of target environments that are representative of the targeted cultivation region. Subsequently, actual selection sites that represent each target environment must be identified for the placement of progeny tests. To-date, a systematic approach is lacking to accomplish the identification of these target environments.
In this case, we propose a brief logic model to identify target environments and corresponding locations for selection. (A) The approach begins by defining the spatial distribution of the soil characteristics required for the commercial production of cultivated selections within the cultivation region via geospatial suitability mapping. (B) Prioritized target environments can be selected from within the suitable areas using criteria to maximize the impact. (C) The specific adaptability traits required to expand tree crop cultivation to the defined targeted environments can be discretely specified and used to guide a breeding program. High-resolution geospatial techniques, such as this, clarify the spatial distribution of each target environment and create the potential to add additional variables (e.g., photoperiod and climate) for recurrent assessment of variation within target environments to test for sub-environments [5,6,135].
Geospatial mapping of soil suitability characteristics is immensely informative for guiding decentralized selection. Foremost, the suitable cultivation areas are discretely identified from within the greater region, which significantly narrows the potential target environments (Figure 1). Additionally, the spatial distribution and predominant clustering of suitable locations immediately enable some prioritization (or de-prioritization) of the adaptability traits to source from wild relatives (e.g., photoperiod or climatic) as well as the general geographies from which wild germplasm should be sourced, if needed.
The geospatial mapping of soil characteristics requires a well-defined suitability index for the tree crop species. Suitability indices estimate productivity (i.e., growth or yield) of cultivated selections from traditional growing regions as a function of soil and climate characteristics. If sufficient data exists, a suitability index can be a quantitative, continuous index [136]. However, discrete suitability classes are currently more common for tree crops ([135], Table A1). The suitability index can then be mapped using existing soil and/or climate geospatial databases. For the Midwest U.S., pre-existing geospatial data is available through the Soil Survey Geographic (SSURGO) Database (NRCS) for suitability mapping [137].
The specific characteristics that negatively affect suitability (and warrant adaptation) vary significantly among and within regions [30]. This variability has the potential to create many discrete target environments within respective local geographies. Consequently, a systematic framework to statistically differentiate these distinct clusters of target environments is needed. A multivariate approach is intriguing in that individual map units can retain their identity and the dataset complexity can be reduced to principal components. Subsequently, cluster analysis could classify individual map units into similar target environments based on the principal component variation representative of different limitations in soil characteristics.
Following the classification of target environments, there are several criteria that can contribute to the prioritization of target environments for breeding.
- (1)
- Spatial extent: As clusters of prospective target environments emerge, their respective sizes and the amenability of their required adaptive traits to systematic improvement can guide the priority in which they are targeted. The target environments should be focused on specific land types and regions that are abundant.
- (2)
- Productivity/profitability of row-crops: A high-resolution index (30 m × 30 m) of row-crop productivity in the Midwest U.S. is available via the National Commodity Crop Productivity Index (NCCPI) [138]. This index can be used in a comparative profitability analysis to ascertain if any of the target environments overlap with low production row-crop environments. Productivity alone, however, is insufficient in determining whether an alternative crop can outcompete row-crops under specific conditions. Instead, high-resolution profitability surfaces are now used to evaluate row-crop suitability [4,5]. In this light, it is important to note that the soil and climate suitability of tree crops and row-crops are not necessarily correlated [6].
- (3)
- Provision of regulatory ecosystem services: The prioritization of target environments could proceed based upon specific land types and regions that provide disproportionally large regulatory ecosystem services. Subsequent analysis of suitable map units could prioritize locations based upon slope, erosion, water quality, and more [5,9].
To proceed with breeding efforts, adequate wild germplasm with adaptability traits corresponding to the identified target environment must be acquired and characterized. Once discrete target environments are identified, they can be surveyed for the presence and availability of local wild relatives. If distinct clusters were produced from the cluster analysis, the geo-reference points nearest to the cluster centroids can serve as starting locations for germplasm collection. Cluster centroids can also guide the placement of progeny within target environments including to locations that best represent the collective map units of the cluster.
6. Discussion and Conclusions
Considerable opportunity exists to diversify the agriculture landscape in the Midwest U.S. and, thereby, increase both agricultural productivity and ecological functioning. Crop diversification is particularly pragmatic when viewed through the lens of economically marginal locations. Such marginal lands often coincide with high priority areas for ecological rehabilitation and disproportional potential returns with land-use change [4,139]. While the term “marginal” can imply a general lack of suitability for crop cultivation, in this context, the term is specific to the profitability of the maize-soybean rotation in the Midwest U.S. Consequently, areas considered marginal under this definition could be productive for other crop species especially those that are low-input. Additionally, these areas are compositionally diverse [30] and, in turn, offer a variety of opportunities for the targeted development of alternate crops that impart ecological benefits, of which select tree crops are ideal candidates.
Tree crops introduce a variety of innate ecological functions (e.g., carbon sequestration and run-off filtration) while providing economic incentive to justify their long-term maintenance [7,12]. Recent breeding efforts (Table 1) have stimulated regionally-specific tree crop adoption [11], and, in the process, this work has built a foundational base of germplasm to enable future breeding endeavors. Moreover, wild relatives of these tree crops occupy broad endemic ranges that are inclusive of the Midwest U.S. and offer the genetic resources needed to expand the cultivated range of existing selections throughout the Midwest U.S. However, while existing germplasm collections are accessible, collection gaps are prevalent. Table 3 highlights the severity of these collection gaps by summarizing the number of existing accessions held at either the USDA-Agriculture Research Station or universities and the relative degrees to which these collections represent the endemic range of the species. While closing collection gaps is a lofty pursuit, we proposed a GIS-based workflow to give the collection structure as well as guide the subsequent use of regionally adapted germplasm in breeding programs via decentralized selection.
The broad integration of the tree crops into the marginal lands of the Midwest U.S. agricultural landscape would contribute substantially towards food security and environmental goals especially to climate change mitigation and adaption. The integration of trees into the temperate agricultural landscape drives considerable carbon sequestration in both woody biomass and soil [42,49] as well as the reduction of non-CO2 greenhouse gases [50,51,142]. Tree biomass within the agricultural landscape is an important resource for carbon storage, comprising 75% of carbon stored (34.2 petagrams C) within the global agricultural landscape [143], even though tree cover only occupies more than 10% of space on roughly 40% of agricultural land. In only the temperate Midwest U.S., Udawatta and Jose [144] estimate the sequestration capacity of conservative, low-density tree integration on just 10% of row-crop lands (or 15.4 million ha) at 52.4 Tg C year−1. Despite the promise, the integration of trees into the agricultural landscape is far from optimized and remains an often overlooked medium for which to increase the landscape’s carbon sequestration capacity [143].
Beyond mitigation functions, tree integration provides a portfolio of climate change adaptation functions to the agricultural landscape. Trees adjacent to row-crops can abate the effect of weather and climate on the crops by blocking wind stress [145], moderating air and soil temperatures through shade and evaporative cooling [146], and reducing evaporation of soil moisture [147]. Furthermore, tree-based systems have apparent advantages under increasing interannual variability in rainfall and heat – two focal challenges for future maize yield stability in the Midwest U.S. [31,148]. First, deep-rooted systems have access to larger areas of water and nutrients, which is beneficial under drought conditions. Similar benefits are transferred to adjacent row crops through increased soil porosity from the tree roots that, in turn, improves water infiltration and storage leading into drought periods [149]. Furthermore, trees have higher evapotranspiration rates than row-crops, increasing the capacity for evaporative cooling under extreme heat but also the aeration of soils following periods of excessive rainfall and flooding [150].
Equally important, diversification via tree crops also introduces a level of economic resiliency against interannual climatic variability. Tree fruits and nuts are often of higher value than commodity grains, and, even when integrated at comparatively small scales at the farm-level, tree crop revenues can buffer against the increasing risks of grain crop losses due to climate change. Additionally, while the consumer preference for horticultural crops can be somewhat plastic, the decentralized approach offers an opportunity to amend the breeding pipeline accordingly. Decentralization is used often as a strategy to integrate selective end-user and grower feedback related to marketability into the breeding pipeline. If systematically integrated, decentralization can be an effective tactic to help breeding objectives evolve with consumer and grower preferences.
The vision to broadly develop tree crop germplasm complimentary to the Midwest U.S. agricultural landscape is tangible and timely. New multi-disciplinary and multi-institutional collaborations are needed to coordinate existing resources and leverage the knowledge-base of regional breeders and tree crop stakeholders (e.g., farmers, buyers, processors). Foremost, the collaborators should prioritize suitability mapping (focusing on soil characteristics) of respective tree crops across the geographic range of the Midwest U.S. Prioritizing suitability mapping will create the foundational structure and direction for new collaborations. For example, the mapping will identify priority regions better suited for respective species, and, thus, where to collect germplasm and to seek out farmer participants to house progenies. With this direction, collaborations can justify comprehensive narratives for seeking funding. The vision’s multifunctional outputs and broad scope has the potential to engage a diversity of funding sources, including foundational and private stakeholders and eventually aspire to committed state-level support. While there is ample opportunity to pursue competitive state and federal grants to support components of the program(s), emphasis should be placed on seeking private industry or foundational funding to support the largely applied breeding component that is core for this endeavor.
Author Contributions
Individual contributions were as follows: conceptualization, R.R., S.L., and T.M.; methodology, R.R. and C.M.; investigation, R.R.; resources, R.R. and S.L.; data curation, R.R. and C.M.; writing—original draft preparation, R.R.; writing—review and editing, R.R., S.L., and T.M., and K.W.; supervision, S.L.; and funding acquisition, R.R. and S.L.
Funding
This research was funded by the Illinois Specialty Crop Block Grant.
Acknowledgments
A special thanks to Erik Stanek for being a great lab mate and sounding board.
Conflicts of Interest
The authors declare no conflict of interest.
Appendix A

{kind=link}
Table A1.
Suitability parameters of cultivated hazelnut (adapted from Kidd et al. [135]).
Table A1.
Suitability parameters of cultivated hazelnut (adapted from Kidd et al. [135]).
| Suitability Class | Soil Depth (cm) | pH (0–15cm) | EC (ds/m) (0–15cm) | Clay % (0–15 cm) | Soil Drainage Class | Stone % (>20 cm) | Rainfall, Mean August (mm) |
|---|---|---|---|---|---|---|---|
| Well suited | >50 | 6.5–6.599 | <0.15 | 30–50 | Well to moderate | <10 | <80 |
| Suited | 40–50 | 5.6–6.499 | <0.15 | 30–50 | Imperfect | 10–20 | <50 |
| Marginally suited | 30–40 | 6.6–7.199 | <0.15 | 30–50 | Imperfect | 10–20 | <50 |
| Unsuited | <30 | <5.599 or >7.2 | <0.15 | >50 or <10 | Poor to very poor | >20 | >50 |
The suitability analysis was performed for Illinois using the National Soil Survey Geographic Database (gSSURGO) that provides soil trait data at 10 × 10 m resolution [137]. Only soil characteristics were used in calculating suitability, with rainfall later included with soil traits for a multivariate statistical analysis. For the geospatial analysis, each soil trait was classified into discrete suitability classes, where a given map unit is assigned a value of 1, 0, −1, or −100 based on how that soil trait relates to successful hazelnut production (Table A1). Suitability was calculated as the unweighted sum of the classified soil traits. If one of the soil traits was classified as Unsuited (−100), then the entire map unit was identified as unsuitable.
The entirety of Illinois is included in the spatial analysis, in that all map units with available data for Illinois from gSSURGO were included. The suitability of a given map unit for actual hazelnut production is subject to whether it is located in an agricultural field.
References
- Jose, S. Agroforestry for ecosystem services and environmental benefits: An overview. Agrofor. Syst. 2009, 76, 1–10. [Google Scholar] [CrossRef]
- Torralba, M.; Fagerholm, N.; Burgess, P.J.; Moreno, G.; Plieninger, T. Do European agroforestry systems enhance biodiversity and ecosystem services? A meta-analysis. Agric. Ecosyst. Environ. 2016, 230, 150–161. [Google Scholar] [CrossRef]
- Tsonkova, P.; Böhm, C.; Quinkenstein, A.; Freese, D. Ecological benefits provided by alley cropping systems for production of woody biomass in the temperate region: A review. Agrofor. Syst. 2012, 85, 133–152. [Google Scholar] [CrossRef]
- Brandes, E.; McNunn, G.S.; Schulte, L.A.; Bonner, I.J.; Muth, D.; Babcock, B.A.; Sharma, B.; Heaton, E.A. Subfield profitability analysis reveals an economic case for cropland diversification. Environ. Res. Lett. 2016, 11, 014009. [Google Scholar] [CrossRef] [Green Version]
- Brandes, E.; McNunn, G.S.; Schulte, L.A.; Muth, D.J.; VanLoocke, A.; Heaton, E.A. Targeted subfield switchgrass integration could improve the farm economy, water quality, and bioenergy feedstock production. GCB Bioenergy 2018, 10, 199–212. [Google Scholar] [CrossRef]
- Wolz, K.J.; DeLucia, E.H. Black walnut alley cropping is economically competitive with row crops in the Midwest USA. Ecol. Appl. 2018. [Google Scholar] [CrossRef]
- Lovell, S.T.; Dupraz, C.; Gold, M.; Jose, S.; Revord, R.; Stanek, E.; Wolz, K.J. Temperate agroforestry research: Considering multifunctional woody polycultures and the design of long-term field trials. Agrofor. Syst. 2018, 92, 1397–1415. [Google Scholar] [CrossRef]
- Mattia, C.M.; Lovell, S.T.; Davis, A. Identifying barriers and motivators for adoption of multifunctional perennial cropping systems by landowners in the Upper Sangamon River Watershed, Illinois. Agrofor. Syst. 2018, 92, 1155–1169. [Google Scholar] [CrossRef]
- Mattia, C.M. Identifying Targets for Multifunctional Perennial Cropping Systems Implementation through a Landowner Adoption Preferences Survey and a Spatial Suitability Analysis. Master’s Thesis, University of Illinois at Urbana, Champaign, IL, USA, 2017. [Google Scholar]
- Molnar, T.J.; Kahn, P.C.; Ford, T.M.; Funk, C.J.; Funk, C.R. Tree crops, a permanent agriculture: Concepts from the past for a sustainable future. Resources 2013, 2, 457–488. [Google Scholar] [CrossRef]
- Mori, G.O.; Gold, M.; Jose, S. Specialty Crops in Temperate Agroforestry Systems: Sustainable Management, Marketing and Promotion for the Midwest Region of the USA. In Integrating Landscapes: Agroforestry for Biodiversity Conservation and Food Sovereignty; Springer: Berlin, Germany, 2017; pp. 331–366. [Google Scholar]
- Rhodes, T.K.; Aguilar, F.X.; Jose, S.; Gold, M. Factors influencing the adoption of riparian forest buffers in the Tuttle Creek Reservoir watershed of Kansas, USA. Agrofor. Syst. 2018, 92, 739–757. [Google Scholar] [CrossRef]
- Wolz, K.J.; DeLucia, E.H. Alley cropping: Global patterns of species composition and function. Agric. Ecosyst. Environ. 2018, 252, 61–68. [Google Scholar] [CrossRef]
- Wolz, K.J.; Lovell, S.T.; Branham, B.E.; Eddy, W.C.; Keeley, K.; Revord, R.S.; Wander, M.M.; Yang, W.H.; DeLucia, E.H. Frontiers in alley cropping: Transformative solutions for temperate agriculture. Glob. Chang. Biol. 2018, 24, 883–894. [Google Scholar] [CrossRef]
- Finn, C.E.; Thomas, A.L.; Byers, P.L.; Serçe, S. Evaluation of American (Sambucus canadensis) and European (S. nigra) elderberry genotypes grown in diverse environments and implications for cultivar development. Hortscience 2008, 43, 1385–1391. [Google Scholar] [CrossRef]
- Thomas, A.; Byers, P.; Avery, J.D., Jr.; Kaps, M.; Gu, S. Horticultural performance of eight American elderberry genotypes at three Missouri locations. Acta Hortic. 2015, 1061, 237–244. [Google Scholar] [CrossRef]
- Brand, M. Aronia: Native shrubs with untapped potential. Arnoldia 2010, 67, 14–25. [Google Scholar]
- Brand, M.H. Breeding Better Aronia Plants©. Acta Hortic. 2013, 1055, 269–272. [Google Scholar] [CrossRef]
- Anagnostakis, S.L. Cultivars of chestnuts. Annu. Rep. North. Nut Grow. Assoc. 1999, 90, 16–31. [Google Scholar]
- Hunt, K.L.; Gold, M.A.; Warmund, M.R. Chinese chestnut cultivar performance in Missouri. Acta Hortic. 2004, 693, 145–148. [Google Scholar] [CrossRef]
- Coggeshall, M. Use of microsatellite markers to develop new eastern black walnut cultivars in Missouri, USA. Acta Hortic. 2010, 918, 221–226. [Google Scholar] [CrossRef]
- Reid, W.; Coggeshall, M.V.; Hunt, K.L. Cultivar evaluation and development for black walnut orchards. In Proceedings of the 6th Walnut Council Research Symposium; Gen. Tech. Rep. NC-243; U.S. Department of Agriculture, Forest Service, North Central Research Station: St. Paul, MN, USA, 2004. [Google Scholar]
- Warmund, M.; Coggeshall, M. Flowering and fruit characteristics of black walnut cultivars at the University of Missouri repository. Acta Hortic. 2009, 861, 177–182. [Google Scholar] [CrossRef]
- Molnar, T.J. Corylus. In Wild Crop Relatives: Genomic and Breeding Resources; Springer: Berlin/Heidelberg, Germany, 2011; pp. 15–48. [Google Scholar]
- Molnar, T.J.; Capik, J. Advances in hazelnut research in North America. Acta Hortic. 2012, 940, 57–65. [Google Scholar] [CrossRef]
- Molnar, T.; Lombardoni, J.; Muehlbauer, M.; Honig, J.; Mehlenbacher, S.; Capik, J. Progress breeding for resistance to eastern filbert blight in the eastern United States. Acta Hortic. 2017, 1226, 79–86. [Google Scholar] [CrossRef]
- Grauke, L.; Wood, B.W.; Harris, M.K. Crop vulnerability: Carya. Hortscience 2016, 51, 653–663. [Google Scholar] [CrossRef]
- Thompson, T.E.; Conner, P.J. Pecan. In Fruit Breeding; Springer: Berlin, Germany, 2012; pp. 771–801. [Google Scholar]
- Fischer, R.; Byerlee, D.; Edmeades, G. Crop Yields and Global Food Security; ACIAR: Canberra, Australia, 2014. [Google Scholar]
- Lobell, D.B.; Azzari, G. Satellite detection of rising maize yield heterogeneity in the US Midwest. Environ. Res. Lett. 2017, 12, 014014. [Google Scholar] [CrossRef]
- Lobell, D.B.; Roberts, M.J.; Schlenker, W.; Braun, N.; Little, B.B.; Rejesus, R.M.; Hammer, G.L. Greater sensitivity to drought accompanies maize yield increase in the U.S. Midwest. Science 2014, 344, 516–519. [Google Scholar] [CrossRef]
- Barbier, E.B. Sustaining agriculture on marginal land: A policy framework. Environ. Sci. Policy Sustain. Dev. 1989, 31, 12–40. [Google Scholar] [CrossRef]
- Kang, S.; Post, W.; Wang, D.; Nichols, J.; Bandaru, V.; West, T. Hierarchical marginal land assessment for land use planning. Land Use Policy 2013, 30, 106–113. [Google Scholar] [CrossRef]
- Wiegmann, K.; Hennenberg, K.J.; Fritsche, U.R. Degraded land and sustainable bioenergy feedstock production. In Proceedings of the Joint International Workshop on High Nature Value Criteria and Potential for Sustainable Use of Degraded Lands, Paris, France, 30 June–1 July 2008. [Google Scholar]
- Niu, X.; Duiker, S.W. Carbon sequestration potential by afforestation of marginal agricultural land in the Midwestern US. For. Ecol. Manag. 2006, 223, 415–427. [Google Scholar] [CrossRef]
- Olsen, J. Nutrient Management Guide—Hazelnut; Oregon State University Extension Service: Corvallis, OR, USA, 2001. [Google Scholar]
- Olsen, J. Orchard nutrition. In Growing Hazelnuts in the Pacific Northwest; Oregon State University Extension Service: Corvallis, OR, USA, 2013; pp. 1–5. [Google Scholar]
- Stanek, E.C.; Lovell, S.T.; Reisner, A. Designing multifunctional woody polycultures according to landowner preferences in Central Illinois. Agrofor. Syst. 2019. [Google Scholar] [CrossRef]
- Smith, J.R. Tree Crops: A Permanent Agriculture; Island Press: Washington, DC, USA, 2013. [Google Scholar]
- Technavio. Global Hazelnut Market 2017–2021; EMIS Intelligence Database; Technavio: London, UK, 2017. [Google Scholar]
- Leakey, R.R.B. The role of trees in agroecology and sustainable agriculture in the tropics. Annu. Rev. Phytopathol. 2014, 52, 113–133. [Google Scholar] [CrossRef]
- Mosquera-Losada, M.; Freese, D.; Rigueiro-Rodríguez, A. Carbon sequestration in European agroforestry systems. In Carbon Sequestration Potential of Agroforestry Systems; Springer: Berlin, Germany, 2011; pp. 43–59. [Google Scholar]
- Rigueiro-Rodríguez, A.; Fernández-Núñez, E.; González-Hernández, P.; McAdam, J.; Mosquera-Losada, M.R. Agroforestry systems in Europe: Productive, ecological and social perspectives. In Agroforestry in Europe; Springer: Berlin, Germany, 2009; pp. 43–65. [Google Scholar]
- Lovell, S.T.; Johnston, D.M. Creating multifunctional landscapes: How can the field of ecology inform the design of the landscape? Front. Ecol. Environ. 2009, 7, 212–220. [Google Scholar] [CrossRef]
- Jose, S. Agroforestry for conserving and enhancing biodiversity. Agrofor. Syst. 2012, 85, 1–8. [Google Scholar] [CrossRef]
- Schoeneberger, M.M. Agroforestry: Working trees for sequestering carbon on agricultural lands. Agrofor. Syst. 2009, 75, 27–37. [Google Scholar] [CrossRef]
- Schoeneberger, M.; Bentrup, G.; De Gooijer, H.; Soolanayakanahally, R.; Sauer, T.; Brandle, J.; Zhou, X.; Current, D. Branching out: Agroforestry as a climate change mitigation and adaptation tool for agriculture. J. Soil Water Conserv. 2012, 67, 128A–136A. [Google Scholar] [CrossRef]
- Schoeneberger, M.M.; Bentrup, G.; Patel-Weynand, T. Agroforestry: Enhancing Resiliency in US Agricultural Landscapes under Changing Conditions; Gen. Tech. Report WO-96; US Department of Agriculture, Forest Service: Washington, DC, USA, 2017.
- Udawatta, R.P.; Jose, S. Agroforestry strategies to sequester carbon in temperate North America. Agrofor. Syst. 2012, 86, 225–242. [Google Scholar] [CrossRef]
- Amadi, C.C.; Van Rees, K.C.; Farrell, R.E. Soil–atmosphere exchange of carbon dioxide, methane and nitrous oxide in shelterbelts compared with adjacent cropped fields. Agric. Ecosyst. Environ. 2016, 223, 123–134. [Google Scholar] [CrossRef]
- Kim, D.-G.; Kirschbaum, M.U.; Beedy, T.L. Carbon sequestration and net emissions of CH4 and N2O under agroforestry: Synthesizing available data and suggestions for future studies. Agric. Ecosyst. Environ. 2016, 226, 65–78. [Google Scholar] [CrossRef]
- Udawatta, R.P.; Krstansky, J.J.; Henderson, G.S.; Garrett, H.E. Agroforestry practices, runoff, and nutrient loss. J. Environ. Qual. 2002, 31, 1214–1225. [Google Scholar] [CrossRef]
- Garrett, H.; McGraw, R.; Walter, W. Alley cropping practices. In North American Agroforestry: An Integrated Science and Practice, 2nd ed.; American Society of Agronomy: Madison, WI, USA, 2009; pp. 133–162. [Google Scholar]
- Allen, S.C.; Jose, S.; Nair, P.K.R.; Brecke, B.J.; Nkedi-Kizza, P.; Ramsey, C.L. Safety-net role of tree roots: Evidence from a pecan (Carya illinoensis K. Koch)–cotton (Gossypium hirsutum L.) alley cropping system in the southern United States. For. Ecol. Manag. 2004, 192, 395–407. [Google Scholar] [CrossRef]
- Bambo, S.K.; Nowak, J.; Blount, A.R.; Long, A.J.; Osiecka, A. Soil nitrate leaching in silvopastures compared with open pasture and pine plantation. J. Environ. Qual. 2009, 38, 1870–1877. [Google Scholar] [CrossRef]
- Muth, D. Profitability versus environmental performance: Are they competing? J. Soil Water Conserv. 2014, 69, 203A–206A. [Google Scholar] [CrossRef]
- Lerch, R.; Kitchen, N.; Kremer, R.; Donald, W.; Alberts, E.; Sadler, E.; Sudduth, K.; Myers, D.; Ghidey, F. Development of a conservation-oriented precision agriculture system: Water and soil quality assessment. J. Soil Water Conserv. 2005, 60, 411–421. [Google Scholar]
- Ford-Lloyd, B.V.; Schmidt, M.; Armstrong, S.J.; Barazani, O.; Engels, J.; Hadas, R.; Hammer, K.; Kell, S.P.; Kang, D.; Khoshbakht, K. Crop wild relatives—Undervalued, underutilized and under threat? BioScience 2011, 61, 559–565. [Google Scholar] [CrossRef]
- Khoury, C.K.; Greene, S.; Wiersema, J.; Maxted, N.; Jarvis, A.; Struik, P.C. An inventory of crop wild relatives of the United States. Crop Sci. 2013, 53, 1496–1508. [Google Scholar] [CrossRef]
- Mayes, S.; Massawe, F.J.; Alderson, P.G.; Roberts, J.A.; Azam-Ali, S.N.; Hermann, M. The potential for underutilized crops to improve security of food production. J. Exp. Bot. 2011, 63, 1075–1079. [Google Scholar] [CrossRef] [Green Version]
- Castañeda-Álvarez, N.P.; Khoury, C.K.; Achicanoy, H.A.; Bernau, V.; Dempewolf, H.; Eastwood, R.J.; Guarino, L.; Harker, R.H.; Jarvis, A.; Maxted, N.; et al. Global conservation priorities for crop wild relatives. Nat. Plants 2016, 2, 16022. [Google Scholar] [CrossRef]
- Khoury, C.K.; Greene, S.L.; Williams, K.A.; Sosa, C.; Richards, C. Conservation Priorities for Tree Crop Wild Relatives in the United States. In Proceedings of the Workshop on Gene Conservation of Tree Species–Banking on the Future, Chicago, IL, USA, 16–19 May 2016. [Google Scholar]
- Dempewolf, H.; Baute, G.; Anderson, J.; Kilian, B.; Smith, C.; Guarino, L. Past and future use of wild relatives in crop breeding. Crop Sci. 2017, 57, 1070–1082. [Google Scholar] [CrossRef]
- Prohens, J.; Gramazio, P.; Plazas, M.; Dempewolf, H.; Kilian, B.; Díez, M.J.; Fita, A.; Herraiz, F.J.; Rodríguez-Burruezo, A.; Soler, S.; et al. Introgressiomics: A new approach for using crop wild relatives in breeding for adaptation to climate change. Euphytica 2017, 213, 158. [Google Scholar] [CrossRef]
- Cernusca, M.; Gold, M. Breaking down market barriers for elderberry growers and producers. Acta Hortic. 2013, 1061, 269–277. [Google Scholar] [CrossRef]
- Lee, J.; Finn, C.E. Anthocyanins and other polyphenolics in American elderberry (Sambucus canadensis) and European elderberry (S. nigra) cultivars. J. Sci. Food Agric. 2007, 87, 2665–2675. [Google Scholar] [CrossRef] [Green Version]
- Özgen, M.; Scheerens, J.C.; Reese, R.N.; Miller, R.A. Total phenolic, anthocyanin contents and antioxidant capacity of selected elderberry (Sambucus canadensis L.) accessions. Pharmacogn. Mag. 2010, 6, 198. [Google Scholar] [CrossRef] [Green Version]
- Charlebois, D.; Byers, P.L.; Finn, C.E.; Thomas, A.L. 4 Elderberry: Botany, Horticulture, Potential. Hortic. Rev. 2010, 37, 213. [Google Scholar]
- Chocolate Confectionery: Comparing Ferrero Group Top 20 Countries against the Market 2011–2016. Euromonitor International. 2016. Available online: Statista.com (accessed on 30 November 2017).
- Xie, M.; Zheng, J.; Wang, D. Achievements and perspective in hazelnut breeding in China. Acta Hortic. 2012, 1052, 41–43. [Google Scholar] [CrossRef]
- Grau, P.; Bastias, R. Productivity and yield efficiency of hazelnut (Corylus avellana L.) cultivars in Chile. Acta Hortic. 2004, 686, 57–64. [Google Scholar] [CrossRef]
- Ellena, M.; Sandoval, P.; Gonzalez, A.; Jequier, J.; Contreras, M.; Grau Beretta, P. Chilean hazelnut situation and perspectives. Acta Hortic. 2012, 1052, 329–342. [Google Scholar] [CrossRef]
- Melhenbacher, S. The hazelnut situation in Oregon. Acta Hortic. 2004, 686, 665–668. [Google Scholar] [CrossRef]
- Baldwin, B.J. The Growth and Productivity of Hazelnut Cultivars (Corylus avellana L.) in Australia. Ph.D. Thesis, University of Sydney, Faculty of Rural Management, Sydney, Australia, 2015. [Google Scholar]
- Gönenc, S.; Tanrıvermis, H.; Bülbül, M. Economic assessment of hazelnut production and the importance of supply management approaches in Turkey. J. Agric. Rural Dev. Trop. Subtrop. (JARTS) 2006, 107, 19–32. [Google Scholar]
- Mehlenbacher, S.A. Genetic resources for hazelnut: State of the art and future perspectives. Acta Hortic. 2008, 845, 33–38. [Google Scholar] [CrossRef]
- Ustaoğlu, B. The Effect of Climatic Conditions on Hazelnut (Corylus avellana) Yield in Giresun (Turkey). 2012. Available online: http://hdl.handle.net/11424/2685 (accessed on 20 March 2018).
- FAOSTAT, Food and Agriculture Organization of the United Nations Cropping Database; FAO: Rome, Italy, 2014.
- Dempewolf, H.; Eastwood, R.J.; Guarino, L.; Khoury, C.K.; Müller, J.V.; Toll, J. Adapting Agriculture to Climate Change: A Global Initiative to Collect, Conserve, and Use Crop Wild Relatives. Agroecol. Sustain. Food Syst. 2014, 38, 369–377. [Google Scholar] [CrossRef] [Green Version]
- Kole, C.; Muthamilarasan, M.; Henry, R.; Edwards, D.; Sharma, R.; Abberton, M.; Batley, J.; Bentley, A.; Blakeney, M.; Bryant, J.; et al. Application of genomics-assisted breeding for generation of climate resilient crops: Progress and prospects. Front. Plant Sci. 2015, 6, 563. [Google Scholar] [CrossRef]
- Warschefsky, E.; Varma Penmetsa, R.; Cook, D.R.; Von Wettberg, E.J.B. Back to the wilds: Tapping evolutionary adaptations for resilient crops through systematic hybridization with crop wild relatives. Am. J. Bot. 2014, 101, 1791–1800. [Google Scholar] [CrossRef] [Green Version]
- Miller, A.J.; Gross, B.L. From forest to field: Perennial fruit crop domestication. Am. J. Bot. 2011, 98, 1389–1414. [Google Scholar] [CrossRef] [Green Version]
- Molnar, T.J.; Honig, J.A.; Mayberry, A.; Revord, R.S.; Lovell, S.T.; Mehlenbacher, S.A.; Capik, J.M. Corylus americana: A valuable genetic resource for developing hazelnuts adapted to the eastern United States. In Proceedings of the IX International Congress on Hazelnut, Atakum, Samsun, Turkey, 15–19 August 2017; Tuncer, P.D.C., Ed.; IX International Congress on Hazelnut—Book of Abstracts. Atakum, Samsun, Turkey; pp. 115–122. [Google Scholar]
- Migicovsky, Z.; Myles, S. Exploiting wild relatives for genomics-assisted breeding of perennial crops. Front. Plant Sci. 2017, 8, 460. [Google Scholar] [CrossRef]
- McClure, K.A.; Sawler, J.; Gardner, K.M.; Money, D.; Myles, S. Genomics: A potential panacea for the perennial problem. Am. J. Bot. 2014, 101, 1780–1790. [Google Scholar] [CrossRef] [Green Version]
- Taheri, R.; Connolly, B.A.; Brand, M.H.; Bolling, B.W. Underutilized chokeberry (Aronia melanocarpa, Aronia arbutifolia, Aronia prunifolia) accessions are rich sources of anthocyanins, flavonoids, hydroxycinnamic acids, and proanthocyanidins. J. Agric. Food Chem. 2013, 61, 8581–8588. [Google Scholar] [CrossRef]
- Brand, M.H.; Connolly, B.A.; Levine, L.H.; Richards, J.T.; Shine, S.M.; Spencer, L.E. Anthocyanins, total phenolics, ORAC and moisture content of wild and cultivated dark-fruited Aronia species. Sci. Hortic. 2017, 224, 332–342. [Google Scholar] [CrossRef]
- Alexander, M.T. Conservation of Castanea Dentata Genetic Resources of the Southeastern United States; University of Tennessee at Chattanooga, Environmental Science: Chattanooga, TN, USA, 2005. [Google Scholar]
- Li, G.-T.; Ai, C.-X.; Zhang, L.-S.; Wei, H.-R.; Liu, Q.-Z. Chestnut genebank in China national clonal plant germplasm repository. Acta Hortic. 2009, 844, 199–206. [Google Scholar]
- Miller, A.C.; Woeste, K.E.; Anagnostakis, S.L.; Jacobs, D.F. Exploration of a rare population of Chinese chestnut in North America: Stand dynamics, health and genetic relationships. AoB Plants 2014, 6, plu065. [Google Scholar] [CrossRef]
- McGranahan, G.; Leslie, C. Breeding walnuts (Juglans regia). In Breeding Plantation Tree Crops: Temperate Species; Springer: Berlin, Germany, 2009; pp. 249–273. [Google Scholar]
- Npgs, U. Genetic Resources Information Service. In GRIN Taxonomy. Available online: https://npgsweb.ars-grin.gov/gringlobal/taxon/taxonomysearch.aspx (accessed on 30 May 2018).
- Thompson, M.M.; Lagerstedt, H.B.; Mehlenbacher, S.A. Hazelnuts. Fruit Breed. 1996, 3, 125–184. [Google Scholar]
- Sathuvalli, V.R.; Mehlenbacher, S.A. Characterization of American hazelnut (Corylus americana) accessions and Corylus americana × Corylus avellana hybrids using microsatellite markers. Genet. Resour. Crop Evol. 2012, 59, 1055–1075. [Google Scholar] [CrossRef]
- Demchik, M.; Kern, A.; Braun, L.; Fischbach, J.; Turnquist, K. Genetic diversity of American hazelnut in the Upper Midwest, USA. Agrofor. Syst. 2018, 92, 1507–1516. [Google Scholar] [CrossRef]
- Molnar, T.J.; Lombardoni, J.J.; Muehlbauer, M.F.; Honig, J.A.; Mehlenbacher, S.A.; Capik, J.M. Progress breeding for resistance to eastern filbert blight in the eastern United States. In Proceedings of the IX International Congress on Hazelnut, Atakum, Samsun, Turkey, 15–19 August 2017; Tuncer, P.D.C., Ed.; IX International Congress on Hazelnut—Book of Abstracts. Atakum, Samsun, Turkey, 2018; pp. 79–86. [Google Scholar]
- Drumke, J.S. A Systematic Survey of Corylus in North America; The University of Tennessee: Knoxville, TN, USA, 1964. [Google Scholar]
- Capik, J.M.; Molnar, T.J. Assessment of host (Corylus sp.) resistance to eastern filbert blight in New Jersey. J. Am. Soc. Hortic. Sci. 2012, 137, 157–172. [Google Scholar] [CrossRef]
- Fuller, A.S. The Nut Culturist; Orange Judd: New York, NY, USA, 1908. [Google Scholar]
- Weschcke, C. Growing Nuts in the North; Webb: St. Paul, MN, USA, 1953. [Google Scholar]
- Johnson, K.; Pinkerton, J. Eastern filbert blight. In Compendium of Nut Crop Diseases in Temperate Zones; APS Press: St. Paul, MN, USA, 2002; pp. 44–46. [Google Scholar]
- Muehlbauer, M.; Tobia, J.; Honig, J.; Hillman, B.; Zhang, N.; Gold, K.M.; Molnar, T. Population differentiation within Anisogramma anomala in North America. Phytopathology 2018. [Google Scholar] [CrossRef]
- Erdogan, V.; Mehlenbacher, S.A. Interspecific hybridization in hazelnut (Corylus). J. Am. Soc. Hortic. Sci. 2000, 125, 489–497. [Google Scholar] [CrossRef]
- Bhattarai, G.; Mehlenbacher, S.A.; Smith, D.C. Eastern filbert blight disease resistance from Corylus americana ‘Rush’ and selection ‘Yoder #5’ maps to linkage group 7. Tree Genet. Genomes 2017, 13, 45. [Google Scholar]
- Cornille, A.; Gladieux, P.; Smulders, M.J.; Roldan-Ruiz, I.; Laurens, F.; Le Cam, B.; Nersesyan, A.; Clavel, J.; Olonova, M.; Feugey, L. New insight into the history of domesticated apple: Secondary contribution of the European wild apple to the genome of cultivated varieties. PLoS Genet. 2012, 8, e1002703. [Google Scholar] [CrossRef]
- Myles, S.; Boyko, A.R.; Owens, C.L.; Brown, P.J.; Grassi, F.; Aradhya, M.K.; Prins, B.; Reynolds, A.; Chia, J.-M.; Ware, D. Genetic structure and domestication history of the grape. Proc. Natl. Acad. Sci. USA 2011, 108, 3530–3535. [Google Scholar] [CrossRef] [Green Version]
- Migicovsky, Z.; Sawler, J.; Money, D.; Eibach, R.; Miller, A.J.; Luby, J.J.; Jamieson, A.R.; Velasco, D.; von Kintzel, S.; Warner, J.; et al. Genomic ancestry estimation quantifies use of wild species in grape breeding. BMC Genom. 2016, 17, 478. [Google Scholar] [CrossRef]
- Öztürk, S.C.; Balık, H.İ.; Balık, S.K.; Kızılcı, G.; Duyar, Ö.; Doğanlar, S.; Frary, A. Molecular genetic diversity of the Turkish national hazelnut collection and selection of a core set. Tree Genet. Genomes 2017, 13, 113. [Google Scholar] [CrossRef]
- Ellstrand, N.C.; Prentice, H.C.; Hancock, J.F. Gene flow and introgression from domesticated plants into their wild relatives. Annu. Rev. Ecol. Syst. 1999, 30, 539–563. [Google Scholar] [CrossRef]
- Maxted, N.; Kell, S.; Ford-Lloyd, B.; Dulloo, E.; Toledo, Á. Toward the Systematic Conservation of Global Crop Wild Relative Diversity. Crop Sci. 2012, 52, 774–785. [Google Scholar] [CrossRef]
- Brumlop, S.; Reichenbecher, W.; Tappeser, B.; Finckh, M.R. What is the SMARTest way to breed plants and increase agrobiodiversity? Euphytica 2013, 194, 53–66. [Google Scholar] [CrossRef]
- Duminil, J.; Fineschi, S.; Hampe, A.; Jordano, P.; Salvini, D.; Vendramin, G.G.; Petit, R.J. Can population genetic structure be predicted from life-history traits? Am. Nat. 2007, 169, 662–672. [Google Scholar]
- Duminil, J.; Hardy, O.J.; Petit, R.J. Plant traits correlated with generation time directly affect inbreeding depression and mating system and indirectly genetic structure. BMC Evolut. Biol. 2009, 9, 177. [Google Scholar] [CrossRef]
- Hamrick, J.L.; Godt, M.J.W.; Sherman-Broyles, S.L. Factors influencing levels of genetic diversity in woody plant species. In Population Genetics of Forest Trees; Springer: Berlin, Germany, 1992; pp. 95–124. [Google Scholar]
- Loveless, M.D.; Hamrick, J.L. Ecological determinants of genetic structure in plant populations. Annu. Rev. Ecol. Syst. 1984, 15, 65–95. [Google Scholar] [CrossRef]
- Petit, R.J.; Hampe, A. Some Evolutionary Consequences of Being a Tree. Annu. Rev. Ecol. Evol. Syst. 2006, 37, 187–214. [Google Scholar] [CrossRef] [Green Version]
- Hedrick, P.W. Genetic polymorphism in heterogeneous environments: The age of genomics. Annu. Rev. Ecol. Evol. Syst. 2006, 37, 67–93. [Google Scholar] [CrossRef]
- Kawecki, T.J.; Ebert, D. Conceptual issues in local adaptation. Ecol. Lett. 2004, 7, 1225–1241. [Google Scholar] [CrossRef] [Green Version]
- Linhart, Y.B.; Grant, M.C. Evolutionary significance of local genetic differentiation in plants. Annu. Rev. Ecol. Syst. 1996, 27, 237–277. [Google Scholar] [CrossRef]
- Pritchard, J.K.; Di Rienzo, A. Adaptation—Not by sweeps alone. Nat. Rev. Gen. 2010, 11, 665. [Google Scholar] [CrossRef]
- Le Corre, V.; Kremer, A. The genetic differentiation at quantitative trait loci under local adaptation. Mol. Ecol. 2012, 21, 1548–1566. [Google Scholar] [CrossRef] [Green Version]
- Aitken, S.N.; Yeaman, S.; Holliday, J.A.; Wang, T.; Curtis-McLane, S. Adaptation, migration or extirpation: Climate change outcomes for tree populations. Evol. Appl. 2008, 1, 95–111. [Google Scholar] [CrossRef]
- Savolainen, O.; Lascoux, M.; Merilä, J. Ecological genomics of local adaptation. Nat. Rev. Gen. 2013, 14, 807. [Google Scholar] [CrossRef]
- Neale, D.B.; Savolainen, O. Association genetics of complex traits in conifers. Trends Plant Sci. 2004, 9, 325–330. [Google Scholar] [CrossRef] [Green Version]
- Brown, G.R.; Gill, G.P.; Kuntz, R.J.; Langley, C.H.; Neale, D.B. Nucleotide diversity and linkage disequilibrium in loblolly pine. Proc. Natl. Acad. Sci. USA 2004, 101, 15255–15260. [Google Scholar] [CrossRef] [Green Version]
- Heuertz, M.; De Paoli, E.; Källman, T.; Larsson, H.; Jurman, I.; Morgante, M.; Lascoux, M.; Gyllenstrand, N. Multilocus patterns of nucleotide diversity, linkage disequilibrium and demographic history of Norway spruce (Picea abies (L.) Karst). Genetics 2006, 174, 2095–2105. [Google Scholar] [CrossRef]
- Varshney, R.K.; Terauchi, R.; McCouch, S.R. Harvesting the promising fruits of genomics: Applying genome sequencing technologies to crop breeding. PLoS Biol. 2014, 12, e1001883. [Google Scholar] [CrossRef]
- Lijavetzky, D.; Cabezas, J.A.; Ibáñez, A.; Rodríguez, V.; Martínez-Zapater, J.M. High throughput SNP discovery and genotyping in grapevine (Vitis vinifera L.) by combining a re-sequencing approach and SNPlex technology. BMC Genom. 2007, 8, 424. [Google Scholar] [CrossRef]
- Parchman, T.L.; Gompert, Z.; Mudge, J.; Schilkey, F.D.; Benkman, C.W.; Buerkle, C.A. Genome-wide association genetics of an adaptive trait in lodgepole pine. Mol. Ecol. 2012, 21, 2991–3005. [Google Scholar] [CrossRef]
- Sork, V.; Aitken, S.; Dyer, R.; Eckert, A.; Legendre, P.; Neale, D. Putting the landscape into the genomics of trees: Approaches for understanding local adaptation and population responses to changing climate. Tree Genet. Genomes 2013, 9, 901–911. [Google Scholar] [CrossRef]
- Simmonds, N. Selection for local adaptation in a plant breeding programme. Theor. Appl. Genet. 1991, 82, 363–367. [Google Scholar] [CrossRef]
- Ceccarelli, S. Efficiency of plant breeding. Crop Sci. 2015, 55, 87–97. [Google Scholar] [CrossRef]
- Atlin, G.N. Breeding for Suboptimal Environments. In Proceedings of the International Workshop on Increased Lowland Rice Production in the Mekong Region, Vientiane, Laos, 30 October–2 November 2000; Fukai, S., Basnayake, J., Eds.; ACIAR: Canberra, Australia; pp. 245–251. [Google Scholar]
- Annicchiarico, P.; Bellah, F.; Chiari, T. Defining subregions and estimating benefits for a specific-adaptation strategy by breeding programs. Crop Sci. 2005, 45, 1741–1749. [Google Scholar] [CrossRef]
- Kidd, D.; Webb, M.; Malone, B.; Minasny, B.; McBratney, A. Digital soil assessment of agricultural suitability, versatility and capital in Tasmania, Australia. Geoderma Reg. 2015, 6, 7–21. [Google Scholar] [CrossRef]
- Wallace, D.C.; Young, F.J. Black walnut suitability index: A natural resources conservation service national soil information system based interpretive model. In Proceedings of the 16th Central Hardwood Forest Conference, West Lafayette, IN, USA, 8–9 April 2008; Gen. Tech. Rep. NRS-P-24. US Department of Agriculture, Forest Service, Northern Research Station: Newtown Square, PA, USA, 2008; pp. 589–595. [Google Scholar]
- Staff, S.S. Soil Survey Geographic (SSURGO) Database for Illinois; Natural Resources Conservation Service, United States Department of Agriculture: Washington, DC, USA, 2017. [Google Scholar]
- Dobos, R.; Sinclair, H.; Hipple, K. National Commodity Crop Productivity Index (NCCPI) User Guide v2. 0; USDA NRCS National Soil Survey Center: Lincoln, NE, USA, 2012.
- Richards, B.K.; Stoof, C.R.; Cary, I.J.; Woodbury, P.B. Reporting on marginal lands for bioenergy feedstock production: A modest proposal. BioEnergy Res. 2014, 7, 1060–1062. [Google Scholar] [CrossRef]
- Bushakra, J.; Bassil, N.; Finn, C.; Hummer, K. Sambucus Genetic Resources at the US National Clonal Germplasm Repository. I Int. Symp. Elderberry 2013, 1061, 135–145. [Google Scholar]
- Byers, P.; Thomas, A. Elderberry research and production in Missouri. Proc. Mo. Small Fruit Veg. Conf. 2005, 25, 91–97. [Google Scholar]
- Wolz, K.J.; Branham, B.E.; DeLucia, E.H. Reduced nitrogen losses after conversion of row crop agriculture to alley cropping with mixed fruit and nut trees. Agric. Ecosyst. Environ. 2018, 258, 172–181. [Google Scholar] [CrossRef]
- Zomer, R.J.; Neufeldt, H.; Xu, J.; Ahrends, A.; Bossio, D.; Trabucco, A.; van Noordwijk, M.; Wang, M. Global Tree Cover and Biomass Carbon on Agricultural Land: The contribution of agroforestry to global and national carbon budgets. Sci. Rep. 2016, 6, 29987. [Google Scholar] [CrossRef] [Green Version]
- Udawatta, R.P.; Jose, S. Carbon sequestration potential of agroforestry practices in temperate North America. In Carbon Sequestration Potential of Agroforestry Systems; Springer: Berlin, Germany, 2011; pp. 17–42. [Google Scholar]
- Böhm, C.; Kanzler, M.; Freese, D. Wind speed reductions as influenced by woody hedgerows grown for biomass in short rotation alley cropping systems in Germany. Agrofor. Syst. 2014, 88, 579–591. [Google Scholar] [CrossRef]
- Lin, B.B. Agroforestry management as an adaptive strategy against potential microclimate extremes in coffee agriculture. Agric. For. Meteorol. 2007, 144, 85–94. [Google Scholar] [CrossRef]
- Siriri, D.; Wilson, J.; Coe, R.; Tenywa, M.; Bekunda, M.; Ong, C.; Black, C. Trees improve water storage and reduce soil evaporation in agroforestry systems on bench terraces in SW Uganda. Agrofor. Syst. 2013, 87, 45–58. [Google Scholar] [CrossRef]
- Lobell, D.B.; Hammer, G.L.; McLean, G.; Messina, C.; Roberts, M.J.; Schlenker, W. The critical role of extreme heat for maize production in the United States. Nat. Clim. Change 2013, 3, 497–501. [Google Scholar] [CrossRef]
- Anderson, S.H.; Udawatta, R.P.; Seobi, T.; Garrett, H.E. Soil water content and infiltration in agroforestry buffer strips. Agrofor. Syst. 2009, 75, 5–16. [Google Scholar] [CrossRef]
- Verchot, L.V.; Van Noordwijk, M.; Kandji, S.; Tomich, T.; Ong, C.; Albrecht, A.; Mackensen, J.; Bantilan, C.; Anupama, K.; Palm, C. Climate change: Linking adaptation and mitigation through agroforestry. Mitig. Adapt. Strateg. Glob. Change 2007, 12, 901–918. [Google Scholar] [CrossRef]
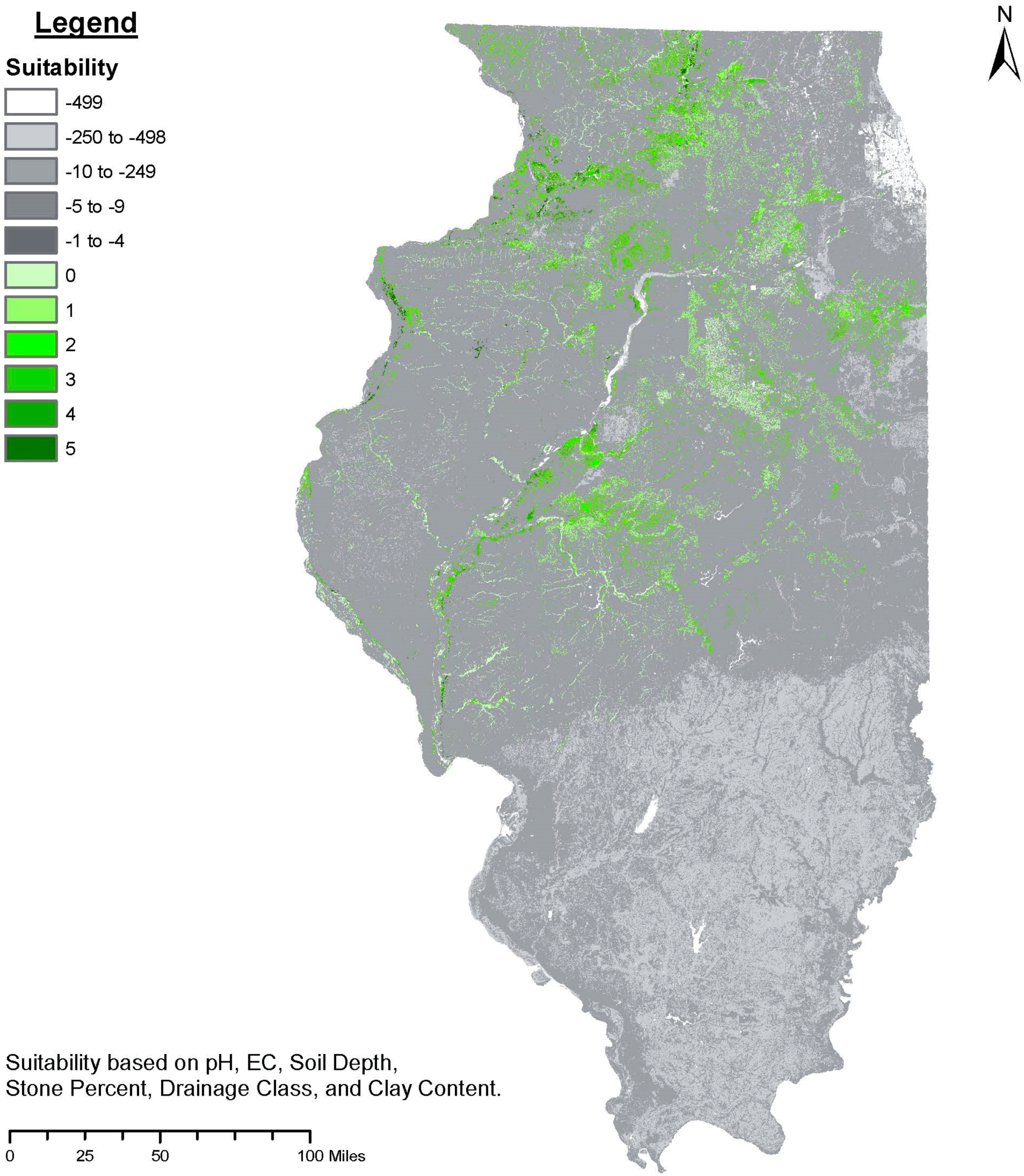
Figure 1.
A suitability analysis of the soil characteristics relevant to the successful cultivation of the European hazelnut Corylus avellana in the state of Illinois in the Midwest U.S. Soil characteristics and corresponding discrete suitability classes are listed in Table A1. Suitability rankings were not weighted, and details of the analysis can be found in Appendix A. Green shaded map units indicate that all six soil characteristics were classified as suited or well-suited. Grey shading indicates marginal or unsuited map units, with rankings below zero.
Figure 1.
A suitability analysis of the soil characteristics relevant to the successful cultivation of the European hazelnut Corylus avellana in the state of Illinois in the Midwest U.S. Soil characteristics and corresponding discrete suitability classes are listed in Table A1. Suitability rankings were not weighted, and details of the analysis can be found in Appendix A. Green shaded map units indicate that all six soil characteristics were classified as suited or well-suited. Grey shading indicates marginal or unsuited map units, with rankings below zero.

Table 1.
Progress to date in breeding underutilized tree crops with relevance to the Midwest U.S.
| Tree Crop | Cultivated Species | Breeding Objective | Breeding Stage | Adapted Regions | References |
|---|---|---|---|---|---|
| Elderberry | Sambucus nigra L. subspecies canadensis R. Bolli | Identify adapted varieties, selection for site specific conditions, fruit quality and yield, reduced inter-annual variability, and late bud break. | Multi-location trials of traditional varieties and new germplasm. | MO. | [15,16] |
| Aronia | Aronia melanocarpa (Michx.) Elliot | Identify adapted varieties, narrower and shorter growth habit, total phenolics and anthocyanins, total yield, and low chilling. | Trials of traditional varieties and new germplasm. | CT, the NE. | [17,18] |
| Chinese chestnut | Castanea mollissima Blume | Identify adapted varieties, kernel size and quality, yield quantity and consistency. | Variety trials; initiated pedigree breeding. | CT; MO. | [11,19,20] |
| Eastern black walnut | Juglans nigra L. | Identify adapted varieties, kernel quality and yield (e.g., nutmeat/shell ratio), alternate bearing, define host resistance to pests and disease, early flowering, and spur-type growth habit. | Multi-location variety trials; pedigree breeding. | MO; KS. | [21,22,23] |
| European hazelnut | Corylus avellana | Eastern filbert blight resistance, cold hardiness, commercial kernel quality and yield. | Screening wild germplasm; modified backcrossing. | NJ; NY. | [24,25,26] |
| Northern Pecan | Carya illinoinensis (Wangenh) K. Koch | High yield, perocity, kernel quality, kernel percentage, ease of shelling, disease resistance, and reduced masting. | Multi-location trials; controlled crossing. | GA; MO; TX. | [27,28] |
Table 2.
Wild relatives and wild utilized species with traits of a known value.
| Species | Known Traits of Value in Wild Relatives 1 |
|---|---|
| Sambucus nigra L. subspecies canadensis R. Bolli | Commercial yields, late flowering, short ripening period, [15], high phenolics and anthocyanins, acylated forms of cyanidin-glycosides, and regional and local adaptation [16]. |
| Aronia arbutifolia (L.) Pers. | Lower chill hours, fruit ripening date, fruit size, and ripe fruit color [18,86]. |
| Aronia melanocarpa (Michx.) Elliot | High anthocyanin and unique profiles [86,87], plant habit (e.g., prostrate), fruit ripening date, fruit size [18], and diverse microclimate adaptation [17]. |
| Aronia prunifolia (Marshall) Rehder | High phenolics [86,87]. |
| Castanea dentata (Marshall) Borkh. | Regional and local adaptation along with rare genetic diversity [88]. |
| Castanea mollissima Blume 2 | Resistance to chestnut blight, winter hardiness, kernel size and quality, yield quantity, late bud break, flowering date, early nut maturity [20,89], naturalize populations [90], and some regional and local adaptation [20]. |
| Juglans nigra L. | Local adaptation, late flowering, anthracnose resistance [91], and rootstock [92]. |
| Corylus americana Walter | Eastern filbert blight resistance [93], local adaptation [24], cold hardiness [94], heat and drought tolerance [24], and genetic diversity [94,95]. |
| Carya illinoinensis (Wangenh) K. Koch | Disease resistance, drought and heat tolerance, cold hardiness, and tree size reduction [27]. |
1 Traits were synthesized from the referenced literature and adapted in part from Khoury et al. [59]. 2 The Chinese chestnut is included because populations of the endemic chestnut (Castanea dentata) have been severely reduced due to Cryphonectria parasitica, and, as a consequence, the Chinese chestnut has been introduced and naturalized in the eastern U.S. [90]. Performance trials of Chinese chestnut accessions also demonstrate suitable adaptability to the Eastern U.S. [19,20].
Table 3.
Collected tree crop wild relatives and wild utilized species endemic to the Eastern U.S.
| Species | Accessions | States 1 Represented in Endemic Range | States Represented in Germplasm Collections | ||
|---|---|---|---|---|---|
| USDA-ARS 2 | University 3 | Populations Collected | |||
| >10 | 1 to 10 | ||||
| Sambucus nigra L. subspecies canadensis R. Bolli | 38 4 | 55 5 | 52 | 1 | 8 |
| Aronia arbutifolia (L.) Pers. | 20 | 19 6 | 29 | 0 | 13 |
| Aronia melanocarpa (Michx.) Elliot | 50 | 57 6 | 36 | 1 | 16 |
| Aronia prunifolia (Marshall) Rehder | 28 | 41 6 | 31 | 2 | 6 |
| Castanea dentata (Marshall) Borkh. | 1 | Not published 7 | 30 | - | - |
| Castanea mollissima Blume 8 | 239 9 | 65 10 | N/A | N/A | N/A |
| Juglans nigra L. | 27 | 64 | 44 | 0 | 8 |
| Corylus americana Walter | 43 | ~100 11 | 39 | 2 | 27 |
| Carya illinoinensis (Wangenh) K. Koch | 3615 | 21 | Data not available | ||
1 U.S. states and Canadian territories. 2 Data was obtained from the USDA-ARS Germplasm Resources Information Network (GRIN) taxonomy for plants database [92]. 3 Figures are based on those collections in which published records were available in the literature. 4 [140]. 5 [141]. 6 [18]. 7 The American Chestnut Foundation has conducted a longstanding backcrossing program to develop blight resistant Castanea dentata. Other germplasm collections also exist at some volume at the University of Connecticut and the University of Tennessee [88]. 8 The Chinese chestnut is included because populations of the endemic chestnut (Castanea dentata) have been severely reduced due to Cryphonectria parasitica, and, as a consequence, the Chinese chestnut has been introduced and naturalized in the Eastern U.S. [90]. Performance trials of Chinese chestnut accessions also demonstrate suitable adaptability to the Eastern U.S. [19,20]. 9 [89]. 10 [11,20]. 11 [94].
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Revord, R.; Lovell, S.; Molnar, T.; Wolz, K.J.; Mattia, C. Germplasm Development of Underutilized Temperate U.S. Tree Crops. Sustainability 2019, 11, 1546. https://doi.org/10.3390/su11061546
AMA Style
Revord R, Lovell S, Molnar T, Wolz KJ, Mattia C. Germplasm Development of Underutilized Temperate U.S. Tree Crops. Sustainability. 2019; 11(6):1546. https://doi.org/10.3390/su11061546
Chicago/Turabian StyleRevord, Ronald, Sarah Lovell, Thomas Molnar, Kevin J. Wolz, and Chloé Mattia. 2019. "Germplasm Development of Underutilized Temperate U.S. Tree Crops" Sustainability 11, no. 6: 1546. https://doi.org/10.3390/su11061546
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.