Co-Activation, Estimated Anterior and Posterior Cruciate Ligament Forces, and Motor Unit Activation Strategies during the Time Course of Fatigue

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Material & Methods
2.1. Subjects
2.2. Protocol
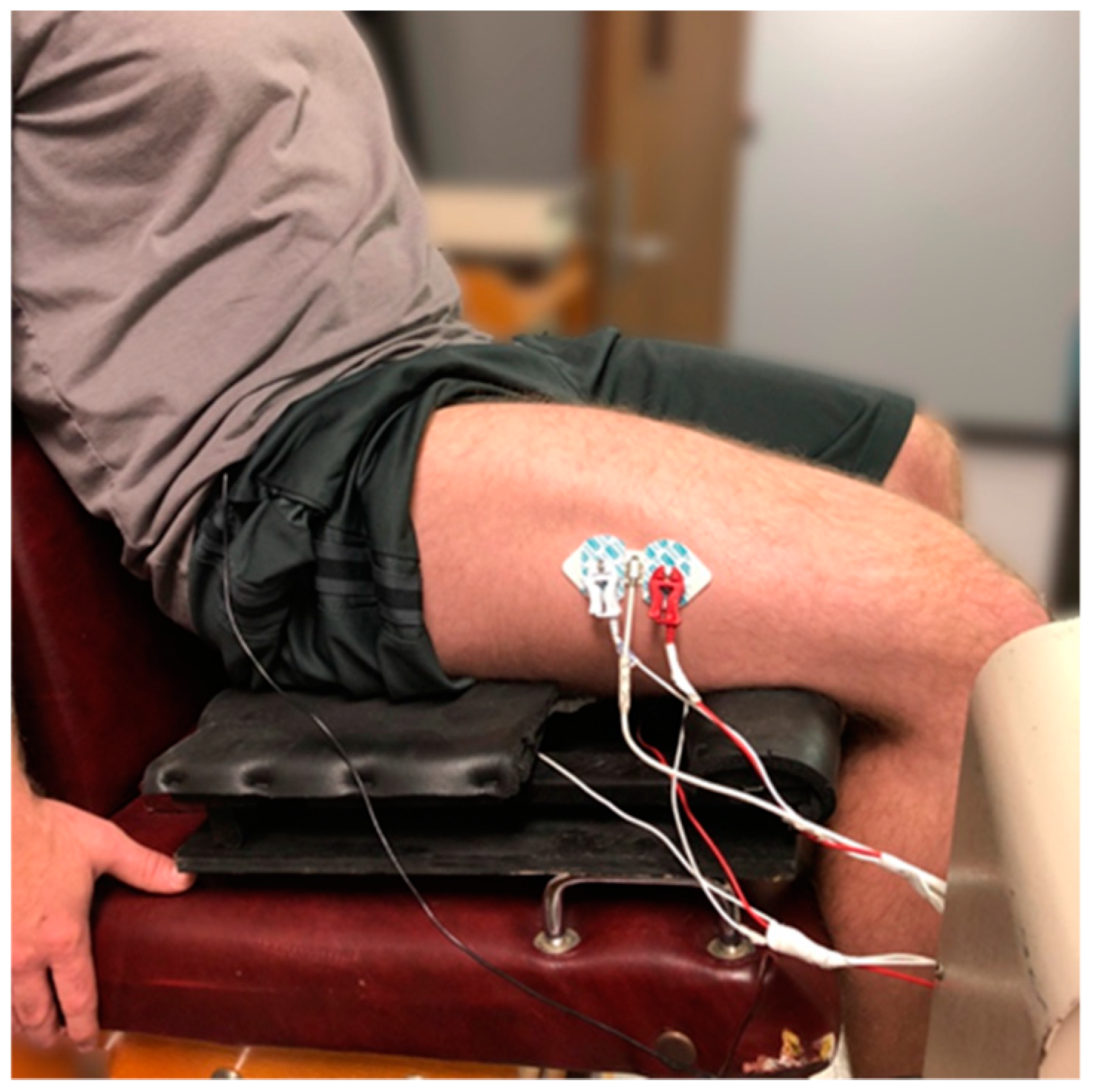
2.3. Electromyographic, Mechanomyographic, and Force Signal Acquisition
2.4. Estimated Anterior and Posterior Cruciate Ligament Knee Forces
2.5. Calculation of Antagonist Force
2.6. Statistical Analysis
3. Results
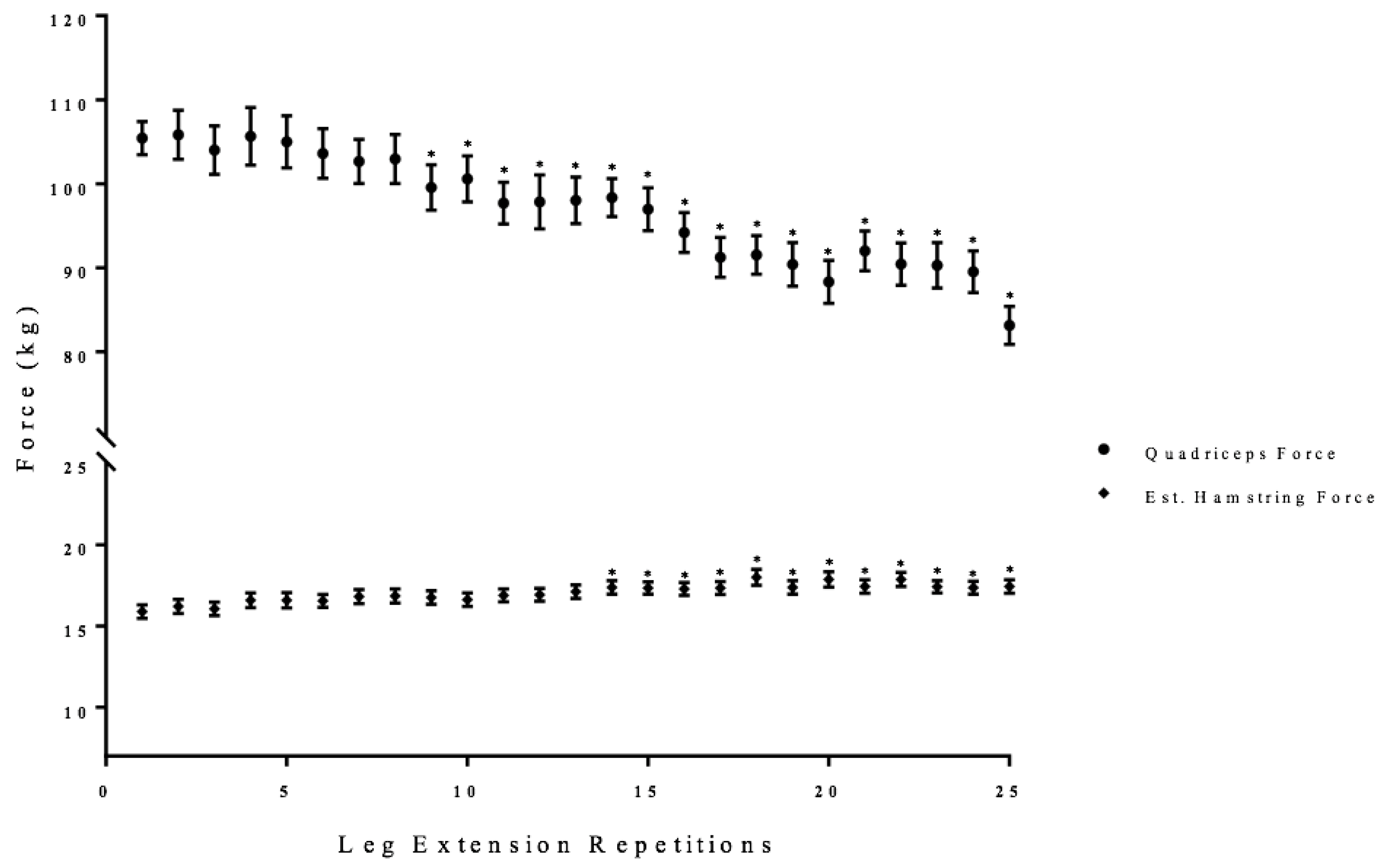
3.1. Leg Extension Quadriceps and Estimated Hamstring Forces
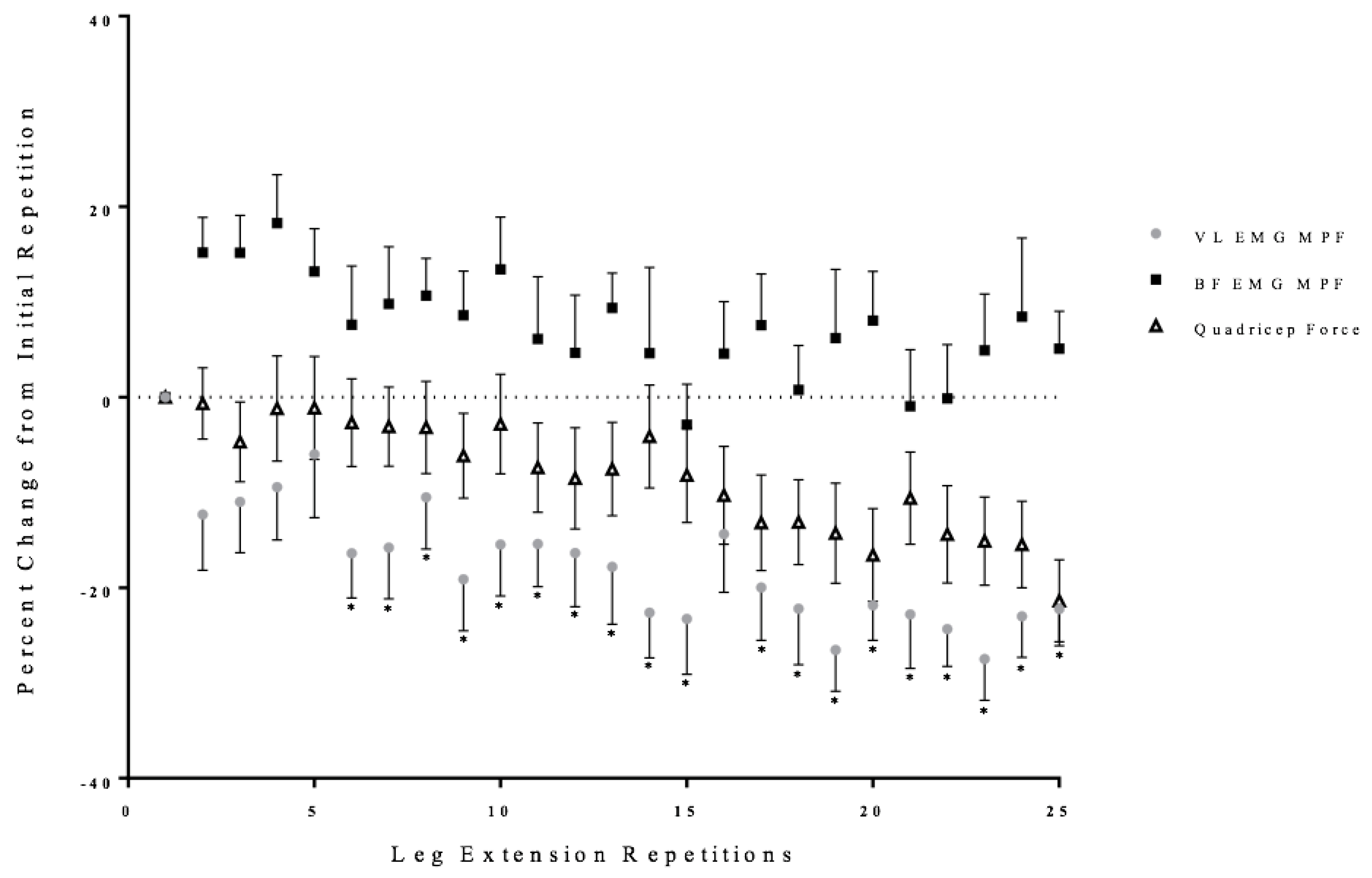
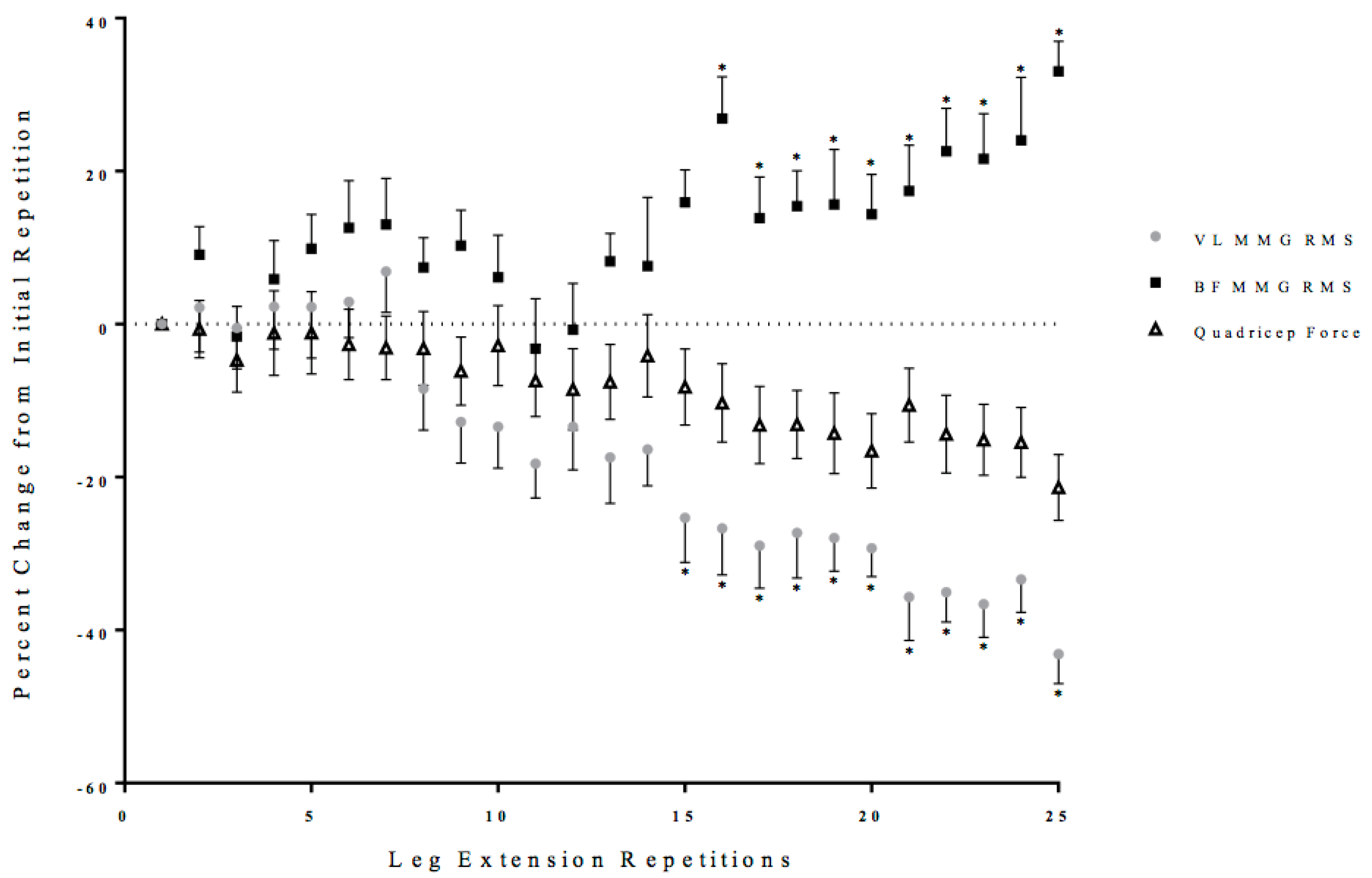
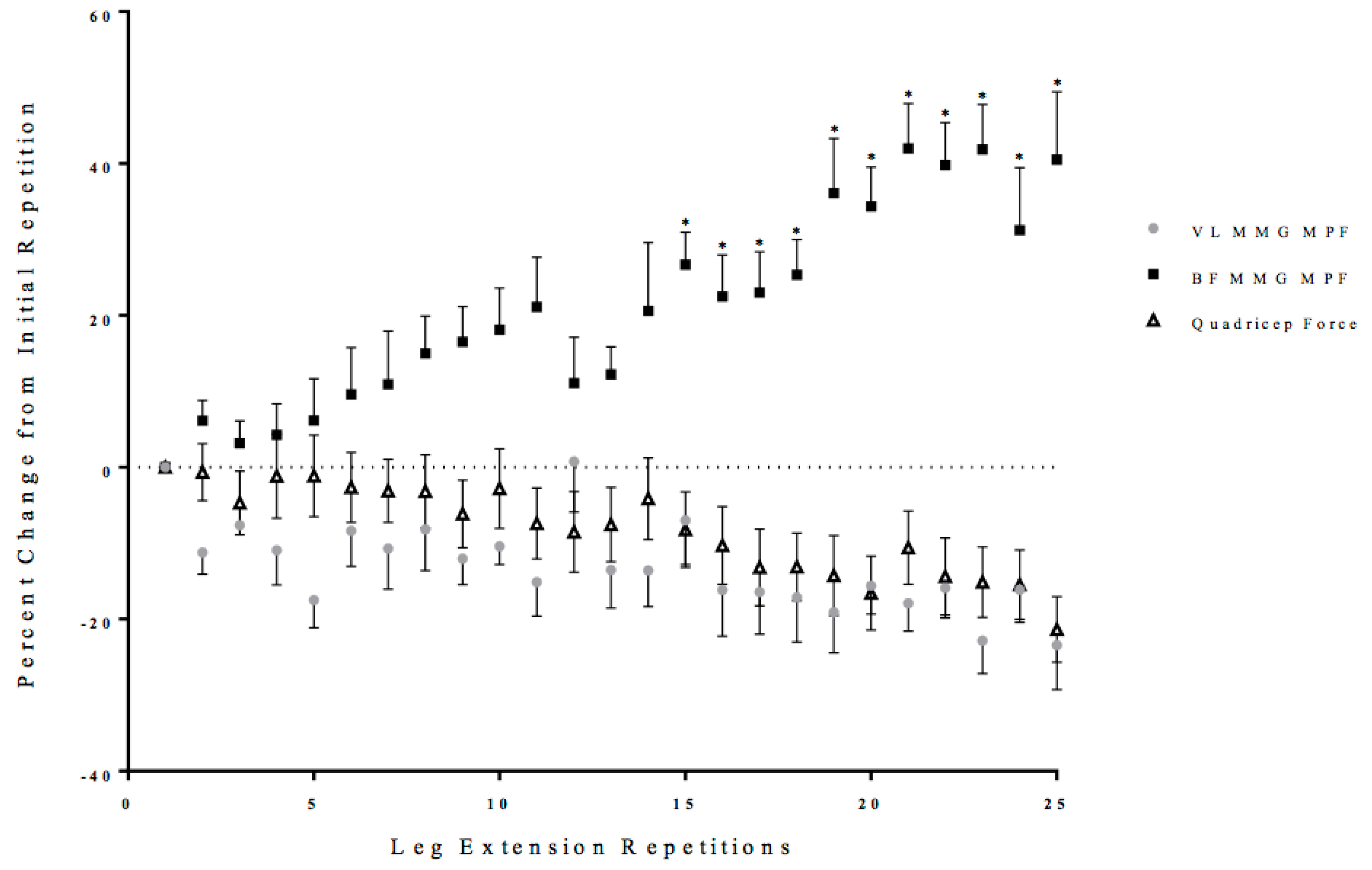
3.2. Neuromuscular Responses
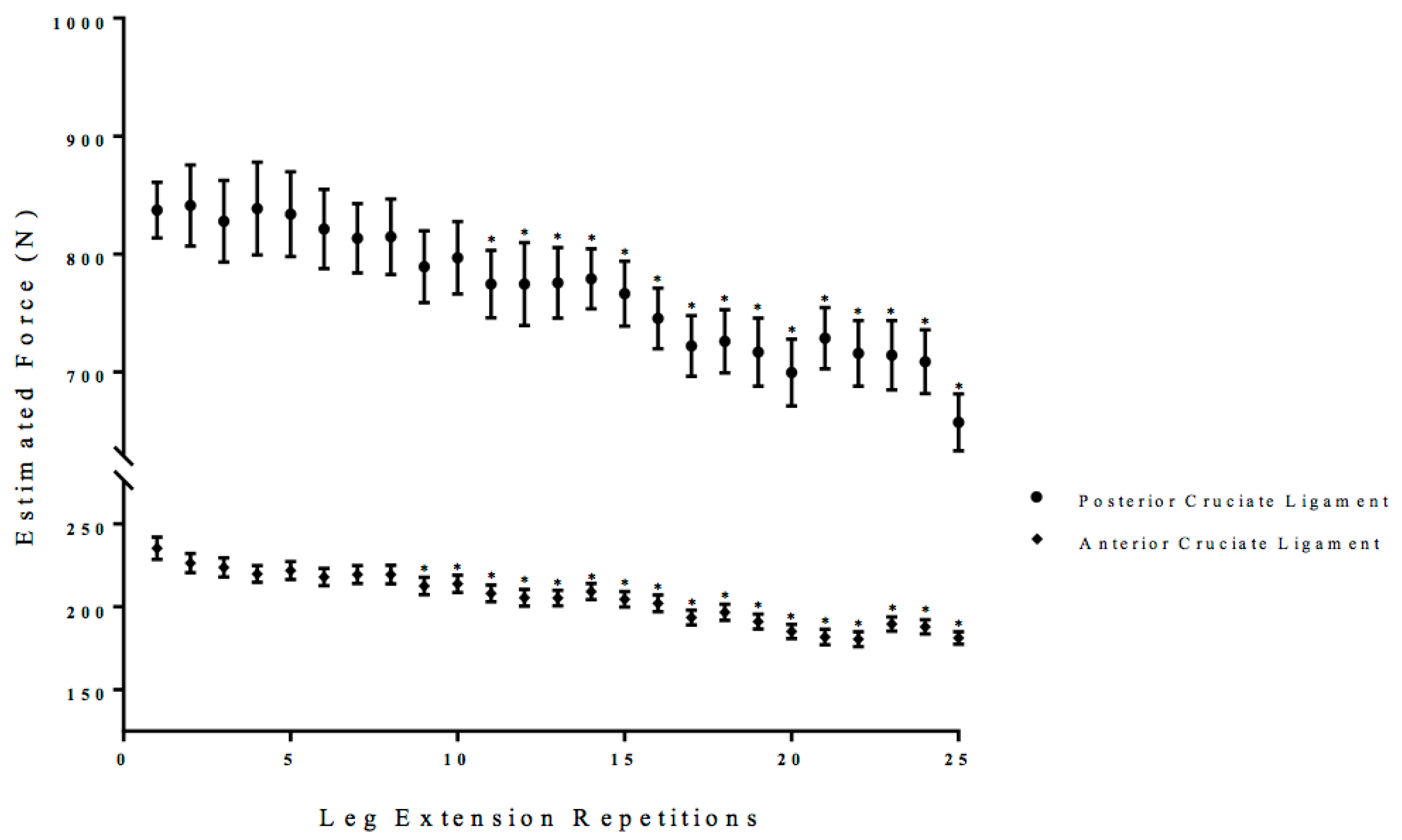
3.3. Estimated Anterior and Posterior Cruciate Ligament Force
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Arendt, E.; Agel, J.; Dick, R. Anterior cruciate ligament injury patterns among collegiate men and women. J. Athl. Train. 1999, 34, 86. [Google Scholar] [PubMed]
- Hughston, J. Posterior Cruciate Ligament in Knee-Joint Stability. J. Bone Joint Surg. 1969, 51, 1045–1046. [Google Scholar]
- Fleming, R.; Blatz, D.; McCarroll, J. Posterior problems in the knee: Posterior cruciate insufficiency and posterolateral rotatory insufficiency. Am. J. Sport Med. 1981, 9, 107–113. [Google Scholar] [CrossRef] [PubMed]
- Zheng, N.; Fleisig, G.; Escamilla, R.; Barrentine, S.W. An analytical model of the knee for estimation of internal forces during exercise. J. Biomech. 1998, 31, 963–967. [Google Scholar] [CrossRef] [Green Version]
- Delp, D. Surgery Simulation: A Computer Graphics System to Analyze and Design Musculoskeletal Reconstructions of the Lower Limb; Stanford University Library: Stanford, CA, USA, 1991. [Google Scholar]
- Ingersoll, C.; Grindstaff, T.; Pietrosimone, B.; Hart, J.M. Neuromuscular consequences of anterior cruciate ligament injury. Clin. Sport Med. 2008, 27, 383–404. [Google Scholar] [CrossRef] [PubMed]
- Brown, L. Isokin Human Performance; Human Kinetics: Champaign, IL, USA, 2000. [Google Scholar]
- Martini, F. Human Anatomy; Pearson/Benjamin Cummings: San Francisco, CA, USA, 2005. [Google Scholar]
- Weir, J.; Keefe, D.; Eaton, J.; Augustine, R.T.; Tobin, D.M. Effect of fatigue on hamstring coactivation during isokinetic knee extensions. Eur. J. Appl. Physiol. 1998, 78, 555–559. [Google Scholar] [CrossRef] [PubMed]
- Draganich, L.; Jaeger, R.; Kralj, A. Coactivation of the hamstrings and quadriceps during extension of the knee. J. Bone Joint Stable 1989, 71, 1075–1081. [Google Scholar] [CrossRef]
- Baratta, R.; Solomonow, M.; Zhou, B.; Letson, D.; Chuinard, R.; D’ambrosia, R. Muscular coactivation: The role of the antagonist musculature in maintaining knee stability. Am. J. Sport Med. 1988, 16, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, J.; De Luca, C. Myoelectric signal versus force relationship in different human muscles. J. Appl. Physiol. 1983, 54, 1653–1659. [Google Scholar] [CrossRef] [PubMed]
- Smith, C.; Housh, T.; Hill, E.; Schmidt, R.J.; Johnson, G.O. Time Course of Changes in Neuromuscular Responses at 30% versus 70% 1 Repetition Maximum during Dynamic Constant External Resistance Leg Extensions to Failure. Intern. J. Exerc. Sci. 2017, 10, 365. [Google Scholar]
- Smith, C. Time Course of Changes in Neuromuscular Parameters during Fatiguing High-Load and Low-Load Concentric Dynamic Constant External Resistance Leg Extension Muscle Actions. Master’s Thesis, University of Nebraska-Lincoln, Lincoln, NE, USA, 12 April 2016. [Google Scholar]
- Farina, D.; Merletti, R.; Enoka, R. The extraction of neural strategies from the surface EMG: An update. J. Appl. Physiol. 2014, 117, 1215–1230. [Google Scholar] [CrossRef] [PubMed]
- De Luca, C.; Contessa, P. Biomechanical benefits of the onion-skin motor unit control scheme. J. Biomech. 2015, 48, 195–203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basmajian, J.; De Luca, C. Muscles Alive: Their Functions Revealed by Electromyography; Williams & Wilkins: Baltimore, MD, USA, 1985. [Google Scholar]
- Orizio, C.; Gobbo, M.; Diemont, B.; Esposito, F.; Veicsteinas, A. The surface mechanomyogram as a tool to describe the influence of fatigue on biceps brachii motor unit activation strategy. Historical basis and novel evidence. Eur. J. Appl. Physiol. 2003, 90, 326–336. [Google Scholar] [CrossRef] [PubMed]
- Orizio, C. Muscle sound: Bases for the introduction of a mechanomyographic signal in muscle studies. Crit. Rev. Biomed. Eng. 1993, 21, 201–243. [Google Scholar] [PubMed]
- Aagaard, P.; Simonsen, E.; Andersen, J.; Magnusson, S.P.; Bojsen-Møller, F.; Dyhre-Poulsen, P. Antagonist muscle coactivation during isokinetic knee extension. Scand. J. Med. Sci. Sports 2000, 10, 58–67. [Google Scholar] [CrossRef] [PubMed]
- Rieb, D. ACSM’s Guidelines for Exercise Testing and Prescription; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2013. [Google Scholar]
- World Medical Association. World Medical Association Declaration of Helsinki: Ethical principles for medical research involving human subjects. JAMA 2013, 310, 2191–2194. [Google Scholar] [CrossRef] [PubMed]
- Abe, T.; Kumagai, K.; Brechue, W. Fascicle length of leg muscles is greater in sprinters than distance runners. Med. Sci. Sport Exerc. 2000, 32, 1125–1129. [Google Scholar] [CrossRef] [Green Version]
- Hermens, H.; Freriks, B.; Merletti, R.; Stegeman, D.; Blok, J.; Rau, G.; Disselhorst-Klug, C.; Hägg, G. European recommendations for surface electromyography. Roessingh Res. Dev. 1999, 8, 13–54. [Google Scholar]
- Barbero, M.; Merletti, R.; Rainoldi, A. Atlas of Muscle Innervation Zones: Understanding Surface Electromyography and Its Applications; Springer Science & Business Media: Berlin, Germany, 2012. [Google Scholar]
- Herzog, W.; Read, L. Lines of action and moment arms of the major force-carrying structures crossing the human knee joint. J. Anat. 1993, 182 Pt 2, 213. [Google Scholar]
- Shelburne, K.; Pandy, M. A musculoskeletal model of the knee for evaluating ligament forces during isometric contractions. J. Biomech. 1997, 30, 163–176. [Google Scholar] [CrossRef]
- Wickiewicz, T.; Roy, R.; Powell, P.; Edgerton, V.R. Muscle architecture of the human lower limb. Clin. Orthop. Relat. Res. 1983, 179, 275–283. [Google Scholar] [CrossRef]
- Smith, C.; Housh, T.; Jenkins, N.; Hill, E.C.; Cochrane, K.C.; Miramonti, A.A.; Schmidt, R.J.; Johnson, G.O. Combining regression and mean comparisons to identify the time course of changes in neuromuscular responses during the process of fatigue. Physiol. Meas. 2016, 37, 1993. [Google Scholar] [CrossRef] [PubMed]
- Salkind, N. Encyclopedia of Research Design; Sage: London, UK, 2010. [Google Scholar]
- Contessa, P.; DeLuca, C.; Kline, J. The compensatory interaction between motor unit firing behavior and muscle force during fatigue. J. Neurophysiol. 2016. [Google Scholar] [CrossRef] [PubMed]
- Arabadzhiev, T.; Dimitrov, V.; Dimitrova, N.; Dimitrov, G.V. Influence of motor unit synchronization on amplitude characteristics of surface and intramuscularly recorded EMG signals. Eur. J. Appl. Physiol. 2009, 108, 227–237. [Google Scholar] [CrossRef] [PubMed]
- Farina, D.; Merletti, R.; Enoka, R. The extraction of neural strategies from the surface EMG. J. Appl. Physiol. 2004, 96, 1486–1495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yao, W.; Fuglevand, R.; Enoka, R. Motor-unit synchronization increases EMG amplitude and decreases force steadiness of simulated contractions. J. Neurophysiol. 2000, 83, 441–452. [Google Scholar] [CrossRef] [PubMed]
- Bouissou, P.; Estrade, P.; Goubel, F.; Guezennec, C.Y.; Serrurier, B. Surface EMG power spectrum and intramuscular pH in human vastus lateralis muscle during dynamic exercise. J. Appl. Physiol. 1989, 67, 1245–1249. [Google Scholar] [CrossRef] [PubMed]
- Barclay, C.; Woledge, R.; Curtin, N. Energy turnover for Ca2+ cycling in skeletal muscle. J. Muscle Res. Cell Motil. 2007, 28, 259–274. [Google Scholar] [CrossRef] [PubMed]
- Bean, J.; Chaffin, D.; Schultz, A. Biomechanical model calculation of muscle contraction forces: A double linear programming method. J. Biomech. 1988, 21, 59–66. [Google Scholar] [CrossRef]
- Hewett, T.; Myer, G.; Ford, K. Anterior cruciate ligament injuries in female athletes. Am. J. Sport Med. 2006, 34, 299–311. [Google Scholar] [CrossRef] [PubMed]







© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Smith, C.M.; Housh, T.J.; Hill, E.C.; Keller, J.L.; Johnson, G.O.; Schmidt, R.J. Co-Activation, Estimated Anterior and Posterior Cruciate Ligament Forces, and Motor Unit Activation Strategies during the Time Course of Fatigue. Sports 2018, 6, 104. https://doi.org/10.3390/sports6040104
Smith CM, Housh TJ, Hill EC, Keller JL, Johnson GO, Schmidt RJ. Co-Activation, Estimated Anterior and Posterior Cruciate Ligament Forces, and Motor Unit Activation Strategies during the Time Course of Fatigue. Sports. 2018; 6(4):104. https://doi.org/10.3390/sports6040104
Chicago/Turabian StyleSmith, Cory M., Terry J. Housh, Ethan C. Hill, Joshua L. Keller, Glen O. Johnson, and Richard J. Schmidt. 2018. "Co-Activation, Estimated Anterior and Posterior Cruciate Ligament Forces, and Motor Unit Activation Strategies during the Time Course of Fatigue" Sports 6, no. 4: 104. https://doi.org/10.3390/sports6040104