1. Introduction
Heat stress (HS) is the most critical physical stressful factor for the poultry industry. HS is caused by high ambient temperature, nutritional stress (nutritional imbalance), and high stocking density (HSD) [
1]. HSD plays a major role in stimulating HS in poultry. Abudabos et al. [
2] showed that the body temperature of chickens is raised at HSD and was higher than that in low stocking density (LSD). Studies on the physiological responses of poultry to stocking density suggested that hens reared at HSD had lower egg performance, biochemical parameters, and immune activities [
3,
4,
5,
6]. Nutritional factors play an important role in alleviating the stress from HSD in laying hens.
A high moisture content of rice grains leads to their germination. Germinated paddy rice (GPR) has enhanced nutritional qualities. Hydrolytic enzymes and other biological components are activated during germination, while starch, polysaccharides, and amino acids are decomposed. The decomposition of polymers in germinated cereal leads to the generation of functional materials [
7]. GPR has many nutritional components, including carbohydrates, protein, oil, dietary fiber, vitamin, γ-oryzanol, and γ-amino butyric acid (GABA) [
7]. GABA acts as a primary inhibitory neurotransmitter in the central nervous system of animals [
8]. Several reports have shown that GABA supplementation from many sources can improve the egg performance, immune response, and alleviate stress of laying hens. Zhang et al. [
9] reported that supplementation of GABA as active ingredient form at 50 mg/kg diet improved the laying performance, egg production, and eggshell breaking strength in laying hens. Egg production, average egg weight, average daily feed intake, feed conversion ratio, percentage of speckled egg, soft shell egg, misshaped egg, and eggshell thickness were significantly improved by the increasing supplementation of a dietary GABA-producing Lactobacillus strain [
10]. In addition, Jeong et al. [
4] noted that dietary GABA as active ingredient form was effective in alleviating stress responses, but the effect was independent of stocking density in broilers. In our previous report, GPR contained 7.48% crude protein, 1.96% ether extract, 67.58% nitrogen free extract, and 12.55% crude fiber as dry matter basis. The gross energy of GPR was 3.77 kcal/g, which was found to be close to the corn. In addition, GPR exhibited a high content of phytonutrients, especially GABA, and this enhanced its antioxidant activities. Therefore, GPR could be a good alternative source of feed ingredients for poultry [
11].
The emergence of novel proteomic techniques in recent years has greatly aided in the understanding of cell functions, physiological and biochemical pathways, and of biological mechanisms [
12,
13]. Proteomics can identify the gene that encodes the protein by combining the amino acid sequence with the genomic database and speculate on the function of the unknown protein. Biomarkers of stress in blood cells which are key components of the immune response, the coagulation cascade, and other functions such as iron and oxygen transportation and protection of blood vessels can be identified [
14]. High-abundance proteins in serum include albumin, immunoglobulins, and transferrin [
15]. Studies on stress proteins, encoded from white blood cells (WBC), have been related to many proteins such as inflammatory protein (NF-κB and TNF-α), interferon (IFN) [
16] and heat shock protein (HSP) [
17]. They also related to the reduction in antioxidant enzymes such as superoxide 4 dismutase (SOD-4), glutathione peroxidase (GPx), and malondialdehyde (MDA) [
18]. It seems that WBC is a good indicator of stress. In addition, the highly complex protein mixtures can be analyzed for proteomic purposes by liquid chromatography coupled with tandem mass spectrometry (LC−MS/MS). Therefore, this study aimed to investigate the effect of dietary germinated paddy rice on egg performance, serum biochemical properties, and proteomic and transcriptomic response of laying hens reared in different stocking densities under chronic heat stress conditions.
4. Discussion
Studies on the physiological responses of poultry to stocking density suggested that certain physiological parameters, biochemical parameters, and expression genes such as GLU, CHO, TG, CORT, Ca, P), LYZ, antioxidant activities, HSP, HMGCR, FASN, and FABP4 could be used to evaluate the chickens under different stocking densities [
4,
5,
9,
10,
32]. HSD affected growth performance, egg production, eggshell quality, health, and immunity [
33]. HSD elevated CORT and heat shock protein 70 (HSP70) expression when compared to broilers raised at LSD [
6].
In this study, increasing stocking densities reduced some parameters such as average daily feed intake, hen-day egg production, egg mass, shell breaking strength, and GABA content in eggs.
Several studies have illustrated the negative effect on body weight gain and feed intake of stocking densities. The review by Wang et al. [
34] indicated that higher stocking densities tended to be associated with reduced feed intake and reduced weight gain in broilers. Jeong et al. [
4] evaluated chickens raised on HSD, reporting that they exhibited a decrease in body weight gain in all phases, feed intake, and an increase in FCR compared with LSD-raised chickens. HSD might contribute to reduced broiler performance because of high temperatures and reduced airflow around the birds. Cage area might affect poultry production, which also affects locomotor activity [
34]. In this study, it was observed that dietary GPR levels significantly improve shell breaking strength and GABA content in eggs. GPR consists of rice husk, which has a high ash content (14–25%). The silica content of the rice husk ash can be as high as 90–98%. Some evidence suggests the involvement of silicon in the formation and stabilization of the extracellular matrix as well as the Ca metabolism. In previous reports, rice hull silicon plays a major role in improving bone-breaking strength and reducing drip and thawing loss of broiler chicken breast and thigh muscles [
35]. Moreover, whole rice hull can be used as a source of insoluble fiber in diets to enhance growth and uniformity of pullet chicks and to improve egg production of laying hens without any harmful impact on egg quality [
36]. In addition, GPR contains phytonutrients, especially GABA, which enhance the antioxidant activities.
Similar findings have been documented by previous researchers. Chand et al. [
37] reported that GABA supplement diets improved the feed intake, body weight, antioxidant status, and immune activity in broilers. El-Naggar et al. [
38] investigated GABA supplementation-increased feed intake by reducing the mRNA expression levels of feed intake-inhibiting neuropeptides, such as proopiomelanocortin (POMC), leptin, chrelin, and cholescystokinin (CCK) by increasing the expression of feed intake-stimulating neuropeptides such as agouti-related protein (AgRP) and neuropeptide (NPY). Zhu et al. [
10] reported that supplementation with 50 mg GABA/kg diet improved average daily feed intake, feed conversion ratio, average egg weight, percentage of speckled egg, soft shell egg, and misshapen egg by enhancing the calcium and phosphorus available for eggshell formation. Zhang et al. [
9] reported that GABA diets in hens could increase amylase, lipase, and trypsin activities in the gastrointestinal tract, thereby increasing egg production, average egg weight, and average daily feed intake. Additionally, Chen et al. [
39] reported that GABA could increase the activity of Ca
2+-Mg
2+-adenosine and Na
+-K
+-adenosine, benefiting the transportation of mineral element. Consequently, GABA improved egg production and egg quality by increasing the total protein concentration and modulating the electrolyte balance [
9].
Serum biochemical parameters are the key indicators for the health of poultry. In this study, diet GPR levels trended to improve the levels of serum CHO, TG and Ca and significantly improved the concentration of GLU, P and CORT hormone (
Table 4). Similar findings have been reported elsewhere. For example, Houshmand et al. [
40] identified increases in GLU and CHO in HSD-raised chickens. Zhigang et al. [
41] reported that levels of serum CHO and TG were increased in cherry valley duck supplemented with 100 mg GABA/kg diet. Curiously, GABA supplementation in chickens reduced lipid deposition and decreased abdominal fat contents, despite the growth in serum triglyceride. This effect may have been due to increased serum levels of GABA, which can stimulate the turnover of fat and the release of free fat acids and glucose into serum to be available to all cells as energy sources [
32].
In contrast, Jeong et al. [
4] noted that blood glucose was decreased in HSD-raised chickens. In addition, CORT is a commonly stress hormone which measured in chickens. In general, the current results confirmed that finding. The levels of serum CORT were higher in chickens raised at HSD than those raised at LSD [
6]. Increasing dietary GPR levels significantly decreased corticosterone hormone. Freeman and Crapo [
42] reported that stress could stimulate the release of CORT, leading to overproduction of oxygen free radicals OH and O
2. However, the body is protected by antioxidant defense systems working in associated with intracellular enzymes such as SOD. SOD in the mitochondria of cells functions to remove superoxide anions by transforming them into H
2O
2 and O
2 [
43]. It should be noted that the levels of total Ig, LZY, and SOD activities were statistically higher in chickens raised at HSD than those in LSD. Dietary at 74 g/kg GPR improved the level of serum ACH50, total Ig, LZY, and SOD activities (
Table 5). Our results are in agreement with Zhang et al. [
9] who reported that GABA significantly improved immune and antioxidant activity by increasing ACH50, total Ig, LZY, and SOD activities. The synthesis of antibodies was improved by adding 50 mg GABA/kg feed, which protected immune cells from the over-production of reactive oxygen species (ROS) caused by heat stress due to the activation of the antioxidant defense system [
10].
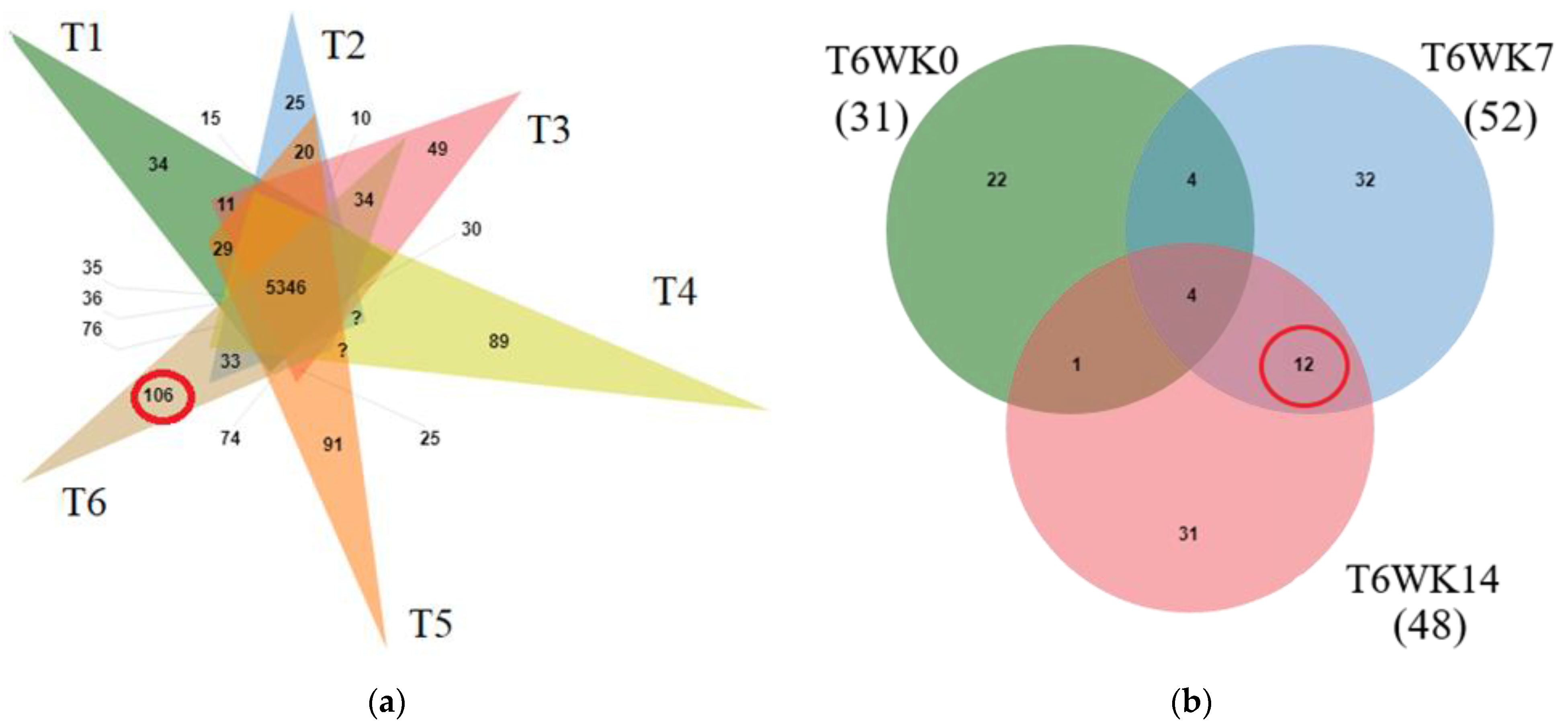
An abundance of differential protein in all treatments at different times was demonstrated (
Table 6). Twelve proteins were present only in T6 during WK7 and WK14 (chickens raised in HSD and dietary at 148 GPR at the middle and late induction times). One protein (TERT; Q3YAE7) was present only in T4WK14 (chickens raised in HSD and dietary 0 g/kg GPR at 14 weeks). One protein (LRP2; G0W2S9) was present only in T6WK14 (chickens raised in HSD and dietary at 148 g/kg GPR at 14 weeks). In addition, we identified four characterized proteins including HSP90, HSP70, HMGCR, FASN, and FABP4 (
Table 6). Telomerase is a ribonucleoprotein enzyme that adds telomeric repeats on telomeres, which are nucleoprotein structures at the ends of eukaryotic chromosomes consisting of tandem repeats of TTAGGG sequences. Telomeres are highly sensitive to damage induced by oxidative stress [
44]. Therefore, telomeric DNA length and telomerase activity are good candidate genes for physiological stress indicators. Sohn et al. [
45] reported that telomere-shortening rate increased in chickens raised under HSD as compared with LSD. HSD had a negative effect on shortening telomere length [
5]. The heat shock proteins (HSP) are a group of conserved proteins that are expressed under thermal and non-thermal stressors. HSP might be stimulated via physiological stresses, pathological stresses, and environmental stresses (such as heat or cold stress, oxidative stress, stocking density stress). HSPs are involved in protein synthesis, and can be classified into several subgroups of molecular chaperones according to their molecular weights (small HSPs, HSP60s, HSP70, HSP90s, and HSP100s). HSPs are considered the protective agents against stress factors [
46]. HMGCR is the rate-controlling enzyme in cholesterol biosynthesis, which in turn is a precursor of cortisol, a universal stress marker. Therefore, increasing cortisol levels in high stocking density can cause negative impact with the synthesis of HMGCR gene under stress situations [
5]. FASN is a key enzyme in fatty acid synthesis which catalyzes the synthesis of long-chain fatty acids. The conversion of acetyl-CoA into malonyl-CoA and then into palmitate is mediated by FASN and ends in triglyceride formation [
47] FABP4 also plays an important role in lipid accumulation during adipogenesis [
48]. It is considered to be important during adipose development because of its role in sequestering fatty acids for triacylglycerol synthesis and as a signaling molecule that regulates activity of enzymes in the adipocyte. Curiously, dietary GPR level supplementation dramatically affected the expression levels of the fat metabolism-related genes
FAS and FABP4, and this impact was linked to reduced fat accumulation in the abdomen of the laying hens. This effect can stimulate the turnover of fat and release of free fat acid and glucose in serum to be available to all cells as energy sources.
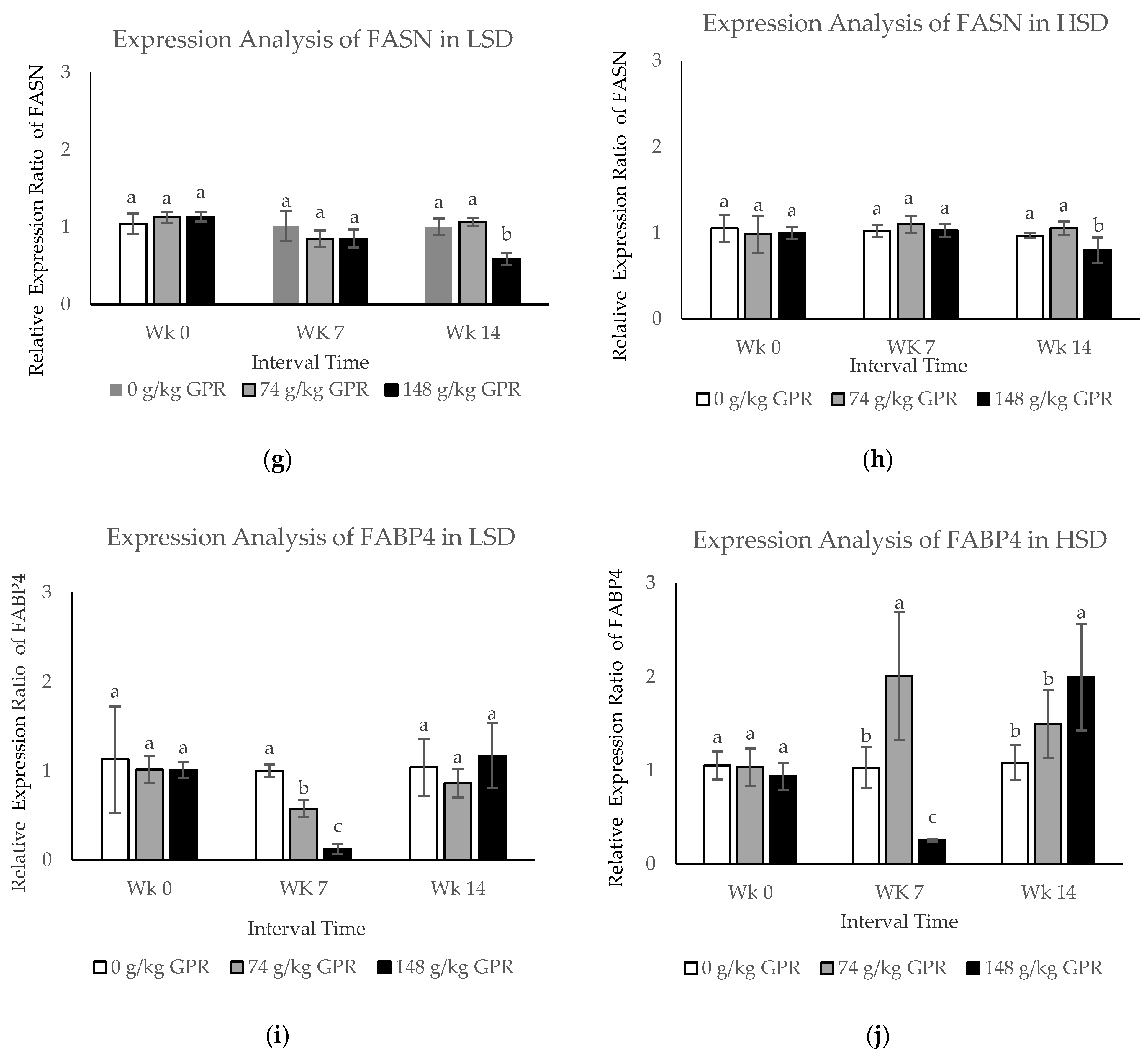
To confirm the mRNA transcription level of identified proteins, the candidate genes were verified by using qPCR. The transcription levels of five genes (HSP70, HSP90, HMGCR, FASN, and FABP4) were determined (
Figure 2). Beloor et al. [
5] reported that the expression levels of HSP70 and HMGCR in blood were increased in HSD (0.0578 m
2/birds) but were not significantly different compared with the LSD (0.116 m
2/birds) and standard (0.077 m
2/birds) groups. However, the stocking density did not affect HSP90. GABA significantly altered FASN gene expression, resulting in significant increase in abdominal fat content in chickens raised normally [
38]. However, the mRNA and protein abundances did not always correlate well. It was often noted that the expression was conducted at the mRNA level, but not at the protein level, because of different regulation operating at transcription and translational steps [
49,
50].
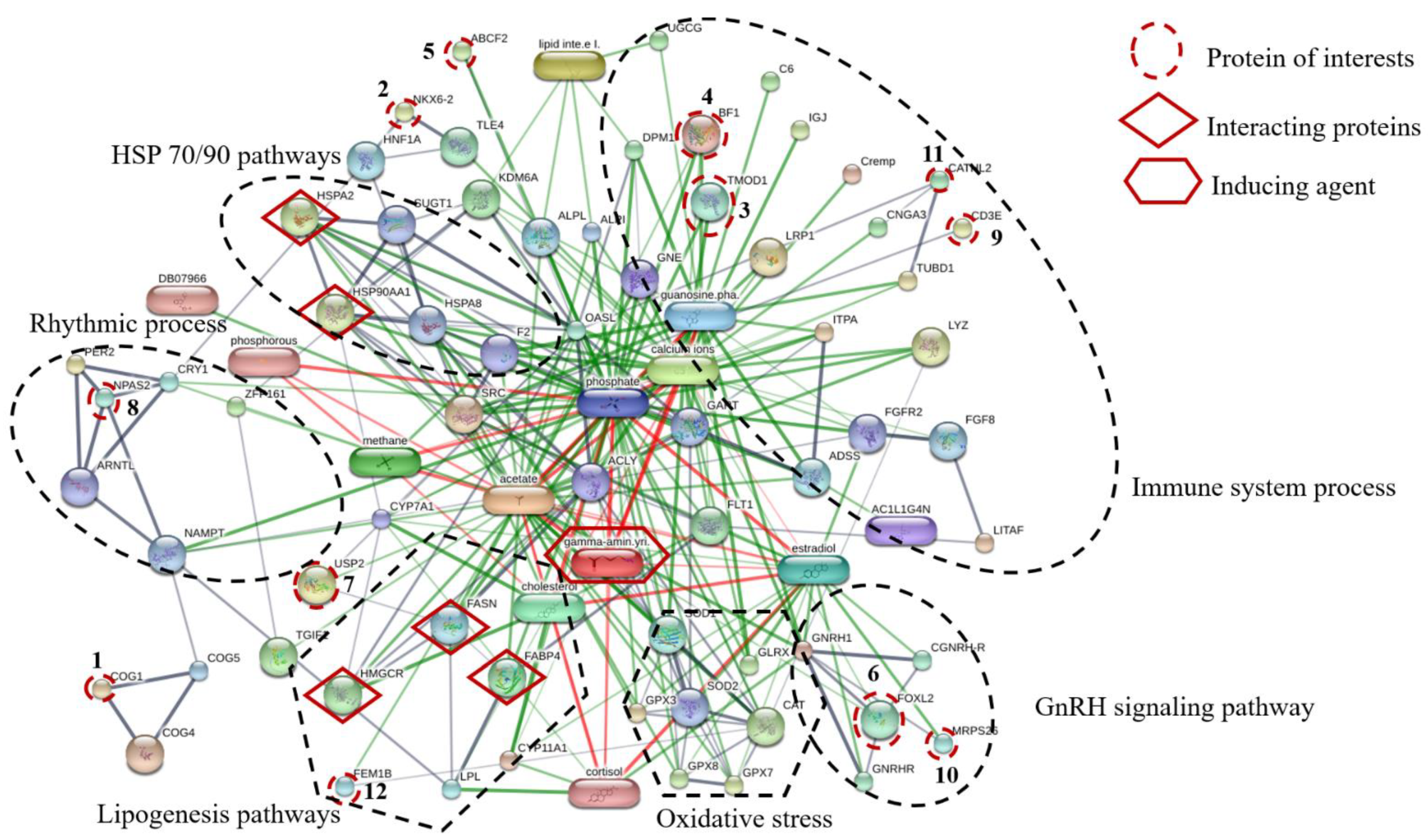
Proteins, protein interactions, and small molecules induced by GABA play major roles in the understanding of molecular and cellular functions. The association between 12 proteins of interest and five interacting proteins (HSP90, HSP70, HMGCR, FASN, and FABP4) were predicted by STITCH version 5.0 using the parameters followed as the organism of
Gallus gallus, medium confidence score (0.4) and active prediction methods (no more than 10 interactions). The results showed that these proteins are associated with proteins involved in HSP70/90 pathways, immune system processes, GnRH signaling pathway, oxidative stress, lipogenesis pathways, and rhythmic process (
Figure 3). The conserved oligomeric Golgi complex subunit 1 (COG1) was associated with COG4, COG5, nicotinamide phosphoribosyltransferase (NAMPT) and neuronal PAS domain-containing protein 2 (NPAS2) which were important in the rhythmic process. The homeobox protein Nkx-6.2 (NKX6.2) was linked to transducin-like enhancer protein 4 (TLE4), hepatocyte nuclear factor 1-alpha (HNF1A) and heat shock 70 kDa protein (HSPA2). Moreover, ATP-binding cassette sub-family F member 2 (ABCF2) was linked to tissue-nonspecific isozyme precursor (ALPL), uncharacterized protein (KDM6A), 59 kDa 2′-5′-oligoadenylate synthase-like protein (OASL), suppressor of G2 allele of SKP1 homolog (SUGT1), heat shock 70 kDa protein (HSPA2), heat shock cognate 71 kDa protein (HSPA8), heat shock protein 90 kDa alpha (cytosolic) class A member 1 (HSP90AA1) and also associated with transducin-like enhancer protein 4 (TLE4) which is known to be involved in HSP70/90 pathways. The forkhead box protein L2 (FOXL2) interacted with mitochondrial ribosomal protein S26 (MRPS26) which is known to be involved in GnRH signaling pathway. Tropomodulin-1 (TMOD1) interacted with MHC BF2 class I precursor (BF1), guanosine diphosphate, low-density lipoprotein receptor-related protein 1 precursor (LRP1), cathelicidin-2 (CATHL2) and T-cell surface glycoprotein CD3 epsilon chain precursor (CD3E), Guanosine phosphate, Calcium ions and GABA which are relevant in the immune system process. Ubiquitin carboxyl-terminal hydrolase 2 (USP2) was associated with FASN and stimulates the secretion of gonadotropins protein fem-1 homolog B (FEM1B), HMGCR, cholesterol, FABP4, lipoprotein lipase (LPL), cortisol hormone and GABA. The mechanism is related to the lipogenesis pathway. In addition, FEM1B was also associated and interacted with catalase (CAT), superoxide dismutase1 (SOD1) and GABA, which are known to be involved in oxidative stress. In this study, dietary GPR levels might affect growth metabolism, lipogenesis, oxidative stress, and immune defense mechanism of laying hens raised at different stoking densities.








{kind=link}
{kind=link}
{kind=link}
{kind=link}