
The Role of Oxidative Stress and the Importance of miRNAs as Potential Biomarkers in the Development of Age-Related Macular Degeneration

 ,
,
Abstract
:1. Introduction
2. AMD—Risk Factors
3. AMD—Enzymatic Antioxidant Capacity
4. AMD—DNA Damage and Repair
5. AMD—miRNA Expression and Diagnostic Markers
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Xiayan, X.; Jing, W.; Xiaoning, Y.; Yelei, T.; Xiajing, T.; Xingchao, S. Regional differences in the global burden of age-related macular degeneration. BMC Public Health 2020, 20, 410–418. [Google Scholar] [CrossRef]
- Shaw, P.X.; Stiles, T.; Douglas, C.; Ho, D.; Fan, W.; Du, H.; Xiao, X. Oxidative stress, innate immunity, and age-related macular degeneration. AIMS Mol. Sci. 2016, 3, 196–221. [Google Scholar] [CrossRef]
- Fleckenstein, M.; Mitchell, P.; Freund, K.B.; Sadda, S.V.; Holz, F.G.; Brittain, C.; Henry, E.C.; Ferrara, D. The Progression of Geographic Atrophy Secondary to Age-Related Macular degeneration. Ophthalmology 2018, 125, 369–390. [Google Scholar] [CrossRef] [PubMed]
- Heesterbeek, T.J.; Lores-Motta, L.; Hoyng, C.B.; Lechanteur, Y.T.E.; den Hollander, A.I. Risk factors for progression of age-related macular degeneration. Ophthalmic. Physiol. Opt. 2020, 40, 140–170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Treister, A.D.; Nesper, P.L.; Fayed, A.E.; Gill, M.K.; Mirza, G.; Fawzi, A.A. Prevalence of Subclinical CNV and Choriocapillaris Nonperfusion in Fellow Eyes of Unilateral Exudative AMD on OCT Angiography. Transl. Vis. Sci. Technol. 2018, 7, 19–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnston, H.; Dickinson, P.; Ivens, A.; Buck, A.H.; Levine, R.D.; Remacle, F.; Campbell, C.J. Intracellular redox potential is correlated with miRNA expression in MCF7 cells under hypoxic conditions. Proc. Natl. Acad. Sci. USA 2019, 116, 19753–19759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Konovalova, J.; Gerasymchuk, D.; Parkkinen, I.; Chmielarz, P.; Domanskyi, A. Interplay between MicroRNAs and oxidative stress in neurodegenerative diseases. Int. J. Mol. Sci. 2019, 20, 6065. [Google Scholar] [CrossRef] [Green Version]
- Algvere, P.V.; Kvanta, A.; Seregard, S. Drusen maculopathy: A risk factor for visual deterioration. Acta Ophthalmol. 2016, 94, 427–433. [Google Scholar] [CrossRef]
- Woodell, A.; Rohrer, B. A mechanistic review of cigarette smoke and agerelated macular degeneration. Adv. Exp. Med. Biol. 2014, 801, 301–307. [Google Scholar]
- Pennington, K.L.; DeAngelis, M.M. Epidemiology of age-related macular degeneration (AMD): Associations with cardiovascular disease phenotypes and lipid factors. Eye Vision 2016, 3, 34–54. [Google Scholar] [CrossRef] [Green Version]
- Zetterberg, M. Age-related eye disease and gender. Maturitas 2016, 83, 19–26. [Google Scholar] [CrossRef]
- Zhang, Q.Y.; Tie, L.J.; Wu, S.S.; Lv, P.L.; Huang, H.W.; Wang, W.Q.; Wang, H.; Ma, L. Overweight, Obesity, and Risk of Age-Related Macular Degeneration. Investig. Opthalmol. Vis. Sci. 2016, 57, 1276–1283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiaohua, L.; Shikun, H.; Mingwei, Z. An Updated Review of the Epigenetic Mechanism Underlying the Pathogenesis of Age-related Macular Degeneration. Aging Dis. 2020, 11, 1219–1234. [Google Scholar]
- Geerlings, M.J.; de Jong, E.K.; den Hollander, A.I. The complement system in age-related macular degeneration: A review of rare genetic variants and implications for personalized treatment. Mol. Immunol. 2017, 84, 65–76. [Google Scholar] [CrossRef]
- Hong, N.; Shen, Y.; Yu, C.Y.; Wang, S.Q.; Tong, J.P. Association of the polymorphism Y402H in the CFH gene with response to anti-VEGF treatment in age-related macular degeneration: A systematic review and meta-analysis. Acta Ophthalmol. 2016, 94, 334–345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clark, S.J.; Bishop, P.N. Role of factor H and related proteins in regulating complement activation in the macula, and relevance to age-related macular degeneration. J. Clin. Med. 2014, 4, 18–31. [Google Scholar] [CrossRef] [Green Version]
- Qian-Qian, Y.; Yong, Y.; Jing, Z.; Xin, B.; Tian-Hua, X.; Chao, S.; Jia, C. Nonsynonymous single nucleotide polymorphisms in the complement component 3 gene are associated with risk of age-related macular degeneration: A meta-analysis. Gene 2015, 561, 249–255. [Google Scholar] [CrossRef]
- Lim, L.S.; Mitchell, P.; Seddon, J.M.; Holz, F.G.; Wong, T.Y. Age-related macular degeneration. Lancet 2012, 379, 1728–1738. [Google Scholar] [CrossRef]
- Hallak, J.A.; de Sisternes, L.; Osborne, A.; Yaspan, B.; Rubin, D.L.; Leng, T. Imaging, Genetic, and Demographic Factors Associated With Conversion to Neovascular Age-Related Macular Degeneration: Secondary Analysis of a Randomized Clinical Trial. JAMA Ophthalmol. 2019, 137, 738–744. [Google Scholar] [CrossRef] [PubMed]
- Wong, W.L.; Su, X.; Li, X.; Cheung, C.M.G.; Klein, R.; Cheng, C.Y.; Wong, T.Y. Global prevalence of age-related macular degeneration and disease burden projection for 2020 and 2040: A systematic review and meta-analysis. Lancet Glob. Health 2014, 2, e106–e116. [Google Scholar] [CrossRef] [Green Version]
- Colijn, J.M.; Buitendijk, G.H.; Prokofyeva, E.; Alves, D.; Cachulo, M.L.; Khawaja, A.P.; Cougnard-Gregoire, A.; Merle, B.M.; Korb, C.; Erke, M.G.; et al. Prevalence of age-related macular degeneration in Europe: The past and the future. Ophthalmology 2017, 124, 1753–1763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Usui, H.; Nishiwaki, A.; Landiev, L.; Kacza, J.; Eichler, W.; Wako, R.; Kato, A.; Takase, N.; Kuwayama, S.; Ohashi, K.; et al. In vitro drusen model – three-dimensional spheroid culture of retinal pigment epithelial cells. J. Cell Sci. Spec. Issue Reconst. Cell Biol. 2019, 132, jcs215798–jcs215805. [Google Scholar] [CrossRef] [Green Version]
- Upadhyay, M.; Milliner, C.; Bell, B.A.; Bonilha, V.L. Oxidative stress in the retina and retinal pigment epithelium (RPE): Role of aging, and DJ-1. Redox Biol. 2020, 37, 101623–101635. [Google Scholar] [CrossRef] [PubMed]
- Renganathan, K.; Gu, J.; Rayborn, M.E.; Crabb, J.S.; Salomon, R.G.; Collier, R.J.; Kapin, M.A.; Romano, C.; Hollyfield, J.G.; Crabb, J.W.; et al. CEP Biomarkers as Potential Tools for Monitoring Therapeutics. PLoS ONE 2013, 8, e76325. [Google Scholar] [CrossRef] [PubMed]
- Cooke, J.P. New Insights Into Tobacco-Induced Vascular Disease: Clinical Ramifications. Methodist Debakey Cardiovasc. J. 2015, 11, 156–159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hou, L.; Yang, G.; Tang, S.; Alcazar, C.; Joshi, P.; Strassberg, Z.; Kim, M.; Kawamura, M.; Woo, Y.J.; Shrager, J.; et al. Small Molecule Derived From Carboxyethylpyrrole Protein Adducts Promotes Angiogenesis in a Mouse Model of Peripheral Arterial Disease. J. Am. Heart Assoc. 2018, 7, e009234–e009245. [Google Scholar] [CrossRef] [Green Version]
- Leńska-Mieciek, M.; Korczak-Kowalska, G.; Bocian, K.; Fiszer, U. Pentosidine, advanced glycation end product, in acute ischaemic stroke patients with and without atrial rhythm disturbances. Neurol. Neurochir. Pol. 2020, 54, 323–328. [Google Scholar] [CrossRef]
- Joo, Y.S.; Areum, S.; Hyunsoo, K.; Youngsoo, K.; Hyeong, G.Y. Carboxymethyl-lysine-modified Plasma Proteins in Age-related Macular Degeneration. Biotechnol. Bioproc. E 2017, 22, 52–59. [Google Scholar] [CrossRef]
- Handa, J.T.; Cano, M.; Wang, L.; Datta, S.; Liu, T. Lipids, Oxidized Lipids, Oxidation-specific Epitopes, and Age-related Macular Degeneration. Biochim. Biophys. Acta. 2017, 1862, 430–440. [Google Scholar] [CrossRef] [Green Version]
- Je-Hyun, B.; Daehan, L.; Park, K.H.; Chae, J.B.; Jang, H.; Lee, J.; Chung, H. Quantitative proteomic analysis of aqueous humor from patients with drusen and reticular pseudodrusen in age-related macular degeneration. BMC Ophthalmol. 2018, 18, 289–303. [Google Scholar] [CrossRef] [Green Version]
- Mrowicka, M.; Mrowicki, J.; Szaflik, J.P.; Szaflik, M.; Ulinska, M.; Szaflik, J.; Majsterek, I. Analysis of antioxidative factors related to AMD risk development in the polish patients. Acta Ophthalmol. 2017, 95, 530–536. [Google Scholar] [CrossRef]
- Yeye, C.; Hezheng, Z. Role of two common SNPs of superoxide dismutase 2 gene in the development of primary open angle glaucoma. Biomed. Res. 2017, 28, 7503–7508. [Google Scholar]
- Abokyi, S.; To, C.H.; Lam, T.T.; Tse, D.Y. Central Role of Oxidative Stress in Age-Related Macular Degeneration: Evidence from a Review of the Molecular Mechanisms and Animal Models. Hindawi Oxidative Med. Cell. Longev. 2020, 2020, 19. [Google Scholar] [CrossRef] [Green Version]
- Saghar, Y. Association between genetic polymorphism of catalase (CAT) C-262T, Cu/Zn superoxide dismutase (SOD1) A251G and risk of age-related macular degeneration. Mol. Bioch. Diagn. J. 2014, 1, 77–88. [Google Scholar]
- Kucukgergin, C.; Gokpinar, M.; Sanli, O.; Tefik, T.; Oktar, T.; Seckin, S. Association between genetic variants in glutathione peroxidase 1 (GPx1) gene, GPx activity and the risk of prostate cancer. Minerva Urol. Nefrol. 2011, 63, 183–190. [Google Scholar] [PubMed]
- Hansen, R.D.; Krath, B.N.; Frederiksen, K.; Tjønneland, A.; Overvad, K.; Roswall, N.; Loft, S.; Dragsted, L.O.; Vogel, U.; Raaschou-Nielsen, O. GPX1 Pro(198)Leu polymorphism, erythrocyte GPX activity, interaction with alcohol consumption and smoking, and risk of colorectal cancer. Mutat. Res. 2009, 664, 13–19. [Google Scholar] [CrossRef]
- Hu, J.; Zhou, G.W.; Wang, N.; Wang, Y.J. GPX1 Pro198Leu polymorphism and breast cancer risk: A meta-analysis. Breast Cancer Res.Treat. 2010, 124, 425–431. [Google Scholar] [CrossRef]
- Zhang, X.; Ng, W.L.; Wang, P.; Tian, L.L.; Werner, E.; Wang, H.; Doetsch, P.; Wang, Y. MicroRNA-21 modulates the levels of reactive oxygen species by targeting SOD3 and TNFα. Canc. Res. 2012, 72, 4707–4713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Q.; Chen, W.; Bai, L.; Chen, W.; Padilla, M.T.; Lin, A.S.; Shi, S.; Wang, X.; Lin, Y. Receptor-interacting protein 1 increases chemoresistance by maintaining inhibitor of apoptosis protein levels and reducing reactive oxygen species through a microRNA-146a-mediated catalase pathway. J. Biol. Chem. 2014, 289, 5654–5663. [Google Scholar] [CrossRef] [Green Version]
- Haque, R.; Chun, E.; Howell, J.C.; Sengupta, T.; Chen, D.; Kim, H. MicroRNA-30b-Mediated Regulation of Catalase Expression in Human ARPE-19 Cells. PLoS ONE 2012, 7, e42542. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Fei, X.; Lu, Y.; Xu, B.; Ma, Y.; Wan, H. miRNA-214 suppresses oxidative stress in diabetic nephropathy via the ROS/Akt/mTOR signaling pathway and uncoupling protein 2. Exp. Ther. Med. 2019, 7, 3530–3538. [Google Scholar] [CrossRef] [Green Version]
- Sangokoya, C.; Telen, M.; Chi, J. microRNA miR-144 modulates oxidative stress tolerance and associates with anemia severity in sickle cell disease. Blood 2010, 116, 4338–4348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szaflik, J.P.; Janik-Papis, K.; Synowiec, E.; Ksiazek, D.; Zaras, M.; Wozniak, K.; Szaflik, J.; Blasiak, J. DNA damage and repair in age-related macular degeneration. Mutat. Res. 2009, 669, 169–176. [Google Scholar] [CrossRef]
- Wozniak, K.; Szaflik, J.P.; Zaras, M.; Sklodowska, A.; Janik-Papis, K.; Poplawski, T.R.; Blasiak, J.; Szaflik, J. DNA damage/repair and polymorphism of the hOGG1 gene in lymphocytes of AMD patients. J. Biomed. Biotechnol. 2009, 2009, 827562. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.; Hao, W.; Pan, L.; Boldogh, I.; Ba, X. The roles of base excision repair enzyme OGG1 in gene expression. Cell. Mol. Life Sci. 2018, 75, 3741–3750. [Google Scholar] [CrossRef] [Green Version]
- Simonelli, V.; Camerini, S.; Mazzei, F.; van Loon, B.; Allione, A.; D’Errico, M.; Barone, F.; Minoprio, A.; Ricceri, F.; Guarrera, S.; et al. Genotype-phenotype analysis of S326C OGG1 polymorphism: A risk factor for oxidative pathologies. Free Radic. Biol. Med. 2013, 63, 401–409. [Google Scholar] [CrossRef]
- Synowiec, E.; Blasiak, J.; Zaras, M.; Szaflik, J.; Szaflik, J.P. Association between polymorphisms of the DNA base excision repair genes MUTYH and hOGG1 and age-related macular degeneration. Exp. Eye Res. 2012, 98, 58–66. [Google Scholar] [CrossRef]
- Shengqun, J.; Nan, H.; Jing, Z.; Junfang, Z.; Ruifang, G.; Jianyan, H.; Huaijin, G. Polymorphisms of the WRN gene and DNA damage of peripheral lymphocytes in age-related cataract in a Han Chinese population. Age 2013, 35, 2435–2444. [Google Scholar] [CrossRef] [Green Version]
- Szaflik, J.P.; Cuchra, M.; Przybylowska-Sygut, K.; Dziki, L.; Kurowska, A.K.; Gacek, M.; Drzewoski, J.; Szaflik, J.; Majsterek, I. Association of the 399Arg/Gln XRCC1, the 194 Arg/Trp XRCC1, the 326Ser/Cys OGG1, and the 324Gln/His MUTYH gene polymorphisms with clinical parameters and the risk for development of primary open-angle glaucoma. Mutat. Res. 2013, 753, 12–22. [Google Scholar] [CrossRef] [PubMed]
- Boal, A.K.; Genereux, J.C.; Sontz, P.A.; Gralnick, J.A.; Newman, D.K.; Barton, J.K. Redox signaling between DNA repair proteins for efficient lesion detection. Proc. Natl. Acad. Sci. USA 2009, 106, 15237–15242. [Google Scholar] [CrossRef] [Green Version]
- Sun, C.; Chen, H.; Guo, W.; Zhang, K.; Qi, Q.; Gu, X.; Zhu, D.; Wang, Y. A common mutation of the MYH gene is associated with increased DNA oxidation and age-related diseases. Free Radic. Biol. Med. 2010, 48, 430–436. [Google Scholar] [CrossRef]
- Synowiec, E.; Wysokinski, D.; Zaras, M.; Kolodziejska, U.; Stoczynska-Fidelus, E.; Janik, K.; Szaflik, J.; Blasiak, J.; Szaflik, J.P. Association between polymorphism of the DNA repair SMUG1 and UNG genes and age-related macular degeneration. Retina 2014, 34, 38–47. [Google Scholar] [CrossRef]
- Mehdizadeh, F.; Bonyadi, M.; Bonyadi, M.H.; Javadzadeh, A. The Association of SMUG1 Gene Polymorphism with Age-related Macular Degeneration in Northwestern Iran. JAMSAT 2016, 2, 257–261. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Lai, Q.; Zhang, S.; Hu, J. Senile cataract and genetic polymorphisms of APE1, XRCC1 and OGG1. Int. J. Clin. Exp. Pathol. 2015, 8, 16036–16045. [Google Scholar]
- Peng, Y.; Li, Z.; Zhang, S.; Xiong, Y.; Cun, Y.; Qian, C.; Li, M.; Ren, T.; Xia, L.; Cheng, Y.; et al. Association of DNA base excision repair genes (OGG1, APE1 and XRCC1) polymorphisms with outcome to platinum-based chemotherapy in advanced nonsmall-cell lung cancer patients. Int. J. Cancer. 2014, 35, 2687–2696. [Google Scholar] [CrossRef] [PubMed]
- Berger, F.; Vaslin, L.; Belin, L.; Asselain, B.; Forlani, S.; Humbert, S.; Durr, A.; Hall, J. The impact of single-nucleotide polymorphisms (SNPs) in OGG1 and XPC on the age at onset of Huntington disease. Mutat. Res. 2013, 755, 115–119. [Google Scholar] [CrossRef] [PubMed]
- Kowalski, M.; Przybylowska, K.; Rusin, P.; Olszewski, J.; Morawiec-Sztandera, A.; Bielecka-Kowalska, A.; Pietruszewska, W.; Mlynarski, W.; Szemaraj, J.; Majsterek, I. Genetic polymorphisms in DNA base excision repair gene XRCC1 and the risk of squamous cell carcinoma of the head and neck. J. Exp. Clin. Cancer Res. 2009, 28, 37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Z.; Zhang, L.; Zhang, W.; Meng, D.; Zhang, H.; Jiang, Y.; Xu, X.; van Meter, M.; Seluanov, A.; Gorbunova, V.; et al. SIRT6 rescues the age related decline in base excision repair in a PARP1-dependent manner. Cell Cycle 2015, 14, 269–276. [Google Scholar] [CrossRef]
- Wang, A.L.; Lukas, T.J.; Yuan, M.; Neufeld, A.H. Increased mitochondrial DNA damage and down-regulation of DNA repair enzymes in aged rodent retinal pigment epithelium and choroid. Mol. Vis. 2008, 14, 644–651. [Google Scholar]
- Wang, A.L.; Lukas, T.J.; Yuan, M.; Neufeld, A.H. Age-related increase in mitochondrial DNA damage and loss of DNA repair capacity in the neural retina. Neurobiol. Aging 2010, 31, 2002–2010. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, L.; Zhang, L.; Bai, J.; Ge, H.; Liu, P. Expression changes in DNA repair enzymes and mitochondrial DNA damage in aging rat lens. Mol. Vis. 2010, 16, 1754–1763. [Google Scholar]
- Santos, J.C.; Funck, A.; Silva-Fernandes, I.J.; Rabenhorst, S.H.; Martinez, C.A.; Ribeiro, M.L. Effect of APE1 T2197G (Asp148Glu) polymorphism on APE1, XRCC1, PARP1 and OGG1 expression in patients with colorectal cancer. Int. J. Mol. Sci. 2014, 29, 17333–17343. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Taniguchi, T. MicroRNAs and DNA damage response: Implications for cancer therapy. Cell Cycle. 2013, 12, 32–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tinaburri, L.; D’Errico, M.; Sileno, S.; Maurelli, R.; Degan, P.; Magenta, A.; Dellambra, E. miR-200a Modulates the Expression of the DNA Repair Protein OGG1 Playing a Role in Aging of Primary Human Keratinocytes. Oxid. Med. Cell. Longev. 2018, 2018, 9147326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, H.L.; Shi, Y.P.; He, H.J.; Wang, Y.H.; Chen, T.; Yang, L.W.; Yang, T.; Chen, J.; Cao, J.; Yao, W.M.; et al. miR-4673 modulates paclitaxel-induced oxidative stress and loss of mitochondrial membrane potential by targeting 8-oxoguanine-DNA glycosylase-1. Cell. Physiol. Biochem. 2017, 42, 889–900. [Google Scholar] [CrossRef] [Green Version]
- Catalanotto, C.; Cogoni, C.; Zardo, G. MicroRNA in Control of Gene Expression: An Overview of Nuclear Functions. Int. J. Mol. Sci. 2016, 17, 1712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, C.; Liu, Q.; Wei, Q.; Cai, W.; He, M.; Du, Y.; Xu, D.; Wu, Y.; Yu, J. Circulating miRNAs as potential biomarkers of age-related macular degeneration. Cell. Physiol. Biochem. 2017, 41, 1413–1423. [Google Scholar] [CrossRef] [PubMed]
- Ulańczyk, Z.; Sobuś, A.; Łuczkowska, K.; Grabowicz, A.; Mozolewska-Piotrowska, K.; Safranow, K.; Kawa, M.P.; Pałucha, A.; Krawczyk, M.; Sikora, P.; et al. Associations of microRNAs, angiogenesis-regulating factors and CFH Y402H polymorphism–an attempt to search for systemic biomarkers in age-related macular degeneration. Int. J. Mol. Sci. 2019, 20, 5750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szemraj, M.; Oszajca, K.; Szemraj, J.; Jurowski, P. MicroRNA expression analysis in serum of patients with congenital hemochromatosis and age-related macular degeneration (AMD). Med. Sci. Monit. 2017, 23, 4050–4060. [Google Scholar] [CrossRef] [Green Version]
- Litwińska, Z.; Sobuś, A.; Łuczkowska, K.; Grabowicz, A.; Mozolewska-Piotrowska, K.; Safranow, K.; Kawa, M.P.; Machaliński, B.; Machalińska, A. The interplay between systemic inflammatory factors and microRNAs in age-related macular degeneration. Front. Aging Neurosci. 2019, 11, 286–298. [Google Scholar] [CrossRef] [PubMed]
- Ménard, C.; Rezende, F.A.; Miloudi, K.; Wilson, A.; Tétreault, N.; Hardy, P.; SanGiovanni, J.P.; de Guire, V.; Sapieha, P. MicroRNA signatures in vitreous humour and plasma of patients with exudative AMD. Oncotarget 2016, 7, 19171–19184. [Google Scholar] [CrossRef] [Green Version]
- Pogue, A.I.; Lukiw, W.J. Up-regulated pro-inflammatory microRNAs (miRNAs) in Alzheimer’s disease and age-related macular degeneration (AMD). Cell. Mol. Neurobiol. 2018, 38, 1021–1031. [Google Scholar] [CrossRef] [PubMed]
- Weber, J.A.; Baxter, D.H.; Zhang, S.; Huang, D.Y.; Huang, K.H.; Lee, M.J.; Galas, D.J.; Wang, K. The microRNA spectrum in 12 body fluids. Clin. Chem. 2010, 56, 1733–1741. [Google Scholar] [CrossRef] [PubMed]
- Ragusa, M.; Caltabiano, R.; Russo, A.; Puzzo, L.; Avitabile, T.; Longo, A.; Toro, M.D.; di Pietro, C.; Purrello, M.; Reibaldi, M. MicroRNAs in vitreus humor from patients with ocular diseases. Mol. Vis. 2013, 19, 430–440. [Google Scholar] [PubMed]
- Elbay, A.; Ercan, Ç.; Akbaş, F.; Bulut, H.; Ozdemir, H. Three new circulating microRNAs may be associated with wet age-related macular degeneration. Scand. J. Clin. Lab. Investig. 2019, 79, 388–394. [Google Scholar] [CrossRef] [PubMed]
- Valadi, H.; Ekstrom, K.; Bossios, A.; Sjostrand, M.; Lee, J.J.; Lotvall, J.O. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat. Cell Biol. 2007, 9, 654–659. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.; Lee, H.; Wang, X.; Groot, M.; Sharma, L.; dela Cruz, C.S.; Jin, Y. A potential role of microvesicle-containing miR-223/142 in lung inflammation. Thorax 2019, 74, 865–874. [Google Scholar] [CrossRef]
- Panshina, D.D.; Kondratov, K.A. The Efficiency of Immunoprecipitation of microRNA/AGO2 Complexes from Human Blood Plasma is Protocol Dependent. Mol. Biol. 2020, 54, 213–219. [Google Scholar] [CrossRef]
- Axmann, M.; Meier, S.M.; Karner, A.; Strobl, W.; Stangl, H.; Plochberger, B. Serum and Lipoprotein Particle miRNA Profile in Uremia Patients. Genes 2018, 9, 533. [Google Scholar] [CrossRef] [Green Version]
- Datta, S.; Cano, M.; Ebrahimi, K.; Wang, L.; Handa, J.T. The Impact of Oxidative Stress and Inflammation on RPE Degeneration in Non-neovascular AMD. Prog. Retin. Eye Res. 2017, 60, 201–218. [Google Scholar] [CrossRef]
- Wei, T.; Jingling, Z.; Shigeo, Y.; Bing, J.; Yedi, Z. The Role of Inflammation in Age-Related Macular Degeneration. Int. J. Biol. Sci. 2020, 16, 2989–3001. [Google Scholar] [CrossRef]
- Knickelbein, J.E.; Chan, C.C.; Sen, H.N.; Ferris, F.L.; Nussenblatt, R.B. Inflammatory mechanisms of age-related macular degeneration. Int. Ophthalmol. Clin. 2015, 55, 63–78. [Google Scholar] [CrossRef] [Green Version]
- Madelaine, R.; Sloan, S.A.; Huber, N.; Notwell, J.H.; Leung, L.C.; Skariah, G.; Halluin, C.; Pasca, S.P.; Bejerano, G.; Krasnow, M.A.; et al. MicroRNA-9 couples brain neurogenesis and angiogenesis. Cell Rep. 2017, 20, 1533–1542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhattacharjee, S.; Zhao, Y.; Dua, P.; Rogaev, E.I.; Lukiw, W.J. microRNA34a-mediated down-regulation of the microglial-enriched triggering receptor and phagocytosis-sensor TREM2 in age-related macular degeneration. PLoS ONE 2016, 11, e0150211. [Google Scholar] [CrossRef]
- Krzywińska, O.; Bracha, M.; Jeanniere, C.; Recchia, E.; Kędziora Kornatowska, K.; Kozakiewicz, M. Meta-Analysis of the Potential Role of miRNA-21 in Cardiovascular System Function Monitoring. BioMed Res. Int. 2020, 2020, 4525410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ertekin, S.; Yildirim, O.; Dinc, E.; Ayaz, L.; Fidanci, S.B.; Tamer, L. Evaluation of circulating miRNAs in wet age-related macular degeneration. Mol. Vis. 2014, 20, 1057–1066. [Google Scholar]
- Lin, S.T.; Fu, Y.H. miR-23 regulation of lamin B1 is crucial for oligodendrocyte development and myelination. Dis. Model Mech. 2009, 2, 178–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romano, G.L.; Platania, C.B.M.; Drago, F.; Salomone, S.; Ragusa, M.; Barbagallo, C.; di Pietro, C.; Purrello, M.; Reibaldi, M.; Avitabile, T.; et al. Retinal and circulating miRNAs in age-related macular degeneration: An in vivo animal and human study. Front. Pharmacol. 2017, 8, 168–177. [Google Scholar] [CrossRef] [Green Version]
- Harris, T.A.; Yamakuchi, M.; Ferlito, M.; Mendell, J.T.; Lowenstein, C.J. MicroRNA-126 regulates endothelial expression of vascular cell adhesion molecule 1. Proc. Natl. Acad. Sci. USA 2008, 105, 1516–1521. [Google Scholar] [CrossRef] [Green Version]
- Fish, J.E.; Santoro, M.M.; Morton, S.U.; Yu, S.; Yeh, R.F.; Wythe, J.D.; Ivey, K.N.; Bruneau, B.G.; Stainier, D.Y.; Srivastava, D. miR-126 regulates angiogenic signaling and vascular integrity. Dev. Cell 2008, 15, 272–284. [Google Scholar] [CrossRef] [Green Version]
- Blasiak, J.; Watala, C.; Tuuminen, R.; Kivinen, N.; Koskela, A.; Uusitalo-Järvinen, H.; Tuulonen, A.; Winiarczyk, M.; Mackiewicz, J.; Zmorzyński, S.; et al. Expression of VEGFA-regulating miRNAs and mortality in wet AMD. J. Cell Mol. Med. 2019, 23, 8464–8471. [Google Scholar] [CrossRef] [Green Version]
- Jadeja, R.N.; Jones, M.A.; Abdelrahman, A.A.; Powell, F.L.; Thounaojam, M.C.; Gutsaeva, D.; Bartoli, M.; Martin, P.M. Inhibiting microRNA-144 potentiates Nrf2-dependent antioxidant signalingin RPE and protects against oxidative stress-induced outer retinaldegeneration. Redox Biol. 2019, 28, 101336. [Google Scholar] [CrossRef] [PubMed]
- Rusca, N.; Monticelli, S. MiR-146a in Immunity and Disease. Mol. Biol. Int. 2011, 2011, 437301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Q.; Bozack, S.N.; Yan, Y.; Boulton, M.E.; Grant, M.B.; Busik, J.V. Regulation of retinal inflammation by rhythmic expression of MiR-146a in diabetic retina. Investig. Ophthalmol. Vis. Sci. 2014, 55, 3986–3994. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.-H.; Sun, Y.; Li, J.; Gong, Y.; Tian, K.T.; Evans, L.P.; Morss, P.C.; Fredrick, T.W.; Saba, N.J.; Chen, J. Endothelial microRNA-150 is an intrinsic suppressor of pathologic ocular neovascularization. Proc. Natl. Acad. Sci. USA 2015, 112, 12163–12168. [Google Scholar] [CrossRef] [Green Version]
- Lin, J.B.; Moolani, H.V.; Sene, A.; Sidhu, R.; Kell, K.; Lin, J.B.; Dong, Z.; Ban, N.; Ory, D.S.; Apte, R.S. Macrophage microRNA-150 promotes pathological angiogenesis as seen in age-related macular degeneration. JCI Insight 2018, 3, e120157. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.; Lee, S.; Lazzaro, D.R.; Aranda, J.; Grant, M.B.; Chaqour, B. Single and Compound Knock-outs of MicroRNA (miRNA)-155 and Its Angiogenic Gene Target CCN1 in Mice Alter Vascular and Neovascular Growth in the Retina via Resident Microglia. J. Biol. Chem. 2015, 290, 23264–23281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lukiw, W.J.; Surjyadipta, B.; Dua, P.; Alexandrov, P.N. Original Article Common micro RNAs (miRNAs) target complement factor H (CFH) regulation in Alzheimer’s disease (AD) and in age-related macular degeneration (AMD). Int. J. Biochem. Mol. Biol. 2012, 3, 105–116. [Google Scholar]
- Lee, H.; Han, S.; Kwon, C.S.; Lee, D. Biogenesis and regulation of the let-7 miRNAs and their functional implications. Protein Cell 2016, 7, 100–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szemraj, M.; Bielecka-Kowalska, A.; Oszajca, K.; Krajewska, M.; Gos, R.; Jurowski, P.; Kowalski, M.; Szemraj, J. Serum MicroRNAs as Potential Biomarkers of AMD. Med. Sci. Monit. 2015, 21, 2734–2742. [Google Scholar] [CrossRef] [Green Version]
- Turchonovich, A.; Weiz, L.; Langheinz, A.; Burwinkel, B. Characterization of extracellular circulating microRNA. Nucleid Acids Res. 2011, 39, 7223–7233. [Google Scholar] [CrossRef]
- Mattes, J.; Yang, M.; Foster, P.S. Regulation of microRNA by antagomirs: A new class of pharmacological antagonists for the specific regulation of gene function? Am. J. Respir. Cell Mol. Biol. 2007, 36, 8–12. [Google Scholar] [CrossRef]
- Sharma, S.; Khadimallah, I.; Corya, A.W.; Ali, Y.O.; Rao, X.; Liu, Y.; Lu, H.C. Presymptomatic change in microRNAs modulates Tau pathology. Sci. Rep. 2018, 8, 9251. [Google Scholar] [CrossRef] [Green Version]
- Rui, S.; Seungil, R.; Wei, Y. In situ hybridization detection of microRNAs. Methods Mol Biol. 2010, 629, 287–294. [Google Scholar] [CrossRef] [Green Version]
- Jiawei, Y.; Mingcheng, X.; Xueke, T.; Sheng, C.; Su, Z. Research advances in the detection of miRNA. J Pharm. Anal. 2019, 9, 217–226. [Google Scholar]
- Sato, F.; Tsuchiya, S.; Terasawa, K.; Tsujimoto, G. Intra-platform repeatability and inter-platform comparability of microRNA microarray technology. PLoS ONE 2009, 4, e5540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berber, P.; Grassmann, F.; Kiel, C.; Weber, B.H. An Eye on Age-Related Macular Degeneration: The Role of MicroRNAs in Disease Pathology. Mol. Diagn. Ther. 2017, 21, 31–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, C.; Yan, X.; Li, H.; Huang, L.; Yin, M.; Yang, Y.; Gao, R.; Hong, L.; Ma, Y.; Shi, C.; et al. Systematic literature review and clinical validation of circulating microRNAs as diagnostic biomarkers for colorectal cancer. Oncotarget 2017, 8, 68317–68328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paradowska, D.; Kucharska, E. Nutrition assessment of patients with AMD. World Sci. News 2020, 139, 221–232. [Google Scholar]
- Jonasson, F.; Fisher, D.E.; Eiriksdottir, G.; Sigurdsson, S.; Klein, R.; Launer, L.J.; Harris, T.; Gudnason, V.; Cotch, M.F. Five-year incidence, progression and risk factors for age-related macular degeneration: The Age, Gene/Environment Susceptibility Study. Ophthalmology 2014, 121, 1766–1772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pugazhendhi, A.; Hubbell, M.; Jairam, P.; Ambati, B. Neovascular Macular Degeneration: A Review of Etiology, Risk Factors, and Recent Advances in Research and Therapy. Int. J. Mol. Sci. 2021, 22, 1170. [Google Scholar] [CrossRef] [PubMed]
- Domènech, E.B.; Marfany, G. The Relevance of Oxidative Stress in the Pathogenesis and Therapy of Retinal Dystrophies. Antioxidants 2020, 9, 347. [Google Scholar] [CrossRef] [PubMed] [Green Version]


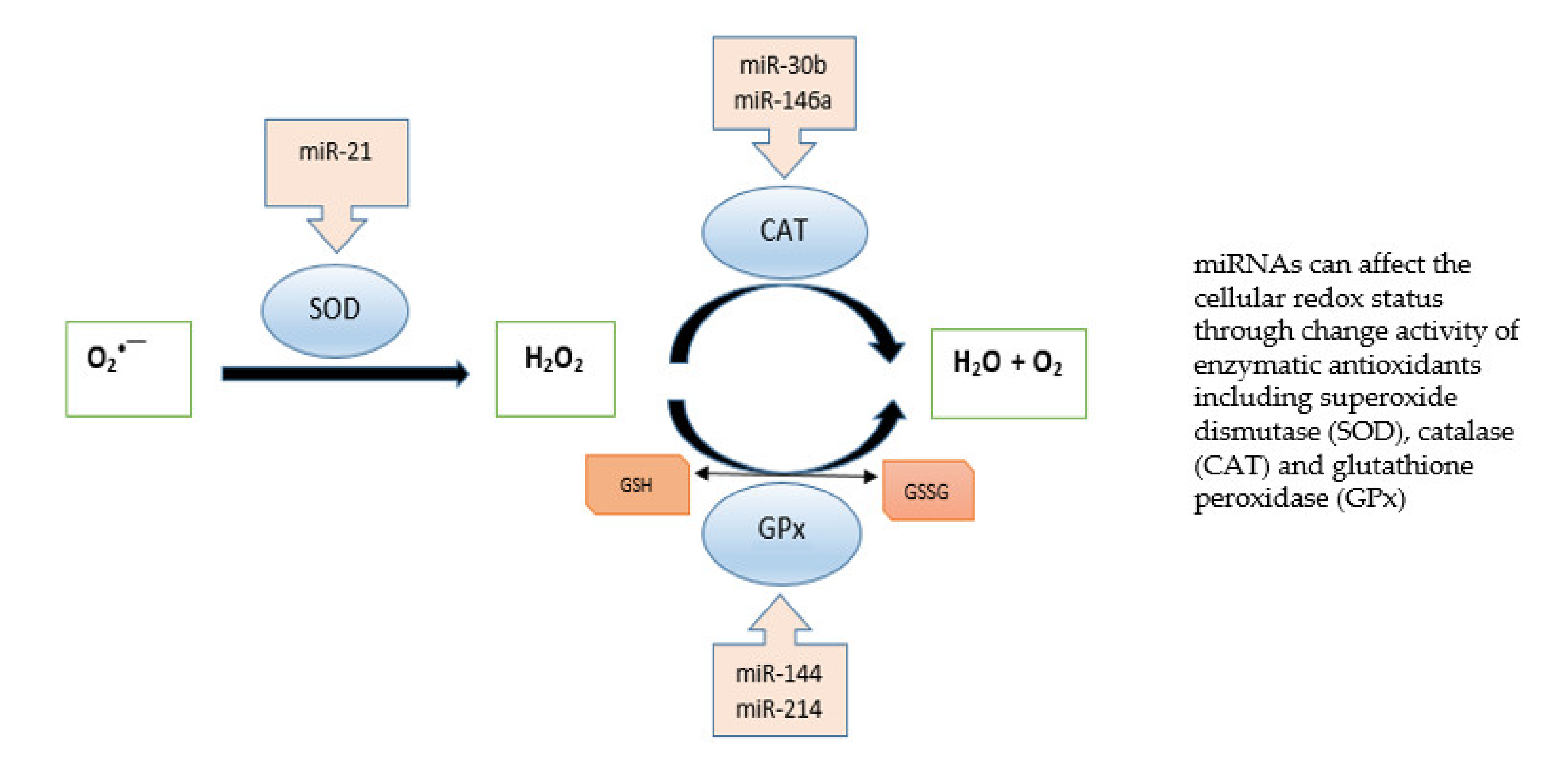{kind=link}
| miRNAs | Sample | Comparison | Effects | Reference |
|---|---|---|---|---|
| miR-9 | Human retinal tissue | Dry AMD vs. HC | Upregulated | Bhattacharjee et al. 2016 |
| miR-21 | Blood plasma | AMD vs. HC | Downregulated | Ulanczyk et al. 2019 |
| miR-23a | PBNCs | Wet AMD vs. HC | Upregulated | Litwińska et al. 2019 |
| Blood serum | Wet AMD vs. HC | Upregulated | Romano et al. 2017 | |
| miR-27a | Blood | Wet vs. dry AMD | Upregulated | Ren et al. 2017 |
| miR-126 | PBNCs | Dry AMD vs. HC | Upregulated | Litwińska et al. 2019 |
| miR-144 | Blood | Wet vs. dry AMD | Upregulated | Ren et al. 2017 |
| miR-146a | PBNCs | Dry AMD vs. HC | Upregulated | Litwińska et al. 2019 |
| Blood plasma | Wet AMD vs. HC | Upregulated | Menard et al. 2016 | |
| Blood serum | Wet AMD vs. HC | Upregulated | Romano et al. 2017 | |
| Retinal tissues | AMD vs. HC | Upregulated | Pogue and Lukiw 2018 | |
| miR-150 | PBNCs | AMD vs. HC | Upregulated | Lin et al. 2018 |
| miR-155 | Human retinal tissue | Dry AMD vs. HC | Upregulated | Bhattacharjee et al. 2016 |
| Let-7 | Blood serum | Wet AMD vs. HC | Upregulated | Szemraj et al. 2015 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mrowicka, M.; Mrowicki, J.; Kucharska, E.; Smigielska, B.; Szaflik, J.P.; Szaflik, J.; Majsterek, I. The Role of Oxidative Stress and the Importance of miRNAs as Potential Biomarkers in the Development of Age-Related Macular Degeneration. Processes 2021, 9, 1328. https://doi.org/10.3390/pr9081328
Mrowicka M, Mrowicki J, Kucharska E, Smigielska B, Szaflik JP, Szaflik J, Majsterek I. The Role of Oxidative Stress and the Importance of miRNAs as Potential Biomarkers in the Development of Age-Related Macular Degeneration. Processes. 2021; 9(8):1328. https://doi.org/10.3390/pr9081328
Chicago/Turabian StyleMrowicka, Malgorzata, Jerzy Mrowicki, Ewa Kucharska, Barbara Smigielska, Jacek Pawel Szaflik, Jerzy Szaflik, and Ireneusz Majsterek. 2021. "The Role of Oxidative Stress and the Importance of miRNAs as Potential Biomarkers in the Development of Age-Related Macular Degeneration" Processes 9, no. 8: 1328. https://doi.org/10.3390/pr9081328