
Improved Foods Using Enzymes from Basidiomycetes


Institute of Food Chemistry, Leibniz University Hannover, Callinstraße 5, 30167 Hannover, Germany
*
Author to whom correspondence should be addressed.
Processes 2022, 10(4), 726; https://doi.org/10.3390/pr10040726
Submission received: 18 March 2022
/
Revised: 4 April 2022
/
Accepted: 6 April 2022
/
Published: 9 April 2022
(This article belongs to the Special Issue Advances in Microbial Fermentation Processes II)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Within the kingdom of fungi, the division Basidiomycota represents more than 30,000 species, some with huge genomes indicating great metabolic potential. The fruiting bodies of many basidiomycetes are appreciated as food (“mushrooms”). Solid-state and submerged cultivation processes have been established for many species. Specifically, xylophilic fungi secrete numerous enzymes but also form smaller metabolites along unique pathways; both groups of compounds may be of interest to the food processing industry. To stimulate further research and not aim at comprehensiveness in the broad field, this review describes some recent progress in fermentation processes and the knowledge of fungal genetics. Processes with potential for food applications based on lipases, esterases, glycosidases, peptidases and oxidoreductases are presented. The formation and degradation of colourants, the degradation of harmful food components, the formation of food ingredients and particularly of volatile and non-volatile flavours serve as examples. In summary, edible basidiomycetes are foods—and catalysts—for food applications and rich donors of genes to construct heterologous cell factories for fermentation processes. Options arise to support the worldwide trend toward greener, more eco-friendly and sustainable processes.
1. Introduction
Basidiomycetes, or taxonomically more correct Basidiomycota, are a diverse group of higher fungi, comprising the subdivisions Pucciniomycetes, Ustilaginomycetes and Agaricomycetes. The latter form a subterranean network of cells in forests, acting as an important sink of carbon. Under suitable conditions, they break through the soil surface, degrading lignocellulosic materials as a source of nutrients while forming fruiting bodies for sexual propagation (“macrofungi”). The thin fungal cells (“hyphae”) can be cultivated in vitro in solid or liquid media and thrive on wood, forest litter and on many side-streams of the agro-industries (peel, bran, etc.). Some basidiomycetous genomes are only slightly shorter than the human genome (3 bil. bp, 20,000 genes) [1], with Phakopsora pachyrhizi (1.3 bil. bp (All bp numbers are given for the haploid genomes.), 22,500 genes) as the current record holder [2,3]. Around 1000 species within the Agaricomycetes are estimated to be edible (“mushrooms”). Since ancient times, they have been appreciated for their pleasant aroma and nutritional value (and many fungi as well for their supposed medicinal applications). Still, food processing is mainly based on enzymes of bacterial, yeast and ascomycetous origins, although a huge and diverse repertoire of basidiomycetous enzymes has been isolated and characterised [4,5]. Their immense potential awaits exploitation in technical applications as a source of processing aids, ingredients and bioactive molecules.
Enzymes secreted by basidiomycetes have to stay active outside protecting cells for extended periods, therefore, they often possess unique stabilities. Many are distinguished by remarkable tolerance toward acidic [6] or alkaline conditions [7] and also show unusual thermostability [8] and resistance against peptidolysis [9,10]. The substrate and reaction specificity typical of enzyme catalysis adds to these features and should contribute to a broader application, not only in the food industries, to promote “green chemistry” processes.
The present review, without aiming at comprehensiveness, presents some more recent food-related examples on the use of basidiomycete cell cultures and enzyme activities (Table S1). First, progress in fermentation processes and genetic advances are reported, followed by the description of processes to change the structure and quality of foods, to use agro-industrial side-streams for added-value creation, to selectively remove unwanted and to increase valuable food constituents and, finally, to show perspectives for future research in the field.
2. Progress in Fermentation Processes
Like the demanding plant cells, basidiomycetes in submerged culture typically grow slowly, a tribute to their genetic complexity. In addition, apart from marine basidiomycetes, these species live in or on solid substrates, where water activity is rather low and oxygen supply rather ample. Thus, large-scale processes for the production of enzymes on the industrial scale, stirred-tank bioreactors or similar reactor types are rarely found, although liquid processes provide fast, up-scalable and compact production. Two alternatives are available: (1) Classic solid-state cultivation and (2) the transfer of structural genes from the basidiomycete donor into a rapidly growing, submerged cultured heterologous host.
Solid-state cultivation of basidiomycetes involves inexpensive, moist substrates, such as wet straw or wood chips and may run under non-controlled, semi-sterile conditions for extended periods. Proven in the production of traditional (Asian) foods, the disadvantages of solid-state processes cannot be neglected: Oxygen supply, temperature and pH value cannot be well regulated, the biomass is less homogeneous, and scale-up is difficult, as is the control of more rapidly growing bacterial contaminants [11].
Nonetheless, solid-state processes have been successfully established, including even the perhaps oldest process in food biotechnology, bacterial aerobic biofilm fermentation (for vinegar production): Phanerochaete chrysosporium or Trametes versicolor was immobilised on Ficus wood chips to treat the vinasse coming from a tequila factory. The biofilter was efficient as determined by the decrease in the chemical oxygen demand of the waste-water, and a laccase activity was proposed to be responsible [12].
Although it is generally accepted that enzyme catalysis is key to the development of more sustainable processes in the food and agro-industries [13], the main focus of recent work on solid-state fermentation is less on the production of enzymes (or valuable metabolites) but on using the substrate to show the degradation and valorisation of lignocellulosic agro-industrial side streams in terms of the bioeconomy concept [14] or merely as a nutrient source, either for the production of fruiting bodies or for specific biotransformations [15].
The in silico or activity-guided identification, amplification and transfer of basidiomycete genes coding for a target enzyme into a heterologous host has become a routine procedure. Functional production of, e.g., a dye-decolorising peroxidase (DyP) was possible without the expression of chaperons or the addition of hemin [16]. Unfortunately, the expression of fungal genes in Escherichia coli, even if corrected for codon usage, is frequently prone to fail. Enzymes commonly used in the food industries, such as amylases or peptidases, were found neither intracellularly nor secreted. The term “toxic gene” has been used to describe this phenomenon. However, one may speculate that the cell recognises “toxic” enzymes, which disturb metabolism right after they have left the ribosome, and eliminates them. A possible solution could be the cis-expression of genes in a fast-growing basidiomycete host [17]. Even if the protein expression is successful, recovery of substantial yields of correctly folded, active enzymes is a recurrent problem when using E. coli as a production host. Both low protein production and solubility have been addressed with varying success [18]. Another inherent disadvantage is the lack of glycosylation of the enzyme product.
Some of these problems can be overcome by using the Komagataella phaffii yeast (formerly known as Pichia pastoris). Chromosomal incorporation of the foreign gene affords stable mutants, and different signal sequences, truncations, tags or codon-optimised constructs are available [18]. While glycosylation may result in higher solubility, thermostability and unexpected catalytic properties, this approach does not necessarily guarantee a glycosylation pattern identical to the wild-type basidiomycete enzyme [19].
Novel fermentation processes for basidiomycetes were reported recently; one of them is the co-cultivation of two fungal strains. For example, the co-cultivation of Pleurotus ostreatus, the edible oyster mushroom, with Aspergillus niger (an ascomycetous producer of citric acid and various enzymes) improved the degradation of aflatoxin B1, a common mycotoxin in nuts and other foods [20]. A recent review showed how co-cultures of different strains of the same or different fungal species may stimulate each other, resulting in the induction of formerly silent polyketide synthase gene clusters, thus offering new options to generate bioactive polyketides [21]. Such observations call for in-depth investigation because many classical food fermentations rely on mixed cultures.
Fungal biotransformation processes often suffer from the low water-solubility and cytotoxicity of the substrate and product. Liquid–liquid or solid–liquid two-phase systems may offer solutions, as they separate the substrate and product from each other or from the producer cells. This applies not only to vegetative cells but also to fungal spores [22]. The author emphasised that spore cells should not be confused with resting cells because they are metabolically active and that spores are often solvent-tolerant enough to survive in the organic phase, which opens intriguing bioprocess options. The solid-liquid approach was successful for the in-situ isolation of hydrophobins from cultures of Schizophyllum commune [23]. Hydrophobins are small bipolar proteins with emulsifying properties, which renders their isolation problematic. Adsorptive binding to polytetrafluoroethylene microparticles allowed the separation from other proteins in the nutrient medium. Drawbacks of adsorption techniques are the sometimes-difficult desorption and the growth of the fungal cells on the adsorbent, which complicates the separation of the bound product from the spent biomass.
If there is no need for subsequent separation of cells and product, microparticles can act as a kind of “condensation nucleus” for the cells. This prevents the typical self-immobilisation and pellet formation, which is regularly observed in submerged fungal cultures. The resulting pellets may reach diameters of 10 mm or more and finally contain a core of autolysing cells with a living coat. These cell clusters offer protection against shear stress in the bioreactor but prevent maximum productivity. Changing the cultivation pH, temperature or the medium composition usually remains ineffective. The addition of artificial condensation nuclei circumvented the problem [24]. Water-insoluble inert organic or inorganic microparticles, such as silicates or ground bran, provided higher product yields and better cell cluster homogeneity at a lower cost [25].
Some requirements for the successful submerged cultivation of basidiomycetes remain to be identified. For example, remarkable effects of the rather unexpected trigger light on metabolism and enzyme production were found with the model wood degrader Phanerochaete chrysosporium [26]. Green light increased the production of fungal biomass and lignin peroxidase. The required light sensing elements, which are coupled to the fungal metabolism, are, as of now, unidentified. More systematic investigations into these and regarding other inducers of enzyme activity are required.
3. Processes Benefit from Deeper Genetic Knowledge
While more than 600 basidiomycete genomes have been sequenced to date [2,3], their annotation often remains incomplete or incorrect, which presents a major obstacle in homology searches, primer design and concerted in silico approaches [27]. With the idea of bioeconomic processes in mind, a lot of scientific attention is paid to genes coding for enzymes involved in the degradation of lignocellulosic side-streams; the same applies to fungi with medicinal uses [28,29]. Comparative bioinformatic genome mining and functional validation require large sets of similar gene sequences. Of more than 1000 putative feruloyl esterase genes, 27 were selected for heterologous production, 20 of which yielded active enzymes [30]: a busy approach but a practicable route to the discovery of misannotated genes.
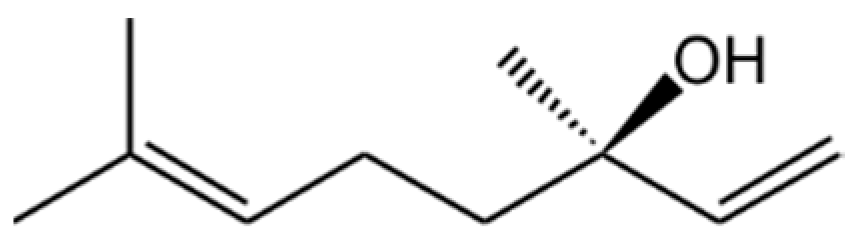
Other examples of the successful validation of predicted genes include the gene for a highly active linalool/nerolidol synthase. It was heterologously expressed in E. coli, where the resulting enzyme converted glucose to enantiopure (R)-linalool, a volatile flavour with attractive flowery notes and widespread applications [31] (Figure 1). Looking at the divergent evolutionary paths to bacterial, fungal and plant terpene synthases, this was a surprisingly successful experiment. Concerted mutations revealed the reasons for the superior catalytic properties of this enzyme.
Investigations at different stages of the fungal life cycle offer further insights. Differential expression of terpene synthase genes and a post-modification network were reported for the mycelia and fruiting body of Antodia cinnamomea, a species with applications in traditional Chinese medicine [32]. Such studies are particularly valuable because they improve our understanding of the factors regulating gene expression during fruiting body formation. Similar work has been carried out on the transcriptome level, and enzymes involved in biomass degradation and oxylipin biosynthesis were characterised [33,34].
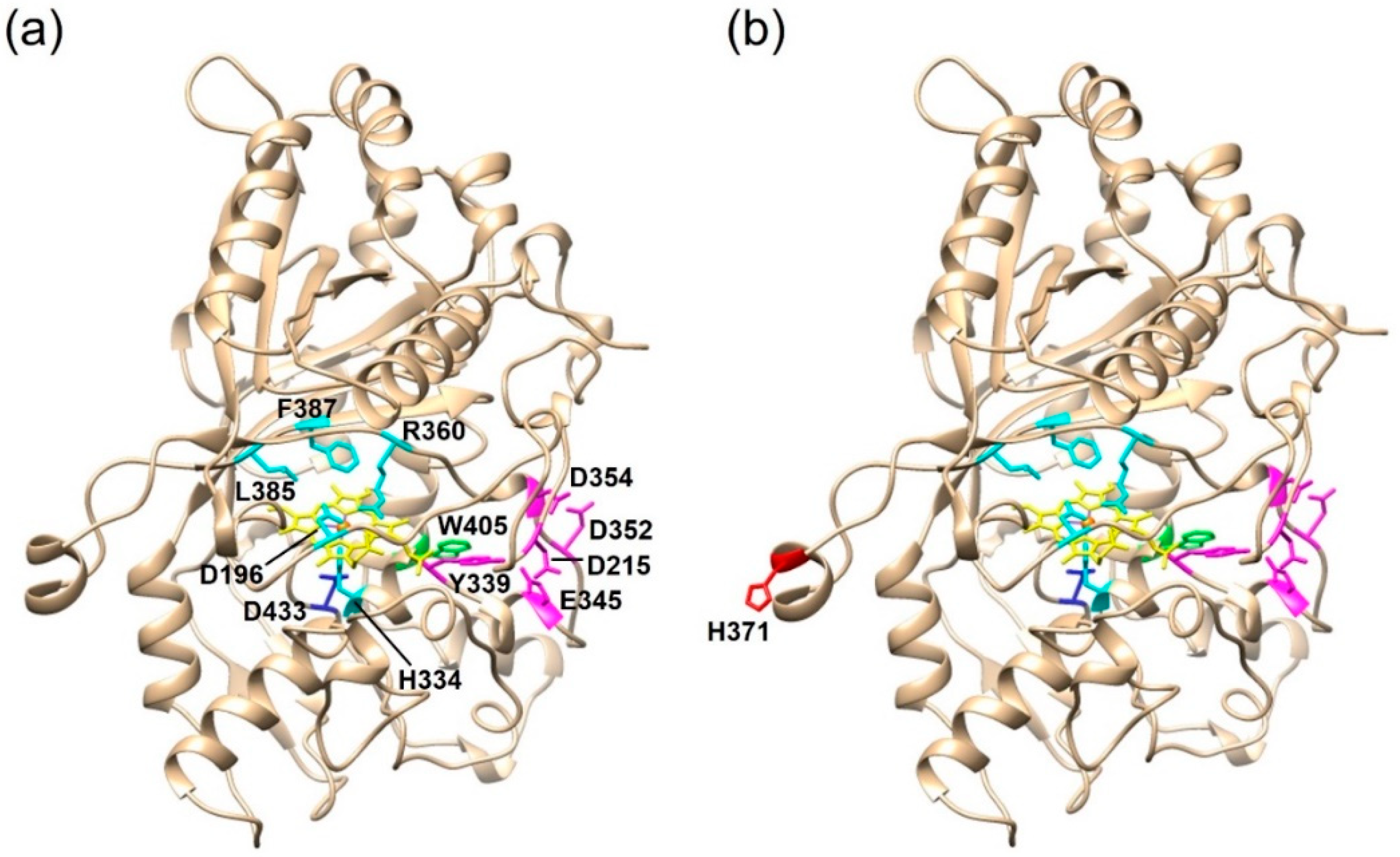
To come one step closer to lifting the riddle of intraspecific variations of basidiomycete strains, 101 monokaryons were derived from a single dikaryotic fruiting body [35]. Using DyP activity as a model reaction, it was found that the cultures could be grouped into low, middle and high-activity strains, which contained three enzyme variants with up to eleven point mutations (Figure 2). Subsequent transcriptional analyses additionally revealed different levels of gene expression. Thus, mutation events and differential gene expression were co-responsible for the observed differences in the peroxidase activity. A different study excluded both for the increase in monokaryotic carotene degrading activity [36]. Thus, no general reason for intraspecific variations in enzyme activity could be identified so far. The respective differences have to be evaluated on a case-by-case basis until more data are available.
4. Lipolytic Processes
Lipases are widely used in the production of tailored food lipids, such as milk fat or cocoa butter substitutes, but also for enhanced cheese ripening, the generation of fruit ester in reverse hydrolytic processes or in detergent formulations [37]. Surprisingly few reports exist on basidiomycetous lipases. A major reason may be that the representatives from Candida and the common porcine pancreas fulfil many of the industrial needs. When contacted with lipid-rich substrates, such as rapeseed press cake, however, even xylophilic Basidiomycota, here Pleurotus ostreatus, started to secrete lipases in solid-state cultures [38]. This quick adaption to environmental conditions is one of the most remarkable features of basidiomycetes. A member of the Ustilaginomycotina, Malassezia globosa, contained an unusual lipase specific for mono- and diacylglycerols [39]. A point mutation converted the wild-type enzyme into a classic triacylglycerol lipase. It may be speculated that a point mutation of the original gene changed the enzyme’s properties, so that the unexpected substrate specificity was the result of an incident without adverse effects on the fungus. Further examples on how single amino acid exchanges result in dramatically altered enzyme properties will be given below.
5. Esterolytic Processes
Hemicelluloses are linked by ester and glucosidic bonds. The associated feruloyl esterases have received a lot of attention. Depending on their preferred substrate, different nomenclatures (e.g., chlorogenic acid, p-coumaroyl esterase) can be found.
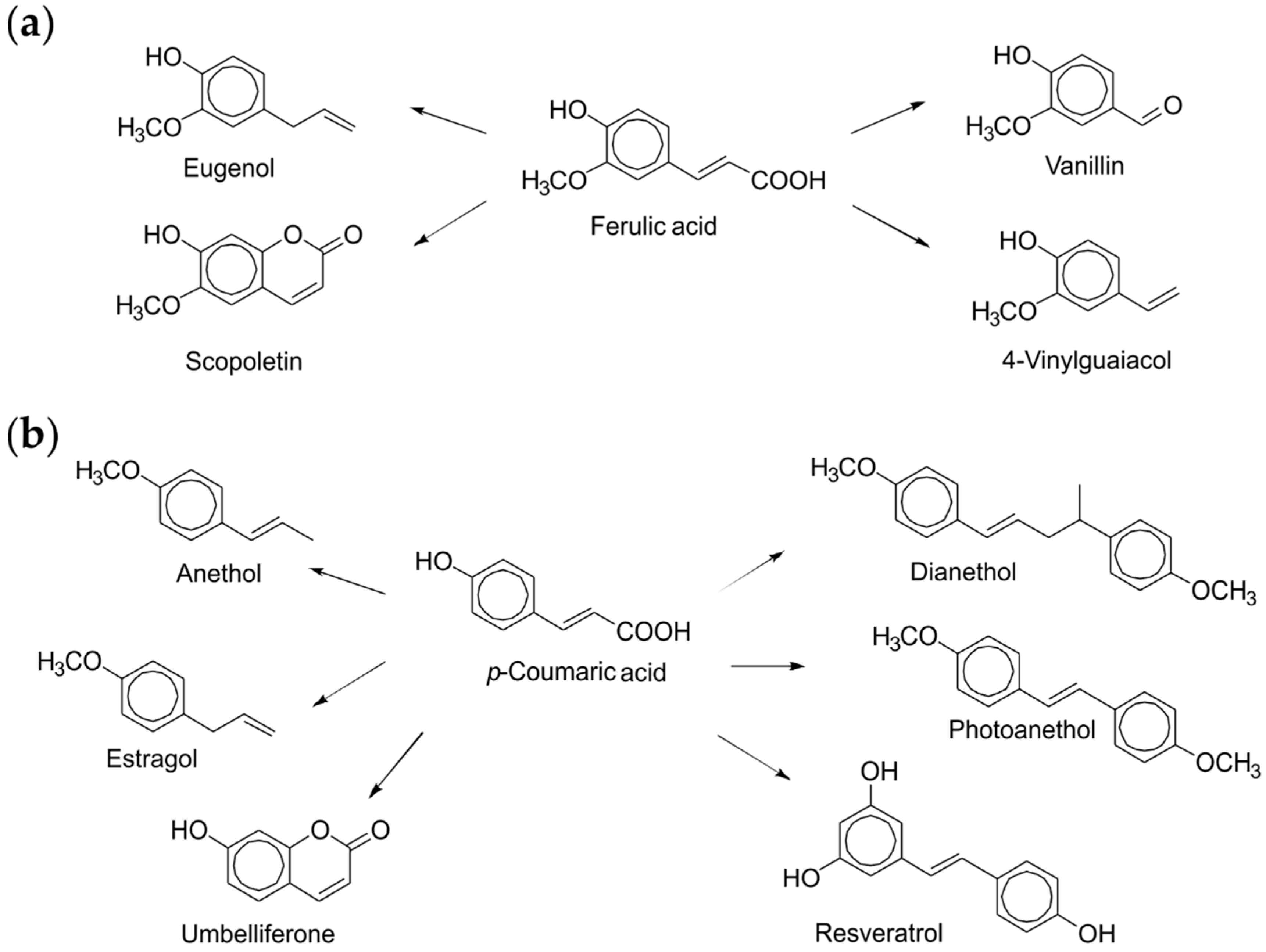
Ustilago maydis, a basidiomycete growing on maize (Zea mays) and consumed as a food delicacy (huitlacoche) in Mexico, produced an intriguing chlorogenic acid esterase when grown submerged. The recombinant enzyme released ferulic, p-coumaric and caffeic acid from renewable substrates, such as destarched wheat bran and coffee pulp. Enzyme-treated complex plant biomass can thus serve as a source of C6C3-building blocks, useful as antioxidants or preservatives in the food, health, cosmetic and pharmaceutical industries (Scheme 1) [40]. The addition of this enzyme to wheat dough resulted in a noticeable softening, offering a new tool for manipulating the texture of bakery and confectionery products without classical chemical baking aids [41].
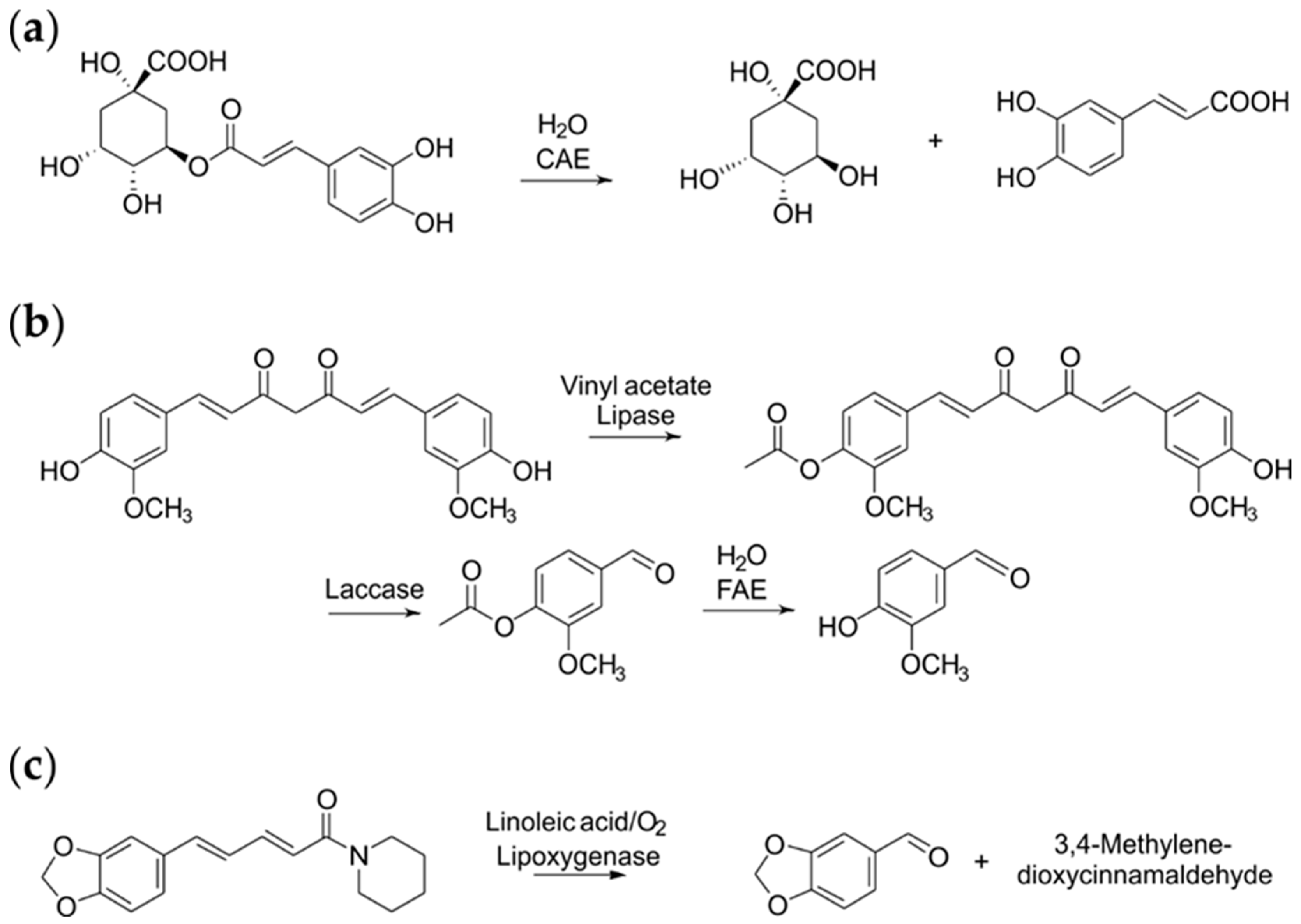
A recombinant feruloyl esterase from Schizophyllum commune accepted various benzoic acid esters [42] and kaempferol 3-O-(2‴-O-sinapoyl-β-sophoroside), the main bitter compound of rapeseed protein isolates, as substrates [43]. As a result, a simple hydrolytic reaction, i.e., the cleavage of the sinapic acid from the sophoroside moiety, turned a feed into a food.
The enzyme class was also capable of hydrolysing chlorogenic acid in coffee powder and beverages. This should contribute to an improved digestibility without compromising the antioxidant capacity (Scheme 2a) [44]. The same reaction principle was successful in apple juice, without affecting the likewise contained fruit esters [45].
Phytic acids are a group of oligo-phosphorylated inositols occurring in plants and decreasing the bioavailability of minerals in food and feed due to their metal ion complexing ability (“anti-nutrients”). Phytases region-specifically cleave phosphate ester bonds and are used especially in feeds for non-ruminants (monogastrics), such as poultry or pigs. A positive side-effect is the decreased phosphate load of the manure. Representatives of these enzymes were found in submerged and solid-state cultures of the Ganoderma species [48,49]. Enzyme solutions were mixed with maltodextrin and spray-dried to yield stable preparations with a half-life of activity for more than one year. Sodium chloride and polyethylene glycol inhibited microbial contamination of the preparations, rendering them applicable to food and feed formulations.
6. Glycolytic Processes
In their natural habitats, basidiomycetes encounter many different saccharides, mainly polysaccharides. Accordingly, their potential to secrete glycosidases ranges from amylases over cellulases and hemicellulases to pectinases and more. Lentinula edodes powder, a traditional food in Japanese cuisine, was mixed with potato tuber, and a rapid release of saccharides was observed under heterogeneous solid-state conditions [50]. This observation proved that a mushroom powder served as a kind of natural immobilisation matrix and preserved enzyme activities. One of the problems is the need for enzyme mixtures for the solubilisation of complex food fractions, such as fruit peel or press cakes, as mixtures are hard to implement in reproducible technological processes.
Like the feruloyl esterases in the field of esterases, α-galactosidases are a focal point in the research on glycosidases; the reason is the recent interest in meat substitutes. Many plant protein sources contain α-galactosyl derivatives of sucrose, such as the trisaccharide raffinose, which are hydrolysed by α-galactosidase, an enzyme missing in the human digestive tract. Multi-step fractionation is required to remove the oligosaccharides; otherwise, they are fermented by bacteria in the large intestine. Short-chain fatty acids (acetic, propanoic, butanoic acids) and gases are produced, leading to the well-known flatulence commonly associated with eating beans and other vegetables.
Fungal α-galactosidases were discovered in Irpex lacteus [51,52], Leucopaxillus tricolor [53] and in Tremella aurantialba [54]. These enzymes, some of which are available in recombinant form, were generally very active, thermostable and working within a broad pH range. It may be supposed that the high substrate specificity of enzymes will help to concertedly remove the oligosaccharides from plant extracts and diminish the number of purification steps to obtain food-grade protein-rich materials.
7. Peptidolytic Processes
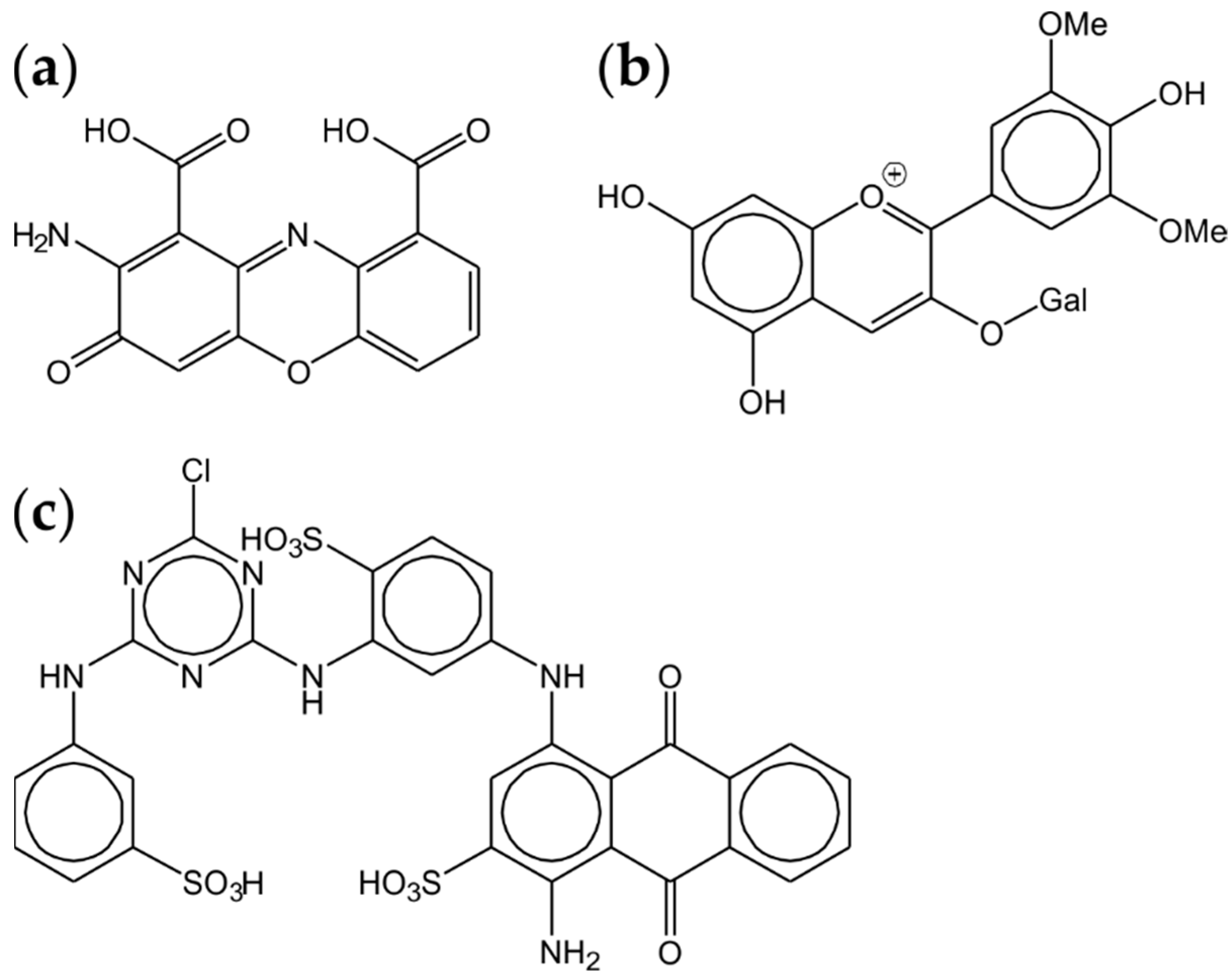
Nitrogen-containing compounds are rare in the natural substrates of xylophilic fungi. To cope with the problem, evolution has equipped basidiomycetes with a multitude of peptidase coding genes. A colourful range of reported applications is the result. As mentioned above, stability under harsh conditions was often observed. A serine-peptidase from Pleurotus sajor-caju, a close relative of the oyster mushroom, showed pH and temperature optima at 9.5 and 70 °C and high stability against detergents and organic solvents [55]. The enzyme was purified by precipitation at 80 °C for 20 min (!) followed by ammonium precipitation and FPLC steps. It may be of interest for high-temperature processes (Figure 3).
The strong increase in worldwide cheese production over the past decades has forced dairy scientists to search for chymosin substitutes. Common peptidases are inapplicable because the non-specific peptidolysis of caseins inevitably results in the generation of very bitter tasting peptides. Recombinant and microbial chymosins are available but not well appreciated by traditional cheese-makers. Aspartic and metallopeptidases isolated from Phanerochaete chrysosporium [56] and Termitomyces clypeatus [57] cleaved off the glycosyl macropeptide of κ-casein and were suggested as chymosin-like milk-clotting substitutes.
Driven by the idea to improve food safety and quality, a gluten-degrading prolyl peptidase was isolated from Flammulina velutipes, the golden needle mushroom (enoki), for the degradation of celiac disease-provoking toxic peptides in cereal proteins [58]. Aqueous extracts from Sarcodon aspratus, an ectomycorrhizal basidiomycete, hydrolysed myosin and actin in bovine muscle tissue and might serve as a sustainable replacement for plant meat tenderisers, such as papain or bromelain [59].
8. Redox Processes
Aiming at the liberation of glucose enclosed in lignocellulosics for biofuel production, a lot of the recent research focused on lignin degradation and consequently on oxidoreductases, such as peroxidases [60,61,62] and laccases [63,64], which are secreted by xylophilic basidiomycetes to secure their nutrient demands.
The second major field for the use of oxidoreductases is the selective removal of only the unwanted constituents in the multi-component matrix of a food, as has been exemplified above. Incidental contamination of food by environmental toxicants may occur during production, processing or post-processing and is difficult if not impossible to control. Taking advantage of the selective (or even specific) action of oxidative enzymes has been widely recognised as a convenient solution for bioremediation in general [65]. Laccases and tyrosinases, for example, from Pleurotus ostreatus or Trametes versicolor, degraded all kinds of phenols in model mixtures, wastewater or sewage. Many promising reports induced the search for food-grade specimens and applications. A recurrent drawback of recent studies is that the desired activity was demonstrated in model systems, in buffered or in immobilised form [66,67], but not in real foods. For example, laccases served the purpose in the presence of artificial mediators, but these do not occur in foods. While it is an intriguing idea to use spent mycelia or even entire fruiting bodies of Agaricus bisporus (button mushroom) or Pleurotus ostreatus as biocatalysts, the enzymology behind such processes sometimes remains obscure [68,69]. On the other hand, a broader range of toxicants may be attacked by a mixture of enzyme activities.
Ascomycetes, such as Aspergillus, Penicillium, Claviceps, Paecilomyces and Fusarium, under special conditions, produce mycotoxins to suppress local bacterial or fungal competitors, with often considerable toxicity to humans as well. Among foods, nuts, crops, cereals and spices are most frequently affected, but feeds, particularly silage, can also be contaminated and even kill animals fed on it. Not surprisingly, many reports on mycotoxin-degrading activities have been published. Aflatoxins, a group of mycotoxins, which resulted in mass killings of people and animals in the past, have received the utmost attention [70,71,72,73]. From spent mycelia over crude extracts, more or less characterised enzyme isolates to recombinant activities, all levels of enzyme purity occur in the literature. Further, mycotoxins, such as deoxynivalenol [74] and zearalenone [75], were also subject to enzymatic degradation. In numerous cases, the enzyme preparation applied did not degrade one mycotoxin specifically. This immediately raises the question of how such preparations would perform within the complex chemistry of a food. Surface contamination may be removed by washing or immersing the food. Liquid foods also lend themselves well to the application of enzymes; beer, wine or juices may be treated with enzyme immobilisates. However, sufficient contact of enzyme and mycotoxin is difficult to achieve, if not impossible, in creams, powders or solid foods.
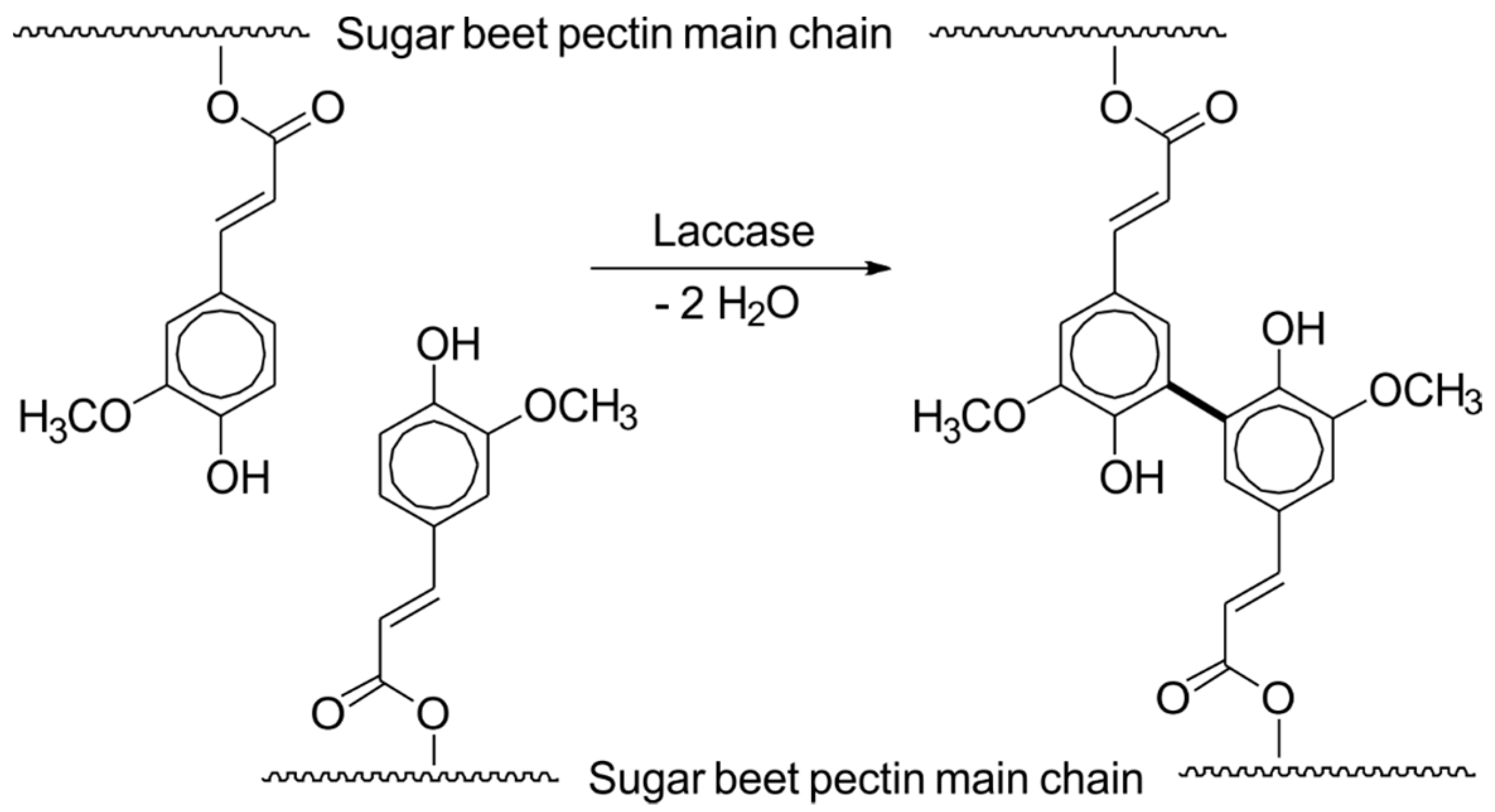
In addition to the removal of unwanted contaminants, oxidoreductases have also been used to generate new food ingredients. Laccases from Funalia trogii and Pleurotus pulmonarius crosslinked feruloylated arabinoxylans, which resulted in stable and solid gels with high water holding capacity, which are comparable to gelatine gels. They thus present an alternative for vegan, halal and kosher food. The soluble arabinoxylan substrate may be obtained from wheat bran or sugar beet fibre, both renewable low-cost side-streams of the agro-industries (Scheme 3) [76,77].
Softening of the cross-linked gel can be achieved using the above-described ferulic acid esterases, while the reverse transferuloylation to yield feruloyl-saccharide esters was shown for mono and disaccharides using a recombinant enzyme from Pleurotus sapidus in a low-water-activity medium [78]. The selective and mediator-free oxidation of lactose to lactobionic acid, a compound useful in food and further applications, was reported using a cellobiose dehydrogenase-laccase system from a Pycnoporus [79]. The authors speculated that 3-hydroxyanthranilic acid, produced by the fungus itself in submerged culture, may have served as an internal mediator for the reaction.
9. Formation and Degradation of Colourants
Without a doubt, colours dominate the sensory attractiveness of foods. Losses due to prolonged storage or high processing call for supplementation. On the other hand, the decolourisation of foods may also be desirable. Examples are the bleaching of annatto in Cheddar cheese whey, achieved by the first basidiomycete enzyme that went commercial (Maxibright), or the bleaching of molasses or of base wines for the production of wine coolers [80,81,82]. While basidiomycetes generate colours through anabolic pathways (shikimate, polyketide, mevalonate), the degradation is left to oxidoreductases, mainly laccases and peroxidases. Examples of both processes will be presented in this paragraph.
A recent review covered fungal colour formation by solid-state processes [83]. Co-lours such as the orange to red laetiporic acids were produced in both solid and liquid cultures [84]. The addition of colourants to food is subject to strict legal regulations in most countries. Novel colours from basidiomycetes might thus be of interest from a scientific point of view but will have to undergo extensive toxicological evaluation before being approved for incorporation into food. Colours from edible fungi would be preferred for obvious reasons.
It is a long-standing practice in food processing to accelerate and steer costly ripening processing by the addition of enzymes, either in the form of starter cultures or as technical enzyme preparations. This idea worked well for green tea, which was treated with a commercial laccase or a tyrosinase from Agaricus bisporus to raise the concentration of yellow-orange theaflavins, the major colourants and tastants in black tea [85]. A two-phase octanol-buffer system improved theaflavin yield and enabled its in-situ extraction [86].
Pycnoporus sanguineus, a wood-rotting saprobiont, produces the deep-red colourant cinnabarinic acid (Figure 4a) [87], but also a laccase, which degraded synthetic dyes, such as Remazol Brilliant Blue R, Reactive Black 5, Reactive Red 195 and Reactive Yellow 145, when cultivated on sugarcane molasses [88]. High-redox potential laccases are also capable of degrading food colours. During screenings for enzymes decolorising stains on laundry, culture supernatants of Pleurotus pulmonarius were found to be particularly active. Laccases were isolated as the responsible activities. Even in the absence of a mediator, the enzymes degraded, among others, malvidin-3-O-galactoside, an anthocyanin widely occurring in berries and major colourant of red wine husks (Figure 4b) [89]. The same objective was achieved using a β-glucosidase from Bjerkandera adusta [90]. The recombinant enzyme possessed a good temperature and pH stability and preferably catalysed the hydrolysis of delphinidin-3-glucoside and cyanidin-3-glucoside and worked in an aqueous solution as well as on stained fabrics.
Versatile peroxidases (vPOX) from basidiomycetes are a major lignolytic activity of Basidiomycota, which combine properties of both manganese and lignin peroxidases. The vPOX of Pleurotus ostreatus oxidised not only Mn2+, but also various colourants, each at different sites of the enzyme and under different conditions [91]. What was developed by nature to allow the fungus to adapt to different environmental conditions may open options of bleaching food colours by adjusting the external conditions, particularly the pH value. Likewise interesting for the decolourisation of food is Bjerkandera adusta. It contains a broad spectrum of peroxidases, which, in addition to a vPOX, lignin and manganese peroxidases, also includes a horseradish-like activity, which was the most active for the decolourisation of melanoidins in sugar beet molasses [92].
While laccases and phenol oxidases as multi-copper enzymes transfer electrons directly on molecular oxygen as the single co-substrate, peroxidases depend on the availability of hydrogen peroxide. This imperative requirement is fulfilled in submerged cultures by complementary flavoenzymes, such as aryl-alcohol oxidases. A recent review summarised the substrates and the problematic heterologous production of these enzymes [93]. While their ecological role in lignin-degraders (white-rot fungi) is to supply the necessary hydrogen peroxide, they are required for the Fenton reaction in carbohydrate-degrading (brown-rot) species [94]. Suggested food applications of aryl-alcohol oxidases range from flavour synthesis over secondary alcohol deracemisation to the oxidation of furfurals for the production of furandicarboxylic acid as a building block for biodegradable packaging materials [95]. A work combining an aryl-alcohol oxidase from Pleurotus sapidus with a DyP from Mycetinis scorodonius proved the principle [96]. The gene from P. sapidus was linked with a promotor from Agaricus bisporus and the expression vector transformed into Coprinus cinerea, another basidiomycete. The two-enzyme system bleached Reactive Blue 5, a large anthraquinone structure (Figure 4c).
10. Formation and Degradation of Food Components
Processes aiming at a single hydrolytic or redox step typically require one enzyme activity only. When it comes to more complex reactions, the use of whole-cell systems is inevitable. Oligo- and polysaccharides are present as nutrients as well as thickeners in many foods. A recent review dealt with the pathways of mushroom polysaccharide composition, formation and its regulation [97].
Some common constituents of foods, such as gluten (cf. “Peptidolytic Processes”), lactose or chlorogenic acids (cf. “Esterolytic Processes”), pose health problems to certain population groups. Phenylketonuria is another example of such a food intolerance. A remedy could be provided by a phenylalanine ammonium lyase isolated from the red basidiomycetous yeast Rhodosporidium toruloides [98]. The enzyme worked on various protein pre-hydrolysates and removed l-phenylalanine efficiently.
l-Asparaginases not only possess medical applications but are used to combat the formation of the undesired acrylamide in roasted foods, such as coffee, toast or French fries. Only two asparaginases have been commercialised so far, both from Aspergillus species. Following a typical activity-guided screening, an l-asparaginase was discovered in Ganoderma australe, a basidiomycete [99].
11. Formation of Volatile Flavours
Volatile and non-volatile flavours determine the freshness, typicity and quality of a food and hence its market success. Flavours are often expensive, which has stimulated a lot of research on their generation and isolation. Traditional fermentation and thermal processes (malt, beer, cocoa, coffee, etc.) result in complex flavour mixtures with hundreds of compounds [100]. Obviously, such mixtures cannot be reproduced by single enzyme processes. Bioprocess development, therefore, has to focus on character impact compounds, which govern the sensory impression by their particular sensory note and have a low detection threshold.
A prime example is vanillin. Countless attempts for biotechnological production reflect its superior market importance. For example, curcumin is a cheap precursor bearing two 4-hydroxy-3-methoxyphenyl substructures. Treatment with laccases, however, yielded rapid oligomerisation. Thus, one of the phenolic hydroxyls was selectively protected using an acetyl donor and a lipase, the resulting monoacetyl curcumin was exposed to a laccase from Funalia trogii, and the acetyl vanillin obtained deprotected using a ferulic acid esterase from Pleurotus eryngii (Scheme 2b) [46]. Various laccases were compared, and it was found that high-potential laccases performed well in this mediator and cofactor-free reaction.
In general, the oxidative cleavage of α,β-double bonds next to an aromatic ring is a route to produce aryl aldehydes with sensory properties useful for the food industry. Thus, p-anisaldehyde, veratraldehyde and acetophenone were generated by a DyP from Pleurotus sapidus, recombinantly produced in E. coli and Komagataella phaffii [101]. Surprisingly, bleaching of annatto and β–carotene was also observed. An analogous oxidation, but through a different mechanism, was observed using piperine, the cheap, major alkaloid of pepper, yielding the vanilla-like flavour compounds piperonal and 3,4-methylenedioxycinnamaldehyde (Scheme 2c) [47]. The reaction was initiated by the hydroperoxidation of an unsaturated fatty acid, such as linoleic acid, by a lipoxygenase from Pleurotus sapidus, followed by the co-oxidation and chain cleavage of piperine.
Lentinula edodes (shiitake) is one of the most popular edible fungi worldwide, and particularly researchers from Asia have studied its flavour meticulously. The fruiting body contained 82 volatile compounds, 25 of which are important for the overall sensory impression [102]. Drought stress stimulated the accumulation of the flavour precursor lenthionine and of the lenthionine precursor cysteine [103]. Transcriptome studies revealed 30 genes associated with histidine, glutathione and unsaturated fatty acid metabolism as related to the formation of the typical volatile compounds [104]. Looking at this wealth of data from the genome, transcriptome and metabolome levels, it should be possible to design submerged cultivation processes with mycelia yielding an optimised production of shiitake volatiles.
The plethora of sesquiterpenes from all classes in basidiomycetes has been known for decades. Main users of these essential oil compounds are the flavour, fragrance, cosmetics and pharmaceutical industries. However, until recently, genes coding for sesquiterpene cyclases, which accept farnesyl diphosphate as a substrate, remained obscure. Genome mining of Agrocybe aegerita (black poplar mushroom) resulted in the identification of nine functional synthases [105]. Based on sequence similarity, 1133 additional putative synthase genes were identified in a species-wide screening. Five new synthases from five fungi were correctly predicted. Recombinant synthases from Coniophora puteana efficiently and highly selectively produced β-copaene and cubebol in E. coli [106]. Single and multi-product synthases were found in Cerrena unicolor [107]. Processes using recombinant E. coli strains could substitute the common essential oil plants, especially where the plants are jeopardised by climate change or extinction by human intervention.
One experimental hurdle is presented by the low similarity of plant and fungal synthases and even between the various clades of basidiomycetes, which impedes data bank searches; another is the lack of knowledge on induction mechanisms of the coding genes. A recent report supposed that polysaccharides occurring in the natural habitat of xylophilic fungi could affect the formation of sesquiterpene synthesis [108]. Tyromyces floriformis, a potent producer of sesquiterpenes, such as α-Ylangene, was chosen as a model system. In contrast to all expectations, the polysaccharides or even a trisaccharide were inhibitory, whereas the presence of the monomeric arabinoxylane building blocks showed no effect. It was concluded that the formation of volatile sesquiterpenes was not associated with the growth substrate but rather connected with yet unknown ecological functions.
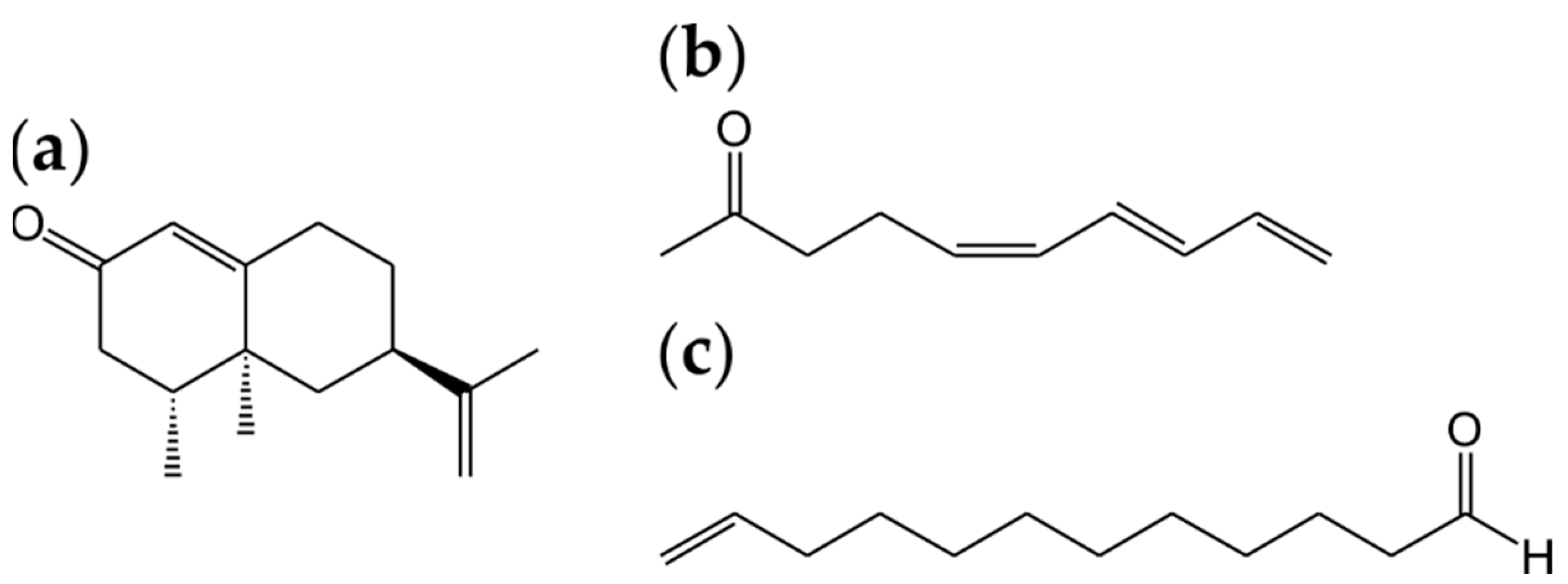
A route to high product yields is offered in cases where an easily available precursor is transformed to the target product, as in the case of (+)-valencene and the high-value grapefruit character impact compound (+)-nootkatone [109]. The combined action of a DyP and a laccase from Funalia trogii resulted in a complete conversion of (+)-valencene within 24 h (Figure 5a).
An excellent proof of the enormous synthetic potential of basidiomycetes was the identification of the (5E/Z,7E,9)-decatrien-2-ones in submerged, cultured Fomitopsis betulina (Figure 5b) [110,111]. The (5Z)-isomer possessed a strong pineapple flavour and belongs to the most potent flavour compounds known. The volatile does not occur in pineapple fruit but shows high structural similarity to 1-(3E,5Z)-undecatriene, a genuine character impact hydrocarbon of this fruit. Likewise unusual were dodec-11-enoic acid and di- and tri-unsaturated C16 isomer fatty acids in the lipids of Flammulina velutipes [112]. Their observation was used to generate ω1-unsaturated aldehydes through α-oxidation, with dodec-11-enal possessing a pleasant coriander/leaf-like smell (Figure 5c).
A ferulic acid esterase (see above) from Rhizoctonia solani produced by Komagataella phaffii acting on wheat bran arabinoxylan was combined with an unusual decarboxylase from Schizophyllum commune to yield the smoke flavour impact 4-vinylguaiacol [113,114]. The cold process did not produce any unwanted and toxic side-products, which are inevitably produced during classical smoking. The process was simplified using immobilised enzymes and the in-situ extraction with the adsorbent XAD4.
While the potential of basidiomycetes for the production of volatile flavours is clearly far from being fully exploited, solid-state [115] and submerged cultures [116] were shown to provide suitable experimental systems to valorise renewable biomass from food processing, such as pomace, molasses, leaves or peels. This allows the use of non-food biomass as a cheap source of nutrients and, at the same time, provides a sink for these large volume side-streams. Proverbially, the old dream of the alchemists to turn lead to gold comes true through these novel fermentation processes.
12. Formation of Non-Volatile Flavours
Edible basidiomycetes are not only appreciated for high fibre and mineral concentration and low energy content but also for their taste. Most of the recent research work on non-volatile flavours revolved around peptides with taste (enhancing) properties. Apart from the four well-known gustatory human senses, umami, described as hearty-meaty-spicy, was identified as the fifth one. Triggered by the amino acids l-glutamate and l-aspartate and the nucleotide ions inosinate and guanosinate, the umami receptors T1R1 and T1R3 also respond to the presence of certain peptides, which, for example, also occur in hard cheeses [117].
Shiitake (Lentinula edodes) and members of the genus Pleurotus repeatedly received the highest scores in umami taste evaluations. In fruiting bodies of Pleurotus geesteranus (pocket-sized oyster) the highest levels of umami taste and concentrations of adenosine phosphates (AMP, ADP and ATP) were found in the sample stored at 5 °C [118]. At these low temperatures, fruiting body formation of fungi of the moderate climate zones is often observed in nature. Di- and tripeptides Gly-Cys-Gly, Glu-Pro-Glu, Cys-Met, Val-Phe and Gly-Glu were identified in the low-molecular mass sub-fraction of shiitake, which possessed the strongest umami taste [119]. When the extraction of free glutamate and aspartate from various basidiomycetes was supported by the addition of a β–glucanase and the peptidase cocktail Flavourzyme, detailed amino acid analyses showed that the best yields were obtained from two Agaricus strains [120]. Bioactivities of peptides beyond taste comprise, for example, antihypertensive and antimicrobial effects, as was reviewed recently [121]. To create a more sustainable source of fungal peptides, such results must be transferred to fermentation processes. Peptides could then be isolated from the ever-growing cells or produced using suitable protein substrates and selected peptidases out of the spectrum of activities secreted by the mycelia (cf. “Peptidolytic Processes”).
As excessive sodium consumption may result in severe health problems, such as hypertension, oedema or gastric inflammation, reducing the sodium chloride contents in foods is one of the main aims of Europe-wide research efforts. Salt taste-enhancing l-arginyl dipeptides (which do not possess salt taste on their own) were obtained by incubating lysozyme or caseins with fungal culture supernatants, as shown by LC-MS/MS and sensory evaluations [122]. Supplementation with the fungal hydrolysates compensated for the reduced salt content of a model cheese matrix and of curd cheese.
13. Outlook
Recent years have seen a strong upsurge in interest in food additives or supplements (“nutraceuticals”), which are supposed to exert beneficial effects on human health. Based on the long-standing history of edible fungi in folk medicine, especially in Asia, fermentation processes using basidiomycetes have thus received a new impetus. From low-molecular mass compounds, such as the antiallergic alkaloid hispidin [123] or the sterol ergosterol [124] to the stilbenoid resveratrol [125] (which is seemingly able to cure everything; cf. Scheme 1b), and the anti-aging [126] and anti-cancer exopolysaccharides [127], to mention only a few recent examples, the cupboard of fungal medicines is well stocked. The accumulated knowledge calls for the transfer of the findings to fermentation processes: Fruiting bodies can only be extracted once, which is obviously not a sustainable procedure. With larger amounts of the presumed bioactives at hand, the scientific foundation of many of the claimed effects would also be facilitated.
The gills at the underside of the fruiting body of a typical macrofungus contain millions of column-shaped cells, the name-giving basidia. On appendages from each basidium, four reproductive haploid spores are produced through meiosis. The spores develop into elongated growing cells, which may fuse with a suitable mating-type partner to form dikaryotic cells again. Using a sesquiterpene biotransformation as a model reaction, it was shown that the sexual event created under as well as over-performing monokaryots compared to the dikaryotic parent [128]. The same referred to monokaryotic strains, which produced a β–carotene degrading laccase [36]. The genetically diverse monokaryotic mycelia performed differently and were stable over many sub-cultivations. This opens the chance to select overproducers without genetic manipulations, which are still rejected by most customers.
Each of the fungal processes described above ends with a fermenter full of more or less homogeneous biomass. According to the concept of circular economy, this biomass should find reasonable reuse. As the mycelia are genetically identical to the fruiting body, they may be directly marketed as an ingredient of vegan/vegetarian foods if the species has a history of safe use as an edible mushroom (“generally recognised as safe” in the US). Other ideas include the separation of the biomass into a protein and a polysaccharide fraction to combat the threatening protein crisis, as well as use of the chitin/β-glucan fractions as renewable materials to produce foams, paper, textiles and even leather substitutes [129]. Also proposed were the development of fertilisers, the use as a substrate for other (fungal?) fermentation processes after autolysis, the use as feed additive for animals or as a constituent in packaging and construction materials [130]. Monocoque constructions of entire houses made of composite material of, for example, bamboo fibres and mushroom mycelia, can be admired on some internet pages [131,132,133]. The selection of the most appropriate basidiomycete and fermentation process conditions may be oriented and optimised towards the specific demands of such non-food uses.
Supplementary Materials
The following supporting information can be downloaded at https://www.mdpi.com/article/10.3390/pr10040726/s1, Table S1: Overview of the different basidiomycetous enzymes, their use for foods and production hosts of recombinantly available enzymes.
Author Contributions
Writing—original draft preparation, R.G.B.; writing—review and editing, F.E.; visualization, R.G.B. and F.E.. All authors have read and agreed to the published version of the manuscript.
Funding
This review received no external funding. The publication was funded by the Open Access Fund of the Gottfried Wilhelm Leibniz Universität Hannover.
Acknowledgments
The authors gratefully acknowledge the meticulous work contributed by the head of analytics U. Krings and by the members of their working groups as they occur in the papers cited.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Nurk, S.; Koren, S.; Rhie, A.; Rautiainen, M.; Bzikadze Andrey, V.; Mikheenko, A.; Vollger Mitchell, R.; Altemose, N.; Uralsky, L.; Gershman, A.; et al. The complete sequence of a human genome. Science 2022, 376, 44–53. [Google Scholar] [CrossRef] [PubMed]
- Nordberg, H.; Cantor, M.; Dusheyko, S.; Hua, S.; Poliakov, A.; Shabalov, I.; Smirnova, T.; Grigoriev, I.V.; Dubchak, I. Basidiomycota. Available online: https://mycocosm.jgi.doe.gov/basidiomycota/basidiomycota.info.html (accessed on 8 March 2022).
- Nordberg, H.; Cantor, M.; Dusheyko, S.; Hua, S.; Poliakov, A.; Shabalov, I.; Smirnova, T.; Grigoriev, I.V.; Dubchak, I. The genome portal of the Department of Energy Joint Genome Institute: 2014 updates. Nucleic Acids Res. 2014, 42, D26–D31. [Google Scholar] [CrossRef] [PubMed]
- Arntzen, M.; Bengtsson, O.; Várnai, A.; Delogu, F.; Mathiesen, G.; Eijsink, V.G.H. Quantitative comparison of the biomass-degrading enzyme repertoires of five filamentous fungi. Sci. Rep. 2020, 10, 20267. [Google Scholar] [CrossRef] [PubMed]
- Coconi Linares, N.; Fernández, F.; Loske, A.M.; Gómez-Lim, M.A. Enhanced Delignification of Lignocellulosic Biomass by Recombinant Fungus Phanerochaete chrysosporium Overexpressing Laccases and Peroxidases. J. Mol. Microbiol. Biotechnol. 2018, 28, 1–13. [Google Scholar] [CrossRef]
- Xu, L.; Chen, B.; Geng, X.; Feng, C.; Meng, J.; Chang, M. A protease-resistant α-galactosidase characterized by relatively acid pH tolerance from the Shitake Mushroom Lentinula edodes. Int. J. Biol. Macromol. 2019, 128, 324–330. [Google Scholar] [CrossRef] [PubMed]
- Lanfermann, I.; Linke, D.; Nimtz, M.; Berger, R.G. Manganese peroxidases from Ganoderma applanatum degrade beta-carotene under alkaline conditions. Appl. Biochem. Biotechnol. 2015, 175, 3800–3812. [Google Scholar] [CrossRef]
- Yin, L.; Ye, J.; Kuang, S.; Guan, Y.; You, R. Induction, purification, and characterization of a thermo and pH stable laccase from Abortiporus biennis J2 and its application on the clarification of litchi juice. Biosci. Biotechnol. Biochem. 2017, 81, 1033–1040. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.; Tian, G.; Zhao, L.; Wang, H.; Ng, T.B. A protease-resistant α-galactosidase from Pleurotus djamor with broad pH stability and good hydrolytic activity toward raffinose family oligosaccharides. Int. J. Biol. Macromol. 2017, 94, 122–130. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Zhu, M.; Tian, G.; Zhao, L.; Wang, H.; Ng, T.B. Isolation of a protease-resistant and pH-stable α-galactosidase displaying hydrolytic efficacy toward raffinose family oligosaccharides from the button mushroom Agaricus bisporus. Int. J. Biol. Macromol. 2017, 104, 576–583. [Google Scholar] [CrossRef]
- Soccol, C.R.; Costa, E.S.F.d.; Letti, L.A.J.; Karp, S.G.; Woiciechowski, A.L.; Vandenberghe, L.P.d.S. Recent developments and innovations in solid state fermentation. Biotechnol. Res. Innov. 2017, 1, 52–71. [Google Scholar] [CrossRef]
- Marco Antonio, G.Z.; Angélica Julieta, A.R.; Esperanza, R.C.; Gerardo, B.; Gerardo, D.G.; Edson Baltazar, E.A. Evaluation of Ficus benjamina wood chip-based fungal biofiltration for the treatment of Tequila vinasses. Water Sci. Technol. 2018, 77, 1449–1459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loi, M.; Glazunova, O.; Fedorova, T.; Logrieco, A.F.; Mulè, G. Fungal Laccases: The Forefront of Enzymes for Sustainability. J. Fungi 2021, 7, 1048. [Google Scholar] [CrossRef] [PubMed]
- Tišma, M.; Žnidaršič-Plazl, P.; Šelo, G.; Tolj, I.; Šperanda, M.; Bucić-Kojić, A.; Planinić, M. Trametes versicolor in lignocellulose-based bioeconomy: State of the art, challenges and opportunities. Bioresour. Technol. 2021, 330, 124997. [Google Scholar] [CrossRef] [PubMed]
- Omarini, A.; Dambolena, J.S.; Lucini, E.; Jaramillo Mejía, S.; Albertó, E.; Zygadlo, J.A. Biotransformation of 1,8-cineole by solid-state fermentation of Eucalyptus waste from the essential oil industry using Pleurotus ostreatus and Favolus tenuiculus. Folia Microbiol. 2016, 61, 149–157. [Google Scholar] [CrossRef]
- Behrens, C.J.; Zelena, K.; Berger, R.G. Comparative Cold Shock Expression and Characterization of Fungal Dye-Decolorizing Peroxidases. Appl. Biochem. Biotechnol. 2016, 179, 1404–1417. [Google Scholar] [CrossRef]
- Zhou, J.; Bai, Y.; Dai, R.; Guo, X.; Liu, Z.H.; Yuan, S. Improved Polysaccharide Production by Homologous Co-overexpression of Phosphoglucomutase and UDP Glucose Pyrophosphorylase Genes in the Mushroom Coprinopsis cinerea. J. Agric. Food Chem. 2018, 66, 4702–4709. [Google Scholar] [CrossRef]
- Zelena, K.; Eisele, N.; Berger, R.G. Escherichia coli as a production host for novel enzymes from basidiomycota. Biotechnol. Adv. 2014, 32, 1382–1395. [Google Scholar] [CrossRef]
- Behrens, C.J.; Linke, D.; Allister, A.B.; Zelena, K.; Berger, R.G. Variants of PpuLcc, a multi-dye decolorizing laccase from Pleurotus pulmonarius expressed in Pichia pastoris. Protein Expr. Purif. 2017, 137, 34–42. [Google Scholar] [CrossRef]
- Wang, L.; Huang, W.; Shen, Y.; Zhao, Y.; Wu, D.; Yin, H.; Yang, S.; Yuan, Q.; Liang, W.; Wang, J. Enhancing the degradation of Aflatoxin B(1) by co-cultivation of two fungi strains with the improved production of detoxifying enzymes. Food Chem. 2022, 371, 131092. [Google Scholar] [CrossRef]
- Xu, X.; Qu, R.; Wu, W.; Jiang, C.; Shao, D.; Shi, J. Applications of microbial co-cultures in polyketides production. J. Appl. Microbiol. 2021, 130, 1023–1034. [Google Scholar] [CrossRef]
- Oda, S. Microbial transformation of water-insoluble substrates by two types of novel interface bioprocesses, tacky liquid-liquid interface bioreactor and non-aqueous sporular bioconversion system. World J. Microbiol. Biotechnol. 2020, 36, 57. [Google Scholar] [CrossRef] [PubMed]
- Kupčík, R.; Zelená, M.; Řehulka, P.; Bílková, Z.; Česlová, L. Selective isolation of hydrophobin SC3 by solid-phase extraction with polytetrafluoroethylene microparticles and subsequent mass spectrometric analysis. J. Sep. Sci. 2016, 39, 717–724. [Google Scholar] [CrossRef] [PubMed]
- Tao, T.L.; Cui, F.J.; Chen, X.X.; Sun, W.J.; Huang, D.M.; Zhang, J.; Yang, Y.; Wu, D.; Liu, W.M. Improved mycelia and polysaccharide production of Grifola frondosa by controlling morphology with microparticle Talc. Microb. Cell Fact. 2018, 17, 1. [Google Scholar] [CrossRef] [PubMed]
- Karahalil, E.; Coban, H.B.; Turhan, I. A current approach to the control of filamentous fungal growth in media: Microparticle enhanced cultivation technique. Crit. Rev. Biotechnol. 2019, 39, 192–201. [Google Scholar] [CrossRef]
- Ramírez, D.A.; Muñoz, S.V.; Atehortua, L.; Michel, F.C., Jr. Effects of different wavelengths of light on lignin peroxidase production by the white-rot fungi Phanerochaete chrysosporium grown in submerged cultures. Bioresour. Technol. 2010, 101, 9213–9220. [Google Scholar] [CrossRef]
- Shim, D.; Park, S.G.; Kim, K.; Bae, W.; Lee, G.W.; Ha, B.S.; Ro, H.S.; Kim, M.; Ryoo, R.; Rhee, S.K.; et al. Whole genome de novo sequencing and genome annotation of the world popular cultivated edible mushroom, Lentinula edodes. J. Biotechnol. 2016, 223, 24–25. [Google Scholar] [CrossRef]
- Zhang, Z.; Shah, A.M.; Mohamed, H.; Zhang, Y.; Tsiklauri, N.; Song, Y. Genomic Studies of White-Rot Fungus Cerrena unicolor SP02 Provide Insights into Food Safety Value-Added Utilization of Non-Food Lignocellulosic Biomass. J. Fungi 2021, 7, 835. [Google Scholar] [CrossRef]
- Luo, H.; Qian, J.; Xu, Z.; Liu, W.; Xu, L.; Li, Y.; Xu, J.; Zhang, J.; Xu, X.; Liu, C.; et al. The Wolfiporia cocos Genome and Transcriptome Shed Light on the Formation of Its Edible and Medicinal Sclerotium. Genom. Proteom. Bioinform. 2020, 18, 455–467. [Google Scholar] [CrossRef]
- Dilokpimol, A.; Mäkelä, M.R.; Varriale, S.; Zhou, M.; Cerullo, G.; Gidijala, L.; Hinkka, H.; Brás, J.L.A.; Jütten, P.; Piechot, A.; et al. Fungal feruloyl esterases: Functional validation of genome mining based enzyme discovery including uncharacterized subfamilies. N. Biotechnol. 2018, 41, 9–14. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Chen, X.; Lee, R.T.C.; Maurer-Stroh, S.; Rühl, M. Bioinformatics-aided identification, characterization and applications of mushroom linalool synthases. Commun. Biol. 2021, 4, 223. [Google Scholar] [CrossRef]
- Lin, Y.L.; Ma, L.T.; Lee, Y.R.; Shaw, J.F.; Wang, S.Y.; Chu, F.H. Differential Gene Expression Network in Terpenoid Synthesis of Antrodia cinnamomea in Mycelia and Fruiting Bodies. J. Agric. Food Chem. 2017, 65, 1874–1886. [Google Scholar] [CrossRef] [PubMed]
- Peng, M.; Aguilar-Pontes, M.V.; Hainaut, M.; Henrissat, B.; Hildén, K.; Mäkelä, M.R.; de Vries, R.P. Comparative analysis of basidiomycete transcriptomes reveals a core set of expressed genes encoding plant biomass degrading enzymes. Fungal Genet. Biol. 2018, 112, 40–46. [Google Scholar] [CrossRef] [PubMed]
- Orban, A.; Weber, A.; Herzog, R.; Hennicke, F.; Rühl, M. Transcriptome of different fruiting stages in the cultivated mushroom Cyclocybe aegerita suggests a complex regulation of fruiting and reveals enzymes putatively involved in fungal oxylipin biosynthesis. BMC Genom. 2021, 22, 324. [Google Scholar] [CrossRef] [PubMed]
- Krahe, N.-K.; Berger, R.G.; Witt, M.; Zorn, H.; Omarini, A.B.; Ersoy, F. Monokaryotic Pleurotus sapidus Strains with Intraspecific Variability of an Alkene Cleaving DyP-Type Peroxidase Activity as a Result of Gene Mutation and Differential Gene Expression. Int. J. Mol. Sci. 2021, 22, 1363. [Google Scholar] [CrossRef] [PubMed]
- Linke, D.; Omarini, A.B.; Takenberg, M.; Kelle, S.; Berger, R.G. Long-Term Monokaryotic Cultures of Pleurotus ostreatus var. florida Produce High and Stable Laccase Activity Capable to Degrade ss-Carotene. Appl. Biochem. Biotechnol. 2019, 187, 894–912. [Google Scholar] [CrossRef]
- Mehta, A.; Guleria, S.; Sharma, R.; Gupta, R. The lipases and their applications with emphasis on food industry. In Microbial Biotechnology in Food and Health; Ray, R.C., Ed.; Academic Press: Cambridge, MA, USA, 2021; pp. 143–164. [Google Scholar]
- Rehman, S.; Bhatti, H.N.; Bilal, M.; Asgher, M. Optimization of process variables for enhanced production of extracellular lipase by Pleurotus ostreatus IBL-02 in solid-state fermentation. Pak. J. Pharm. Sci. 2019, 32, 617–624. [Google Scholar]
- Lan, D.; Popowicz, G.M.; Pavlidis, I.V.; Zhou, P.; Bornscheuer, U.T.; Wang, Y. Conversion of a Mono- and Diacylglycerol Lipase into a Triacylglycerol Lipase by Protein Engineering. Chembiochem 2015, 16, 1431–1434. [Google Scholar] [CrossRef]
- Nieter, A.; Haase-Aschoff, P.; Kelle, S.; Linke, D.; Krings, U.; Popper, L.; Berger, R.G. A chlorogenic acid esterase with a unique substrate specificity from Ustilago maydis. Appl. Environ. Microbiol. 2015, 81, 1679–1688. [Google Scholar] [CrossRef] [Green Version]
- Nieter, A.; Kelle, S.; Takenberg, M.; Linke, D.; Bunzel, M.; Popper, L.; Berger, R.G. Heterologous production and characterization of a chlorogenic acid esterase from Ustilago maydis with a potential use in baking. Food Chem. 2016, 209, 1–9. [Google Scholar] [CrossRef]
- Nieter, A.; Kelle, S.; Linke, D.; Berger, R.G. Feruloyl esterases from Schizophyllum commune to treat food industry side-streams. Bioresour. Technol. 2016, 220, 38–46. [Google Scholar] [CrossRef]
- Siebert, M.; Krings, U.; Gunther, T.; Fragalas, A.; Berger, R.G. Enzymatic hydrolysis of kaempferol 3-O-(2″″-O-sinapoyl-beta-sophoroside), the key bitter compound of rapeseed (Brassica napus L.) protein isolate. J. Sci. Food Agric. 2021, 102, 2179–2182. [Google Scholar] [CrossRef] [PubMed]
- Siebert, M.; Berger, R.G.; Nieter, A. Enzymatic mitigation of 5-O-chlorogenic acid for an improved digestibility of coffee. Food Chem. 2018, 258, 124–128. [Google Scholar] [CrossRef] [PubMed]
- Siebert, M.; Berger, R.G.; Pfeiffer, F. Hydrolysis of chlorogenic acid in apple juice using a p-coumaryl esterase of Rhizoctonia solani. J. Sci. Food Agric. 2019, 99, 6644–6648. [Google Scholar] [CrossRef] [Green Version]
- Esparan, V.; Krings, U.; Struch, M.; Berger, R.G. A three-enzyme-system to degrade curcumin to natural vanillin. Molecules 2015, 20, 6640–6653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krahe, N.K.; Berger, R.G.; Kahlert, L.; Ersoy, F. Co-Oxidative Transformation of Piperine to Piperonal and 3,4-Methylenedioxycinnamaldehyde by a Lipoxygenase from Pleurotus sapidus. Chembiochem 2021, 22, 2857–2861. [Google Scholar] [CrossRef]
- Spier, M.R.; Behsnilian, D.; Zielinski, A.; Konietzny, U.; Greiner, R. Studies towards the stabilisation of a mushroom phytase produced by submerged cultivation. Protein J. 2015, 34, 367–379. [Google Scholar] [CrossRef] [PubMed]
- Spier, M.R.; Siepmann, F.B.; Staack, L.; Souza, P.Z.; Kumar, V.; Medeiros, A.B.; Soccol, C.R. Impact of microbial growth inhibition and proteolytic activity on the stability of a new formulation containing a phytate-degrading enzyme obtained from mushroom. Prep. Biochem. Biotechnol. 2016, 46, 725–733. [Google Scholar] [CrossRef] [PubMed]
- Tatsumi, E.; Konishi, Y.; Tsujiyama, S. Application of residual polysaccharide-degrading enzymes in dried shiitake mushrooms as an enzyme preparation in food processing. Biotechnol. Lett. 2016, 38, 1923–1928. [Google Scholar] [CrossRef]
- Guo, Y.; Song, Y.; Qiu, Y.; Shao, X.; Wang, H.; Song, Y. Purification of thermostable α-galactosidase from Irpex lacteus and its use for hydrolysis of oligosaccharides. J. Basic Microbiol. 2016, 56, 448–458. [Google Scholar] [CrossRef]
- Jang, J.M.; Yang, Y.; Wang, R.; Bao, H.; Yuan, H.; Yang, J. Characterization of a high performance α-galactosidase from Irpex lacteus and its usage in removal of raffinose family oligosaccharides from soymilk. Int. J. Biol. Macromol. 2019, 131, 1138–1146. [Google Scholar] [CrossRef]
- Geng, X.; Fan, J.; Xu, L.; Wang, H.; Ng, T.B. Hydrolysis of oligosaccharides by a fungal α-galactosidase from fruiting bodies of a wild mushroom Leucopaxillus tricolor. J. Basic Microbiol. 2018, 58, 1043–1052. [Google Scholar] [CrossRef] [PubMed]
- Geng, X.; Yang, D.; Zhang, Q.; Chang, M.; Xu, L.; Cheng, Y.; Wang, H.; Meng, J. Good hydrolysis activity on raffinose family oligosaccharides by a novel α-galactosidase from Tremella aurantialba. Int. J. Biol. Macromol. 2020, 150, 1249–1257. [Google Scholar] [CrossRef] [PubMed]
- Omrane Benmrad, M.; Mechri, S.; Zaraî Jaouadi, N.; Ben Elhoul, M.; Rekik, H.; Sayadi, S.; Bejar, S.; Kechaou, N.; Jaouadi, B. Purification and biochemical characterization of a novel thermostable protease from the oyster mushroom Pleurotus sajor-caju strain CTM10057 with industrial interest. BMC Biotechnol. 2019, 19, 43. [Google Scholar] [CrossRef] [PubMed]
- da Silva, R.R.; de Oliveira, L.C.; Juliano, M.A.; Juliano, L.; de Oliveira, A.H.; Rosa, J.C.; Cabral, H. Biochemical and milk-clotting properties and mapping of catalytic subsites of an extracellular aspartic peptidase from basidiomycete fungus Phanerochaete chrysosporium. Food Chem. 2017, 225, 45–54. [Google Scholar] [CrossRef] [PubMed]
- Majumder, R.; Banik, S.P.; Khowala, S. Purification and characterisation of κ-casein specific milk-clotting metalloprotease from Termitomyces clypeatus MTCC 5091. Food Chem. 2015, 173, 441–448. [Google Scholar] [CrossRef] [PubMed]
- Schulz, K.; Giesler, L.; Linke, D.; Berger, R.G. A prolyl endopeptidase from Flammulina velutipes for the possible degradation of celiac disease provoking toxic peptides in cereal proteins. Process Biochem. 2018, 73, 47–55. [Google Scholar] [CrossRef]
- Lee, K.H.; Kim, H.K.; Kim, S.H.; Kim, K.H.; Choi, Y.M.; Jin, H.H.; Lee, S.J.; Ryu, Y.C. Effects of mushroom extract on textural properties and muscle protein degradation of bovine longissimus dorsi muscle. Biosci. Biotechnol. Biochem. 2017, 81, 558–564. [Google Scholar] [CrossRef] [Green Version]
- Biko, O.D.V.; Viljoen-Bloom, M.; van Zyl, W.H. Microbial lignin peroxidases: Applications, production challenges and future perspectives. Enzyme Microb. Technol. 2020, 141, 109669. [Google Scholar] [CrossRef]
- Knop, D.; Yarden, O.; Hadar, Y. The ligninolytic peroxidases in the genus Pleurotus: Divergence in activities, expression, and potential applications. Appl. Microbiol. Biotechnol. 2015, 99, 1025–1038. [Google Scholar] [CrossRef]
- Imami, A.; Riemer, S.; Schulze, M.; Amelung, F.; Gorshkov, V.; Rühl, M.; Ammenn, J.; Zorn, H. Depolymerization of lignosulfonates by submerged cultures of the basidiomycete Irpex consors and cloning of a putative versatile peroxidase. Enzyme Microb. Technol. 2015, 81, 8–15. [Google Scholar] [CrossRef]
- Upadhyay, P.; Shrivastava, R.; Agrawal, P.K. Bioprospecting and biotechnological applications of fungal laccase. 3 Biotech 2016, 6, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patel, N.; Shahane, S.; Shivam; Majumdar, R.; Mishra, U. Mode of Action, Properties, Production, and Application of Laccase: A Review. Recent Pat. Biotechnol. 2019, 13, 19–32. [Google Scholar] [CrossRef] [PubMed]
- Martínková, L.; Kotik, M.; Marková, E.; Homolka, L. Biodegradation of phenolic compounds by Basidiomycota and its phenol oxidases: A review. Chemosphere 2016, 149, 373–382. [Google Scholar] [CrossRef] [PubMed]
- Taghizadeh, T.; Talebian-Kiakalaieh, A.; Jahandar, H.; Amin, M.; Tarighi, S.; Faramarzi, M.A. Biodegradation of bisphenol A by the immobilized laccase on some synthesized and modified forms of zeolite Y. J. Hazard. Mater. 2020, 386, 121950. [Google Scholar] [CrossRef]
- Mayans, B.; Camacho-Arévalo, R.; García-Delgado, C.; Antón-Herrero, R.; Escolástico, C.; Segura, M.L.; Eymar, E. An assessment of Pleurotus ostreatus to remove sulfonamides, and its role as a biofilter based on its own spent mushroom substrate. Environ Sci. Pollut. Res. Int. 2021, 28, 7032–7042. [Google Scholar] [CrossRef]
- García-Delgado, C.; Yunta, F.; Eymar, E. Bioremediation of multi-polluted soil by spent mushroom (Agaricus bisporus) substrate: Polycyclic aromatic hydrocarbons degradation and Pb availability. J. Hazard. Mater. 2015, 300, 281–288. [Google Scholar] [CrossRef]
- Zhang, C.; Li, M.; Chen, X.; Li, M. Edible fungus degrade bisphenol A with no harmful effect on its fatty acid composition. Ecotoxicol. Environ. Saf. 2015, 118, 126–132. [Google Scholar] [CrossRef]
- Zhou, Z.; Li, R.; Ng, T.B.; Lai, Y.; Yang, J.; Ye, X. A New Laccase of Lac 2 from the White Rot Fungus Cerrena unicolor 6884 and Lac 2-Mediated Degradation of Aflatoxin B(1). Toxins 2020, 12, 476. [Google Scholar] [CrossRef]
- Branà, M.T.; Sergio, L.; Haidukowski, M.; Logrieco, A.F.; Altomare, C. Degradation of Aflatoxin B(1) by a Sustainable Enzymatic Extract from Spent Mushroom Substrate of Pleurotus eryngii. Toxins 2020, 12, 49. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Mao, H.; Woldemariam Yohannes, K.; Wan, Z.; Cao, Y.; Tron, T.; Lin, J.; Jiang, Y.; Li, H.; Wang, J. Degradation of aflatoxin B(1) by a recombinant laccase from Trametes sp. C30 expressed in Saccharomyces cerevisiae: A mechanism assessment study in vitro and in vivo. Food Res. Int. 2021, 145, 110418. [Google Scholar] [CrossRef]
- Yang, P.; Xiao, W.; Lu, S.; Jiang, S.; Zheng, Z.; Zhang, D.; Zhang, M.; Jiang, S.; Jiang, S. Recombinant Expression of Trametes versicolor Aflatoxin B(1)-Degrading Enzyme (TV-AFB(1)D) in Engineering Pichia pastoris GS115 and Application in AFB(1) Degradation in AFB(1)-Contaminated Peanuts. Toxins 2021, 13, 349. [Google Scholar] [CrossRef] [PubMed]
- Tso, K.H.; Lumsangkul, C.; Ju, J.C.; Fan, Y.K.; Chiang, H.I. The Potential of Peroxidases Extracted from the Spent Mushroom (Flammulina velutipes) Substrate Significantly Degrade Mycotoxin Deoxynivalenol. Toxins 2021, 13, 72. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Bai, Y.; Huang, H.; Tu, T.; Wang, Y.; Wang, Y.; Luo, H.; Yao, B.; Su, X. Degradation of Aflatoxin B(1) and Zearalenone by Bacterial and Fungal Laccases in Presence of Structurally Defined Chemicals and Complex Natural Mediators. Toxins 2019, 11, 609. [Google Scholar] [CrossRef] [Green Version]
- Khalighi, S.; Berger, R.G.; Ersoy, F. Cross-linking of fibrex gel by fungal laccase: Gel rheological and structural characteristics. Processes 2020, 8, 16. [Google Scholar] [CrossRef] [Green Version]
- Khalighi, S.; Berger, R.G.; Ersoy, F. Cross-linking of wheat bran arabinoxylan by fungal laccases yields firm gels. Processes 2020, 8, 36. [Google Scholar] [CrossRef] [Green Version]
- Kelle, S.; Nieter, A.; Krings, U.; Zelena, K.; Linke, D.; Berger, R.G. Heterologous production of a feruloyl esterase from Pleurotus sapidus synthesizing feruloyl-saccharide esters. Biotechnol. Appl. Biochem. 2016, 63, 852–862. [Google Scholar] [CrossRef]
- Tian, Q.; Feng, Y.; Huang, H.; Zhang, J.; Yu, Y.; Guan, Z.; Cai, Y.; Liao, X. Production of lactobionic acid from lactose using the cellobiose dehydrogenase-3-HAA-laccase system from Pycnoporus sp. SYBC-L10. Lett. Appl. Microbiol. 2018, 67, 589–597. [Google Scholar] [CrossRef]
- Szweda, R.T.; Schmidt, K.; Zorn, H. Bleaching of colored whey and milk by a multiple-enzyme system. Eur. Food Res. Technol. 2013, 237, 377–384. [Google Scholar] [CrossRef]
- Kahraman, S.; Yeşilada, O. Decolorization and bioremediation of molasses wastewater by white-rot fungi in a semi-solid-state condition. Folia Microbiol. 2003, 48, 525–528. [Google Scholar] [CrossRef]
- Rodríguez Couto, S.; Toca Herrera, J.L. Industrial and biotechnological applications of laccases: A review. Biotechnol. Adv. 2006, 24, 500–513. [Google Scholar] [CrossRef]
- Meruvu, H.; Dos Santos, J.C. Colors of life: A review on fungal pigments. Crit. Rev. Biotechnol. 2021, 41, 1153–1177. [Google Scholar] [CrossRef] [PubMed]
- Zschätzsch, M.; Steudler, S.; Reinhardt, O.; Bergmann, P.; Ersoy, F.; Stange, S.; Wagenführ, A.; Walther, T.; Berger, R.G.; Werner, A. Production of natural colorants by liquid fermentation with Chlorociboria aeruginascens and Laetiporus sulphureus and prospective applications. Eng. Life Sci. 2021, 21, 270–282. [Google Scholar] [CrossRef] [PubMed]
- Verloop, A.J.; Gruppen, H.; Bisschop, R.; Vincken, J.P. Altering the phenolics profile of a green tea leaves extract using exogenous oxidases. Food Chem. 2016, 196, 1197–1206. [Google Scholar] [CrossRef] [PubMed]
- Narai-Kanayama, A.; Uekusa, Y.; Kiuchi, F.; Nakayama, T. Efficient Synthesis of Theaflavin 3-Gallate by a Tyrosinase-Catalyzed Reaction with (-)-Epicatechin and (-)-Epigallocatechin Gallate in a 1-Octanol/Buffer Biphasic System. J. Agric. Food Chem. 2018, 66, 13464–13472. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.; Jia, G.; Sun, H.; Sun, T.; Hou, D. Genome sequence of the fungus Pycnoporus sanguineus, which produces cinnabarinic acid and pH- and thermo-stable laccases. Gene 2020, 742, 144586. [Google Scholar] [CrossRef] [PubMed]
- Marim, R.A.; Oliveira, A.C.; Marquezoni, R.S.; Servantes, J.P.; Cardoso, B.K.; Linde, G.A.; Colauto, N.B.; Valle, J.S. Use of sugarcane molasses by Pycnoporus sanguineus for the production of laccase for dye decolorization. Genet Mol. Res. 2016, 15, gmr15048972. [Google Scholar] [CrossRef] [PubMed]
- Detering, T.; Berger, R.G. Formation of 2,6-DMBQ in anthocyanin-rich foods. LWT 2020, 134, 110178. [Google Scholar] [CrossRef]
- Behrens, C.J.; Krahe, N.K.; Linke, D.; Berger, R.G. BadGluc, a beta-glucosidase from Bjerkandera adusta with anthocyanase properties. Bioprocess Biosyst. Eng. 2018, 41, 1391–1401. [Google Scholar] [CrossRef]
- Knop, D.; Levinson, D.; Makovitzki, A.; Agami, A.; Lerer, E.; Mimran, A.; Yarden, O.; Hadar, Y. Limits of Versatility of Versatile Peroxidase. Appl. Environ. Microbiol. 2016, 82, 4070–4080. [Google Scholar] [CrossRef] [Green Version]
- Korniłłowicz-Kowalska, T.; Rybczyńska-Tkaczyk, K. Decolorization and biodegradation of melanoidin contained in beet molasses by an anamorphic strain of Bjerkandera adusta CCBAS930 and its mutants. World J. Microbiol. Biotechnol. 2020, 37, 1. [Google Scholar] [CrossRef]
- Urlacher, V.B.; Koschorreck, K. Pecularities and applications of aryl-alcohol oxidases from fungi. Appl. Microbiol. Biotechnol. 2021, 105, 4111–4126. [Google Scholar] [CrossRef] [PubMed]
- Serrano, A.; Carro, J.; Martínez, A.T. Reaction mechanisms and applications of aryl-alcohol oxidase. Enzymes 2020, 47, 167–192. [Google Scholar] [CrossRef] [PubMed]
- Viña-Gonzalez, J.; Martinez, A.T.; Guallar, V.; Alcalde, M. Sequential oxidation of 5-hydroxymethylfurfural to furan-2,5-dicarboxylic acid by an evolved aryl-alcohol oxidase. Biochim. Biophys. Acta Proteins Proteom. 2020, 1868, 140293. [Google Scholar] [CrossRef] [PubMed]
- Galperin, I.; Javeed, A.; Luig, H.; Lochnit, G.; Rühl, M. An aryl-alcohol oxidase of Pleurotus sapidus: Heterologous expression, characterization, and application in a 2-enzyme system. Appl. Microbiol. Biotechnol. 2016, 100, 8021–8030. [Google Scholar] [CrossRef]
- Wang, Q.; Wang, F.; Xu, Z.; Ding, Z. Bioactive Mushroom Polysaccharides: A Review on Monosaccharide Composition, Biosynthesis and Regulation. Molecules 2017, 22, 955. [Google Scholar] [CrossRef] [Green Version]
- Castañeda, M.T.; Adachi, O.; Hours, R.A. Reduction of L-phenylalanine in protein hydrolysates using L-phenylalanine ammonia-lyase from Rhodosporidium toruloides. J. Ind. Microbiol. Biotechnol. 2015, 42, 1299–1307. [Google Scholar] [CrossRef]
- Chakraborty, M.; Shivakumar, S. Bioprospecting of the agaricomycete Ganoderma australe GPC191 as novel source for L-asparaginase production. Sci. Rep. 2021, 11, 6192. [Google Scholar] [CrossRef]
- Bel-Rhlid, R.; Berger, R.G.; Blank, I. Bio-mediated generation of food flavors—Towards sustainable flavor production inspired by nature. Trends Food Sci. Technol. 2018, 78, 134–143. [Google Scholar] [CrossRef]
- Krahe, N.K.; Berger, R.G.; Ersoy, F. A DyP-type peroxidase of pleurotus sapidus with alkene cleaving activity. Molecules 2020, 25, 1536. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Wang, J.; Chen, W.; Yang, Y.; Zhang, J.; Feng, J.; Yu, H.; Li, Q. Analysis of volatile compounds of Lentinula edodes grown in different culture substrate formulations. Food Res. Int. 2019, 125, 108517. [Google Scholar] [CrossRef]
- Wang, Y.; Yang, Z.; Chen, X.; Han, D.; Han, J.; Wang, L.; Ren, A.; Yu, H.; Zhao, M. Lenthionine, a Key Flavor Substance in Lentinula edodes, Is Regulated by Cysteine under Drought Stress. J. Agric. Food Chem. 2021, 69, 12645–12653. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Chen, W.C.; Wang, J.B.; Feng, J.; Wu, D.; Zhang, Z.; Zhang, J.S.; Yang, Y. Screening candidate genes related to volatile synthesis in shiitake mushrooms and construction of regulatory networks to effectively improve mushroom aroma. J. Sci. Food Agric. 2021, 101, 5618–5626. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Chen, X.; Orban, A.; Shukal, S.; Birk, F.; Too, H.P.; Rühl, M. Agrocybe aegerita Serves As a Gateway for Identifying Sesquiterpene Biosynthetic Enzymes in Higher Fungi. ACS Chem. Biol. 2020, 15, 1268–1277. [Google Scholar] [CrossRef] [PubMed]
- Mischko, W.; Hirte, M.; Fuchs, M.; Mehlmer, N.; Brück, T.B. Identification of sesquiterpene synthases from the Basidiomycota Coniophora puteana for the efficient and highly selective β-copaene and cubebol production in E. coli. Microb. Cell Fact. 2018, 17, 164. [Google Scholar] [CrossRef]
- Püth, N.; Ersoy, F.; Krings, U.; Berger, R.G. Sesquiterpene Cyclases from the Basidiomycete Cerrena unicolor. Catalysts 2021, 11, 1361. [Google Scholar] [CrossRef]
- Grosse, M.; Strauss, E.; Krings, U.; Berger, R.G. Response of the sesquiterpene synthesis in submerged cultures of the Basidiomycete Tyromyces floriformis to the medium composition. Mycologia 2019, 111, 885–894. [Google Scholar] [CrossRef] [PubMed]
- Kolwek, J.; Behrens, C.; Linke, D.; Krings, U.; Berger, R.G. Cell-free one-pot conversion of (+)-valencene to (+)-nootkatone by a unique dye-decolorizing peroxidase combined with a laccase from Funalia trogii. J. Ind. Microbiol. Biotechnol. 2018, 45, 89–101. [Google Scholar] [CrossRef]
- Grosse, M.; Pendzialeck, T.; Fohrer, J.; Berger, R.G.; Krings, U. (5E/Z,7E,9)-Decatrien-2-ones, Pineapple-like Flavors from Fomitopsis betulina-Structure Elucidation and Sensorial Properties. J. Agric. Food Chem. 2020, 68, 10329–10335. [Google Scholar] [CrossRef]
- Grosse, M.; Wu, S.; Krings, U.; Berger, R.G. Formation of Decatrienones with a Pineapple-like Aroma from 1-(13)C-Acetate by Cell Cultures of the Birch Polypore, Fomitopsis betulina. J. Agric. Food Chem. 2020, 68, 1678–1683. [Google Scholar] [CrossRef]
- Hammer, A.K.; Emrich, N.O.; Ott, J.; Birk, F.; Fraatz, M.A.; Ley, J.P.; Geissler, T.; Bornscheuer, U.T.; Zorn, H. Biotechnological Production and Sensory Evaluation of ω1-Unsaturated Aldehydes. J. Agric. Food Chem. 2021, 69, 345–353. [Google Scholar] [CrossRef]
- Günther, T.; Schoppe, L.; Ersoy, F.; Berger, R.G. Alternative Routes for the Production of Natural 4-Vinylguaiacol from Sugar Beet Fiber Using Basidiomycetous Enzymes. Catalysts 2021, 11, 631. [Google Scholar] [CrossRef]
- Detering, T.; Mundry, K.; Berger, R.G. Generation of 4-vinylguaiacol through a novel high-affinity ferulic acid decarboxylase to obtain smoke flavours without carcinogenic contaminants. PLoS ONE 2020, 15, e0244290. [Google Scholar] [CrossRef] [PubMed]
- Hadj Saadoun, J.; Bertani, G.; Levante, A.; Vezzosi, F.; Ricci, A.; Bernini, V.; Lazzi, C. Fermentation of Agri-Food Waste: A Promising Route for the Production of Aroma Compounds. Foods 2021, 10, 707. [Google Scholar] [CrossRef] [PubMed]
- Burger, F.; Koch, M.; Fraatz, M.A.; Omarini, A.B.; Berger, R.G.; Zorn, H. Production of an Anise- and Woodruff-like Aroma by Monokaryotic Strains of Pleurotus sapidus Grown on Citrus Side Streams. Molecules 2022, 27, 651. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Venkitasamy, C.; Pan, Z.; Liu, W.; Zhao, L. Novel Umami Ingredients: Umami Peptides and Their Taste. J. Food Sci. 2017, 82, 16–23. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Zhang, X.; Xin, G.; Gong, X.; Wang, Y.; Wang, L.; Sun, B. Umami taste and its association with energy status in harvested Pleurotus geesteranus stored at different temperatures. Food Chem. 2019, 279, 179–186. [Google Scholar] [CrossRef] [PubMed]
- Kong, Y.; Zhang, L.L.; Zhao, J.; Zhang, Y.Y.; Sun, B.G.; Chen, H.T. Isolation and identification of the umami peptides from shiitake mushroom by consecutive chromatography and LC-Q-TOF-MS. Food Res. Int. 2019, 121, 463–470. [Google Scholar] [CrossRef]
- Poojary, M.M.; Orlien, V.; Passamonti, P.; Olsen, K. Enzyme-assisted extraction enhancing the umami taste amino acids recovery from several cultivated mushrooms. Food Chem. 2017, 234, 236–244. [Google Scholar] [CrossRef]
- Zhou, J.; Chen, M.; Wu, S.; Liao, X.; Wang, J.; Wu, Q.; Zhuang, M.; Ding, Y. A review on mushroom-derived bioactive peptides: Preparation and biological activities. Food Res. Int. 2020, 134, 109230. [Google Scholar] [CrossRef]
- Harth, L.; Krah, U.; Linke, D.; Dunkel, A.; Hofmann, T.; Berger, R.G. Salt Taste Enhancing l-Arginyl Dipeptides from Casein and Lysozyme Released by Peptidases of Basidiomycota. J. Agric. Food Chem. 2018, 66, 2344–2353. [Google Scholar] [CrossRef]
- Tamrakar, S.; Fukami, K.; Parajuli, G.P.; Shimizu, K. Antiallergic Activity of the Wild Mushrooms of Nepal and the Pure Compound Hispidin. J. Med. Food 2019, 22, 225–227. [Google Scholar] [CrossRef] [PubMed]
- Kawai, J.; Mori, K.; Hirasawa, N. Grifola frondosa extract and ergosterol reduce allergic reactions in an allergy mouse model by suppressing the degranulation of mast cells. Biosci. Biotechnol. Biochem. 2019, 83, 2280–2287. [Google Scholar] [CrossRef] [PubMed]
- Kang, L.; Li, Q.; Lin, J.; Guo, L. Biosynthesis of Resveratrol in Blastospore of the Macrofungus Tremella fuciformis. Mol. Biotechnol. 2015, 57, 675–684. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Zhao, H.; Gao, Z.; Song, X.; Wang, W.; Yuan, F.; Feng, Y.; Zhang, Y.; Zhang, J.; Zhang, S.; et al. The Antioxidant and Anti-Aging Effects of Acetylated Mycelia Polysaccharides from Pleurotus djamor. Molecules 2019, 24, 2698. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Jiao, Y.; Lu, H.; Shu, X.; Chen, Q. Chemical characterization, antioxidant properties and anticancer activity of exopolysaccharides from Floccularia luteovirens. Carbohydr. Polym. 2020, 229, 115432. [Google Scholar] [CrossRef]
- Omarini, A.B.; Plagemann, I.; Schimanski, S.; Krings, U.; Berger, R.G. Crosses between monokaryons of Pleurotus sapidus or Pleurotus florida show an improved biotransformation of (+)-valencene to (+)-nootkatone. Bioresour. Technol. 2014, 171, 113–119. [Google Scholar] [CrossRef]
- Gandia, A.; van den Brandhof, J.G.; Appels, F.V.W.; Jones, M.P. Flexible Fungal Materials: Shaping the Future. Trends Biotechnol. 2021, 39, 1321–1331. [Google Scholar] [CrossRef]
- Grimm, D.; Wösten, H.A.B. Mushroom cultivation in the circular economy. Appl. Microbiol. Biotechnol. 2018, 102, 7795–7803. [Google Scholar] [CrossRef] [Green Version]
- Roth-Johnson, L. Dreams of a Fungal Future. Available online: https://www.kqed.org/quest/71171/a-house-made-from-mushrooms-an-artist-dreams-of-a-fungal-future (accessed on 14 March 2022).
- Scheuerle, S.; Landgraf, M. Cultivated Building Materials for Cities of the Future. Available online: https://www.kit.edu/kit/english/pi_2017_121_cultivated-building-materials-for-cities-of-the-future.php (accessed on 14 March 2022).
- Frearson, A. Tower of “Grown” Bio-Bricks by the Living Opens at MoMA PS1. Available online: https://www.dezeen.com/2014/07/01/tower-of-grown-bio-bricks-by-the-living-opens-at-moma-ps1-gallery/ (accessed on 14 March 2022).
Figure 1.
(R)-(-)-linalool (Licareol), a potent flavour with flowery-wood odour notes.

Figure 2.
Structural models of PsaPOX variants generated with SWISS-MODEL using the X-ray structure of Pleos-DyP4 (PDB-ID 6FSK). (a) Overall fold of PsaPOX_DK showing the typical ferredoxin-like fold, with the heme cofactor in yellow and the heme iron in orange. Conserved amino acids of the active site/hydrogen peroxide pocket (proximal H334, distal D196, R360, L385 and F387) are shown in cyan. D433, which forms a hydrogen bond with H334 to stabilise compound I (oxidised heme after transfer of two electrons to H2O2), is presented in blue, and the exposed W405 potentially involved in long-range electron transfer is shown in green. Important amino acids for Mn2+-oxidation (D215, Y339, E345, D352 and D354) are highlighted in magenta. (b) PsaPOX_high with a single-point mutant (S371H) in comparison to PsaPOX_DK and a ca. 1.5-fold increased activity. Reprinted from [35].
Figure 2.
Structural models of PsaPOX variants generated with SWISS-MODEL using the X-ray structure of Pleos-DyP4 (PDB-ID 6FSK). (a) Overall fold of PsaPOX_DK showing the typical ferredoxin-like fold, with the heme cofactor in yellow and the heme iron in orange. Conserved amino acids of the active site/hydrogen peroxide pocket (proximal H334, distal D196, R360, L385 and F387) are shown in cyan. D433, which forms a hydrogen bond with H334 to stabilise compound I (oxidised heme after transfer of two electrons to H2O2), is presented in blue, and the exposed W405 potentially involved in long-range electron transfer is shown in green. Important amino acids for Mn2+-oxidation (D215, Y339, E345, D352 and D354) are highlighted in magenta. (b) PsaPOX_high with a single-point mutant (S371H) in comparison to PsaPOX_DK and a ca. 1.5-fold increased activity. Reprinted from [35].

Scheme 1.
(a) Ferulic acid, liberated from lignified plant materials, is not only a powerful antioxidant, but also a precursor to other valuable compounds. (b) p-Coumaric acid may likewise be transformed into other bioactive compounds.
Scheme 1.
(a) Ferulic acid, liberated from lignified plant materials, is not only a powerful antioxidant, but also a precursor to other valuable compounds. (b) p-Coumaric acid may likewise be transformed into other bioactive compounds.

Scheme 2.
Biotransformations using enzymes from basidiomycetes. (a) Chlorogenic acid, a possibly stomach-irritating compound in coffee, was selectively degraded to quinic and caffeic acid using a p-coumaroyl esterase (CAE) [44]. (b) Vanillin can be generated from curcumin using a three-step enzyme process. FAE: ferulic acid esterase. Modified after [46]. (c) The vanilla and almond-like flavour piperonal can be generated by co-oxidation from the abundant pepper alkaloid piperine. Modified after [47].
Scheme 2.
Biotransformations using enzymes from basidiomycetes. (a) Chlorogenic acid, a possibly stomach-irritating compound in coffee, was selectively degraded to quinic and caffeic acid using a p-coumaroyl esterase (CAE) [44]. (b) Vanillin can be generated from curcumin using a three-step enzyme process. FAE: ferulic acid esterase. Modified after [46]. (c) The vanilla and almond-like flavour piperonal can be generated by co-oxidation from the abundant pepper alkaloid piperine. Modified after [47].

Figure 3.
A peptidase from Pleurotus sajor-caju (SPPS) (a) showed remarkable temperature stability, which was improved through the addition of Ca2Cl and sorbitol. (b) SPPS showed improved hydrolysis of casein in comparison to other peptidases (SPTC: peptidase from Trametes cingulata, Flavourzyme 500 L: commercial peptidase mix from Aspergillus oryzae, Thermolysin type X: commercial peptidase from Geobacillus stearothermophilus). Reprinted from [55].
Figure 3.
A peptidase from Pleurotus sajor-caju (SPPS) (a) showed remarkable temperature stability, which was improved through the addition of Ca2Cl and sorbitol. (b) SPPS showed improved hydrolysis of casein in comparison to other peptidases (SPTC: peptidase from Trametes cingulata, Flavourzyme 500 L: commercial peptidase mix from Aspergillus oryzae, Thermolysin type X: commercial peptidase from Geobacillus stearothermophilus). Reprinted from [55].

Scheme 3.
Oxidative gelation of arabinoxylan using laccases. Modified after [76].
Scheme 3.
Oxidative gelation of arabinoxylan using laccases. Modified after [76].

Figure 4.
Structures of industrially interesting colourants, which are produced or degraded by basidiomycetes. (a) Cinnabarinic acid, a deep-red colourant; (b) malvidin-3-O-galactoside, a major colourant of red wine husks; (c) Reactive Blue 5, an anthraquinone dye.
Figure 4.
Structures of industrially interesting colourants, which are produced or degraded by basidiomycetes. (a) Cinnabarinic acid, a deep-red colourant; (b) malvidin-3-O-galactoside, a major colourant of red wine husks; (c) Reactive Blue 5, an anthraquinone dye.

Figure 5.
Structures of industrially interesting volatile flavours, which are produced by basidiomycetes. (a) (+)-Nootkatone, the character impact compound of grapefruit; (b) (5Z,7E,9)-decatriene-2-one, volatile with a very strong pineapple flavour; (c) dodec-11-enal, a flavour with a coriander/leaf-like smell.
Figure 5.
Structures of industrially interesting volatile flavours, which are produced by basidiomycetes. (a) (+)-Nootkatone, the character impact compound of grapefruit; (b) (5Z,7E,9)-decatriene-2-one, volatile with a very strong pineapple flavour; (c) dodec-11-enal, a flavour with a coriander/leaf-like smell.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Berger, R.G.; Ersoy, F. Improved Foods Using Enzymes from Basidiomycetes. Processes 2022, 10, 726. https://doi.org/10.3390/pr10040726
AMA Style
Berger RG, Ersoy F. Improved Foods Using Enzymes from Basidiomycetes. Processes. 2022; 10(4):726. https://doi.org/10.3390/pr10040726
Chicago/Turabian StyleBerger, Ralf G., and Franziska Ersoy. 2022. "Improved Foods Using Enzymes from Basidiomycetes" Processes 10, no. 4: 726. https://doi.org/10.3390/pr10040726
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.