
Gallic Acid Enhances the Anti-Cancer Effect of Temozolomide in Human Glioma Cell Line via Inhibition of Akt and p38-MAPK Pathway

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Line, Reagents, and Chemicals
2.2. Drug Treatment and Cell Culture
2.3. Cellular Viability Assay
2.4. Intracellular ROS Analysis
2.5. Western Blot Analysis
2.6. Statistical Analysis
3. Results
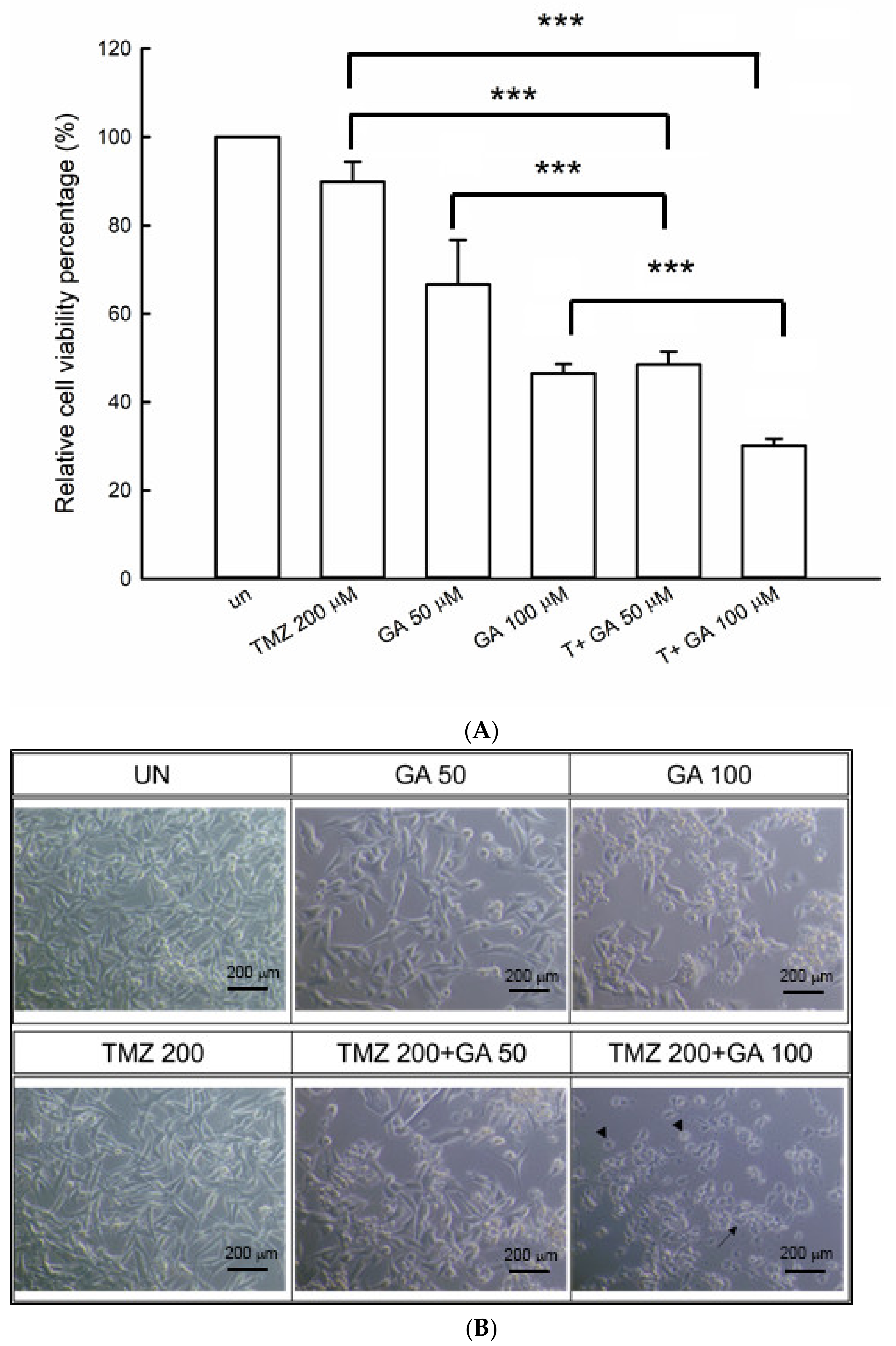
3.1. Ga Enhanced the Cell Viability Inhibition during the TMZ Treatment of U87MG Glioma Cells
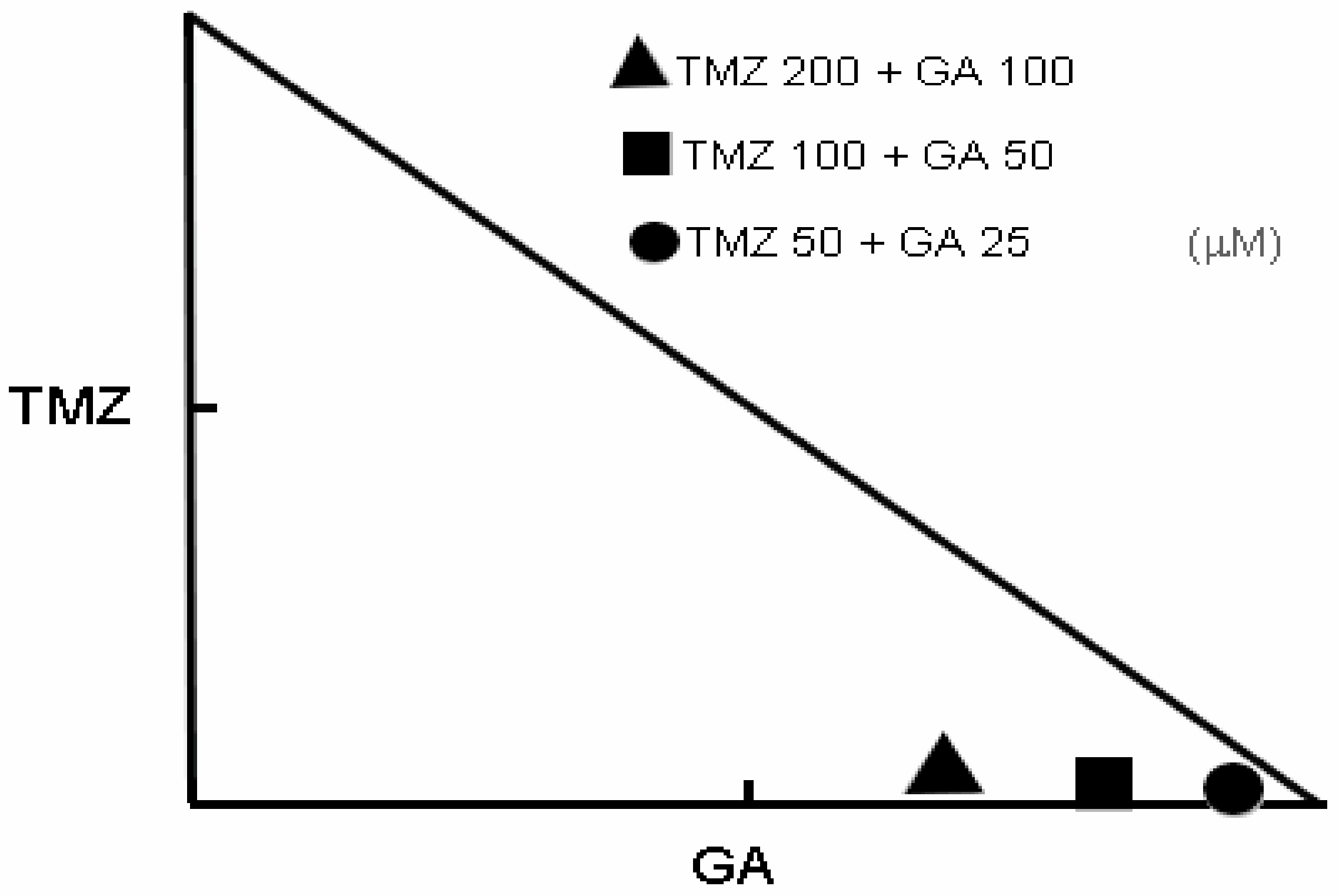
3.2. Synergism of Ga and TMZ Combination in Cell Viability Inhibition
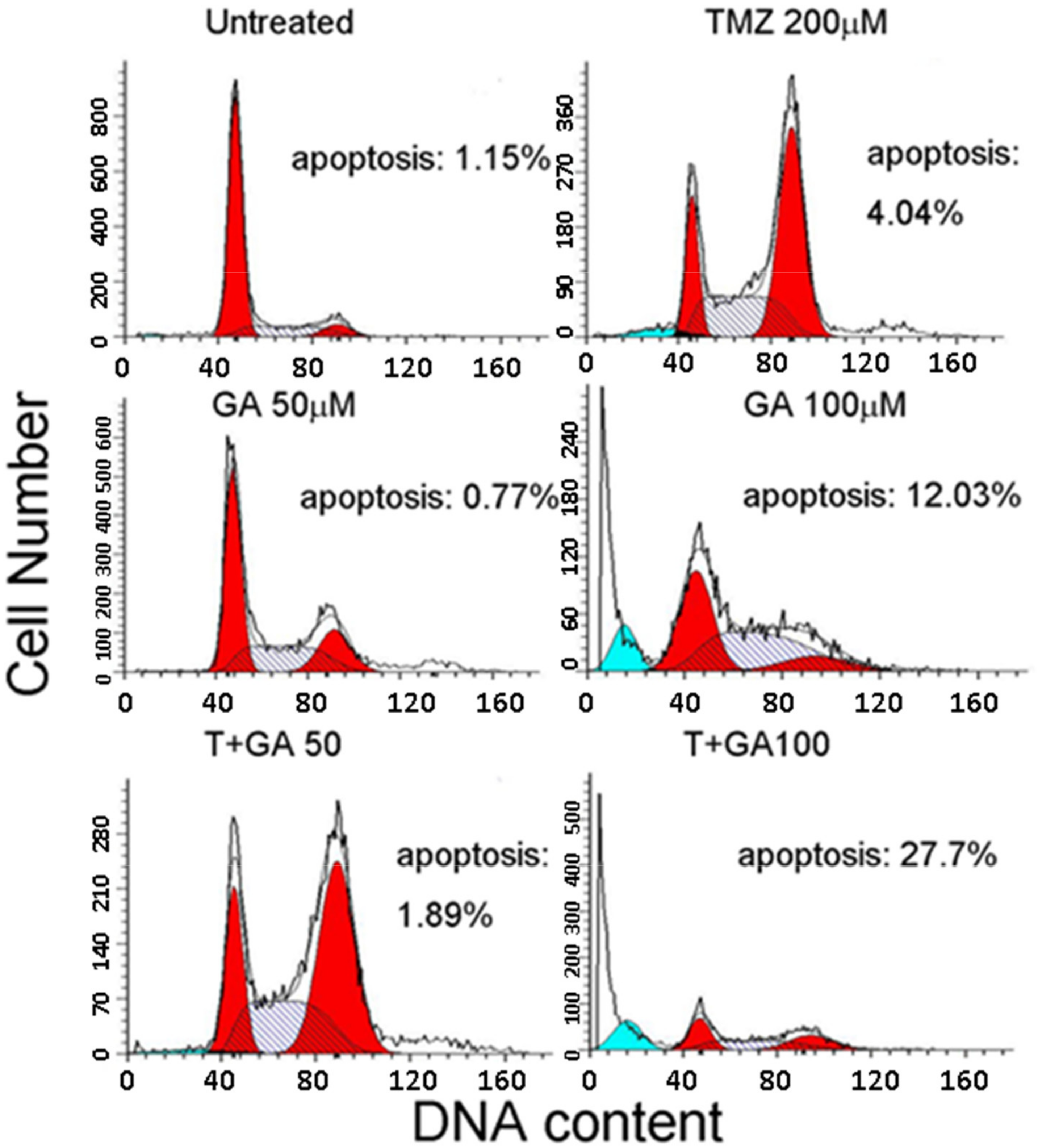
3.3. Ga Enhanced the Apoptosis during TMZ Treatment of U87MG Glioma Cells
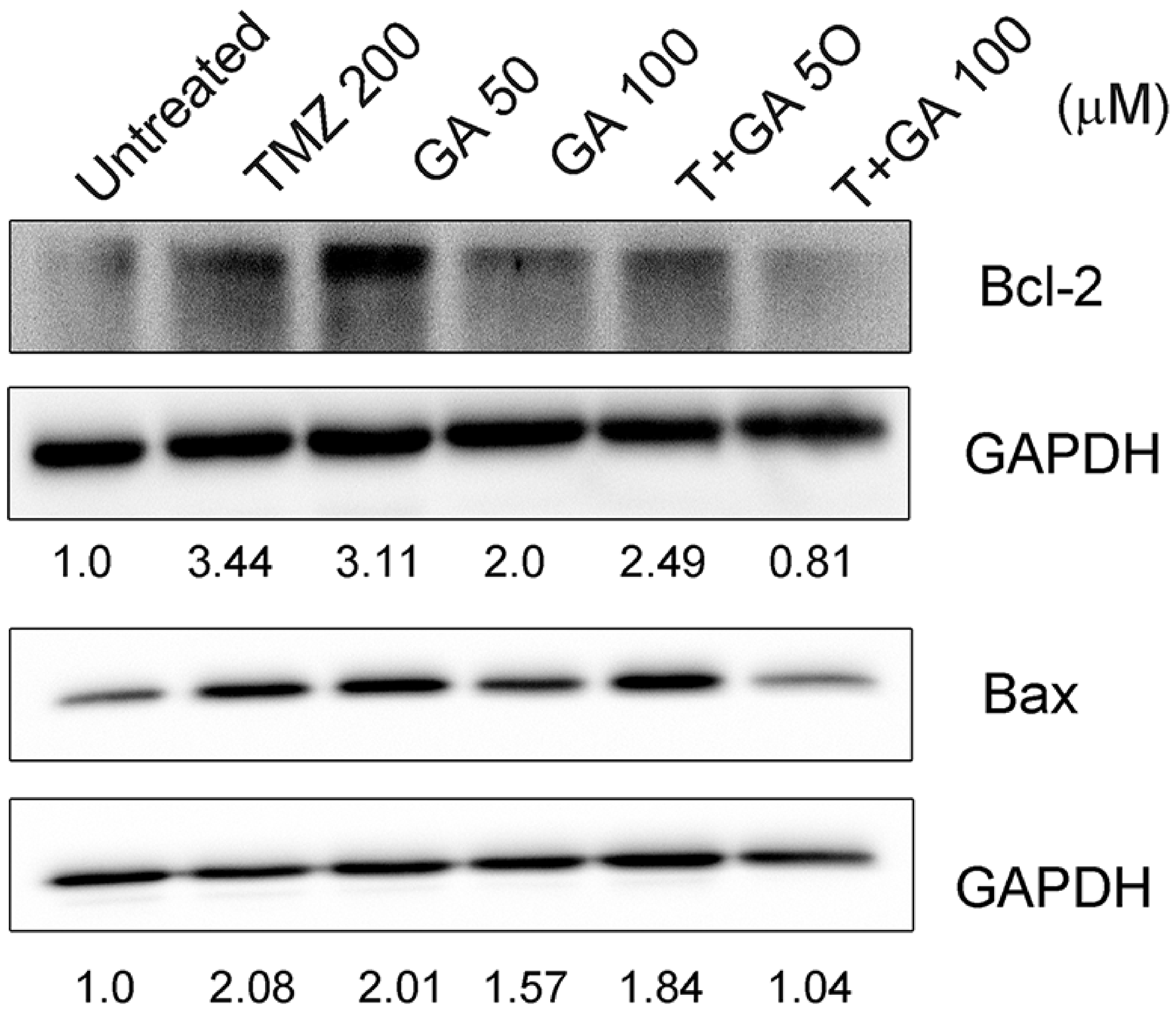
3.4. Ga Enhanced the Bcl-2 Inhibition of TMZ in U87MG Glioma Cell Line
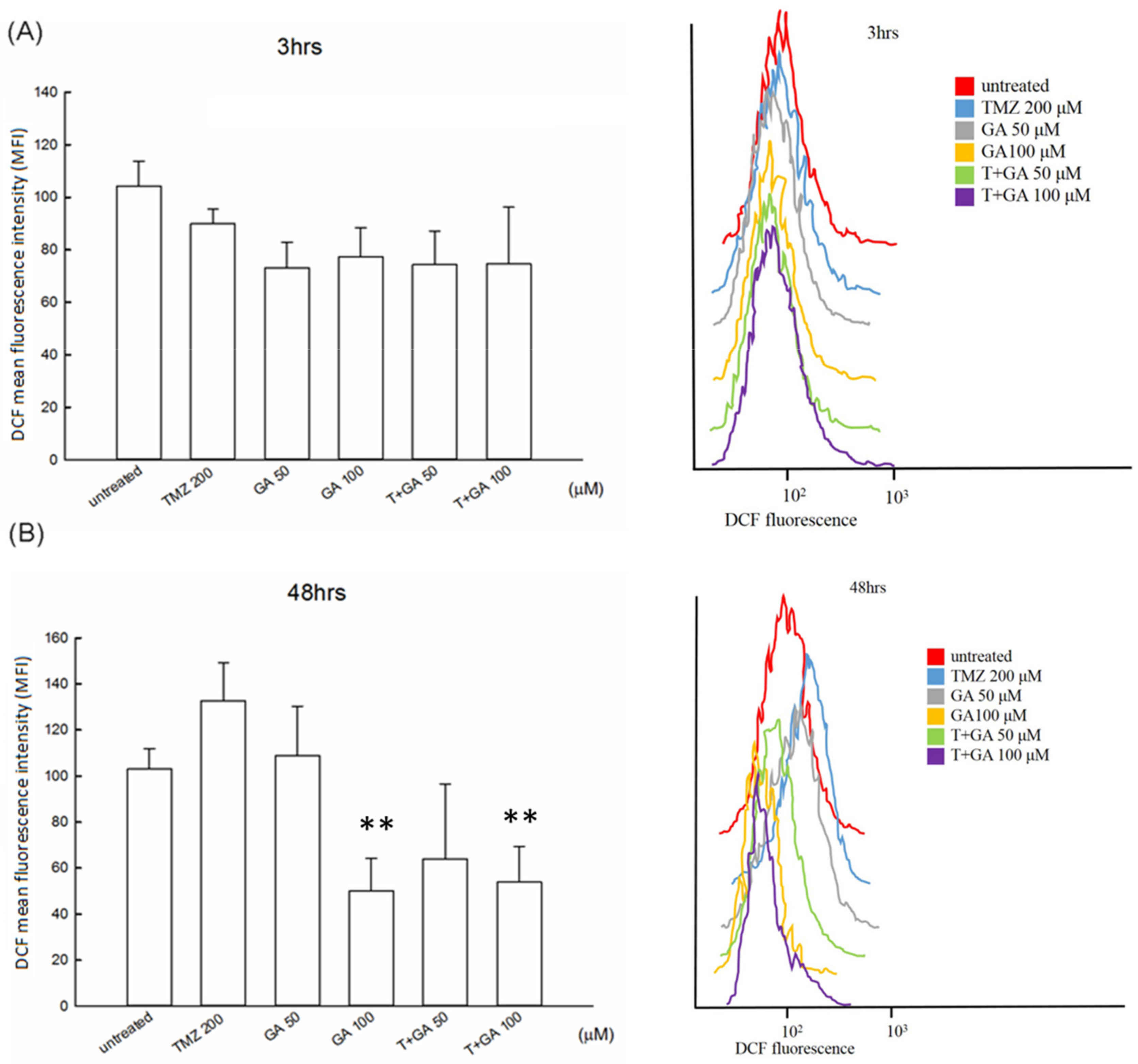
3.5. ROS Level in the Ga/TMZ Treatment
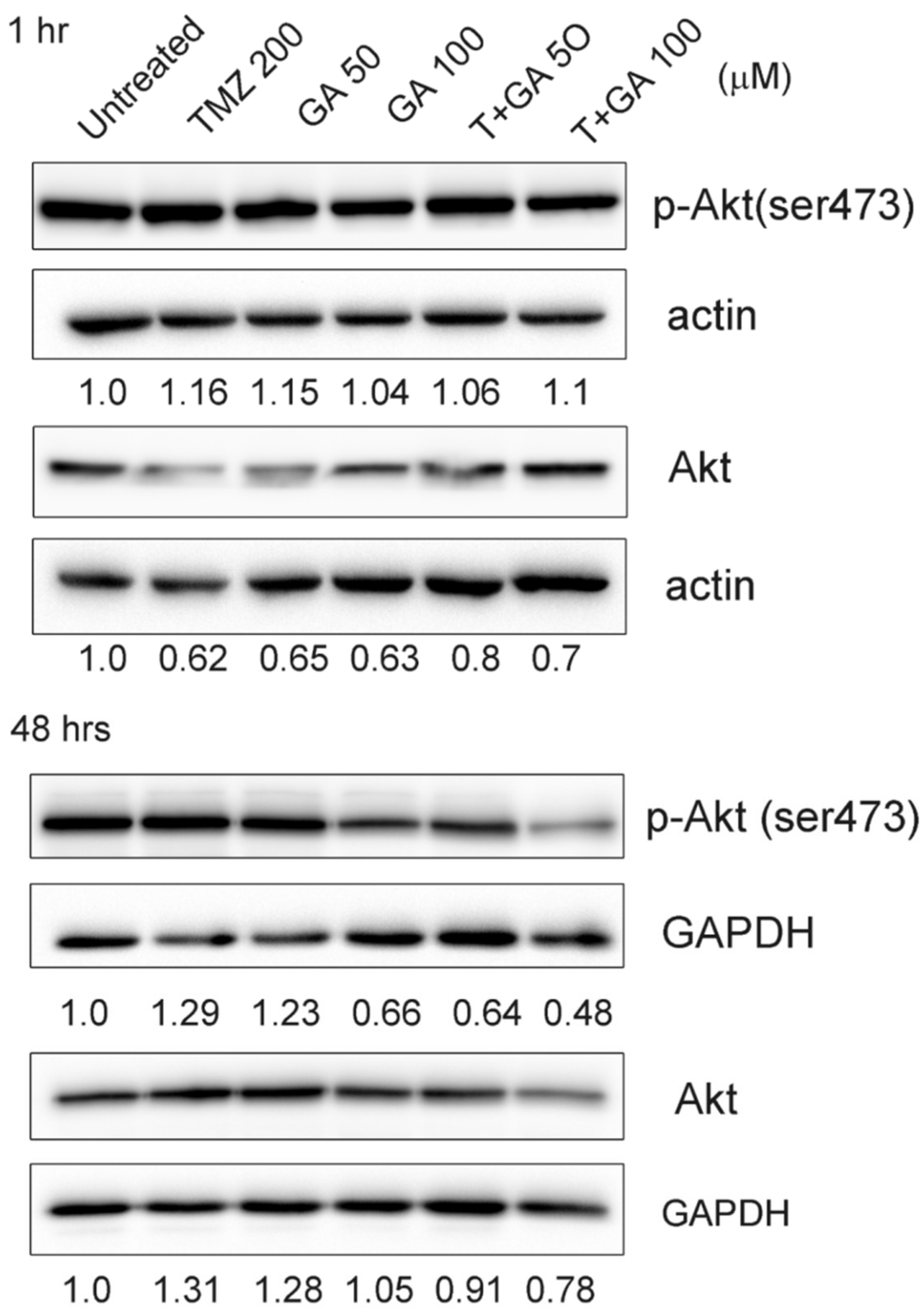
3.6. Ga Increased the Repression of Akt Phosphorylation on TMZ Incubation in U87MG Glioma Cells
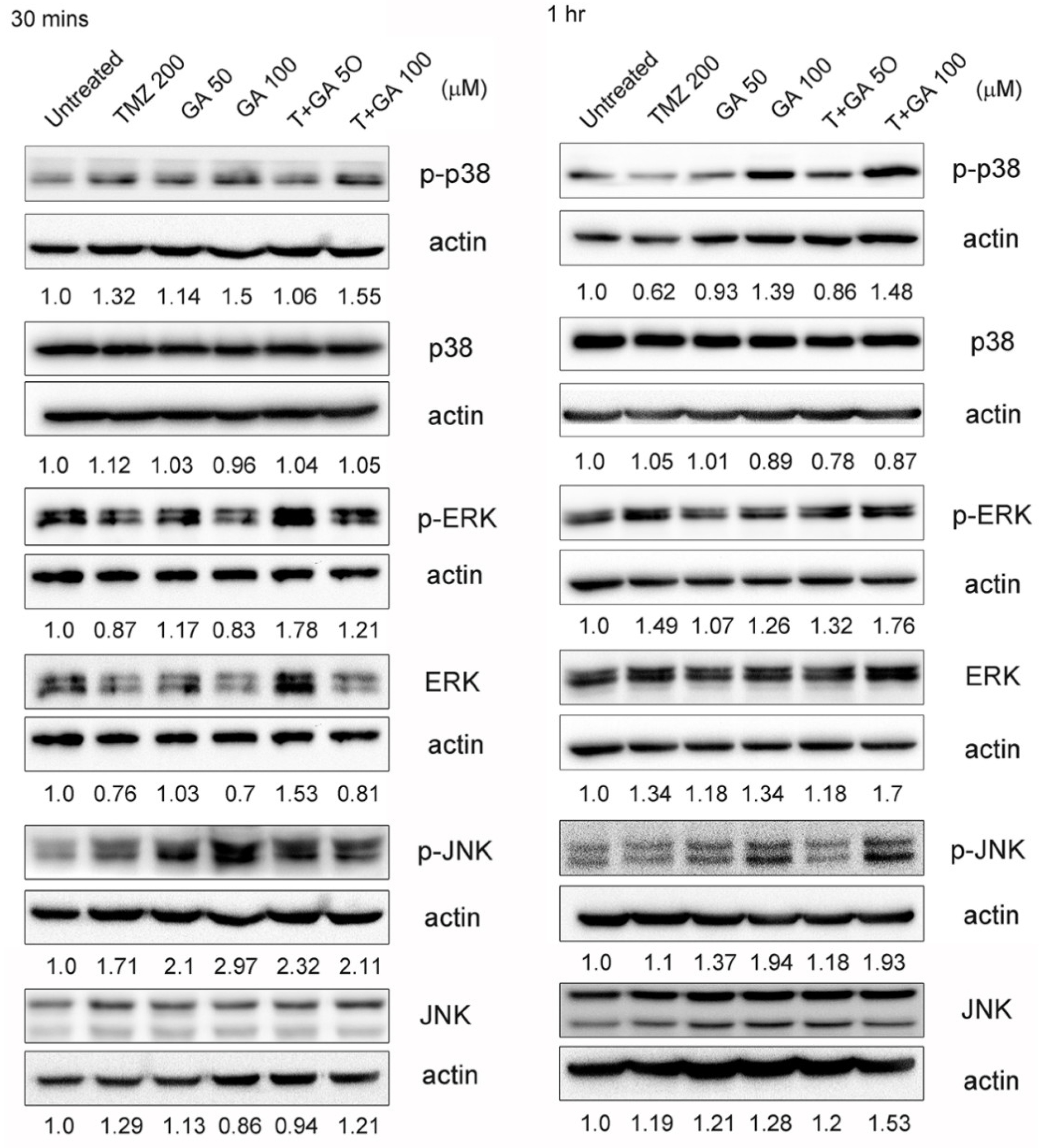
3.7. MAPK Expression in the Ga/TMZ Treatment
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lombardi, F.; Augello, F.R.; Artone, S.; Gugu, M.K.; Cifone, M.G.; Cique, B.; Palumbo, P. Up-regulation of cyclooxygenase-2 (COX-2) expression by temozolomide (TMZ) in human glioblastoma (GBM) cell lines. Int. J. Mol. Sci. 2022, 23, 1545. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Lu, C.; Zhou, P.; Zhao, L.; Lyu, X.; Yin, J.; Shi, Z.; You, Y. EIF4A3-induced circular RNA ASAP1 promotes tumorigenesis and temozolomide resistance of glioblastoma via NRAS/MEK1/ERK1-2 signaling. Neuro-Oncology 2021, 23, 611–624. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Pastorino, F.; Berry, P.; Bonner, J.; Kirk, C.; Wood, K.M.; Thomas, H.D.; Zhao, Y.; Daga, A.; Veal, G.J.; et al. Preclinical evaluation of the first intravenous small molecule MDM2 antagonist alone and in combination with temozolomide in neuroblastoma. Int. J. Cancer 2019, 144, 3146–3159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliveira, K.A.; Dal-Cim, T.A.; Lopes, F.G.; Nedel, C.B.; Tasca, C.I. Guanosine promotes cytotoxicity via adenosine receptors and induces apoptosis in temozolomide-treated A172 glioma cells. Purinergic Signal. 2017, 13, 305–318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.; Cong, D.; Li, Y.; Wang, D.; Li, Q.; Hu, S. Combination of sonodynamic with temozolomide inhibits C6 glioma migration and promotes mitochondrial pathway apoptosis via suppressing NHE-1 expression. Ultrason. Sonochemistry 2017, 39, 654–661. [Google Scholar] [CrossRef]
- Chio, C.-C.; Chen, K.-Y.; Chang, C.-K.; Chuang, J.-Y.; Liu, C.-C.; Liu, S.-H.; Chen, R.-M. Improved effects of honokiol on temozolomide-induced autophagy and apoptosis of drug-sensitive and-tolerant glioma cells. BMC Cancer 2018, 18, 379. [Google Scholar] [CrossRef] [Green Version]
- Ma, J.; Yang, Y.-R.; Chen, W.; Chen, M.-H.; Wang, H.; Wang, X.-D.; Sun, L.-L.; Wang, F.-Z.; Wang, D.-C. Fluoxetine synergizes with temozolomide to induce the CHOP-dependent endoplasmic reticulum stress-related apoptosis pathway in glioma cells. Oncol. Rep. 2016, 36, 676–684. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.-H.; Chang, Y.-J.; Ku, M.S.; Chung, K.-T.; Yang, J.-T. Enhancement of temozolomide-induced apoptosis by valproic acid in human glioma cell lines through redox regulation. J. Mol. Med. 2011, 89, 303–315. [Google Scholar] [CrossRef]
- Yang, J.-T.; Lee, I.; Lu, F.-J.; Chung, C.-Y.; Lee, M.-H.; Cheng, Y.-C.; Chen, K.-T.; Chen, C.-H. Propyl gallate exerts an antimigration effect on temozolomide-treated malignant glioma cells through inhibition of ROS and the NF-κB pathway. J. Immunol. Res. 2017, 2017, 9489383. [Google Scholar] [CrossRef] [Green Version]
- Kumar, P.P.; Madhuri, D.; Reddy, L.S.S.; Reddy, Y.D.; Somasekhar, G.; Sirisha, N.V.L.; Nagaraju, K.; Shouib, M.S.; Rizwaan, A.S. A new cerebral ischemic injury model in rats, preventive effect of gallic acid and in silico approaches. Saudi J. Biol. Sci. 2021, 28, 5204–5213. [Google Scholar] [CrossRef]
- Lu, Y.; Jiang, F.; Jiang, H.; Wu, K.; Zheng, X.; Cai, Y.; Katakowski, M.; Chopp, M.; To, S.-S.T. Gallic acid suppresses cell viability, proliferation, invasion and angiogenesis in human glioma cells. Eur. J. Pharmacol. 2010, 641, 102–107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsu, S.-S.; Chou, C.-T.; Liao, W.-C.; Shieh, P.; Kuo, D.-H.; Kuo, C.-C.; Jan, C.-R.; Liang, W.-Z. The effect of gallic acid on cytotoxicity, Ca2+ homeostasis and ROS production in DBTRG-05MG human glioblastoma cells and CTX TNA2 rat astrocytes. Chem.-Biol. Interact. 2016, 252, 61–73. [Google Scholar] [CrossRef] [PubMed]
- Paolini, A.; Curti, V.; Pasi, F.; Mazzini, G.; Nano, R.; Capelli, E. Gallic acid exerts a protective or an anti-proliferative effect on glioma T98G cells via dose-dependent epigenetic regulation mediated by miRNAs. Int. J. Oncol. 2015, 46, 1491–1497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jing, Z.; Li, M.; Wang, H.; Yang, Z.; Zhou, S.; Ma, J.; Meng, E.; Zhang, H.; Liang, W.; Hu, W.; et al. Gallic acid-gold nanoparticles enhance radiation-induced cell death of human glioma U251 cells. IUBMB Life 2021, 73, 398–407. [Google Scholar] [CrossRef] [PubMed]
- Liang, C.-Z.; Zhang, X.; Li, H.; Tao, Y.-Q.; Tao, L.-J.; Yang, Z.-R.; Zhou, X.-P.; Shi, Z.-L.; Tao, H.-M. Gallic acid induces the apoptosis of human osteosarcoma cells in vitro and in vivo via the regulation of mitogen-activated protein kinase pathways. Cancer Biother. Radiopharm. 2012, 27, 701–710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moghtaderi, H.; Sepehri, H.; Delphi, L.; Attari, F. Gallic acid and curcumin induce cytotoxicity and apoptosis in human breast cancer cell MDA-MB-231. Bioimpacts 2018, 8, 185–194. [Google Scholar] [CrossRef] [Green Version]
- Shahrzad, S.; Aoyagi, K.; Winter, A.; Koyama, A.; Bitsch, I. Pharmacokinetics of Gallic Acid and Its Relative Bioavailability from Tea in Healthy Humans. J. Nutrition 2001, 131, 1207–1210. [Google Scholar] [CrossRef]
- Wang, R.; Ma, L.; Weng, D.; Yao, J.; Liu, X.; Jin, F. Gallic acid induces apoptosis and enhances the anticancer effects of cisplatin in human small cell lung cancer H446 cell line via the ROS-dependent mitochondrial apoptotic pathway. Oncol. Rep. 2016, 35, 3075–3083. [Google Scholar] [CrossRef] [Green Version]
- Jeong, C.-H.; Joo, S.-H. Downregulation of reactive oxygen species in apoptosis. J. Cancer Prev. 2016, 21, 13–20. [Google Scholar] [CrossRef] [Green Version]
- George, S.; Abrahamse, H. Redox potential of antioxidants in cancer progression and prevention. Antioxidants 2020, 9, 1156. [Google Scholar] [CrossRef]
- Dehkordi, M.H.; Tashakor, A.; O’Connell, E.; Fearnhead, H.O. Apoptosome-dependent myotube formation involves activation of caspase-3 in differentiating myoblasts. Cell Death Dis. 2020, 11, 308. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Ma, L.; Wu, P.; Li, W.; Li, T.; Gu, R.; Dan, X.; Li, Z.; Fan, X.; Xiao, Z. Gallic acid has anticancer activity and enhances the anticancer effects of cisplatin in non-small cell lung cancer A549 cells via the JAK/STAT3 signaling pathway. Oncol. Rep. 2019, 41, 1779–1788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, R.; Zhang, M.; Meng, H.; Xu, D.; Xie, Y. Gallic acid targets acute myeloid leukemia via Akt/mTOR-dependent mitochondrial respiration inhibition. Biomed. Pharmacother. 2018, 105, 491–497. [Google Scholar] [CrossRef]
- Zeng, M.; Su, Y.; Li, K.; Jin, D.; Li, Q.; Li, Y.; Zhou, B. Gallic acid inhibits bladder cancer T24 cell progression through mitochondrial dysfunction and PI3K/Akt/NF-κB signaling suppression. Front. Pharmacol. 2020, 11, 1222. [Google Scholar]
- Chen, Y.-J.; Lin, K.-N.; Jhang, L.-M.; Huang, C.-H.; Lee, Y.-C.; Chang, L.-S. Gallic acid abolishes the EGFR/Src/Akt/Erk-mediated expression of matrix metalloproteinase-9 in MCF-7 breast cancer cells. Chem.-Biol. Interact. 2016, 252, 131–140. [Google Scholar] [CrossRef]
- Demiroglu-Zergeroglu, A.; Candemir, G.; Turhanlar, E.; Sagir, F.; Ayvali, N. EGFR-dependent signalling reduced and p38 dependent apoptosis required by Gallic acid in Malignant Mesothelioma cells. Biomed. Pharmacother. 2016, 84, 2000–2007. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, J.-T.; Lee, I.-N.; Chen, C.-H.; Lu, F.-J.; Chung, C.-Y.; Lee, M.-H.; Cheng, Y.-C.; Chen, K.-T.; Peng, J.-Y.; Chen, C.-H. Gallic Acid Enhances the Anti-Cancer Effect of Temozolomide in Human Glioma Cell Line via Inhibition of Akt and p38-MAPK Pathway. Processes 2022, 10, 448. https://doi.org/10.3390/pr10030448
Yang J-T, Lee I-N, Chen C-H, Lu F-J, Chung C-Y, Lee M-H, Cheng Y-C, Chen K-T, Peng J-Y, Chen C-H. Gallic Acid Enhances the Anti-Cancer Effect of Temozolomide in Human Glioma Cell Line via Inhibition of Akt and p38-MAPK Pathway. Processes. 2022; 10(3):448. https://doi.org/10.3390/pr10030448
Chicago/Turabian StyleYang, Jen-Tsung, I-Neng Lee, Chun-Han Chen, Fung-Jou Lu, Chiu-Yen Chung, Ming-Hsueh Lee, Yu-Ching Cheng, Kuo-Tai Chen, Jyun-Yu Peng, and Ching-Hsein Chen. 2022. "Gallic Acid Enhances the Anti-Cancer Effect of Temozolomide in Human Glioma Cell Line via Inhibition of Akt and p38-MAPK Pathway" Processes 10, no. 3: 448. https://doi.org/10.3390/pr10030448