Salycilic Acid Induces Exudation of Crocin and Phenolics in Saffron Suspension-Cultured Cells

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Results
2.1. Cell Suspension Culture and SA Effects on Cell Growth
2.2. Analysis of Crocetin Esters and PC Profile Following the Imposition of SA
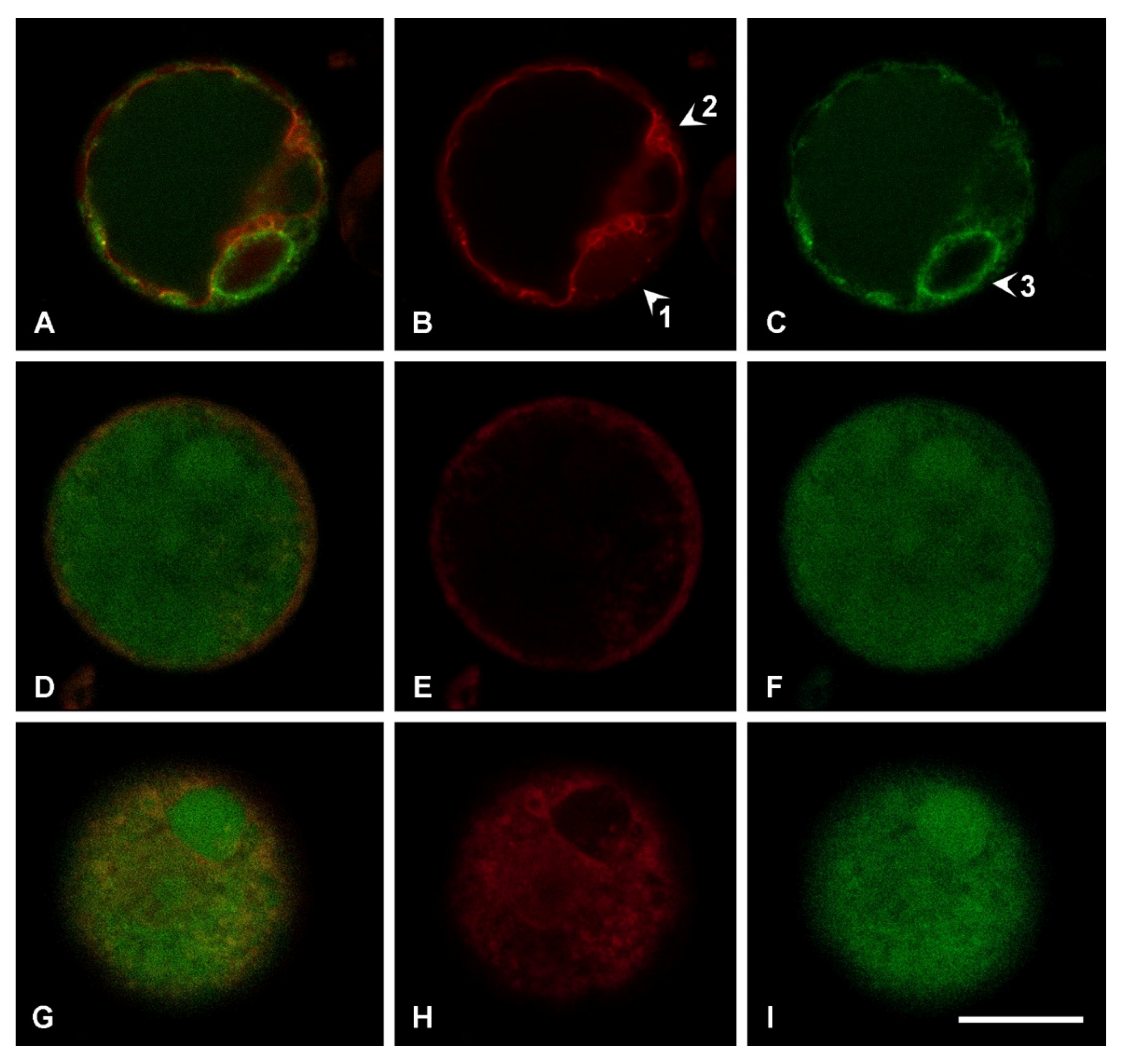
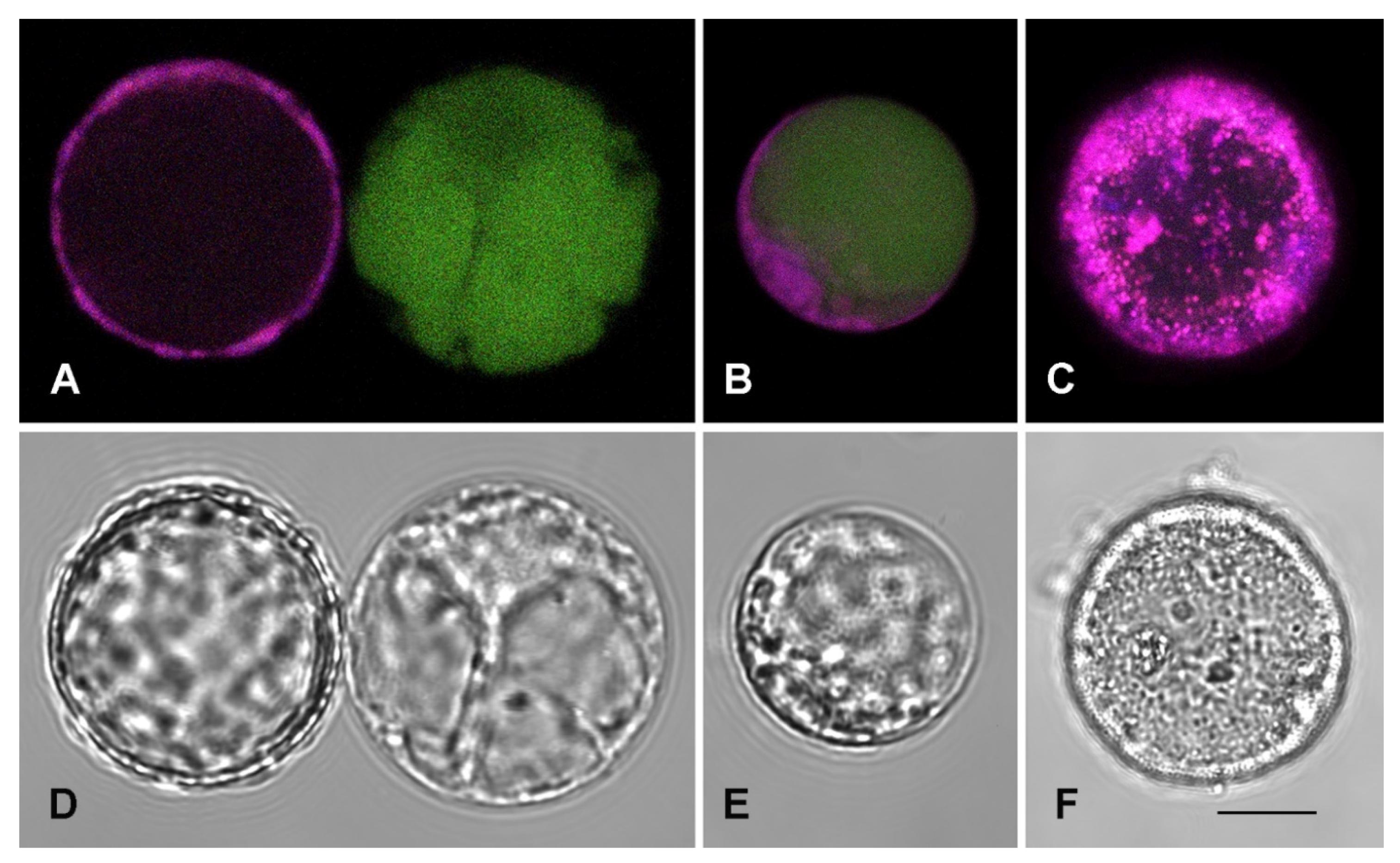
2.3. SA Effect on Subcellular Compartmentalization
2.4. Optimization of the Elicitation Time and Harvest Time for Metabolite Productions
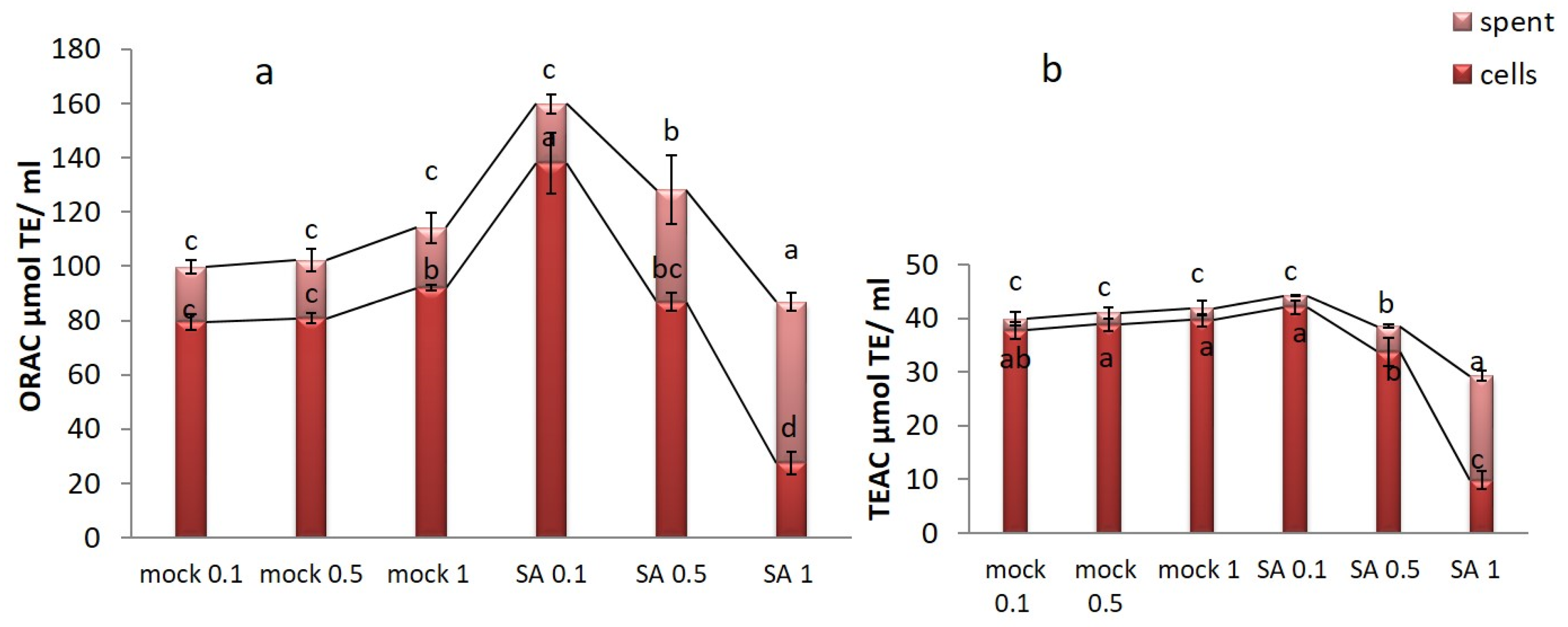
2.5. Antioxidant Activity of Saffron Cell Culture Extracts
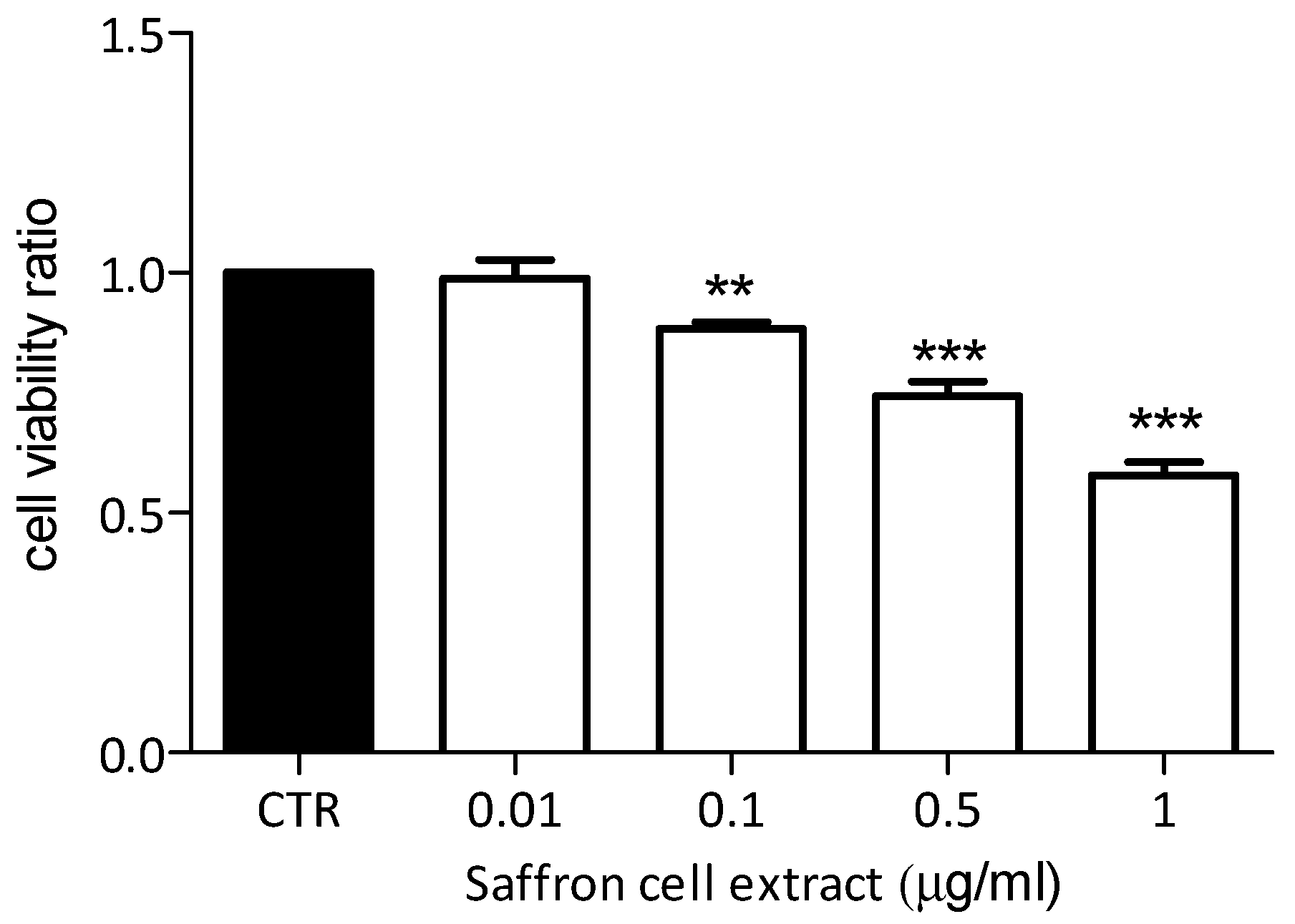
2.6. Effects of Saffron Cell Suspension Extract on Breast Cancer Cells
3. Discussion
4. Materials and Methods
4.1. Establishment of Cell Suspension Cultures and SA Treatment
4.2. Extraction of Crocins from Cells
4.3. Extraction of Crocins and Phenolic Compounds from the Spent Medium of Suspension Cultures
4.4. Extraction of Phenolic Compounds from Cells
4.5. Total Phenolic Content
4.6. HPLC-DAD Analysis
4.7. Protoplasts Preparation, Transformation, and Analysis
4.8. Trolox Equivalent Antioxidant Capacity (TEAC) Assay
4.9. Oxygen Radical Absorbance Capacity (ORAC) Assay
4.10. 1-Diphenyl-2-picrylhydrazyl (DPPH) Radical Scavenging Assay
4.11. MTT Assay
4.12. Data Acquisition and Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Fernandez, J.A. Genetic resources of saffron and allies (Crocus spp.). Acta Hortic. 2007, 739, 167–185. [Google Scholar] [CrossRef]
- Rahimi, M. Chemical and medicinal properties of saffron. Bull. Environ. Pharmacol. Life Sci. 2015, 4, 69–81. [Google Scholar]
- Abe, K.; Saito, H. Effects of saffron and its constituent crocin on learning behavior and long-term potentiation. Phytother. Res. 2000, 14, 149–152. [Google Scholar] [CrossRef]
- Angaji, S.A.; Mousavi, A.; Babapour, E. Antioxidants: A few key points. Ann. Biol. Res. 2012, 3, 3968–3977. [Google Scholar]
- Kawabata, K.; Tung, N.H.; Shoyama, Y.; Sugie, S.; Mori, T.; Tanaka, T. Dietary crocin inhibits colitis and colitis-associated colorectal carcinogenesis in male ICR mice. Evid. Based. Complement. Altern. Med. 2012, 2012, 820415. [Google Scholar] [CrossRef]
- Veisi, A.; Akbari, G.; Mard, S.A.; Badfar, G.; Zarezade, V.; Mirshekar, M.A. Role of crocin in several cancer cell lines: An updated review. Iran. J. Basic Med. Sci. 2019, 23, 3–12. [Google Scholar]
- Jam, I.N.; Sahebkar, A.H.; Eslami, S.; Mokhber, N.; Nosrati, M.; Khademi, M.; Foroutan-Tanha, M.; Ghayour-Mobarhan, M.; Hadizadeh, F.; Ferns, G.; et al. The effects of crocin on the symptoms of depression in subjects with metabolic syndrome. Adv. Clin. Exp. Med. 2017, 26, 925–930. [Google Scholar]
- Adabizadeh, M.; Mehri, S.; Rajabpour, M.; Abnous, K.; Rashedinia, M.; Hosseinzadeh, H. The effects of crocin on spatial memory impairment induced by hyoscine: Role of NMDA, AMPA, ERK, and CaMKII proteins in rat hippocampus. Iran. J. Basic. Med. Sci. 2019, 22, 601–609. [Google Scholar]
- Smetanska, I. Production of secondary metabolites using plant cell cultures. Adv. Biochem. Eng. Biotechnol. 2008, 111, 187–228. [Google Scholar]
- Caretto, S.; Paradiso, A.; D’Amico, L.; De Gara, L. Ascorbate and glutathione metabolism in two sunflower cell lines of differing α-tocopherol biosynthetic capability. Plant Physiol. Biochem. 2002, 40, 509–513. [Google Scholar] [CrossRef]
- Yue, W.; Ming, Q.L.; Lin, B.; Rahman, K.; Zheng, C.J.; Han, T.; Qin, L.P. Medicinal plant cell suspension cultures: Pharmaceutical applications and high-yielding strategies for the desired secondary metabolites. Crit. Rev. Biotechnol. 2016, 36, 215–232. [Google Scholar] [CrossRef] [PubMed]
- Caretto, S.; Nisi, R.; Paradiso, A.; De Gara, L. Tocopherol production in plant cell cultures. Mol. Nutr. Food Res. 2010, 54, 726–730. [Google Scholar] [CrossRef] [PubMed]
- Almagro, L.; Tudela, L.R.; Sabater-Jara, A.B.; Miras-Moreno, B.; Pedreño, M.A. Cyclodextrins increase phytosterol and tocopherol levels in suspension-cultured cells obtained from mung beans and safflower. Biotechnol. Prog. 2017, 33, 1662–1665. [Google Scholar] [CrossRef] [PubMed]
- Caretto, S.; Quarta, A.; Durante, M.; Nisi, R.; De Paolis, A.; Blando, F.; Mita, G. Methyl jasmonate and miconazole differently affect arteminisin production and gene expression in Artemisia annua suspension cultures. Plant Biol. 2011, 13, 51–58. [Google Scholar] [CrossRef]
- Malik, S.; Cusidó, R.M.; Mirjalili, M.H.; Moyano, E.; Palazón, J.; Bonfill, M. Production of the anticancer drug taxol in Taxus baccata suspension cultures: A review. Process Biochem. 2011, 46, 23–34. [Google Scholar] [CrossRef]
- Guo, B.; Liang, Y.C.; Zhu, Y.G.; Zhao, F.J. Role of salicylic acid in alleviating oxidative damage in rice roots (Oryza sativa) subjected to cadmium stress. Environ. Pollut. 2007, 147, 743–749. [Google Scholar] [CrossRef]
- Hayat, Q.; Hayat, S.; Irfan, M.; Ahmad, A. Effect of exogenous salicylic acid under changing environment: A review. Environ. Exp. Bot. 2010, 68, 14–25. [Google Scholar] [CrossRef]
- Gorelick, J.; Bernstein, N. Chapter Five—Elicitation: An Underutilized Tool in the Development of Medicinal Plants as a Source of Therapeutic Secondary Metabolites. Adv. Agron. 2014, 124, 201–230. [Google Scholar]
- Mendoza, D.; Cuaspuda, O.; Ariasa, J.P.; Ruizc, O.; Ariasa, M. Effect of salicylic acid and methyl jasmonate in the production of phenolic compounds in plant cell suspension cultures of Thevetia peruviana. Biotechnol. Rep. 2018, 19, e00273. [Google Scholar] [CrossRef]
- Thiruvengadam, M.; Rekha, K.; Rajakumar, G.; Lee, T.J.; Kim, S.H.; Chung, I.M. Enhanced Production of Anthraquinones and Phenolic Compounds and Biological Activities in the Cell Suspension Cultures of Polygonum multiflorum. Int. J. Mol. Sci. 2016, 17, 1912. [Google Scholar] [CrossRef] [Green Version]
- Mahalakshmi, R.; Eganathan, P.; Parida, A. Salicylic acid elicitation on the production of secondary metabolite by cell cultures of Jatropha curcas L. Int. J. Pharm. Pharm. Sci. 2013, 5, 655–659. [Google Scholar]
- Moradi, A.; Zarinkamar, F.; Caretto, S.; Azadi, P. Influence of thidiazuron on callus induction and crocin production in corm and style explants of Crocus sativus L. Acta Physiol. Plant 2018, 40, 185. [Google Scholar] [CrossRef]
- Di Sansebastiano, G.P.; Barozzi, F. Transient Secretory Enzyme Expression in Leaf Protoplasts to Characterize SNARE Functional Classes in Conventional and Unconventional Secretion. In Plant Protein Secretion. Methods in Molecular Biology; Jiang, L., Ed.; Humana Press: New York, NY, USA, 2017; Volume 1662, pp. 209–221. [Google Scholar]
- Stigliano, E.; Faraco, M.; Neuhaus, J.M.; Montefusco, A.; Dalessandro, G.; Piro, G.; Di Sansebastiano, G.P. Two glycosylated vacuolar GFPs are new markers for ER-to-vacuole sorting. Plant Physiol. Biochem. 2013, 73, 337–343. [Google Scholar] [CrossRef]
- De Benedictis, M.; Bleve, G.; Faraco, M.; Stigliano, E.; Grieco, F.; Piro, G.; Dalessandro, G.; Di Sansebastiano, G.P. AtSYP51/52 functions diverge in the post-Golgi traffic and differently affect vacuolar sorting. Mol. Plant 2013, 6, 916–930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Sansebastiano, G.P.; Fornaciari, S.; Barozzi, F.; Piro, G.; Arru, L. New insights on plant cell elongation: A role for acetylcholine. Int. J. Mol. Sci. 2014, 15, 4565–4582. [Google Scholar] [CrossRef] [Green Version]
- Davalos, A.; Gómez-Cordovés, C.; Artolomé, B. Extending applicability of the oxygen radical absorbance capacity (ORAC− fluorescein) assay. J. Agric. Food Chem. 2004, 52, 48–54. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Blois, M.S. Antioxidant determinations by the use of a stable free radical. Nature 1958, 29, 1199–1200. [Google Scholar] [CrossRef]
- Negbi, M. (Ed.) Saffron cultivation. In Saffron: Crocus sativus L.; CRC Press: Boca Raton, FL, USA, 1999; pp. 1–17. [Google Scholar]
- Azadi, P.; Bagheri, K.; Gholami, M.; Mirmasoumi, M.; Moradi, A.; Sharafi, A. Thin cell layer, a suitable explant for in vitro regeneration of saffron (Crocus sativus L.). J. Agric. Sci. Technol. 2017, 19, 1429–1435. [Google Scholar]
- Sharifi, G.; Ebrahimzadeh, H.; Ghareyazie, B.; Karimi, M. Globular embryo-like structures and highly efficient thidiazuron-induced multiple shoot formation in saffron (Crocus sativus L.). In Vitro Cell Dev. Biol. Plant 2010, 46, 274–280. [Google Scholar] [CrossRef]
- Mir, J.I.; Ahmed, N.; Wani, S.H.; Rashid, R.; Mir, H.; Sheikh, M.A. In vitro development of microcorms and stigma like structures in saffron (Crocus sativus L.). Physiol. Mol. Biol. Plants 2010, 16, 369–373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parray, J.A.; Kamili, A.N.; Hamid, R.; Husaini, A.M. In vitro cormlet production of saffron (Crocus sativus L. Kashmirianus) and their flowering response under greenhouse. GM Crops Food 2012, 3, 289–295. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.A.; Wang, X.; Zhao, B.; Yuan, X.; Wang, Y. Production of crocin using Crocus sativus callus by two-stage culture system. Biotechnol. Lett. 2003, 25, 1235–1238. [Google Scholar] [CrossRef] [PubMed]
- Namin, M.; Ebrahimzadeh, H.; Ghareyazie, B.; Radjabian, T.; Namin, H. Initiation and origin of stigma-like structures (SLS) on ovary and style explants of saffron in tissue culture. Acta Biol. Crac. Ser. Bot. 2010, 52, 55–60. [Google Scholar] [CrossRef]
- Rezaei, A.; Ghanati, F.; Dehaghi, M.A. Stimulation of taxol production by combined salicylic acid elicitation and sonication in Taxus baccata cell culture. In Proceedings of the 2011 International Conference on Life Science and Technology IPCBEE, Singapore, 7–9 January 2011; pp. 193–197. [Google Scholar]
- Dong, J.; Wan, G.; Liang, Z. Accumulation of salicylic acid-induced phenolic compounds and raised activities of secondary metabolic and antioxidative enzymes in Salvia miltiorrhiza cell culture. J. Biotechnol. 2010, 148, 99–104. [Google Scholar] [CrossRef] [PubMed]
- Conceiçao, L.F.; Ferreres, F.; Tavares, R.M.; Dias, A.C. Induction of phenolic compounds in Hypericum perforatum L. cells by Colletotrichum gloeosporioides elicitation. Phytochemistry 2006, 67, 149–155. [Google Scholar] [CrossRef]
- Shukor, M.F.A.; Ismail, I.; Zainal, Z.; Noor, N.M. Development of a Polygonum minus cell suspension culture system and analysis of secondary metabolites enhanced by elicitation. Acta Physiol. Plant 2013, 35, 1675–1689. [Google Scholar] [CrossRef]
- Cai, Z.; Kastell, A.; Knorr, D.; Smetanska, I. Exudation: An expanding technique for continuous production and release of secondary metabolites from plant cell suspension and hairy root cultures. Plant Cell Rep. 2012, 31, 461–477. [Google Scholar] [CrossRef]
- Nielsen, E.; Temporiti, M.E.; Cella, R. Improvement of phytochemical production by plant cells and organ culture and by genetic engineering. Plant Cell Rep. 2019, 38, 1199–1215. [Google Scholar] [CrossRef]
- Barber, M.S.; McConnell, V.S.; DeCaux, B.S. Antimicrobial intermediates of the general phenylpropanoid and lignin specific pathways. Phytochemistry 2000, 54, 53–56. [Google Scholar] [CrossRef]
- Dixon, R.A.; Achnine, L.; Kota, P.; Liu, C.J.; Reddy, M.S.S.; Wang, L. The phenylpropanoid pathway and plant defence-a genomics perspective. Mol. Plant Pathol. 2002, 3, 371–390. [Google Scholar] [CrossRef] [PubMed]
- Gadzovska, S.; Maury, S.; Delaunay, A.; Spasenoski, M.; Hagege, D.; Courtois, D.; Joseph, C. The influence of salicylic acid elicitation of shoots, callus, and cell suspension cultures on production of naphtodianthrones and phenylpropanoids in Hypericum perforatum L. Plant Cell Tiss. Organ Cult. 2013, 113, 25–39. [Google Scholar] [CrossRef]
- Riedel, H.; Akumo, D.N.; Saw, N.M.M.T.; Kütük, O.; Neubauer, P.; Smetanska, I. Elicitation and precursor feeding influence phenolic acids composition in Vitis vinifera suspension culture. Afr. J. Biotechnol. 2012, 11, 3000–3008. [Google Scholar]
- Hao, W.; Guo, H.; Zhang, J.; Hu, G.; Yao, Y.; Dong, J. Hydrogen peroxide is involved in Salicylic acid-elicited rosmarinic acid production in Salvia miltiorrhiza cell cultures. Sci. World J. 2014, 2014, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barozzi, F.; Papadia, P.; Stefano, G.; Renna, L.; Brandizzi, F.; Migoni, D.; Fanizzi, F.P.; Piro, G.; Di Sansebastiano, G.P. Variation in Membrane Trafficking Linked to SNARE AtSYP51 Interaction with Aquaporin NIP1;1. Front. Plant Sci. 2019, 9, 1949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Caroli, M.; Lenucci, M.S.; Di Sansebastiano, G.P.; Dalessandro, G.; De Lorenzo, G.; Piro, G. Dynamic protein traf- ficking to the cell wall. Plant Signal Behav. 2011, 6, 94–97. [Google Scholar] [CrossRef] [Green Version]
- Stigliano, E.; Di Sansebastiano, G.P.; Neuhaus, J.M. Contribution of chitinase A’s C-terminal vacuolar sorting determinant to the study of soluble protein compartmentation. Int. J. Mol. Sci. 2014, 15, 11030–11039. [Google Scholar] [CrossRef] [Green Version]
- Mashmoul, M.; Azlan, A.; Khaza’ai, H.; Yusof, B.N.M.; Noor, S.M. Saffron: A natural potent antioxidant as a promising anti-obesity drug. Antioxidants 2013, 2, 293–308. [Google Scholar] [CrossRef] [Green Version]
- Rahaiee, S.; Moini, S.; Hashemi, M.; Shojaosadati, S.A. Evaluation of antioxidant activities of bioactive compounds and various extracts obtained from saffron (Crocus sativus L.): A review. J. Food. Sci. Technol. 2015, 52, 1881–1888. [Google Scholar] [CrossRef] [Green Version]
- Parray, J.A.; Kamili, A.N.; Hamid, R.; Reshi, Z.A.; Qadri, R.A. Antibacterial and antioxidant activity of methanol extracts of Crocus sativus L. cv. Kashmirianus. Front. Life Sci. 2015, 8, 40–46. [Google Scholar] [CrossRef]
- Karimi, E.; Oskoueian, E.; Hendra, R.; Jaafar, H.Z. Evaluation of Crocus sativus L. stigma phenolic and flavonoid compounds and its antioxidant activity. Molecules 2010, 15, 6244–6256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baba, S.A.; Malik, A.H.; Wani, Z.A.; Mohiuddin, T.; Shah, Z.; Abbas, N.; Ashraf, N. Phytochemical analysis and antioxidant activity of different tissue types of Crocus sativus and oxidative stress alleviating potential of saffron extract in plants, bacteria, and yeast. S. Afr. J. Bot. 2015, 99, 80–87. [Google Scholar] [CrossRef]
- Gismondi, A.; Serio, M.; Canuti, L.; Canini, A. Biochemical, antioxidant and antineoplastic properties of Italian saffron (Crocus sativus L.). Am. J. Plant Sci. 2012, 3, 1573–1580. [Google Scholar] [CrossRef] [Green Version]
- Assimopoulou, A.; Sinakos, Z.; Papageorgiou, V. Radical scavenging activity of Crocus sativus L. extract and its bioactive constituents. Phytother. Res. 2005, 19, 997–1000. [Google Scholar] [CrossRef] [PubMed]
- Bhandari, P.R. Crocus sativus L. (saffron) for cancer chemoprevention: A mini review. J. Tradit. Complement. Med. 2015, 5, 81–87. [Google Scholar] [CrossRef] [PubMed]
- Tosun, M.; Ercisli, S.; Sengul, M.; Ozer, H.; Polat, T.; Ozturk, E. Antioxidant properties and total phenolic content of eight Salvia species from Turkey. Biol. Res. 2009, 42, 175–181. [Google Scholar] [CrossRef] [PubMed]
- Katalinic, V.; Milos, M.; Kulisic, T.; Jukic, M. Screening of 70 medicinal plant extracts for antioxidant capacity and total phenols. Food Chem. 2006, 94, 550–557. [Google Scholar] [CrossRef]
- Borneo, R.; León, A.; Aguirre, A.; Ribotta, P.; Cantero, J. Antioxidant capacity of medicinal plants from the Province of Córdoba (Argentina) and their in vitro testing in a model food system. Food Chem. 2009, 112, 664–670. [Google Scholar] [CrossRef]
- Stankovic, M.S. Total phenolic content, flavonoid concentration and antioxidant activity of Marrubium peregrinum L. extracts. Kragujevac J. Sci. 2011, 33, 63–72. [Google Scholar]
- Bukhari, S.I.; Din, I.; Grewal, S.; Dhar, M.K. Antiproliferative effect of Saffron and its constituents on different cancerous cell lines. Pharmacogn. Res. 2018, 10, 291–295. [Google Scholar] [CrossRef]
- Giudetti, A.; De Domenico, S.; Ragusa, A.; Lunetti, P.; Gaballo, A.; Franck, J.; Simeone, P.; Nicolardi, G.; De Nuccio, F.; Santino, A.; et al. A specific lipid metabolic profile is associated with the epithelial mesenchymal transition program. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2019, 1864, 344–357. [Google Scholar] [CrossRef] [PubMed]
- Widholm, J.M. The use of fluorescein diacetate and phenosafranine for determining the viability of cultured plant cells. Stain. Technol. 1972, 47, 189–194. [Google Scholar] [CrossRef] [PubMed]
- Hoshyar, R.; Bathaie, S. Quantitative, HPLC analysis of monoter-pene aldehydes in different sources of Iranian saffron. In Proceedings of the 9th Iranian Congress of Biochemistry and the 2nd International Congress of Biochemistry and Molecular Biology, Shiraz, Iran, 29 October–1 November 2007; University of Shiraz: Shiraz, Iran. [Google Scholar]
- Schweiggert, U.; Kammerer, D.R.; Carle, R.; Schieber, A. Characterization of carotenoids and carotenoid esters in red pepper pods (Capsicum annuum L.) by high-performance liquid chromatography/atmospheric pressure chemical ionization mass spectrometry. Rapid Commun. Mass Spectrom. 2005, 19, 2617–2628. [Google Scholar] [CrossRef] [PubMed]
- Ranganna, S. Handbook of Analysis and Quality Control for Fruit and Vegetable Products; Tata McGraw-Hill Edu Publishing Co., Ltd.: New Delhi, India, 1986. [Google Scholar]
- Pinelo, M.; Rubilar, M.; Sineiro, J.; Nunez, M. Extraction of antioxidant phenolics from almond hulls (Prunus amygdalus) and pine sawdust (Pinus pinaster). Food Chem. 2004, 85, 267–273. [Google Scholar] [CrossRef]
- Di Sansebastiano, G.P.; Rizzello, F.; Durante, M.; Caretto, S.; Nisi, R.; De Paolis, A.; Faraco, M.; Montefusco, A.; Piro, G.; Mita, G. Subcellular compartmentalization in protoplasts from Artemisia annua cell cultures: Engineering attempts using a modified SNARE protein. J. Biotechnol. 2015, 202, 146–152. [Google Scholar] [CrossRef] [Green Version]
- Vergara, D.; Simeone, P.; Toraldo, D.; Del Boccio, P.; Vergaro, V.; Leporatti, S.; Pieragostino, D.; Tinelli, A.; De Domenico, S.; Alberti, S.; et al. Resveratrol downregulates Akt/GSK and ERK signalling pathways in OVCAR-3 ovarian cancer cells. Mol. Biosyst. 2012, 8, 1078–1087. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | I% | IC 50 (µg mL−1) | |||
|---|---|---|---|---|---|
| 20 µg mL−1 | 30 µg mL−1 | 40 µg mL−1 | 50 µg mL−1 | ||
| Ascorbic acid | 66 | 81.66 | 90.33 | 97.66 | 6.79 |
| 0.1 mock | 26.13 | 33.16 | 41.62 | 49.83 | 55.41 |
| 0.5 mock | 27.72 | 34.92 | 41.59 | 48.91 | 54.19 |
| 1 mock | 29.72 | 37.49 | 43.10 | 51.51 | 49.37 |
| 0.1 SA | 39.19 | 56.06 | 73.23 | 85.13 | 19.47 |
| 0.5 SA | 26.53 | 39.58 | 58.51 | 81.53 | 31.25 |
| 1 SA | 19.31 | 30.68 | 50.65 | 72.79 | 42.77 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moradi, A.; Zarinkamar, F.; De Domenico, S.; Mita, G.; Di Sansebastiano, G.P.; Caretto, S. Salycilic Acid Induces Exudation of Crocin and Phenolics in Saffron Suspension-Cultured Cells. Plants 2020, 9, 949. https://doi.org/10.3390/plants9080949
Moradi A, Zarinkamar F, De Domenico S, Mita G, Di Sansebastiano GP, Caretto S. Salycilic Acid Induces Exudation of Crocin and Phenolics in Saffron Suspension-Cultured Cells. Plants. 2020; 9(8):949. https://doi.org/10.3390/plants9080949
Chicago/Turabian StyleMoradi, Azar, Fatemeh Zarinkamar, Stefania De Domenico, Giovanni Mita, Gian Pietro Di Sansebastiano, and Sofia Caretto. 2020. "Salycilic Acid Induces Exudation of Crocin and Phenolics in Saffron Suspension-Cultured Cells" Plants 9, no. 8: 949. https://doi.org/10.3390/plants9080949