Evolutionary Understanding of Aquaporin Transport System in the Basal Eudicot Model Species Aquilegia coerulea

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Genome-Wide Identification of Aquaporins in Aquilegia coerulea
2.2. Classification and Phylogeny of Aquilegia coerulea Aquaporins
2.3. Gene Structure and Conserved Motif Analysis of Aquaporins in Aquilegia coerulea
2.4. Identification of Subcellular Localization and Structural Characterizations of Aquilegia Aquaporins
2.5. Protein Structure Prediction
2.6. RNA-Seq Data Analysis
3. Results
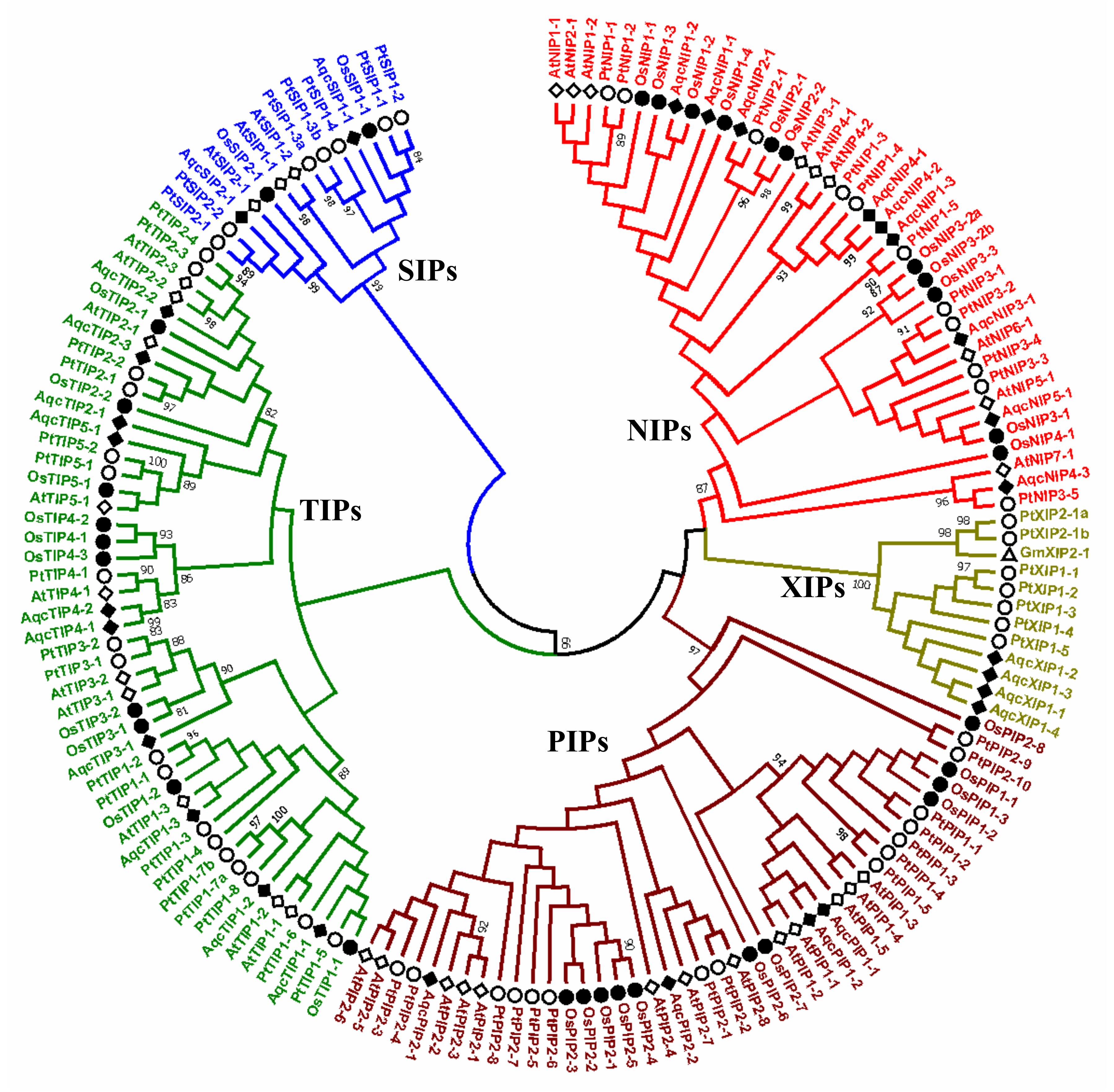
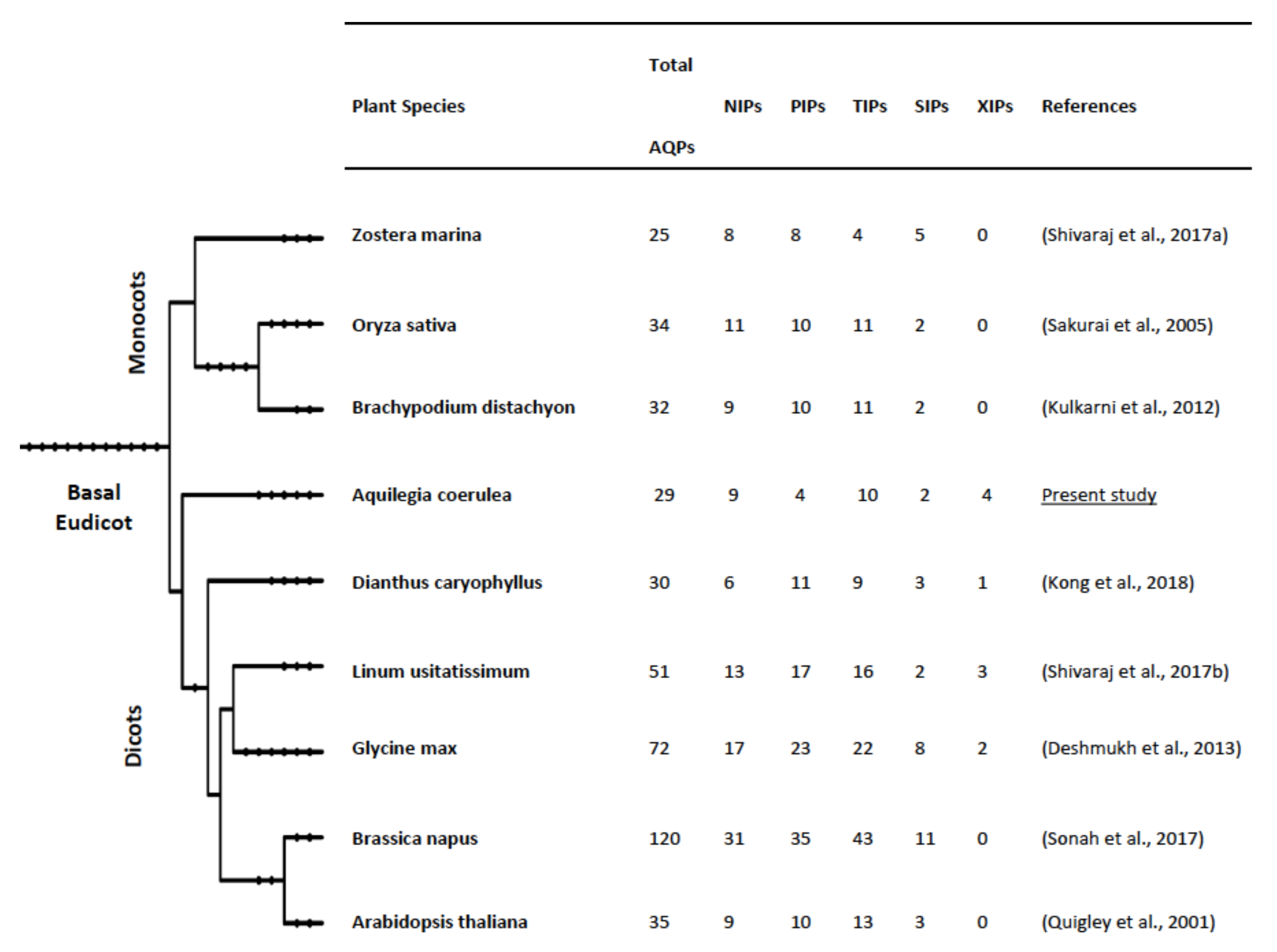
3.1. Genome-Wide Identification, Classification and Phylogenetic Distribution of Aquaporins in Aquilegia coerulea
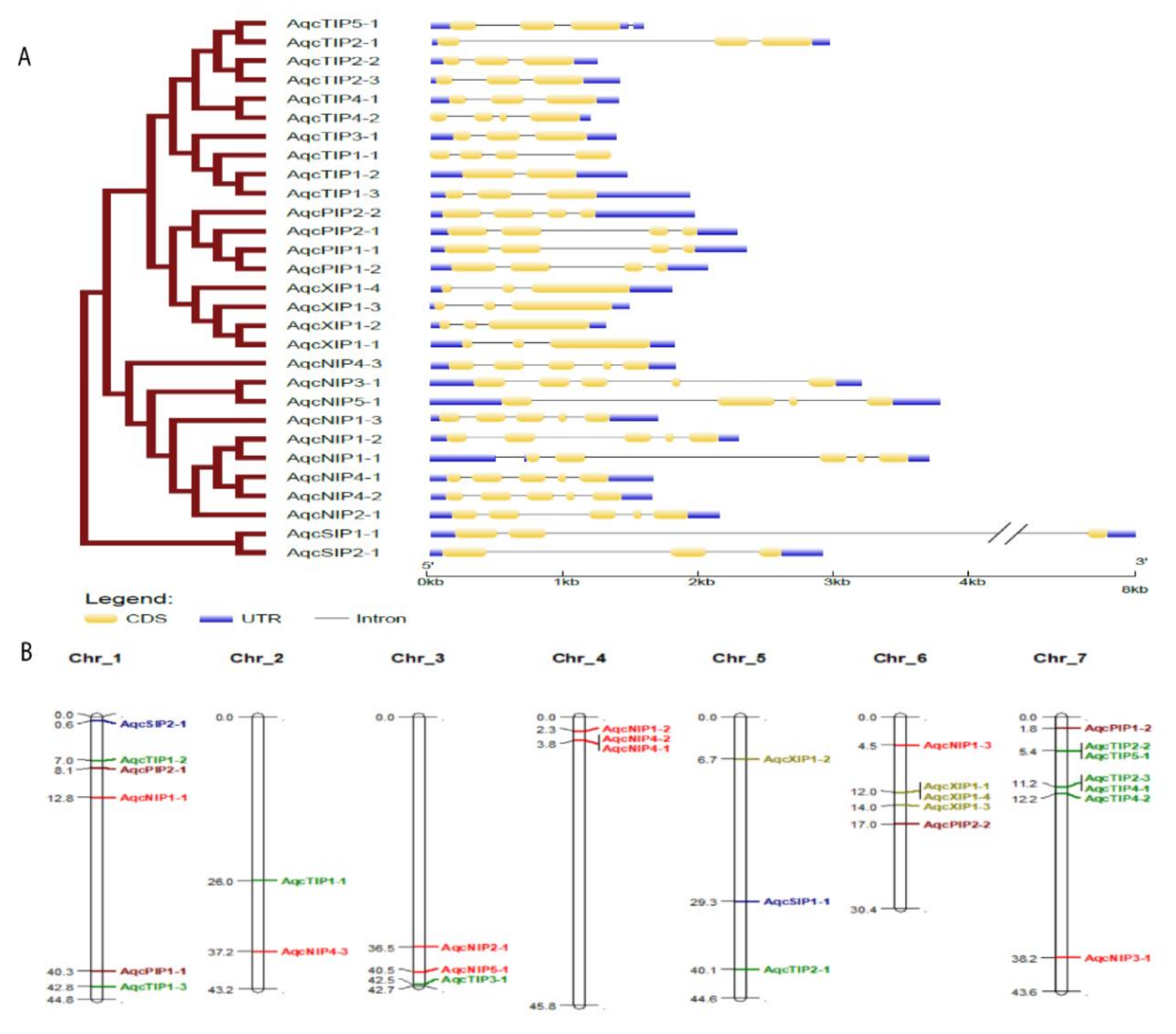
3.2. Gene Structure and Conserved Motif Analysis of Aquaporins in Aquilegia coerulea
3.3. Subcellular Localization and Exon–Intron Organization of Aquilegia Aquaporins
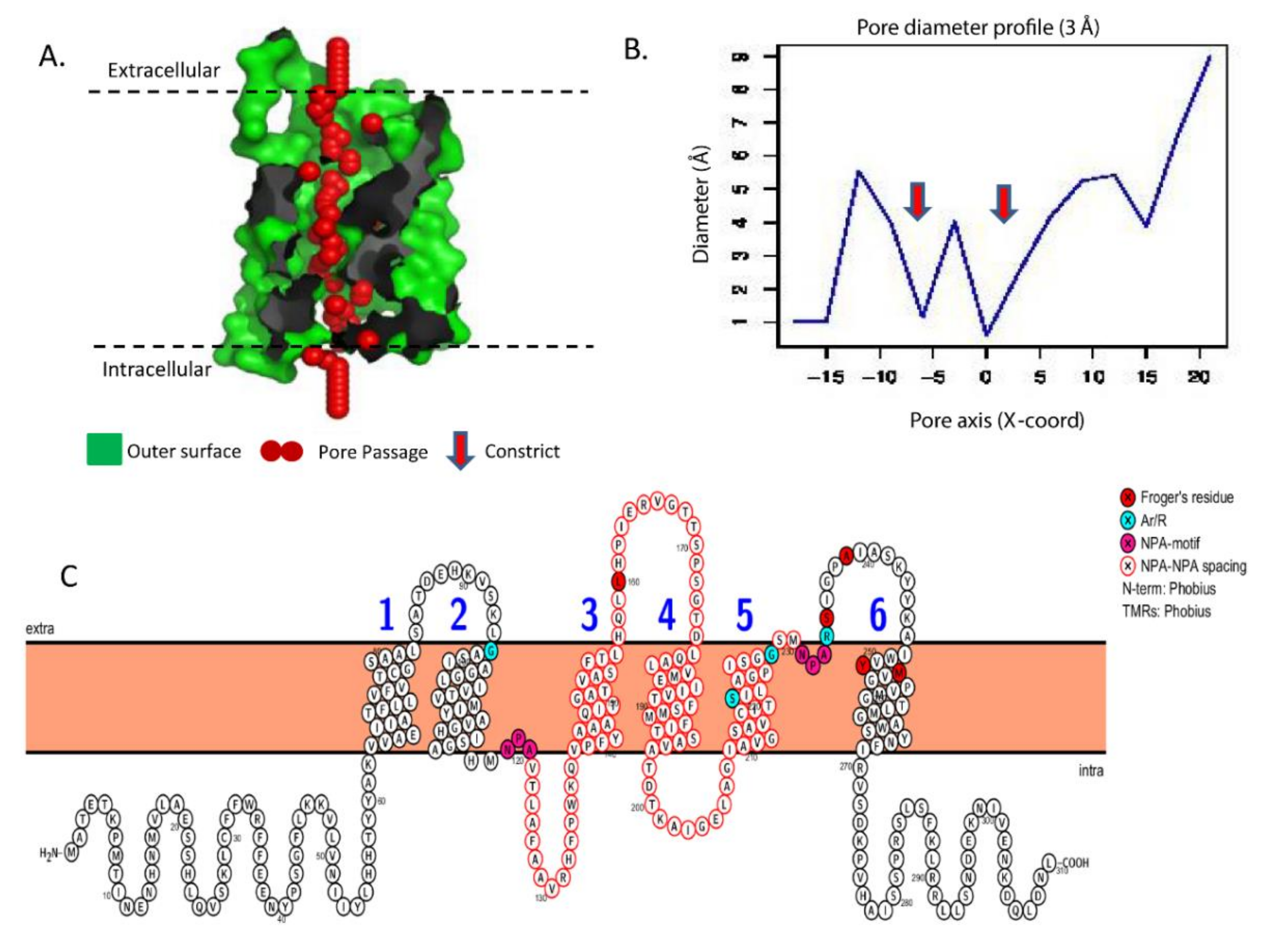
3.4. Characteristic Secondary and Tertiary Protein Structure of Aquilegia Aquaporins
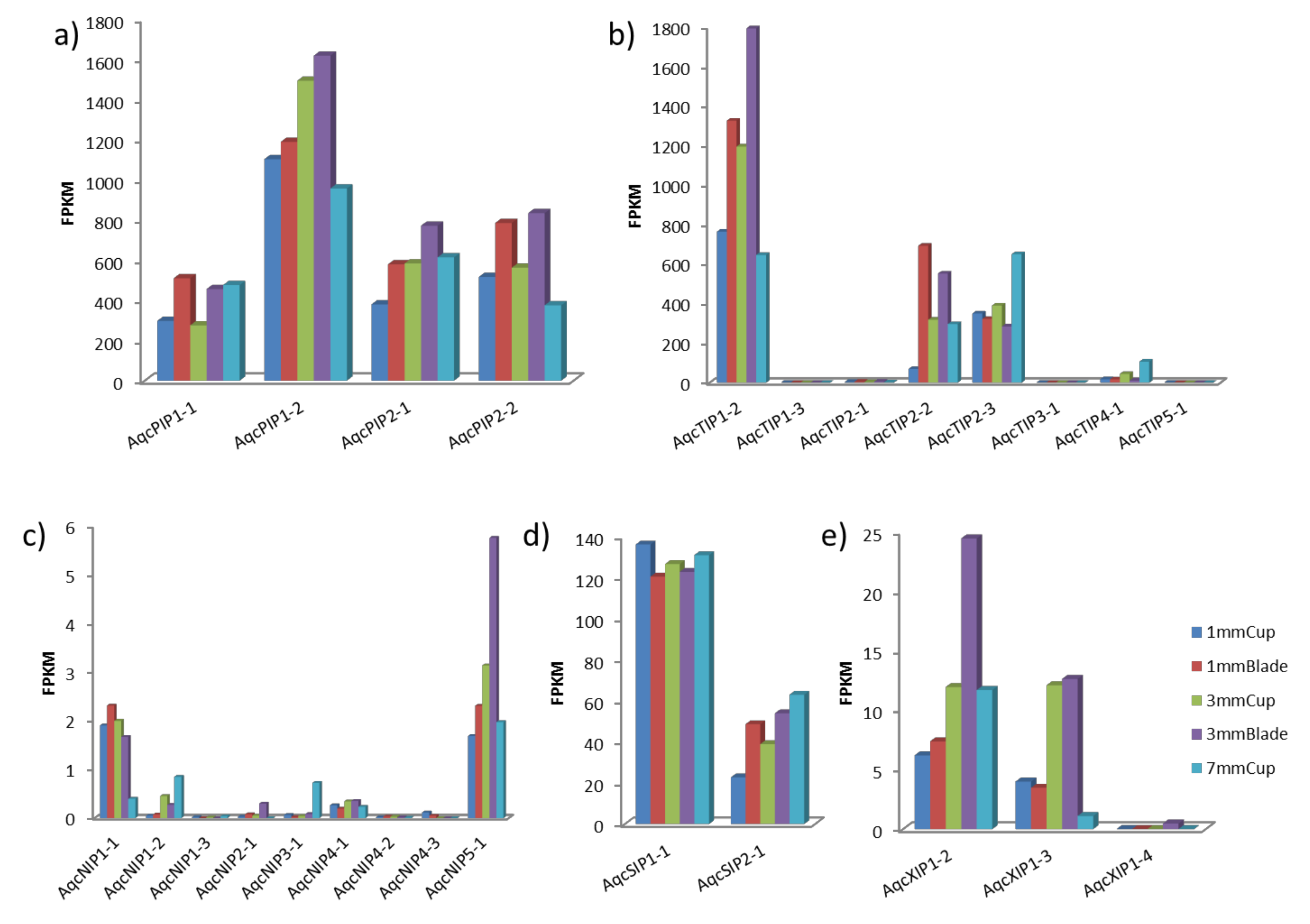
3.5. RNA-Seq Data Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Deshmukh, R.K.; Vivancos, J.; Ramakrishnan, G.; Guérin, V.; Carpentier, G.; Sonah, H.; Labbé, C.; Isenring, P.; Belzile, F.J.; Bélanger, R.R. A precise spacing between the NPA domains of aquaporins is essential for silicon permeability in plants. Plant J. 2015, 83, 489–500. [Google Scholar] [CrossRef] [PubMed]
- Hove, R.M.; Bhave, M. Plant aquaporins with non-aqua functions: Deciphering the signature sequences. Plant Mol. Biol. 2011, 75, 413–430. [Google Scholar] [CrossRef] [PubMed]
- Maurel, C.; Javot, H.; Lauvergeat, V.; Gerbeau, P.; Tournaire, C.; Santoni, V.; Heyes, J. Molecular physiology of aquaporins in plants. In International Review of Cytology; Elsevier: Amsterdam, The Netherlands, 2002; Volume 215, pp. 105–148. [Google Scholar]
- Deshmukh, R.K.; Nguyen, H.T.; Belanger, R.R. Editorial: Aquaporins: Dynamic Role and Regulation. Front. Plant Sci. 2017, 8, 1420. [Google Scholar] [CrossRef] [PubMed]
- Laloux, T.; Junqueira, B.; Maistriaux, L.C.; Ahmed, J.; Jurkiewicz, A.; Chaumont, F. Plant and Mammal Aquaporins: Same but Different. Int. J. Mol. Sci. 2018, 19, 521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Preston, G.M.; Agre, P. Isolation of the cDNA for erythrocyte integral membrane protein of 28 kilodaltons: Member of an ancient channel family. Proc. Natl. Acad. Sci. USA 1991, 88, 11110–11114. [Google Scholar] [CrossRef] [Green Version]
- Preston, G.M.; Carroll, T.P.; Guggino, W.B.; Agre, P. Appearance of water channels in Xenopus oocytes expressing red cell CHIP28 protein. Science 1992, 256, 385–387. [Google Scholar] [CrossRef] [Green Version]
- Papadopoulos, M.C.; Verkman, A.S. Aquaporin water channels in the nervous system. Nat. Rev. Neurosci. 2013, 14, 265. [Google Scholar] [CrossRef] [Green Version]
- Benga, O.; Huber, V.J. Brain water channel proteins in health and disease. Mol. Asp. Med. 2012, 33, 562–578. [Google Scholar] [CrossRef]
- King, L.S.; Kozono, D.; Agre, P. From structure to disease: The evolving tale of aquaporin biology. Nat. Rev. Mol. Cell Biol. 2004, 5, 687. [Google Scholar] [CrossRef]
- Singh, R.K.; Deshmukh, R.; Muthamilarasan, M.; Rani, R.; Prasad, M. Versatile roles of aquaporin in physiological processes and stress tolerance in plants. Plant Physiol. Biochem. 2020, 149, 178–189. [Google Scholar] [CrossRef]
- Deshmukh, R.K.; Sonah, H.; Bélanger, R.R. Plant Aquaporins: Genome-wide identification, transcriptomics, proteomics, and advanced analytical tools. Front. Plant Sci. 2016, 7, 1896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sonah, H.; Deshmukh, R.K.; Labbé, C.; Bélanger, R.R. Analysis of aquaporins in Brassicaceae species reveals high-level of conservation and dynamic role against biotic and abiotic stress in canola. Sci. Rep. 2017, 7, 2771. [Google Scholar] [CrossRef] [PubMed]
- Quigley, F.; Rosenberg, J.M.; Shachar-Hill, Y.; Bohnert, H.J. From genome to function: The Arabidopsis aquaporins. Genome Biol. 2001, 3, 1. [Google Scholar] [CrossRef] [PubMed]
- Vishwakarma, K.; Mishra, M.; Patil, G.; Mulkey, S.; Ramawat, N.; Singh V, P.; Deshmukh, R.; Kumar Tripathi, D.K.; Nguyen, H.T.; Sharma, S. Avenues of the membrane transport system in adaptation of plants to abiotic stresses. Crit. Rev. Biotech. 2019, 39, 861–883. [Google Scholar] [CrossRef]
- Guan, X.G.; Su, W.H.; Yi, F.; Zhang, D.; Hao, F.; Zhang, H.G.; Liu, Y.J.; Feng, X.C.; Ma, T.H. NPA motifs play a key role in plasma membrane targeting of aquaporin-4. IUBMB Life 2010, 62, 222–226. [Google Scholar] [CrossRef]
- Santoni, V.; Joëlle, V.; Pflieger, D.; Sommerer, N.; Maurel, C. A proteomic study reveals novel insights into the diversity of aquaporin forms expressed in the plasma membrane of plant roots. Biochem. J. 2003, 373, 289–296. [Google Scholar] [CrossRef]
- Shivaraj, S.; Deshmukh, R.; Sonah, H.; Bélanger, R.R. Identification and characterization of aquaporin genes in Arachis duranensis and Arachis ipaensis genomes, the diploid progenitors of peanut. BMC Genom. 2019, 20, 222. [Google Scholar] [CrossRef] [Green Version]
- Maurel, C.; Boursiac, Y.; Luu, D.-T.; Santoni, V.; Shahzad, Z.; Verdoucq, L. Aquaporins in plants. Physiol. Rev. 2015, 95, 1321–1358. [Google Scholar] [CrossRef]
- Vera-Estrella, R.; Barkla, B.J.; Amezcua-Romero, J.C.; Pantoja, O. Day/night regulation of aquaporins during the CAM cycle in Mesembryanthemum crystallinum. Plant Cell Environ. 2012, 35, 485–501. [Google Scholar] [CrossRef]
- Törnroth-Horsefield, S.; Wang, Y.; Hedfalk, K.; Johanson, U.; Karlsson, M.; Tajkhorshid, E.; Neutze, R.; Kjellbom, P. Structural mechanism of plant aquaporin gating. Nature 2006, 439, 688–694. [Google Scholar] [CrossRef]
- Eriksson, U.K.; Fischer, G.; Friemann, R.; Enkavi, G.; Tajkhorshid, E.; Neutze, R. Subangstrom resolution X-ray structure details aquaporin-water interactions. Science 2013, 340, 1346–1349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kramer, E.M. Aquilegia: A new model for plant development, ecology, and evolution. Annu. Rev. Plant Biol. 2009, 60, 261–277. [Google Scholar] [CrossRef] [PubMed]
- Filiault, D.L.; Ballerini, E.S.; Mandáková, T.; Aköz, G.; Derieg, N.J.; Schmutz, J.; Jenkins, J.; Grimwood, J.; Shu, S.; Hayes, R.D. The Aquilegia genome provides insight into adaptive radiation and reveals an extraordinarily polymorphic chromosome with a unique history. eLife 2018, 7, e36426. [Google Scholar] [CrossRef] [PubMed]
- Brunet, J.; Eckert, C. Effects of floral morphology and display on outcrossing in blue columbine, Aquilegia caerulea (Ranunculaceae). Funct. Ecol. 1998, 12, 596–606. [Google Scholar] [CrossRef]
- Miller, R.B. Hawkmoths and the geographic patterns of floral variation in Aquilegia caerulea. Evolution 1981, 35, 763–774. [Google Scholar] [CrossRef]
- Sharma, B.; Kramer, E.M. The MADS-box gene family of the basal eudicot and hybrid Aquilegia coerulea ‘Origami’ (Ranunculaceae). Ann. Mo. Bot. Gard. 2014, 99, 313–322. [Google Scholar] [CrossRef]
- Thairu, M.W.; Brunet, J. The role of pollinators in maintaining variation in flower colour in the Rocky Mountain columbine, Aquilegia coerulea. Ann. Bot. 2015, 115, 971–979. [Google Scholar] [CrossRef] [Green Version]
- Yant, L.; Collani, S.; Puzey, J.; Levy, C.; Kramer, E.M. Molecular basis for three-dimensional elaboration of the Aquilegia petal spur. Proc. R. Soc. Lond. B Biol. Sci. 2015, 282, 20142778. [Google Scholar] [CrossRef] [Green Version]
- Higuchi, T.; Suga, S.; Tsuchiya, T.; Hisada, H.; Morishima, S.; Okada, Y.; Maeshima, M. Molecular cloning, water channel activity and tissue specific expression of two isoforms of radish vacuolar aquaporin1. Plant Cell Physiol. 1998, 39, 905–913. [Google Scholar] [CrossRef] [Green Version]
- Ishibashi, K.; Kondo, S.; Hara, S.; Morishita, Y. The evolutionary aspects of aquaporin family. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2010, 300, R566–R576. [Google Scholar] [CrossRef]
- Siefritz, F.; Otto, B.; Bienert, G.P.; Van Der Krol, A.; Kaldenhoff, R. The plasma membrane aquaporin NtAQP1 is a key component of the leaf unfolding mechanism in tobacco. Plant J. 2004, 37, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Martre, P.; Morillon, R.; Barrieu, F.; North, G.B.; Nobel, P.S.; Chrispeels, M.J. Plasma membrane aquaporins play a significant role during recovery from water deficit. Plant Physiol. 2002, 130, 2101–2110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, A.B.; Sankararamakrishnan, R. Genome-wide analysis of major intrinsic proteins in the tree plant Populus trichocarpa: Characterization of XIP subfamily of aquaporins from evolutionary perspective. BMC Plant Biol. 2009, 9, 134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goodstein, D.M.; Shu, S.; Howson, R.; Neupane, R.; Hayes, R.D.; Fazo, J.; Mitros, T.; Dirks, W.; Hellsten, U.; Putnam, N. Phytozome: A comparative platform for green plant genomics. Nucleic Acids Res. 2011, 40, D1178–D1186. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. In Nucleic Acids Symposium Series; Information Retrieval Ltd.: London, UK, 1999; pp. 95–98. [Google Scholar]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Bernsel, A.; Viklund, H.; Hennerdal, A.; Elofsson, A. TOPCONS: Consensus prediction of membrane protein topology. Nucleic Acids Res. 2009, 37, W465–W468. [Google Scholar] [CrossRef] [Green Version]
- Bailey, T.L.; Williams, N.; Misleh, C.; Li, W.W. MEME: Discovering and analyzing DNA and protein sequence motifs. Nucleic Acids Res. 2006, 34, W369–W373. [Google Scholar] [CrossRef]
- Emanuelsson, O.; Nielsen, H.; Brunak, S.; Von Heijne, G. Predicting subcellular localization of proteins based on their N-terminal amino acid sequence. J. Mol. Biol. 2000, 300, 1005–1016. [Google Scholar] [CrossRef] [Green Version]
- Horton, P.; Park, K.-J.; Obayashi, T.; Fujita, N.; Harada, H.; Adams-Collier, C.; Nakai, K. WoLF PSORT: Protein localization predictor. Nucleic Acids Res. 2007, 35, W585–W587. [Google Scholar] [CrossRef] [Green Version]
- Yu, C.S.; Lin, C.J.; Hwang, J.K. Predicting subcellular localization of proteins for Gram-negative bacteria by support vector machines based on n-peptide compositions. Protein Sci. 2004, 13, 1402–1406. [Google Scholar] [CrossRef] [Green Version]
- Guo, A.; Zhu, Q.; Chen, X.; Luo, J.C. GSDS: A gene structure display server. Yi Chuan= Hered. 2007, 29, 1023–1026. [Google Scholar] [CrossRef]
- Voorrips, R.E. MapChart: Software for the graphical presentation of linkage maps and QTLs. J. Hered. 2002, 93, 77–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Omasits, U.; Ahrens, C.H.; Müller, S.; Wollscheid, B. Protter: Interactive protein feature visualization and integration with experimental proteomic data. Bioinformatics 2013, 30, 884–886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kelley, L.A.; Mezulis, S.; Yates, C.M.; Wass, M.N.; Sternberg, M.J.E. The Phyre2 web portal for protein modeling, prediction and analysis. Nat. Protoc. 2015, 10, 845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berka, K.; Hanák, O.; Sehnal, D.; Banáš, P.; Navratilova, V.; Jaiswal, D.; Ionescu, C.-M.; Svobodová Vařeková, R.; Koča, J.; Otyepka, M. MOLE online 2.0: Interactive web-based analysis of biomacromolecular channels. Nucleic Acids Res. 2012, 40, W222–W227. [Google Scholar] [CrossRef] [Green Version]
- Pellegrini-Calace, M.; Maiwald, T.; Thornton, J.M. PoreWalker: A novel tool for the identification and characterization of channels in transmembrane proteins from their three-dimensional structure. PLoS Comput. Biol. 2009, 5, e1000440. [Google Scholar] [CrossRef] [Green Version]
- Patil, G.; Valliyodan, B.; Deshmukh, R.; Prince, S.; Nicander, B.; Zhao, M.; Sonah, H.; Song, L.; Lin, L.; Chaudhary, J. Soybean (Glycine max) SWEET gene family: Insights through comparative genomics, transcriptome profiling and whole genome re-sequence analysis. BMC Genom. 2015, 16, 1. [Google Scholar] [CrossRef] [Green Version]
- Shivaraj, S.M.; Deshmukh, R.; Bhat, J.A.; Sonah, H.; Bélanger, R.R. Understanding Aquaporin Transport System in Eelgrass (Zostera marina L.), an Aquatic Plant Species. Front. Plant Sci. 2017, 8. [Google Scholar] [CrossRef] [Green Version]
- Danielson, J.Å.; Johanson, U. Unexpected complexity of the aquaporin gene family in the moss Physcomitrella patens. BMC Plant Biol. 2008, 8, 45. [Google Scholar] [CrossRef] [Green Version]
- Guerriero, G.; Deshmukh, R.; Sonah, H.; Sergeant, K.; Hausman, J.-F.; Lentzen, E.; Valle, N.; Siddiqui, K.S.; Exley, C. Identification of the aquaporin gene family in Cannabis sativa and evidence for the accumulation of silicon in its tissues. Plant Sci. 2019, 287, 110167. [Google Scholar] [CrossRef]
- Bhat, J.A.; Shivaraj, S.; Singh, P.; Navadagi, D.B.; Tripathi, D.K.; Dash, P.K.; Solanke, A.U.; Sonah, H.; Deshmukh, R. Role of silicon in mitigation of heavy metal stresses in crop plants. Plants 2019, 8, 71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deshmukh, R.K.; Ma, J.F.; Bélanger, R.R. Role of silicon in plants. Front. Plant Sci. 2017, 8, 1858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Majeed Zargar, S.; Nazir, M.; Kumar Agrawal, G.; Kim, D.-W.; Rakwal, R. Silicon in plant tolerance against environmental stressors: Towards crop improvement using omics approaches. Curr. Proteom. 2010, 7, 135–143. [Google Scholar] [CrossRef]
- Vivancos, J.; Deshmukh, R.; Grégoire, C.; Rémus-Borel, W.; Belzile, F.; Bélanger, R.R. Identification and characterization of silicon efflux transporters in horsetail (Equisetum arvense). J. Plant Physiol. 2016, 200, 82–89. [Google Scholar] [CrossRef]
- Grégoire, C.; Rémus-Borel, W.; Vivancos, J.; Labbé, C.; Belzile, F.; Bélanger, R.R. Discovery of a multigene family of aquaporin silicon transporters in the primitive plant Equisetum arvense. Plant J. 2012, 72, 320–330. [Google Scholar] [CrossRef]
- Coskun, D.; Deshmukh, R.; Sonah, H.; Menzies, J.G.; Reynolds, O.; Ma, J.F.; Kronzucker, H.J.; Bélanger, R.R. The controversies of silicon’s role in plant biology. New Phytol. 2018. [Google Scholar] [CrossRef]
- Deshmukh, R.; Bélanger, R.R. Molecular evolution of aquaporins and silicon influx in plants. Funct. Ecol. 2016, 30, 1277–1285. [Google Scholar] [CrossRef]
- Shivaraj, S.; Deshmukh, R.K.; Rai, R.; Bélanger, R.; Agrawal, P.K.; Dash, P.K. Genome-wide identification, characterization, and expression profile of aquaporin gene family in flax (Linum usitatissimum). Sci. Rep. 2017, 7, 46137. [Google Scholar] [CrossRef] [Green Version]
- Zargar, S.M.; Nagar, P.; Deshmukh, R.; Nazir, M.; Wani, A.A.; Masoodi, K.Z.; Agrawal, G.K.; Rakwal, R. Aquaporins as potential drought tolerance inducing proteins: Towards instigating stress tolerance. J. Proteom. 2017, 169, 233–238. [Google Scholar] [CrossRef]
- Deshmukh, R.K.; Sonah, H.; Singh, N.K. Intron gain, a dominant evolutionary process supporting high levels of gene expression in rice. J. Plant Biochem. Biotechnol. 2016, 25, 142–146. [Google Scholar] [CrossRef]
- Kozlowski, L.P. IPC–isoelectric point calculator. Biol. Direct 2016, 11, 55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puzey, J.R.; Gerbode, S.J.; Hodges, S.A.; Kramer, E.M.; Mahadevan, L. Evolution of spur-length diversity in Aquilegia petals is achieved solely through cell-shape anisotropy. Proc. R. Soc. Lond. B Biol. Sci. 2012, 279, 1640–1645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uehlein, N.; Fileschi, K.; Eckert, M.; Bienert, G.P.; Bertl, A.; Kaldenhoff, R. Arbuscular mycorrhizal symbiosis and plant aquaporin expression. Phytochemistry 2007, 68, 122–129. [Google Scholar] [CrossRef] [PubMed]
- Ludevid, D.; Höfte, H.; Himelblau, E.; Chrispeels, M.J. The expression pattern of the tonoplast intrinsic protein γ-TIP in Arabidopsis thaliana is correlated with cell enlargement. Plant Physiol. 1992, 100, 1633–1639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suga, S.; Maeshima, M. Water channel activity of radish plasma membrane aquaporins heterologously expressed in yeast and their modification by site-directed mutagenesis. Plant Cell Physiol. 2004, 45, 823–830. [Google Scholar] [CrossRef]
- Eisenbarth, D.A.; Weig, A.R. Dynamics of aquaporins and water relations during hypocotyl elongation in Ricinus communis L. seedlings. J. Exp. Bot. 2005, 56, 1831–1842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barrieu, F.; Chaumont, F.; Chrispeels, M.J. High Expression of the Tonoplast Aquaporin ZmTIP1in Epidermal and Conducting Tissues of Maize. Plant Physiol. 1998, 117, 1153–1163. [Google Scholar] [CrossRef] [Green Version]
- Chaumont, F.; Barrieu, F.; Herman, E.M.; Chrispeels, M.J. Characterization of a maize tonoplast aquaporin expressed in zones of cell division and elongation. Plant Physiol. 1998, 117, 1143–1152. [Google Scholar] [CrossRef] [Green Version]
- Kong, W.; Bendahmane, M.; Fu, X. Genome-wide identification and characterization of aquaporins and their role in the flower opening processes in carnation (Dianthus caryophyllus). Molecules 2018, 23, 1895. [Google Scholar] [CrossRef] [Green Version]
- Azad, A.K.; Sawa, Y.; Ishikawa, T.; Shibata, H. Phosphorylation of plasma membrane aquaporin regulates temperature-dependent opening of tulip petals. Plant Cell Physiol. 2004, 45, 608–617. [Google Scholar] [CrossRef] [Green Version]
- Azad, A.K.; Katsuhara, M.; Sawa, Y.; Ishikawa, T.; Shibata, H. Characterization of four plasma membrane aquaporins in tulip petals: A putative homolog is regulated by phosphorylation. Plant Cell Physiol. 2008, 49, 1196–1208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, N.; Xue, J.; Li, Y.; Liu, X.; Dai, F.; Jia, W.; Luo, Y.; Gao, J. Rh-PIP2; 1, a rose aquaporin gene, is involved in ethylene-regulated petal expansion. Plant Physiol. 2008, 148, 894–907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Q.; Chen, H.; He, M.; Zhao, Z.; Cai, H.; Ding, G.; Shi, L.; Xu, F. The boron transporter BnaC4. BOR1; 1c is critical for inflorescence development and fertility under boron limitation in Brassica napus. Plant Cell Environ. 2017, 40, 1819–1833. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID | NPA Motifs | NPA Spacing | Ar/R Selectivity Filters | Froger’s Residues | Mitani’s Residue | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| NPA (LB) | NPA (LE) | H2 | H5 | LE1 | LE2 | P1 | P2 | P3 | P4 | P5 | |||
| AqcNIP1-1 | NPA | NPA | 109 | W | V | A | R | F | S | A | Y | I | P |
| AqcNIP1-2 | NPA | NPA | 109 | W | V | A | R | F | S | A | Y | I | P |
| AqcNIP1-3 | NPA | NPA | 112 | W | V | S | R | Y | S | A | Y | L | P |
| AqcNIP2-1 | NPA | NPA | 108 | G | S | G | R | L | S | A | Y | M | P |
| AqcNIP3-1 | NPA | NPA | 108 | T | I | A | R | F | S | A | Y | L | P |
| AqcNIP4-1 | NPA | NPA | 109 | W | V | A | R | F | S | A | Y | M | P |
| AqcNIP4-3 | NPS | NPA | 108 | G | I | G | R | Y | S | A | Y | I | P |
| AqcNIP4-2 | NPA | NPA | 109 | W | V | A | R | F | S | A | Y | I | P |
| AqcNIP5-1 | NPS | NPV | 108 | A | I | G | R | F | T | A | Y | L | P |
| AqcPIP1-1 | NPA | NPA | 118 | F | H | T | R | Q | S | A | H | W | W |
| AqcPIP1-2 | NPA | NPA | 118 | F | H | T | R | Q | S | A | H | W | W |
| AqcPIP2-1 | NPA | NPA | 118 | F | H | T | R | Q | S | A | Q | W | W |
| AqcPIP2-2 | NPA | NPA | 118 | F | H | T | R | M | S | A | H | W | W |
| AqcSIP1-1 | NPT | NPA | 22 | V | G | L | A | Q | I | G | E | Y | Q |
| AqcSIP2-1 | NPL | NPA | 40 | S | A | L | K | L | Q | V | E | E | E |
| AqcTIP1-2 | NPA | NPA | 111 | H | I | A | V | T | A | A | Y | W | P |
| AqcTIP1-1 | NPA | NPA | 130 | A | I | A | V | T | S | A | Y | W | P |
| AqcTIP1-3 | NPA | NPA | 111 | H | I | A | V | T | S | A | Y | W | P |
| AqcTIP2-1 | NPA | NPA | 110 | H | I | G | R | T | S | A | Y | W | P |
| AqcTIP2-3 | NPA | NPA | 111 | H | I | G | R | T | S | A | Y | W | P |
| AqcTIP2-2 | NPA | NPA | 111 | H | I | G | R | T | S | A | Y | W | P |
| AqcTIP3-1 | NPA | NPA | 111 | H | I | A | R | T | A | A | Y | W | P |
| AqcTIP4-1 | NPA | NPA | 111 | H | I | A | R | T | S | S | Y | W | P |
| AqcTIP4-2 | NPA | NPA | 96 | H | I | A | R | S | S | S | Y | W | P |
| AqcTIP5-1 | NPA | NPA | 110 | S | V | G | C | T | S | A | Y | W | P |
| AqcXIP1-2 | NPV | NPA | 135 | I | V | V | R | V | C | A | W | V | V |
| AqcXIP1-1 | NPV | HPA | 135 | V | V | V | R | I | C | A | W | I | V |
| AqcXIP1-4 | NPT | NPA | 134 | V | V | V | R | V | C | T | W | I | V |
| AqcXIP1-3 | NPA | NPA | 135 | I | V | V | R | V | C | A | W | V | V |
| Aquaporins | Length(Å) | Hydropathy | Bottleneck | Charge | Polarity | Mutability | LogP | LogD | LogS | Ionizable |
|---|---|---|---|---|---|---|---|---|---|---|
| AqcNIP1-1 | 56.40 | 0.63 | 0.80 | 1.00 | 9.26 | 91.00 | 0.61 | 0.35 | −0.19 | 3.00 |
| AqcNIP1-2 | 49.80 | 0.98 | 1.00 | 2.00 | 7.05 | 84.00 | 0.74 | 0.54 | −0.39 | 4.00 |
| AqcNIP1-3 | 38.80 | 1.08 | 0.30 | 1.00 | 6.10 | 91.00 | 0.65 | 0.43 | −0.22 | 3.00 |
| AqcNIP2-1 | 57.40 | −0.03 | 1.20 | 2.00 | 9.50 | 91.00 | 0.27 | 0.00 | 0.32 | 4.00 |
| AqcNIP3-1 | 51.40 | 0.08 | 1.30 | 1.00 | 6.02 | 91.00 | 0.26 | 0.06 | 0.03 | 1.00 |
| AqcNIP4-1 | 57.60 | 0.31 | 0.60 | −1.00 | 10.39 | 87.00 | 0.47 | 0.12 | 0.02 | 5.00 |
| AqcNIP4-3 | 38.70 | 0.37 | −0.10 | −2.00 | 8.08 | 90.00 | 0.17 | −0.16 | 0.34 | 2.00 |
| AqcNIP4-2 | 84.70 | 0.16 | 0.70 | −1.00 | 6.68 | 91.00 | 0.30 | 0.10 | 0.15 | 3.00 |
| AqcNIP5-1 | 46.00 | 0.98 | 0.20 | −1.00 | 4.22 | 86.00 | 0.59 | 0.46 | −0.03 | 1.00 |
| AqcPIP1-1 | 83.20 | 0.38 | 1.10 | 0.00 | 11.40 | 82.00 | 0.70 | 0.30 | −0.17 | 8.00 |
| AqcPIP1-2 | 35.50 | 0.39 | 0.20 | −1.00 | 5.21 | 81.00 | 0.44 | 0.39 | −0.12 | 1.00 |
| AqcPIP2-1 | 38.40 | 0.14 | 0.60 | 1.00 | 8.44 | 83.00 | 0.90 | 0.71 | −0.46 | 3.00 |
| AqcPIP2-2 | 74.90 | 0.22 | 0.00 | 1.00 | 8.60 | 89.00 | 0.66 | 0.34 | −0.14 | 3.00 |
| AqcSIP1-1 | 45.50 | −0.85 | 0.00 | −1.00 | 3.89 | 81.00 | 0.30 | 0.28 | −0.09 | 1.00 |
| AqcSIP2-1 | 43.40 | −1.83 | 0.40 | 8.00 | 18.48 | 85.00 | N/A | −0.73 | 0.62 | 8.00 |
| AqcTIP1-2 | 34.40 | 1.09 | 1.20 | 2.00 | 7.19 | 82.00 | 0.93 | 0.79 | −0.64 | 2.00 |
| AqcTIP1-1 | 47.30 | 1.21 | 1.00 | 0.00 | 4.79 | 92.00 | 0.61 | 0.55 | −0.28 | 2.00 |
| AqcTIP1-3 | 53.20 | 1.11 | 0.50 | 1.00 | 6.09 | 85.00 | 0.58 | 0.50 | −0.23 | 3.00 |
| AqcTIP2-1 | 48.70 | −0.10 | 0.00 | 1.00 | 10.98 | 85.00 | 0.32 | −0.02 | 0.07 | 3.00 |
| AqcTIP2-3 | 61.80 | 0.54 | 0.50 | 2.00 | 8.66 | 86.00 | 0.49 | 0.34 | −0.21 | 2.00 |
| AqcTIP2-2 | 44.10 | 0.89 | 0.50 | 2.00 | 8.70 | 84.00 | 0.58 | 0.39 | −0.29 | 2.00 |
| AqcTIP3-1 | 58.50 | 0.81 | 0.50 | 3.00 | 8.74 | 86.00 | 0.62 | 0.42 | −0.25 | 5.00 |
| AqcTIP4-1 | 60.70 | 0.36 | 0.60 | −1.00 | 9.79 | 87.00 | 0.47 | 0.14 | −0.01 | 5.00 |
| AqcTIP4-2 | 98.20 | −0.04 | 0.70 | −2.00 | 8.54 | 90.00 | 0.33 | 0.04 | 0.20 | 6.00 |
| AqcTIP5-1 | 60.10 | 0.67 | 0.60 | 1.00 | 4.05 | 84.00 | 0.32 | 0.28 | −0.09 | 1.00 |
| AqcXIP1-2 | 94.10 | 0.24 | 1.00 | 4.00 | 10.58 | 84.00 | 0.59 | 0.12 | −0.04 | 6.00 |
| AqcXIP1-1 | 55.70 | 0.60 | 0.70 | 1.00 | 9.16 | 86.00 | 0.81 | 0.60 | −0.35 | 3.00 |
| AqcXIP1-4 | 70.00 | −0.31 | 0.60 | 4.00 | 10.05 | 86.00 | 0.44 | 0.03 | 0.04 | 4.00 |
| AqcXIP1-3 | 47.20 | 0.00 | 0.20 | 1.00 | 7.66 | 89.00 | 0.23 | 0.06 | 0.12 | 1.00 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Singh, S.; Bhatt, V.; Kumar, V.; Kumawat, S.; Khatri, P.; Singla, P.; Shivaraj, S.M.; Nadaf, A.; Deshmukh, R.; Sharma, T.R.; et al. Evolutionary Understanding of Aquaporin Transport System in the Basal Eudicot Model Species Aquilegia coerulea. Plants 2020, 9, 799. https://doi.org/10.3390/plants9060799
Singh S, Bhatt V, Kumar V, Kumawat S, Khatri P, Singla P, Shivaraj SM, Nadaf A, Deshmukh R, Sharma TR, et al. Evolutionary Understanding of Aquaporin Transport System in the Basal Eudicot Model Species Aquilegia coerulea. Plants. 2020; 9(6):799. https://doi.org/10.3390/plants9060799
Chicago/Turabian StyleSingh, Shweta, Vacha Bhatt, Virender Kumar, Surbhi Kumawat, Praveen Khatri, Pankaj Singla, S.M. Shivaraj, Altaf Nadaf, Rupesh Deshmukh, Tilak Raj Sharma, and et al. 2020. "Evolutionary Understanding of Aquaporin Transport System in the Basal Eudicot Model Species Aquilegia coerulea" Plants 9, no. 6: 799. https://doi.org/10.3390/plants9060799