Assembly and Analysis of the Complete Mitochondrial Genome of Capsella bursa-pastoris

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
2.1. Sequencing and Assembly of the Complete Mitogenome of C. bursa-pastoris
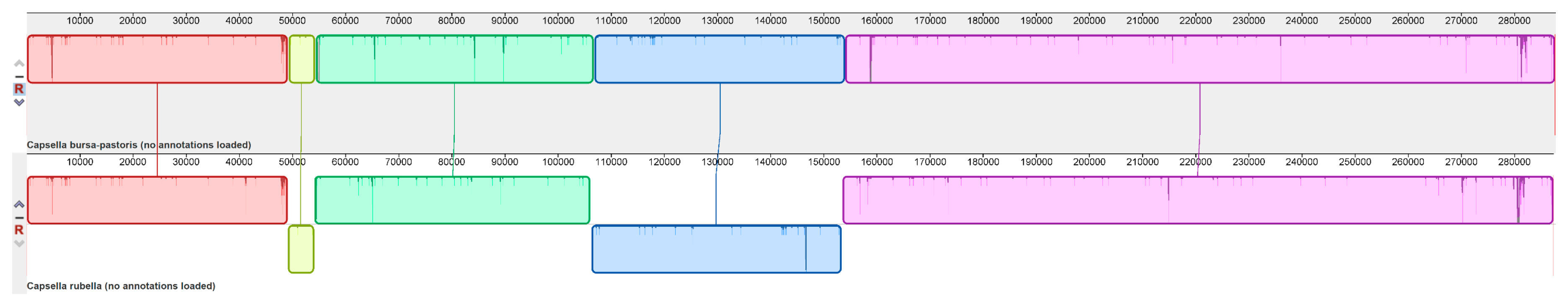
2.2. Repeats and Structural Variation Analysis
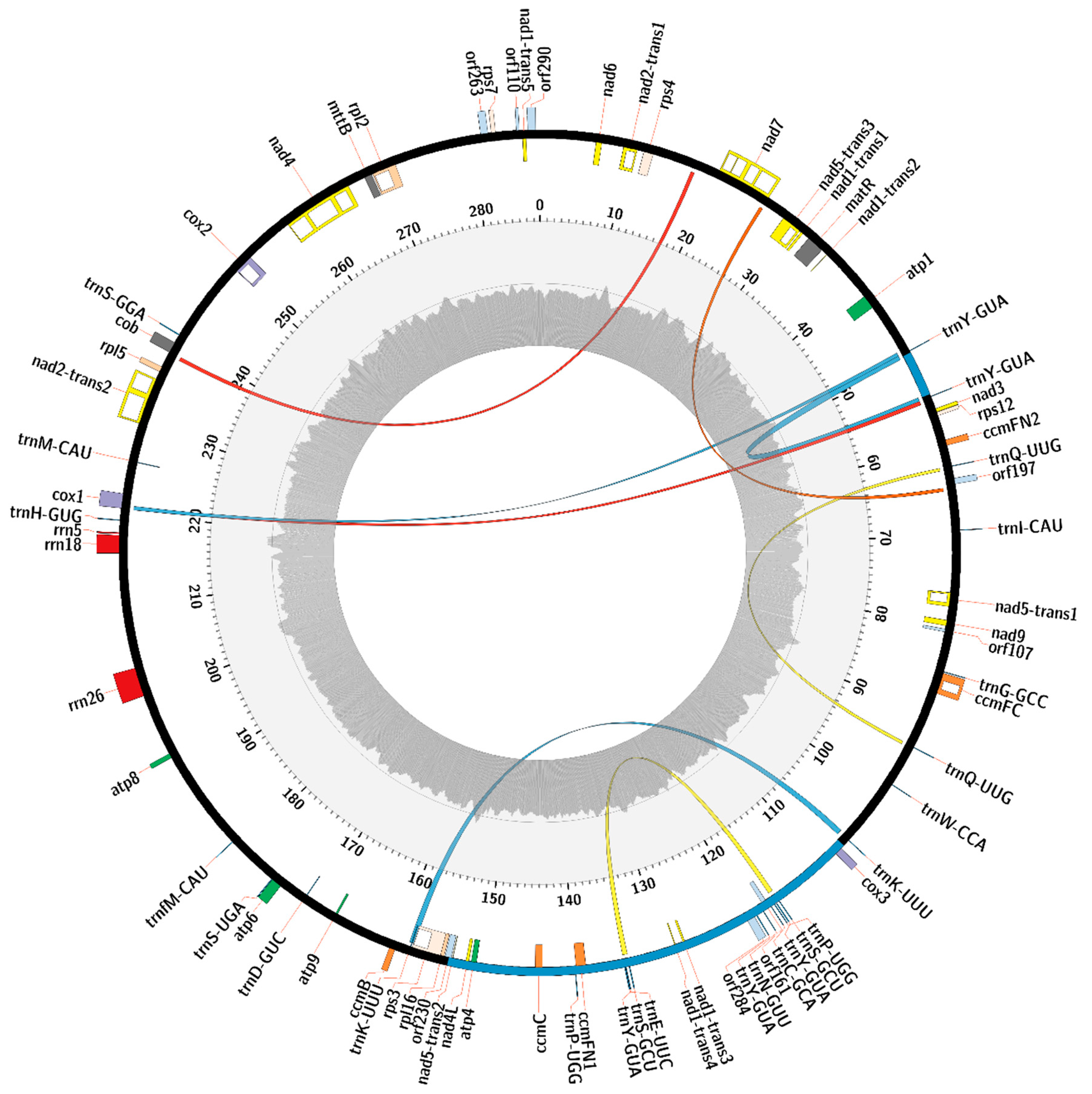
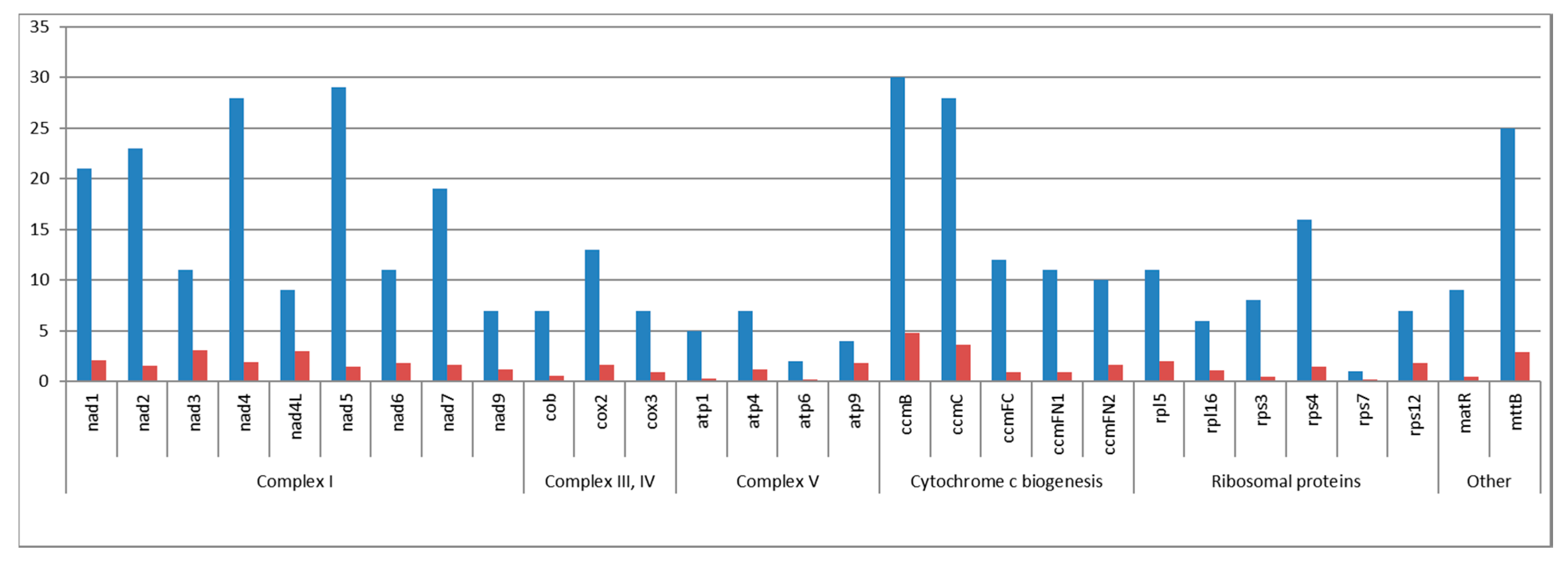
2.3. Gene Content of the C. bursa-pastoris Mitogenome and Comparison with C. rubella and A. thaliana
2.4. RNA Editing
3. Discussion
4. Materials and Methods
4.1. DNA Extraction and Sequencing
4.2. Mitochondrial and Chloroplast Genome Assembly
4.3. Repeats and Structural Variation Analysis
4.4. RNA Extraction, Sequencing, and Analysis
4.5. Genome Annotation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Hintz, M.; Bartholmes, C.; Nutt, P.; Ziermann, J.; Hameister, S.; Neuffer, B.; Theissen, G. Catching a “hopeful monster”: Shepherd’s purse (Capsella bursa-pastoris) as a model system to study the evolution of flower development. J. Exp. Bot. 2006, 57, 3531–3542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, T.-S.; Wu, Q.; Hou, X.-H.; Li, Z.-W.; Zou, Y.-P.; Ge, S.; Guo, Y.-L. Frequent Introgressions from Diploid Species Contribute to the Adaptation of the Tetraploid Shepherd’s Purse (Capsella bursa-pastoris). Mol. Plant 2015, 8, 427–438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bock, R.; Knoop, V. (Eds.) Genomics of Chloroplasts and Mitochondria; Advances in Photosynthesis and Respiration; Springer: Dordrecht, The Netherlands, 2012; ISBN 978-94-007-2919-3. [Google Scholar]
- Davila, J.I.; Arrieta-Montiel, M.P.; Wamboldt, Y.; Cao, J.; Hagmann, J.; Shedge, V.; Xu, Y.-Z.; Weigel, D.; Mackenzie, S.A. Double-strand break repair processes drive evolution of the mitochondrial genome in Arabidopsis. BMC Biol. 2011, 9, 64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maréchal, A.; Brisson, N. Recombination and the maintenance of plant organelle genome stability. New Phytol. 2010, 186, 299–317. [Google Scholar] [CrossRef]
- Skippington, E.; Barkman, T.J.; Rice, D.W.; Palmer, J.D. Miniaturized mitogenome of the parasitic plant Viscum scurruloideum is extremely divergent and dynamic and has lost all nad genes. Proc. Natl. Acad. Sci. USA 2015, 112, E3515–E3524. [Google Scholar] [CrossRef] [Green Version]
- Sloan, D.B.; Alverson, A.J.; Chuckalovcak, J.P.; Wu, M.; McCauley, D.E.; Palmer, J.D.; Taylor, D.R. Rapid Evolution of Enormous, Multichromosomal Genomes in Flowering Plant Mitochondria with Exceptionally High Mutation Rates. PLoS Biol. 2012, 10, e1001241. [Google Scholar] [CrossRef] [Green Version]
- Fauron, C.; Allen, J.; Clifton, S.; Newton, K. Plant Mitochondrial Genomes. In Molecular Biology and Biotechnology of Plant Organelles: Chloroplasts and Mitochondria; Daniell, H., Chase, C., Eds.; Springer: Dordrecht, The Netherlands, 2004; pp. 151–177. ISBN 978-1-4020-3166-3. [Google Scholar]
- Kovar, L.; Nageswara-Rao, M.; Ortega-Rodriguez, S.; Dugas, D.V.; Straub, S.; Cronn, R.; Strickler, S.R.; Hughes, C.E.; Hanley, K.A.; Rodriguez, D.N.; et al. PacBio-Based Mitochondrial Genome Assembly of Leucaena trichandra (Leguminosae) and an Intrageneric Assessment of Mitochondrial RNA Editing. Genome Biol. Evol. 2018, 10, 2501–2517. [Google Scholar] [CrossRef]
- Shearman, J.R.; Sonthirod, C.; Naktang, C.; Pootakham, W.; Yoocha, T.; Sangsrakru, D.; Jomchai, N.; Tragoonrung, S.; Tangphatsornruang, S. The two chromosomes of the mitochondrial genome of a sugarcane cultivar: Assembly and recombination analysis using long PacBio reads. Sci. Rep. 2016, 6, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Dong, S.; Zhao, C.; Chen, F.; Liu, Y.; Zhang, S.; Wu, H.; Zhang, L.; Liu, Y. The complete mitochondrial genome of the early flowering plant Nymphaea colorata is highly repetitive with low recombination. BMC Genom. 2018, 19, 614. [Google Scholar] [CrossRef]
- Gui, S.; Wu, Z.; Zhang, H.; Zheng, Y.; Zhu, Z.; Liang, D.; Ding, Y. The mitochondrial genome map of Nelumbo nucifera reveals ancient evolutionary features. Sci. Rep. 2016, 6, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Liao, X.; Zhao, Y.; Kong, X.; Khan, A.; Zhou, B.; Liu, D.; Kashif, M.H.; Chen, P.; Wang, H.; Zhou, R. Complete sequence of kenaf (Hibiscus cannabinus) mitochondrial genome and comparative analysis with the mitochondrial genomes of other plants. Sci. Rep. 2018, 8, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Mower, J.P.; Sloan, D.B.; Alverson, A.J. Plant Mitochondrial Genome Diversity: The Genomics Revolution. In Plant Genome Diversity Volume 1: Plant Genomes, their Residents, and their Evolutionary Dynamics; Wendel, J.F., Greilhuber, J., Dolezel, J., Leitch, I.J., Eds.; Springer: Vienna, Austria, 2012; pp. 123–144. ISBN 978-3-7091-1130-7. [Google Scholar]
- Lin, H.; Bai, D. The complete mitochondrial genome of a highly selfing species Capsella rubella. Mitochondrial DNA Part B 2019, 4, 1907–1908. [Google Scholar] [CrossRef] [Green Version]
- Tao, Y.-T.; Suo, F.; Tusso, S.; Wang, Y.-K.; Huang, S.; Wolf, J.B.W.; Du, L.-L. Intraspecific Diversity of Fission Yeast Mitochondrial Genomes. Genome Biol. Evol. 2019, 11, 2312–2329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Twyford, A.D.; Ness, R.W. Strategies for complete plastid genome sequencing. Mol. Ecol. Resour. 2017, 17, 858–868. [Google Scholar] [CrossRef] [PubMed]
- Palmer, J.D.; Herbon, L.A. Plant mitochondrial DNA evolves rapidly in structure, but slowly in sequence. J. Mol. Evol. 1988, 28, 87–97. [Google Scholar] [CrossRef]
- Wolfe, K.H.; Li, W.H.; Sharp, P.M. Rates of nucleotide substitution vary greatly among plant mitochondrial, chloroplast, and nuclear DNAs. Proc. Natl. Acad. Sci. USA 1987, 84, 9054–9058. [Google Scholar] [CrossRef] [Green Version]
- Wynn, E.L.; Christensen, A.C. Repeats of Unusual Size in Plant Mitochondrial Genomes: Identification, Incidence and Evolution. G3 (Bethesda) 2018, 9, 549–559. [Google Scholar] [CrossRef] [Green Version]
- André, C.; Levy, A.; Walbot, V. Small repeated sequences and the structure of plant mitochondrial genomes. Trends Genet. 1992, 8, 128–132. [Google Scholar] [CrossRef]
- Douglas, G.M.; Gos, G.; Steige, K.A.; Salcedo, A.; Holm, K.; Josephs, E.B.; Arunkumar, R.; Ågren, J.A.; Hazzouri, K.M.; Wang, W.; et al. Hybrid origins and the earliest stages of diploidization in the highly successful recent polyploid Capsella bursa-pastoris. Proc. Natl. Acad. Sci. USA 2015, 112, 2806–2811. [Google Scholar] [CrossRef] [Green Version]
- Richardson, A.O.; Rice, D.W.; Young, G.J.; Alverson, A.J.; Palmer, J.D. The “fossilized” mitochondrial genome of Liriodendron tulipifera: Ancestral gene content and order, ancestral editing sites, and extraordinarily low mutation rate. BMC Biol. 2013, 11, 29. [Google Scholar] [CrossRef] [Green Version]
- Adams, K.L.; Qiu, Y.-L.; Stoutemyer, M.; Palmer, J.D. Punctuated evolution of mitochondrial gene content: High and variable rates of mitochondrial gene loss and transfer to the nucleus during angiosperm evolution. Proc. Natl. Acad. Sci. USA 2002, 99, 9905–9912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sloan, D.B.; Alverson, A.J.; Štorchová, H.; Palmer, J.D.; Taylor, D.R. Extensive loss of translational genes in the structurally dynamic mitochondrial genome of the angiosperm Silene latifolia. BMC Evol. Biol. 2010, 10, 274. [Google Scholar] [CrossRef] [Green Version]
- Chase, C.D.; Gabay-Laughnan, S. Cytoplasmic Male Sterility and Fertility Restoration by Nuclear Genes. In Molecular Biology and Biotechnology of Plant Organelles; Daniell, H., Chase, C., Eds.; Springer: Dordrecht, The Netherlands, 2004; pp. 593–621. ISBN 978-1-4020-2713-0. [Google Scholar]
- Hanson, M.R.; Bentolila, S. Interactions of Mitochondrial and Nuclear Genes That Affect Male Gametophyte Development. Plant Cell 2004, 16, S154–S169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Handa, H. The complete nucleotide sequence and RNA editing content of the mitochondrial genome of rapeseed (Brassica napus L.): Comparative analysis of the mitochondrial genomes of rapeseed and Arabidopsis thaliana. Nucleic Acids Res. 2003, 31, 5907–5916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finster, S.; Legen, J.; Qu, Y.; Schmitz-Linneweber, C. Land Plant RNA Editing or: Don’t Be Fooled by Plant Organellar DNA Sequences. In Genomics of Chloroplasts and Mitochondria; Bock, R., Knoop, V., Eds.; Advances in Photosynthesis and Respiration; Springer: Dordrecht, The Netherlands, 2012; pp. 293–321. ISBN 978-94-007-2920-9. [Google Scholar]
- Li, J.; Bi, C.; Tu, J.; Lu, Z. The complete mitochondrial genome sequence of Boechera stricta. Mitochondrial DNA Part B 2018, 3, 896–897. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Bi, C. The complete mitochondrial genome sequence of an alpine plant Arabis alpina. Mitochondrial DNA Part B 2018, 3, 725–727. [Google Scholar] [CrossRef] [Green Version]
- Kasianov, A.S.; Klepikova, A.V.; Kulakovskiy, I.V.; Gerasimov, E.S.; Fedotova, A.V.; Besedina, E.G.; Kondrashov, A.S.; Logacheva, M.D.; Penin, A.A. High-quality genome assembly of Capsella bursa-pastoris reveals asymmetry of regulatory elements at early stages of polyploid genome evolution. Plant J. 2017, 91, 278–291. [Google Scholar] [CrossRef] [Green Version]
- Li, H. Minimap2: Pairwise alignment for nucleotide sequences. Bioinformatics 2018, 34, 3094–3100. [Google Scholar] [CrossRef]
- Koren, S.; Walenz, B.P.; Berlin, K.; Miller, J.R.; Bergman, N.H.; Phillippy, A.M. Canu: Scalable and accurate long-read assembly via adaptive k-mer weighting and repeat separation. Genome Res. 2017, 27, 722–736. [Google Scholar] [CrossRef] [Green Version]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef] [Green Version]
- Sedlazeck, F.J.; Rescheneder, P.; Smolka, M.; Fang, H.; Nattestad, M.; von Haeseler, A.; Schatz, M.C. Accurate detection of complex structural variations using single-molecule sequencing. Nat. Methods 2018, 15, 461–468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darling, A.E.; Mau, B.; Perna, N.T. progressiveMauve: Multiple Genome Alignment with Gene Gain, Loss and Rearrangement. PLoS ONE 2010, 5, e11147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thorvaldsdóttir, H.; Robinson, J.T.; Mesirov, J.P. Integrative Genomics Viewer (IGV): High-performance genomics data visualization and exploration. Brief. Bioinform. 2013, 14, 178–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.; Paggi, J.M.; Park, C.; Bennett, C.; Salzberg, S.L. Graph-based genome alignment and genotyping with HISAT2 and HISAT-genotype. Nat. Biotechnol. 2019, 37, 907–915. [Google Scholar] [CrossRef]
- Li, H. A statistical framework for SNP calling, mutation discovery, association mapping and population genetical parameter estimation from sequencing data. Bioinformatics 2011, 27, 2987–2993. [Google Scholar] [CrossRef] [Green Version]
- Danecek, P.; Schiffels, S.; Durbin, R. Multiallelic calling model in bcftools (-m). Available online: https://samtools.github.io/bcftools/call-m.pdf (accessed on 23 January 2020).
- Wu, S.; Liu, W.; Aljohi, H.A.; Alromaih, S.A.; Alanazi, I.O.; Lin, Q.; Yu, J.; Hu, S. REDO: RNA Editing Detection in Plant Organelles Based on Variant Calling Results. J. Comput. Biol. 2018, 25, 509–516. [Google Scholar] [CrossRef]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. 1000 Genome Project Data Processing Subgroup The Sequence Alignment/Map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [Green Version]
- Alverson, A.J.; Wei, X.; Rice, D.W.; Stern, D.B.; Barry, K.; Palmer, J.D. Insights into the Evolution of Mitochondrial Genome Size from Complete Sequences of Citrullus lanatus and Cucurbita pepo (Cucurbitaceae). Mol. Biol. Evol. 2010, 27, 1436–1448. [Google Scholar] [CrossRef] [Green Version]
- Shi, L.; Chen, H.; Jiang, M.; Wang, L.; Wu, X.; Huang, L.; Liu, C. CPGAVAS2, an integrated plastome sequence annotator and analyzer. Nucleic Acids Res. 2019, 47, W65–W73. [Google Scholar] [CrossRef]
- Krzywinski, M.; Schein, J.; Birol, I.; Connors, J.; Gascoyne, R.; Horsman, D.; Jones, S.J.; Marra, M.A. Circos: An information aesthetic for comparative genomics. Genome Res. 2009, 19, 1639–1645. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Repeat ID | Structural Variant Type | Number of Supporting CCS Reads | Repeat Length (bp) | First Repeat Unit Position (bp) | Second Repeat Unit Position (bp) | Repeat Units’ Sequence Identity |
|---|---|---|---|---|---|---|
| Rep_1 | inversion | 13 | 854 | 54,219–55,072 | 49,506–48,653 | 98.9% |
| Rep_2 | inversion | 8 | 635 | 158,147–158,781 | 106,515–105,881 | 100.0% |
| Rep_3 | duplication | 6 | 538 | 134,019–134,556 | 116,143–116,679 | 99.8% |
| Rep_7 | deletion | 5 | 418 | 220,620–221,037 | 54,655–55,072 | 99.8% |
| Rep_10 | inversion | 4 | 420 | 220,623–221,042 | 49,067–48,648 | 97.9% |
| Rep_11 | duplication | 3 | 356 | 64,729–65,084 | 26,047–26,402 | 100.0% |
| Rep_12 | duplication | 3 | 327 | 93,763–94,089 | 62,446–62,772 | 100.0% |
| Rep_9 | deletion | 2 | 404 | 238,171–238,573 | 17,371–17,774 | 99.8% |
| Rep_11 | deletion | 2 | 356 | 64,729–65,084 | 26,047–26,402 | 100.0% |
| Name * | InterProScan Predictions | BLASTp Similarity |
|---|---|---|
| orf290 | Cytochrome c oxidase, subunit II (cox2) domain with 2 transmembrane regions within | cox2 |
| orf197 | 1 transmembrane region | atp6 |
| orf107 | Nothing found | hypothetical protein |
| orf161 | Member of Protein TIC214 (ycf1) InterPro family. Ycf1 domain and 4 transmembrane regions within the domain | ycf1 |
| orf284 | Signal peptide (located 1–17 aa) and 2 transmembrane regions | atp9 |
| orf230 | Member of Ribosomal protein L2 (rpl2) InterPro family. Ribosomal_L2 domain and 1 transmembrane region | rpl2 |
| orf263 | Member of ATP synthase, F0 complex, subunit C (atp9) InterPro family. ATP synthase, subunit C, isoform a domain and 3 transmembrane regions within the domain | atp9 |
| orf110 | Nothing found | hypothetical protein |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Omelchenko, D.O.; Makarenko, M.S.; Kasianov, A.S.; Schelkunov, M.I.; Logacheva, M.D.; Penin, A.A. Assembly and Analysis of the Complete Mitochondrial Genome of Capsella bursa-pastoris. Plants 2020, 9, 469. https://doi.org/10.3390/plants9040469
Omelchenko DO, Makarenko MS, Kasianov AS, Schelkunov MI, Logacheva MD, Penin AA. Assembly and Analysis of the Complete Mitochondrial Genome of Capsella bursa-pastoris. Plants. 2020; 9(4):469. https://doi.org/10.3390/plants9040469
Chicago/Turabian StyleOmelchenko, Denis O., Maxim S. Makarenko, Artem S. Kasianov, Mikhail I. Schelkunov, Maria D. Logacheva, and Aleksey A. Penin. 2020. "Assembly and Analysis of the Complete Mitochondrial Genome of Capsella bursa-pastoris" Plants 9, no. 4: 469. https://doi.org/10.3390/plants9040469