The 21st Century Agriculture: When Rice Research Draws Attention to Climate Variability and How Weedy Rice and Underutilized Grains Come in Handy

 and
and
Abstract
:1. Introduction
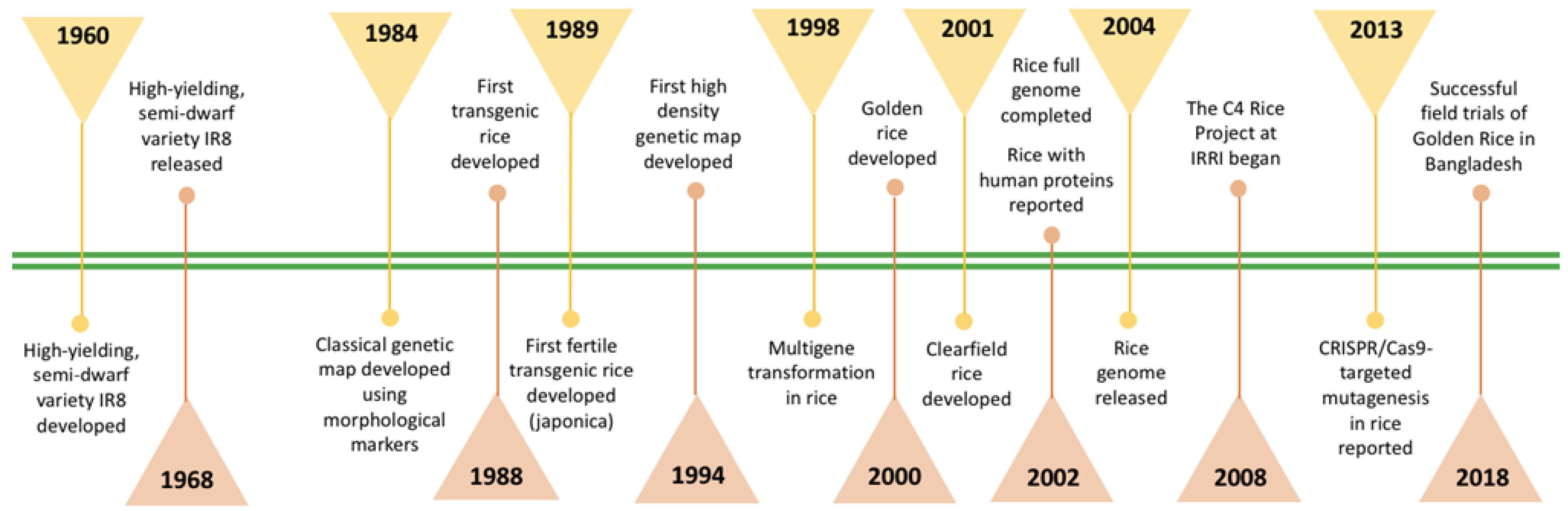
2. Highlights of Rice Research since the Green Revolution
2.1. Early Green Revolution
2.2. Late Green Revolution
2.3. 21st Century
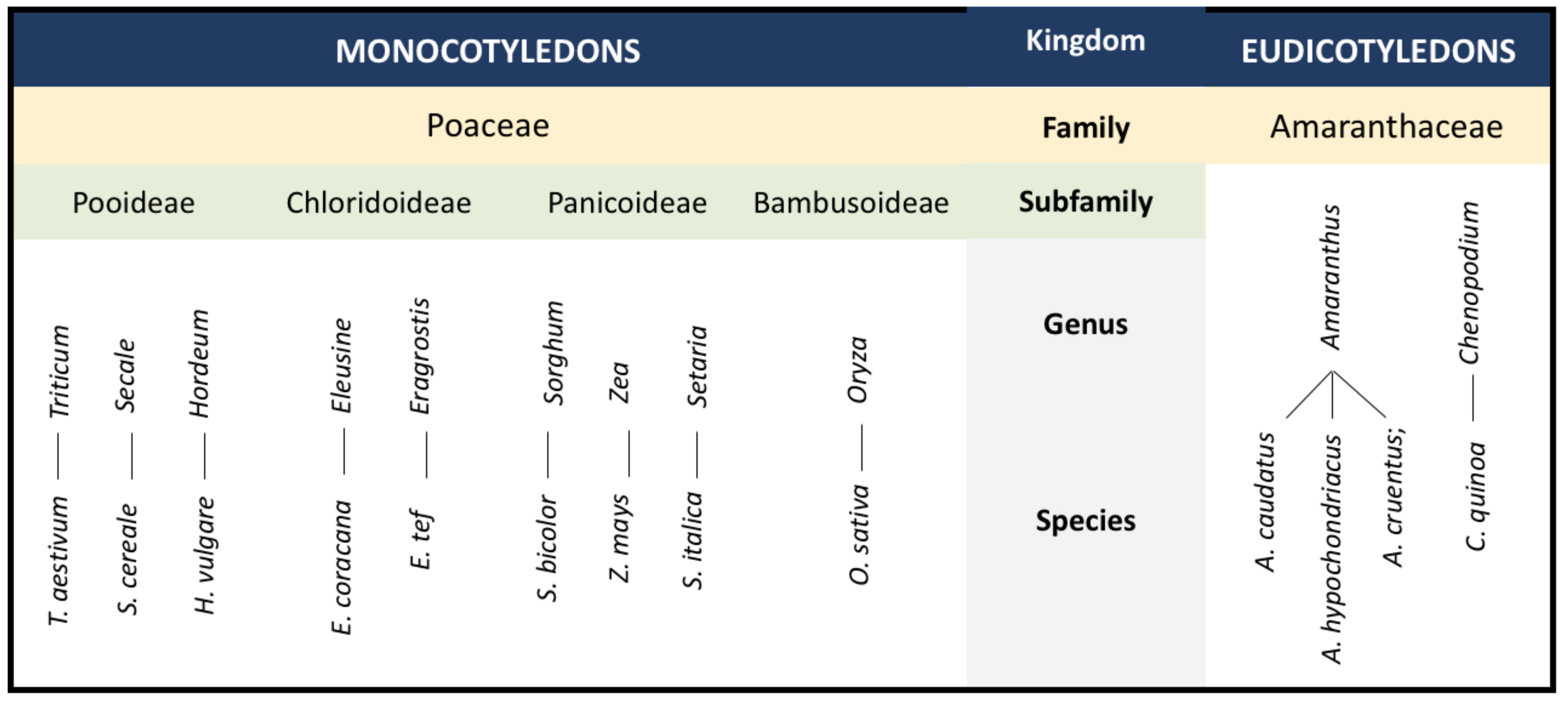
3. Weedy Rice and Underutilized Grain Crops as Potential Complement to Existing Rice Research
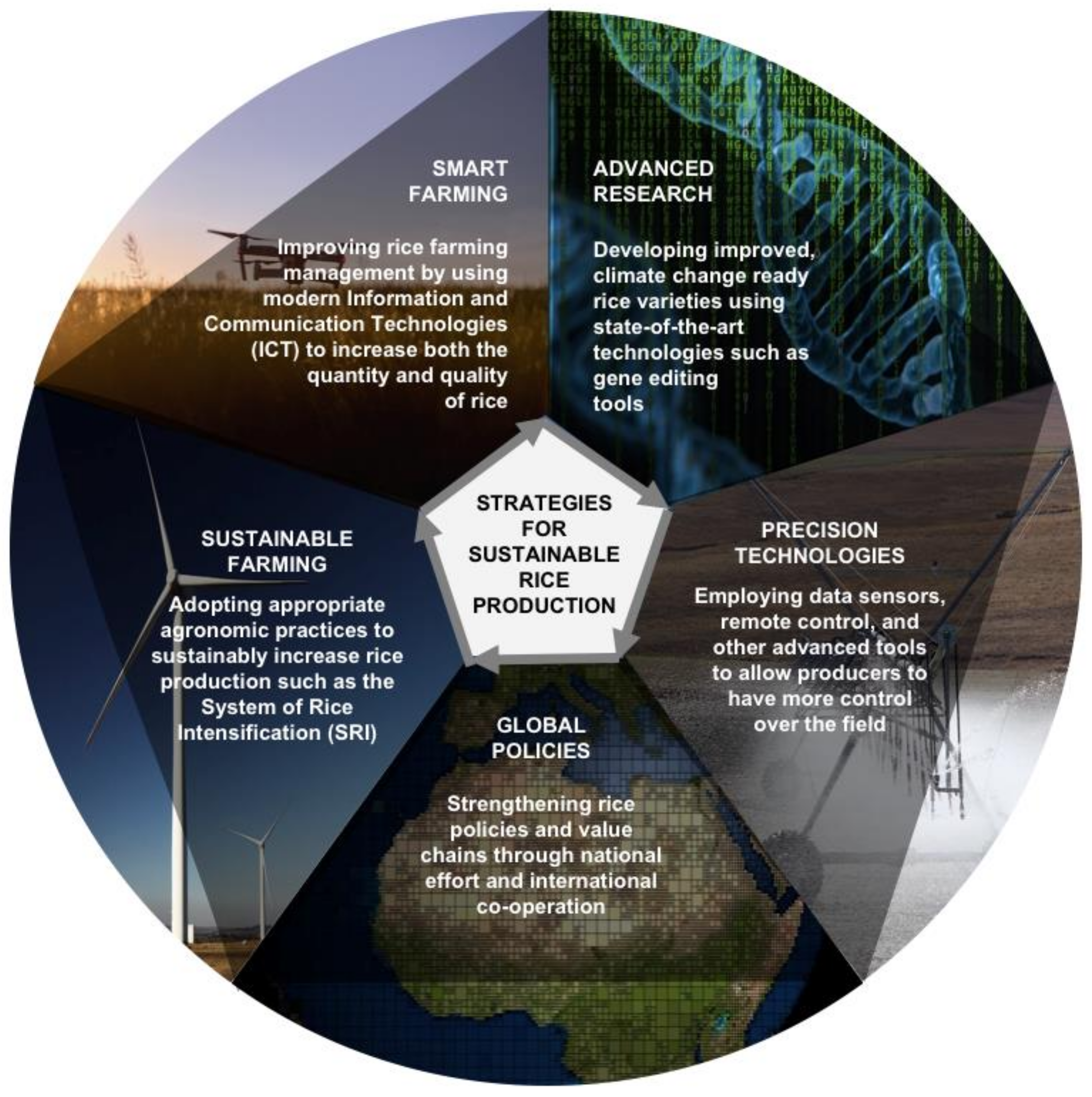
4. Laying the Route to Sustainable Rice Production: What Can We Possibly Do?
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Cheng, A. Shaping a sustainable food future by rediscovering long-forgotten ancient grains. Plant Sci. 2018, 269, 136–142. [Google Scholar] [CrossRef]
- World Health Organization. Health in 2015: From MDGs, Millennium Development Goals to SDGs, Sustainable Development Goals; WHO Press: Geneva, Switzerland, 2015. [Google Scholar]
- Keesstra, S.D.; Bouma, J.; Wallinga, J.; Tittonell, P.; Smith, P.; Cerdà, A.; Montanarella, L.; Quinton, J.N.; Pachepsky, Y.; Van Der Putten, W.H. The significance of soils and soil science towards realization of the United Nations Sustainable Development Goals. Soil 2016. [Google Scholar] [CrossRef] [Green Version]
- Khush, G.S. Green revolution: The way forward. Nat. Rev. Genet. 2001, 2, 815–822. [Google Scholar] [CrossRef] [PubMed]
- Lobell, D.B.; Schlenker, W.; Costa-Roberts, J. Climate trends and global crop production since 1980. Science 2011, 333, 616–620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thornton, P.K.; Ericksen, P.J.; Herrero, M.; Challinor, A.J. Climate variability and vulnerability to climate change: A review. Global Change Biol. 2014, 20, 3313–3328. [Google Scholar] [CrossRef]
- Adhikari, V.R.; Devkota, N.; Phuyal, R.K. Impact of climate variation in paddy production in Nepal. Int. J. Econ. Perspect. 2017, 11, 1084–1092. [Google Scholar]
- Aditya, J.; Bhartiya, A.; Chahota, R.K.; Joshi, D.; Chandra, N.; Kant, L.; Pattanayak, A. Ancient orphan legume horse gram: A potential food and forage crop of future. Planta 2019, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Pattanayak, A.; Roy, S.; Sood, S.; Iangrai, B.; Banerjee, A.; Gupta, S.; Joshi, D.C. Rice bean: A lesser known pulse with well-recognized potential. Planta 2019, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Gibson, D.J. Grasses and Grassland Ecology; Oxford University Press: New York, NY, USA, 2009. [Google Scholar]
- Khush, G.S. Origin, dispersal, cultivation and variation of rice. Plant Mol. Biol. 1997, 35, 25–34. [Google Scholar] [CrossRef]
- Abraham, A.; Mathew, A.K.; Sindhu, R.; Pandey, A.; Binod, P. Potential of rice straw for bio-refining: An overview. Bioresour. Technol. 2016, 215, 29–36. [Google Scholar] [CrossRef]
- Ladha, J.; Tirol-Padre, A.; Reddy, K.; Ventura, W. Prospects and problems of biological nitrogen fixation in rice production: A critical assessment. In New Horizons in Nitrogen Fixation; Palacios, R., Mora, J., Newton, W., Eds.; Springer: Cancun, Mexico, 1993; pp. 677–682. [Google Scholar]
- Zeigler, R.S.; Barclay, A. The relevance of rice. Rice 2008, 1, 3–10. [Google Scholar] [CrossRef] [Green Version]
- USDA. Overview. Rice. Available online: https://www.ers.usda.gov/topics/crops/rice.aspx (accessed on 26 June 2019).
- FAO. A Regional Strategy for Sustainable Hybrid Rice Development in Asia; Food and Agriculture Organization: Rome, Italy, 2014. [Google Scholar]
- Atwell, B.J.; Wang, H.; Scafaro, A.P. Could abiotic stress tolerance in wild relatives of rice be used to improve Oryza sativa? Plant Sci. 2014, 215, 48–58. [Google Scholar] [CrossRef] [PubMed]
- Sweeney, M.; McCouch, S. The complex history of the domestication of rice. Ann. Bot. 2007, 100, 951–957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kasem, S. Evaluating Grain Food Potential of Wild Relatives of Rice. Ph.D. Thesis, Southern Cross University, East Lismore, NSW, Australia, 2012. [Google Scholar]
- Londo, J.P.; Chiang, Y.-C.; Hung, K.-H.; Chiang, T.-Y.; Schaal, B.A. Phylogeography of Asian wild rice, Oryza rufipogon, reveals multiple independent domestications of cultivated rice, Oryza sativa. Proc. Natl. Acad. Sci. USA 2006, 103, 9578–9583. [Google Scholar] [CrossRef] [Green Version]
- Lu, B.-R.; Snow, A.A. Gene flow from genetically modified rice and its environmental consequences. BioScience 2005, 55, 669–678. [Google Scholar] [CrossRef] [Green Version]
- Hamilton, R.W.; Ammayao, A. The Art of Rice: Spirit and Sustenance in Asia; University of California: Los Angeles, CA, USA, 2003. [Google Scholar]
- Izawa, T.; Shimamoto, K. Becoming a model plant: The importance of rice to plant science. Trends Plant Sci. 1996, 1, 95–99. [Google Scholar] [CrossRef]
- Chen, M.; Presting, G.; Barbazuk, W.B.; Goicoechea, J.L.; Blackmon, B.; Fang, G.; Kim, H.; Frisch, D.; Yu, Y.; Sun, S. An integrated physical and genetic map of the rice genome. Plant Cell 2002, 14, 537–545. [Google Scholar] [CrossRef]
- Kumar, S.; Balyan, H.S.; Gupta, P.K. Comparative DNA sequence analysis involving wheat, Brachypodium and rice genomes using mapped wheat ESTs. Triticeae Genom. Genet. 2012, 3. [Google Scholar] [CrossRef]
- Jena, K.K. The species of the genus Oryza and transfer of useful genes from wild species into cultivated rice, O. sativa. Breeding Sci. 2010, 60, 518–523. [Google Scholar] [CrossRef] [Green Version]
- Harlan, J.R.; De Wet, J.M. Toward a rational classification of cultivated plants. Taxon 1971, 20, 509–517. [Google Scholar] [CrossRef]
- Shan, Q.; Wang, Y.; Li, J.; Gao, C. Genome editing in rice and wheat using the CRISPR/Cas system. Nat. Protoc. 2014, 9, 2395. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zhang, J.; Wei, P.; Zhang, B.; Gou, F.; Feng, Z.; Mao, Y.; Yang, L.; Zhang, H.; Xu, N. The CRISPR/Cas9 system produces specific and homozygous targeted gene editing in rice in one generation. Plant Biotechnol. J. 2014, 12, 797–807. [Google Scholar] [CrossRef] [PubMed]
- Jackson, S.A. Rice: The first crop genome. Rice 2016, 9, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, J.; Qian, Q.; Ma, D.R.; Xu, Z.J.; Liu, D.; Du, H.B.; Chen, W.F. Introgression and selection shaping the genome and adaptive loci of weedy rice in northern China. New Phytol. 2013, 197, 290–299. [Google Scholar] [CrossRef] [PubMed]
- Jia, Y.; Gealy, D. Weedy red rice has novel sources of resistance to biotic stress. Crop J. 2018, 6, 443–450. [Google Scholar] [CrossRef]
- Hargrove, T.R.; Cabanilla, V.L. The impact of semidwarf varieties on Asian rice-breeding programs. BioScience 1979, 29, 731–735. [Google Scholar] [CrossRef]
- Massawe, F.; Mayes, S.; Cheng, A. Crop diversity: An unexploited treasure trove for food security. Trends Plant Sci. 2016, 21, 365–368. [Google Scholar] [CrossRef]
- Khush, G.S. Rice breeding: Past, present and future. J. Genet. 1987, 66, 195–216. [Google Scholar] [CrossRef]
- Khush, G.S. Modern varieties—their real contribution to food supply and equity. GeoJournal 1995, 35, 275–284. [Google Scholar] [CrossRef]
- Syme, J. A high-yielding Mexican semi-dwarf wheat and the relationship of yield to harvest index and other varietal characteristics. Aust. J. Exp. Agric. 1970, 10, 350–353. [Google Scholar] [CrossRef]
- Welch, V.A. Semi-dwarf Maize and Method. U.S. Patent US4368592A.
- Dalrymple, D.G. Development and Spread of High-Yielding Rice Varieties in Developing Countries; Metrotec Inc.: Washington, DC, USA, 1986. [Google Scholar]
- Matson, P.A.; Parton, W.J.; Power, A.; Swift, M. Agricultural intensification and ecosystem properties. Science 1997, 277, 504–509. [Google Scholar] [CrossRef] [Green Version]
- Ameen, A.; Raza, S. Green revoultion: A review. Int. J. Adv. Sci. Res. 2017, 3, 129–137. [Google Scholar] [CrossRef] [Green Version]
- Gollin, D.; Morris, M.; Byerlee, D. Technology adoption in intensive post-green revolution systems. Am. J. Agric. Econ. 2005, 87, 1310–1316. [Google Scholar] [CrossRef]
- Toriyama, K.; Arimoto, Y.; Uchimiya, H.; Hinata, K. Transgenic rice plants after direct gene transfer into protoplasts. Bio/technology 1988, 6, 1072–1074. [Google Scholar] [CrossRef]
- Zhang, H.; Yang, H.; Rech, E.; Golds, T.; Davis, A.; Mulligan, B.; Cocking, E.; Davey, M. Transgenic rice plants produced by electroporation-mediated plasmid uptake into protoplasts. Plant Cell Rep. 1988, 7, 379–384. [Google Scholar]
- Giri, C.; Laxmi, G.V. Production of transgenic rice with agronomically useful genes: An assessment. Biotechnol. Adv. 2000, 18, 653–683. [Google Scholar] [CrossRef]
- Duan, X.; Li, X.; Xue, Q.; Abo-EI-Saad, M.; Xu, D.; Wu, R. Transgenic rice plants harboring an introduced potato proteinase inhibitor II gene are insect resistant. Nat. Biotechnol. 1996, 14, 494. [Google Scholar] [CrossRef] [PubMed]
- Pinto, Y.M.; Kok, R.A.; Baulcombe, D.C. Resistance to rice yellow mottle virus (RYMV) in cultivated African rice varieties containing RYMV transgenes. Nat. Biotechnol. 1999, 17, 702. [Google Scholar] [CrossRef] [PubMed]
- Uchimiya, H.; Iwata, M.; Nojiri, C.; Samarajeewa, P.K.; Takamatsu, S.; Ooba, S.; Anzai, H.; Christensen, A.H.; Quail, P.H.; Toki, S. Bialaphos treatment of transgenic rice plants expressing a bar gene prevents infection by the sheath blight pathogen (Rhizoctonia solani). Bio/technology 1993, 11, 835–836. [Google Scholar] [CrossRef]
- Song, W.-Y.; Wang, G.-L.; Chen, L.-L.; Kim, H.-S.; Pi, L.-Y.; Holsten, T.; Gardner, J.; Wang, B.; Zhai, W.-X.; Zhu, L.-H. A receptor kinase-like protein encoded by the rice disease resistance gene, Xa21. Science 1995, 270, 1804–1806. [Google Scholar] [CrossRef] [Green Version]
- Tu, J.; Zhang, G.; Datta, K.; Xu, C.; He, Y.; Zhang, Q.; Khush, G.S.; Datta, S.K. Field performance of transgenic elite commercial hybrid rice expressing Bacillus thuringiensis δ-endotoxin. Nat. Biotechnol. 2000, 18, 1101. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.-Y.; Mei, F.; Zhang, W.; Shen, Z.; Fang, J. Creation of Bt rice expressing a fusion protein of Cry1Ac and Cry1I-like using a green tissue-specific promoter. J. Econ. Entomol. 2014, 107, 1674–1679. [Google Scholar] [CrossRef] [PubMed]
- Ye, X.; Zhang, G.; Al-Babili, S.; Kloti, A.; Zhang, J.; Lucca, P.; Beyer, P.; Potrykus, I. Field performance of transgenic elite commercial hybrid rice expressing Bacillus thuringiensis δ-endotoxin. Engineering the provitamin A (β-carotene) biosynthetic pathway into (carotenoid-free) rice endosperm. Science 2000, 287, 303–305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- ISAAA. Philippines Approves Golden Rice for Direct Use as Food and Feed, or for Processing. Available online: http://www.isaaa.org/kc/cropbiotechupdate/article/default.asp?ID=17900 (accessed on 12 February 2020).
- Cornell Alliance for Science. US FDA Approves Golden Rice. Available online: https://allianceforscience.cornell.edu/blog/2018/05/us-fda-approves-golden-rice (accessed on 14 February 2020).
- Babujee, L.; Gnanamanickam, S. Molecular tools for characterization of rice blast pathogen (Magnaporthe grisea) population and molecular marker-assisted breeding for disease resistance. Curr. Sci. 2000, 78, 248–257. [Google Scholar]
- Pradhan, S.K.; Pandit, E.; Pawar, S.; Baksh, S.Y.; Mukherjee, A.K.; Mohanty, S.P. Development of flash-flood tolerant and durable bacterial blight resistant versions of mega rice variety ‘Swarna’ through marker-assisted backcross breeding. Sci. Rep. 2019, 9, 12810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Z.; Young, N.D.; Reagon, M.; Hyma, K.E.; Olsen, K.M.; Jia, Y.; Caicedo, A.L. All roads lead to weediness: Patterns of genomic divergence reveal extensive recurrent weedy rice origins from South Asian Oryza. Mol. Ecol. 2017, 26, 3151–3167. [Google Scholar] [CrossRef]
- Yano, K.; Yamamoto, E.; Aya, K.; Takeuchi, H.; Lo, P.-c.; Hu, L.; Yamasaki, M.; Yoshida, S.; Kitano, H.; Hirano, K. Genome-wide association study using whole-genome sequencing rapidly identifies new genes influencing agronomic traits in rice. Nat. Genet. 2016, 48, 927–934. [Google Scholar] [CrossRef]
- Bajaj, S.; Mohanty, A. Recent advances in rice biotechnology—towards genetically superior transgenic rice. Plant Biotechnol. J. 2005, 3, 275–307. [Google Scholar] [CrossRef]
- Sasaki, T.; Matsumoto, T.; Antonio, B.A.; Nagamura, Y. From mapping to sequencing, post-sequencing and beyond. Plant Cell Physiol. 2005, 46, 3–13. [Google Scholar] [CrossRef]
- Quackenbush, J.; Cho, J.; Lee, D.; Liang, F.; Holt, I.; Karamycheva, S.; Parvizi, B.; Pertea, G.; Sultana, R.; White, J. The TIGR Gene Indices: Analysis of gene transcript sequences in highly sampled eukaryotic species. Nucleic Acids Res. 2001, 29, 159–164. [Google Scholar] [CrossRef] [Green Version]
- Somerville, C.; Koornneef, M. A fortunate choice: The history of Arabidopsis as a model plant. Nat. Rev. Genet. 2002, 3, 883. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Guo, L.; Li, Y.; Wang, Z. Systematic comparison of C3 and C4 plants based on metabolic network analysis. BMC Syst. Biol. 2012, 6, S9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, P.; Vlad, D.; Langdale, J.A. Finding the genes to build C4 rice. Curr. Opin. Plant Biol. 2016, 31, 44–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mishra, S.; Singh, M.K.; Snehal, S.; Pathak, H. C4 Rice-Tweaking Rice Physiology for Second Green Revolution. Int. J. Curr. Microbiol. Appl. Sci. 2017, 6, 1161–1176. [Google Scholar] [CrossRef]
- Miao, J.; Guo, D.; Zhang, J.; Huang, Q.; Qin, G.; Zhang, X.; Wan, J.; Gu, H.; Qu, L.-J. Targeted mutagenesis in rice using CRISPR-Cas system. Cell Res. 2013, 23, 1233–1236. [Google Scholar] [CrossRef] [Green Version]
- Jiang, W.; Zhou, H.; Bi, H.; Fromm, M.; Yang, B.; Weeks, D.P. Demonstration of CRISPR/Cas9/sgRNA-mediated targeted gene modification in Arabidopsis, tobacco, sorghum and rice. Nucleic Acids Res. 2013, 41, e188. [Google Scholar] [CrossRef]
- Endo, M.; Mikami, M.; Toki, S. Multigene knockout utilizing off-target mutations of the CRISPR/Cas9 system in rice. Plant Cell Physiol. 2015, 56, 41–47. [Google Scholar] [CrossRef] [Green Version]
- Fiaz, S.; Ahmad, S.; Noor, M.A.; Wang, X.; Younas, A.; Riaz, A.; Riaz, A.; Ali, F. Applications of the CRISPR/Cas9 system for rice grain quality improvement: Perspectives and opportunities. Int. J. Mol. Sci. 2019, 20, 888. [Google Scholar] [CrossRef] [Green Version]
- Wani, S.; Sah, S. Biotechnology and abiotic stress tolerance in rice. Rice Res. 2014. [Google Scholar] [CrossRef]
- Cantrell, R.P.; Hettel, G.P. Research Strategy for Rice in the 21st Century; Copyright International Rice Research Institute: Los Baños, Phillipines, 2005; p. 26. [Google Scholar]
- Tadele, Z. Orphan crops: Their importance and the urgency of improvement. Planta 2019, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Tadele, Z.; Bartels, D. Promoting Orphan Crops Research and Development; Springer: Berlin, Germany, 2019. [Google Scholar]
- FAO; IFAD; UNICEF; WFP; WHO. The State of Food Security and Nutrition in the World 2017; FAO: Rome, Italy, 2017. [Google Scholar]
- Singh, K.; Kumar, V.; Saharawat, Y.; Gathala, M.; Ladha, J.; Chauhan, B. Weedy rice: An emerging threat for direct-seeded rice production systems in India. Rice Res. 2013. [Google Scholar] [CrossRef]
- Vidotto, F.; Ferrero, A. Germination behaviour of red rice (Oryza sativa L.) seeds in field and laboratory conditions. Agronomie 2000, 20, 375–382. [Google Scholar] [CrossRef]
- Suh, H. Characterization of Weedy Rice Germplasm; Wild Crop Germplasm Bank; Yeungnam University: Joyeong-dong, Korea.
- Oka, H.-I. Origin of Cultivated Rice; Elsevier: Tokyo, Japan, 2012. [Google Scholar]
- Kanapeckas, K.L.; Vigueira, C.C.; Ortiz, A.; Gettler, K.A.; Burgos, N.R.; Fischer, A.J.; Lawton-Rauh, A.L. Escape to ferality: The endoferal origin of weedy rice from crop rice through de-domestication. PLoS ONE 2016. [Google Scholar] [CrossRef] [PubMed]
- Ziska, L.H.; Gealy, D.R.; Tomecek, M.B.; Jackson, A.K.; Black, H.L. Recent and projected increases in atmospheric CO2 concentration can enhance gene flow between wild and genetically altered rice (Oryza sativa). PLoS ONE 2012, 7, e37522. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Qi, X.; Gealy, D.R.; Olsen, K.M.; Caicedo, A.L.; Jia, Y. QTL analysis for resistance to blast disease in US weedy rice. Mol. Plant-Microbe Interact. 2015, 28, 834–844. [Google Scholar] [CrossRef] [Green Version]
- Nadir, S.; Xiong, H.-B.; Zhu, Q.; Zhang, X.-L.; Xu, H.-Y.; Li, J.; Dongchen, W.; Henry, D.; Guo, X.-Q.; Khan, S. Weedy rice in sustainable rice production. A review. Agron. Sustain. Dev. 2017, 37, 46. [Google Scholar] [CrossRef]
- Chen, L.J.; Lee, D.S.; Song, Z.P.; Suh, H.S.; LU, B.R. Gene flow from cultivated rice (Oryza sativa) to its weedy and wild relatives. Ann. Botany 2004, 93, 67–73. [Google Scholar] [CrossRef] [Green Version]
- He, Q.; Kim, K.W.; Park, Y.J. Population genomics identifies the origin and signatures of selection of Korean weedy rice. Plant Biotechnol. J. 2016, 15, 357–366. [Google Scholar] [CrossRef]
- Qiu, J.; Zhou, Y.; Mao, L.; Ye, C.; Wang, W.; Zhang, J.; Yu, Y.; Fu, F.; Wang, Y.; Qian, F. Genomic variation associated with local adaptation of weedy rice during de-domestication. Nat. Commun. 2017. [Google Scholar] [CrossRef]
- Zhang, Y.; Fang, J.; Wu, X.; Dong, L. Na+/K+ balance and transport regulatory mechanisms in weedy and cultivated rice (Oryza sativa L.) under salt stress. BMC Plant Biol. 2018, 18, 375. [Google Scholar] [CrossRef]
- Bevilacqua, C.B.; Basu, S.; Pereira, A.; Tseng, T.-M.; Zimmer, P.D.; Burgos, N.R. Analysis of stress-responsive gene expression in cultivated and weedy rice differing in cold stress tolerance. PLoS ONE 2015, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.; Jia, Y.; Jia, M.; Gealy, D.R.; Olsen, K.M.; Caicedo, A.L. Molecular evolution of the rice blast resistance gene Pi-ta in invasive weedy rice in the USA. PLoS ONE 2011, 6, e26260. [Google Scholar] [CrossRef] [PubMed]
- Kabisch, N.; Haase, D. Diversifying European agglomerations: Evidence of urban population trends for the 21st century. Popul. Space Place 2011, 17, 236–253. [Google Scholar] [CrossRef]
- Reardon, T.; Tschirley, D.; Dolislager, M.; Snyder, J.; Hu, C.; White, S. Urbanization, Diet Change, and Ttransformation of Food Supply Chains in Asia; Global Center for Food Systems Innovation: East Lansing, MI, USA, 2014. [Google Scholar]
- Ruel, M.T.; Garrett, J.; Yosef, S.; Olivier, M. Urbanization, food security and nutrition. In Nutrition and Health in a Developing World; Springer: Berlin, Germany, 2017; pp. 705–735. [Google Scholar]
- Padulosi, S.; Hodgkin, T.; Williams, J.; Haq, N.; Engles, J.; Rao, V.; Brown, A.; Jackson, M. Underutilized Crops: Trends, Challenges and Opportunities in the 21st Century; CAB International: Wallingford, UK, 2002; p. 323. [Google Scholar]
- Adhikari, L.; Tuladhar, S.; Hussain, A.; Aryal, K. Are Traditional Food Crops Really ‘Future Smart Foods?’A Sustainability Perspective. Sustainability 2019, 11, 5236. [Google Scholar] [CrossRef] [Green Version]
- Mabhaudhi, T.; Chimonyo, V.G.P.; Hlahla, S.; Massawe, F.; Mayes, S.; Nhamo, L.; Modi, A.T. Prospects of orphan crops in climate change. Planta 2019, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chanyalew, S.; Ferede, S.; Damte, T.; Fikre, T.; Genet, Y.; Kebede, W.; Tolossa, K.; Tadele, Z.; Assefa, K. Significance and prospects of an orphan crop tef. Planta 2019, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joshi, D.; Chaudhari, G.V.; Sood, S.; Kant, L.; Pattanayak, A.; Zhang, K.; Fan, Y.; Janovská, D.; Meglič, V.; Zhou, M. Revisiting the versatile buckwheat: Reinvigorating genetic gains through integrated breeding and genomics approach. Planta 2019, 1–19. [Google Scholar] [CrossRef]
- Arendt, E.K.; Zannini, E. Cereal Grains for the Food and Beverage Industries; Elsevier: Amsterdam, The Netherlands, 2013. [Google Scholar]
- Asfaw, K.G.; Danno, F.I. Effects of salinity on yield and yield components of tef [Eragrostis tef (Zucc.) Trotter] accessions and varieties. Curr. Res. J. Biol. Sci. 2011, 3, 289–299. [Google Scholar]
- Caruso, C.; Maucieri, C.; Berruti, A.; Borin, M.; Barbera, A. Responses of different Panicum miliaceum L. genotypes to saline and water stress in a marginal Mediterranean environment. Agronomy 2018, 8, 8. [Google Scholar] [CrossRef] [Green Version]
- Danielsen, S.; Bonifacio, A.; Ames, T. Diseases of quinoa (Chenopodium quinoa). Food. Rev. Int. 2003, 19, 43–59. [Google Scholar] [CrossRef]
- Habiyaremye, C.; Barth, V.; Highet, K.; Coffey, T.; Murphy, K. Phenotypic responses of twenty diverse proso millet (Panicum miliaceum L.) accessions to irrigation. Sustainability 2017, 9, 389. [Google Scholar] [CrossRef] [Green Version]
- Hinojosa, L.; González, J.; Barrios-Masias, F.; Fuentes, F.; Murphy, K. Quinoa abiotic stress responses: A review. Plants 2018, 7, 106. [Google Scholar] [CrossRef] [Green Version]
- National Research Council. National Science Education Standards; National Academies Press: Washington, DC, USA, 1996. [Google Scholar]
- Wilson, R.; Courteau, J. Search of plant introduction proso millets for fall armyworm resistance. J. Econ. Entomol. 1984, 77, 171–173. [Google Scholar] [CrossRef]
- Chakraborty, S.; Chakraborty, N.; Datta, A. Increased nutritive value of transgenic potato by expressing a nonallergenic seed albumin gene from Amaranthus hypochondriacus. Proc. Natl. Acad. Sci. USA 2000, 97, 3724–3729. [Google Scholar] [CrossRef] [PubMed]
- Cannarozzi, G.; Plaza-wuthrich, S.; Esfeld, K.; Larti, S.; Wilson, Y.S.; Girma, D.; Castro, E.; Chanyalew, S.; Blosch, R.; Farinelli, L.; et al. Genome and transcriptome sequencing identifies breeding targets in the orphan crop tef (Eragrostis tef). BMC Genom. 2014, 15, 581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ravet, K.; Patterson, E.L.; Krahmer, H.; Hamouzova, K.; Fan, L.; Jasieniuk, M.; Lawton-Rauh, A.; Malone, J.M.; McElroy, J.S.; Merotto, A., Jr.; et al. The power and potential of genomics in weed biology and management. Pest Manag. Sci. 2018, 74, 2216–2225. [Google Scholar] [CrossRef]
- Brodt, S.; Six, J.; Feenstra, G.; Ingels, C.; Campbell, D. Sustainable agriculture. Nat. Educ. Knowl. 2011, 3. [Google Scholar]
- Sherwood, S.; Uphoff, N. Soil health: Research, practice and policy for a more regenerative agriculture. Appl. Soil Ecol. 2000, 15, 85–97. [Google Scholar] [CrossRef]
- Gerber, J.M. Farmer participation in research: A model for adaptive research and education. Am. J. Altern. Agric. 1992, 7, 118–121. [Google Scholar] [CrossRef]
- Shiming, L.; Gliessman, S.R. Agroecology in China: Science, Practice, and Sustainable Management; CRC Press: Boca Raton, FL, USA, 2017. [Google Scholar]
- Chel, A.; Kaushik, G. Renewable energy for sustainable agriculture. Agron. Sustain. Dev. 2011, 31, 91–118. [Google Scholar] [CrossRef]
- Godfray, H.C.J.; Beddington, J.R.; Crute, I.R.; Haddad, L.; Lawrence, D.; Muir, J.F.; Pretty, J.; Robinson, S.; Thomas, S.M.; Toulmin, C. Food security: The challenge of feeding 9 billion people. Science 2010, 327, 812–818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osei, M.; Asante, M.; Agyeman, A.; Adebayo, M.; Adu-Dapaah, H. Plant breeding: A tool for achieving food sufficiency. In Sustainable Horticultural Systems; Springer: Berlin, Germany, 2014; pp. 253–274. [Google Scholar]
- Varshney, R.K.; Singh, V.K.; Kumar, A.; Powell, W.; Sorrells, M.E. Can genomics deliver climate-change ready crops? Curr. Opin. Plant Biol. 2018, 45, 205–211. [Google Scholar] [CrossRef] [PubMed]
- Brush, S.B. Rethinking crop genetic resource conservation. Conserv. Biol. 1989, 3, 19–29. [Google Scholar] [CrossRef]
- Elijah, O.; Rahman, T.A.; Orikumhi, I.; Leow, C.Y.; Hindia, M.N. An overview of Internet of things (IoT) and data analytics in agriculture: Benefits and challenges. IEEE Internet Things J. 2018, 5, 3758–3773. [Google Scholar] [CrossRef]
- Ruiz-Garcia, L.; Lunadei, L.; Barreiro, P.; Robla, I. A review of wireless sensor technologies and applications in agriculture and food industry: State of the art and current trends. Sensors 2009, 9, 4728–4750. [Google Scholar] [CrossRef] [Green Version]
- Pivoto, D.; Waquil, P.D.; Talamini, E.; Finocchio, C.P.S.; Dalla Corte, V.F.; De Vargas Mores, G. Scientific development of smart farming technologies and their application in Brazil. Inf. Process. Agric. 2018, 5, 21–32. [Google Scholar] [CrossRef]
- McBride, W.D.; Daberkow, S.G. Information and the adoption of precision farming technologies. J. Agribus. 2003, 21, 21. [Google Scholar]
- Theriault, V.; Smale, M.; Haider, H. Economic incentives to use fertilizer on maize under differing agro-ecological conditions in Burkina Faso. Food Secur. 2018, 10, 1263–1277. [Google Scholar] [CrossRef] [Green Version]
- Tilman, D.; Cassman, K.G.; Matson, P.A.; Naylor, R.; Polasky, S. Agricultural sustainability and intensive production practices. Nat. Biotechnol. 2002, 418, 671. [Google Scholar] [CrossRef]
- Hirochika, H.; Guiderdoni, E.; An, G.; Hsing, Y.-i.; Eun, M.Y.; Han, C.-d.; Upadhyaya, N.; Ramachandran, S.; Zhang, Q.; Pereira, A. Rice mutant resources for gene discovery. Plant. Mol. Biol. 2004, 54, 325–334. [Google Scholar] [CrossRef]
- Magdoff, F.; Foster, J.B.; Buttel, F.H. Hungry for Profit: The Agribusiness Threat to Farmers, Food, and the Environment; NYU Press: New York, NY, USA, 2000. [Google Scholar]
- Tran, D.V. World Rice Production: Main Issues and Technical Possibilities; Cahiers Options Méditerranéennes: Paris, France, 1998; Volume 24. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Gene(s) | Biotic or Abiotic Stress | Reference |
|---|---|---|
| Asr1 | Salinity stress | [83] |
| Bar | Basta herbicide | [84] |
| EXPA3 | Salinity tolerance | [85] |
| HKT, NHX1 and SOS1 | Salinity stress | [86] |
| OVP1 | Cold stress | [87] |
| PDR8 | Non-host resistance | [83] |
| Pi-ta and Ptr(t) | Blast | [88] |
| Rc, Bh4 and Phr1 | Aging | [31] |
| Snl6 | Bacterial blight | [83] |
| Cereal | Pseudo-cereal | |||
|---|---|---|---|---|
| Teff | Proso Millet | Quinoa | Amaranth | |
| Centre of diversity | Eastern Africa | China | Latin America | South America |
| Family | Poaceae | Poaceae | Amaranthaceae | Amaranthaceae |
| Cultivated species | Eragrostis tef | Panicum miliaceum | Chenopodium quinoa | Amaranthus caudatus; A. cruentus; A. hypochondriacus |
| Genome size | ca. 730 Mbp | ca. 1020 Mbp | ca. 1450 Mbp | ca. 500 Mbp |
| Chromosome number | 2n = 4x = 40 | 2n = 4x =36 | 2n = 4x =36 | 2n = 2x = 32 or 2n = 2x =3 4 |
| Photosynthetic pathway | C4 | C4 | C3 | C4 |
| Salinity tolerance | Broad intraspecific variation | Tolerant | Tolerant | Tolerant |
| Cold tolerance | Tolerant | Sensitive | Tolerant | Sensitive |
| Drought tolerance | Moderately tolerant | Tolerant | Tolerant | Tolerant |
| Heat tolerance | Tolerant | Tolerant | Tolerant | Tolerant |
| Waterlogging tolerance | Tolerant | Sensitive | Sensitive | Sensitive |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mohd Hanafiah, N.; Mispan, M.S.; Lim, P.E.; Baisakh, N.; Cheng, A. The 21st Century Agriculture: When Rice Research Draws Attention to Climate Variability and How Weedy Rice and Underutilized Grains Come in Handy. Plants 2020, 9, 365. https://doi.org/10.3390/plants9030365
Mohd Hanafiah N, Mispan MS, Lim PE, Baisakh N, Cheng A. The 21st Century Agriculture: When Rice Research Draws Attention to Climate Variability and How Weedy Rice and Underutilized Grains Come in Handy. Plants. 2020; 9(3):365. https://doi.org/10.3390/plants9030365
Chicago/Turabian StyleMohd Hanafiah, Noraikim, Muhamad Shakirin Mispan, Phaik Eem Lim, Niranjan Baisakh, and Acga Cheng. 2020. "The 21st Century Agriculture: When Rice Research Draws Attention to Climate Variability and How Weedy Rice and Underutilized Grains Come in Handy" Plants 9, no. 3: 365. https://doi.org/10.3390/plants9030365